
Size Effects of Copper Oxide Nanoparticles on Boosting Soybean Growth via Differentially Modulating Nitrogen Assimilation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nanoparticle Characterization
2.2. Pot Experiment
2.2.1. Concentration Optimization of CuO NPs
2.2.2. Pot Experiment with Optimal Concentrations of CuO NPs
2.3. Enzyme Activity Related to Nitrogen Assimilation
2.3.1. Nitrite Reductase
2.3.2. Glutamate Synthase
2.4. Determination of the Content of Nitrogenous Compounds
2.4.1. Nitrate-N
2.4.2. Ammonia-N
2.4.3. Amino Acid and Protein
2.5. Soil Enzyme Activity
2.5.1. Soil Urease
2.5.2. Soil Neutral Phosphatase
2.5.3. Soil Polyphenol Oxidase
2.5.4. Soil Sucrase
2.6. Element Content Measurement
2.7. Statistical Analysis
3. Results and Discussion
3.1. Effects of CuO NPs on Soybean Biomass
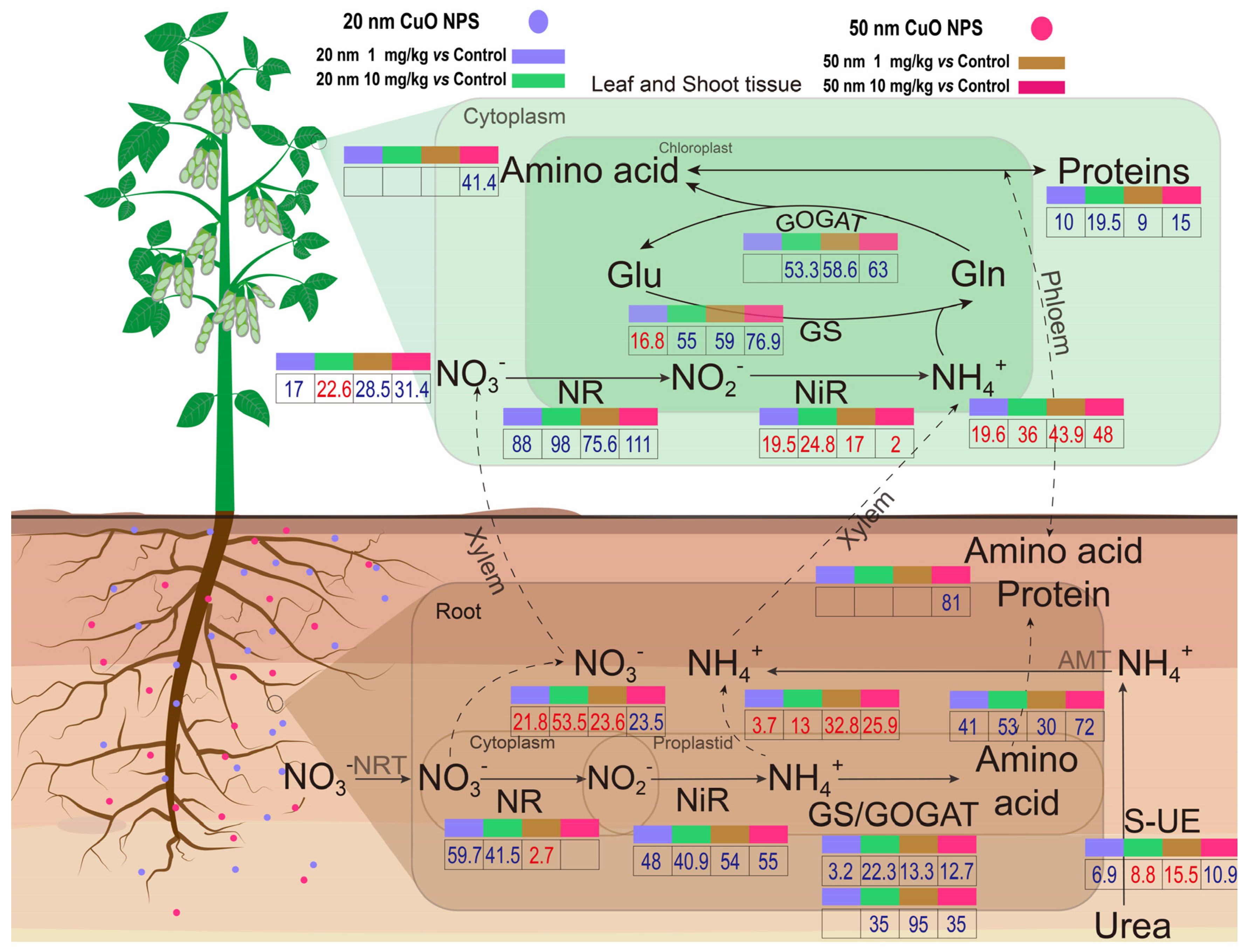
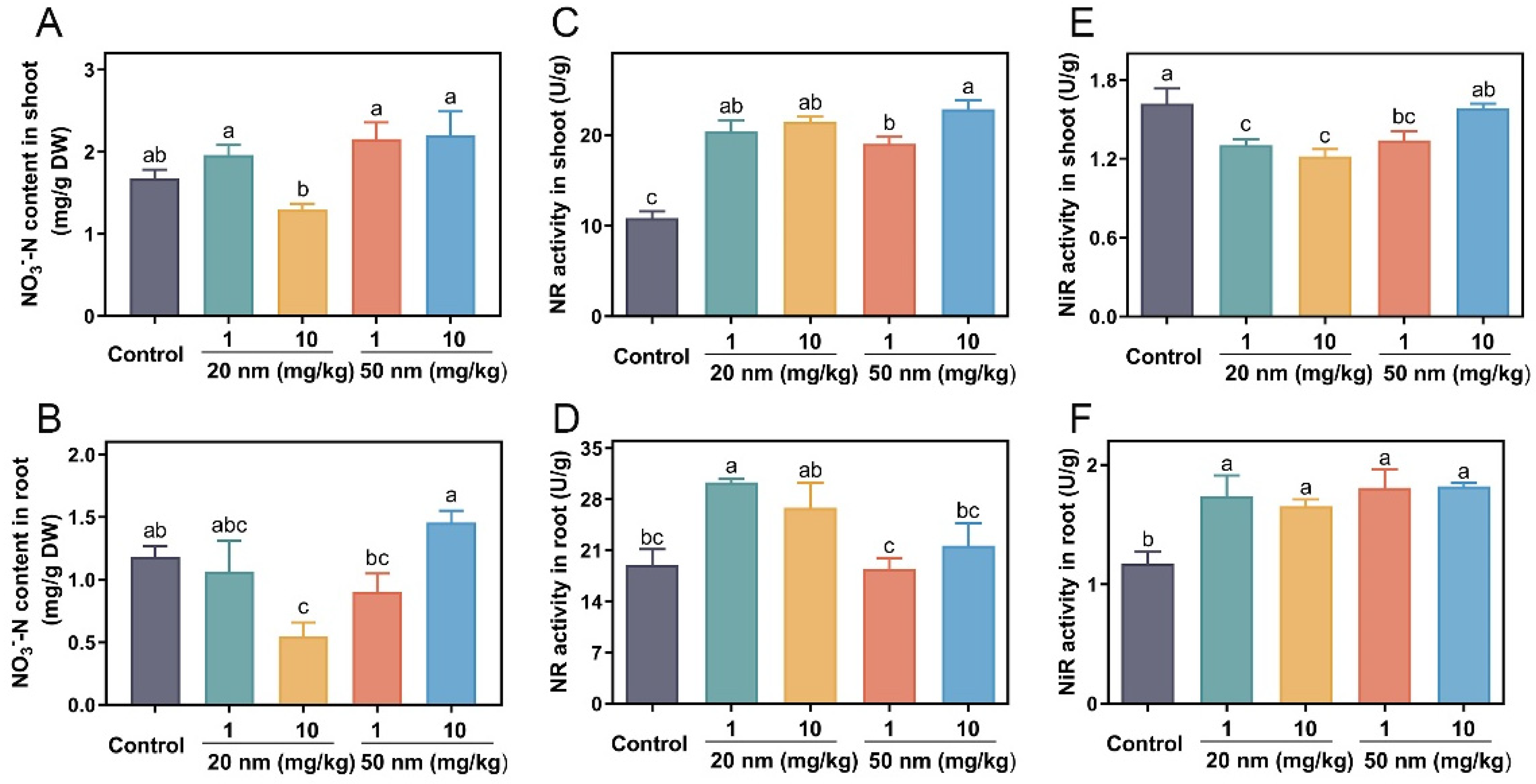
3.2. Effects of CuO NPs on Nitrate Reduction in Soybeans
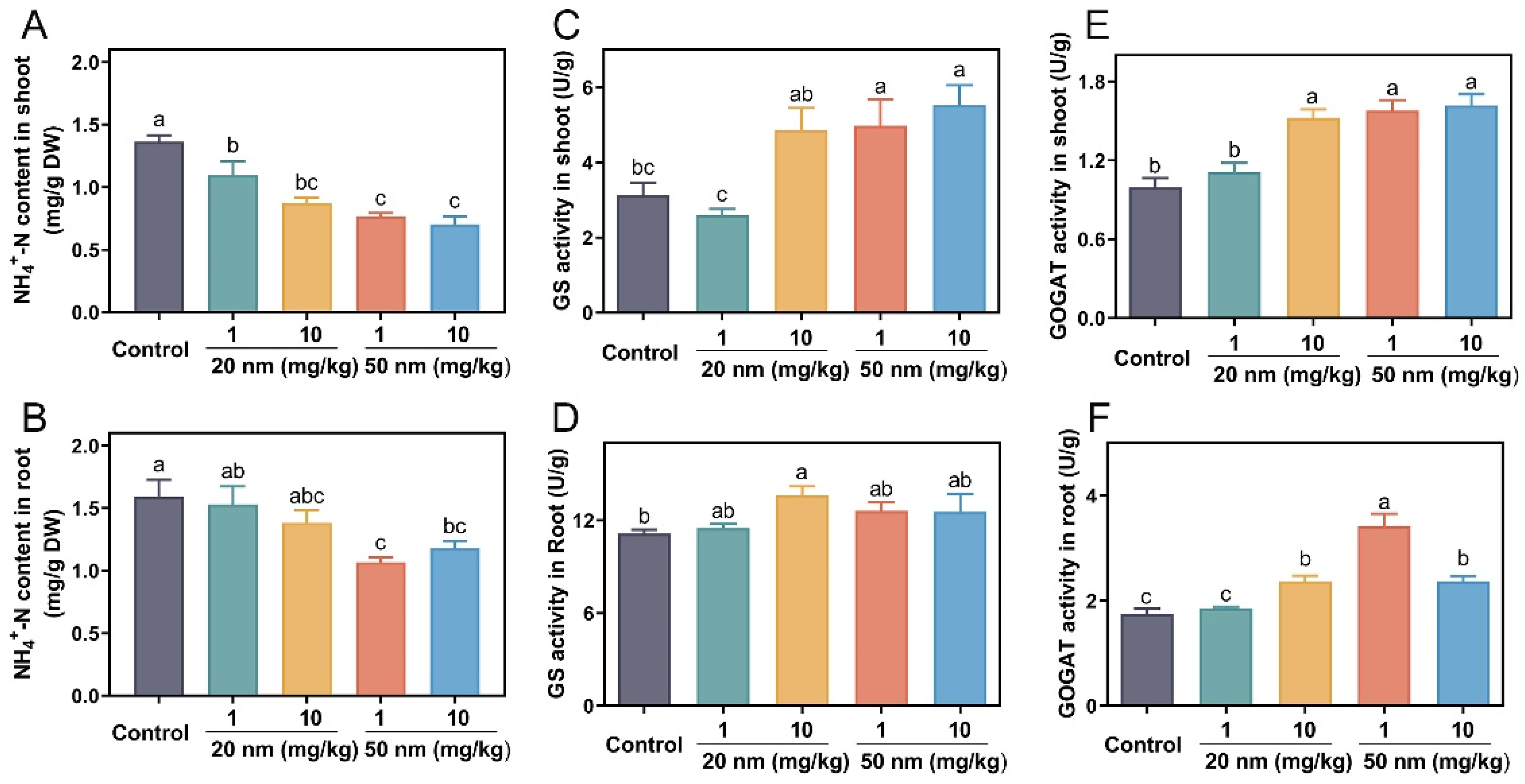
3.3. Effects of CuO NPs on Ammonia Assimilation in Soybeans
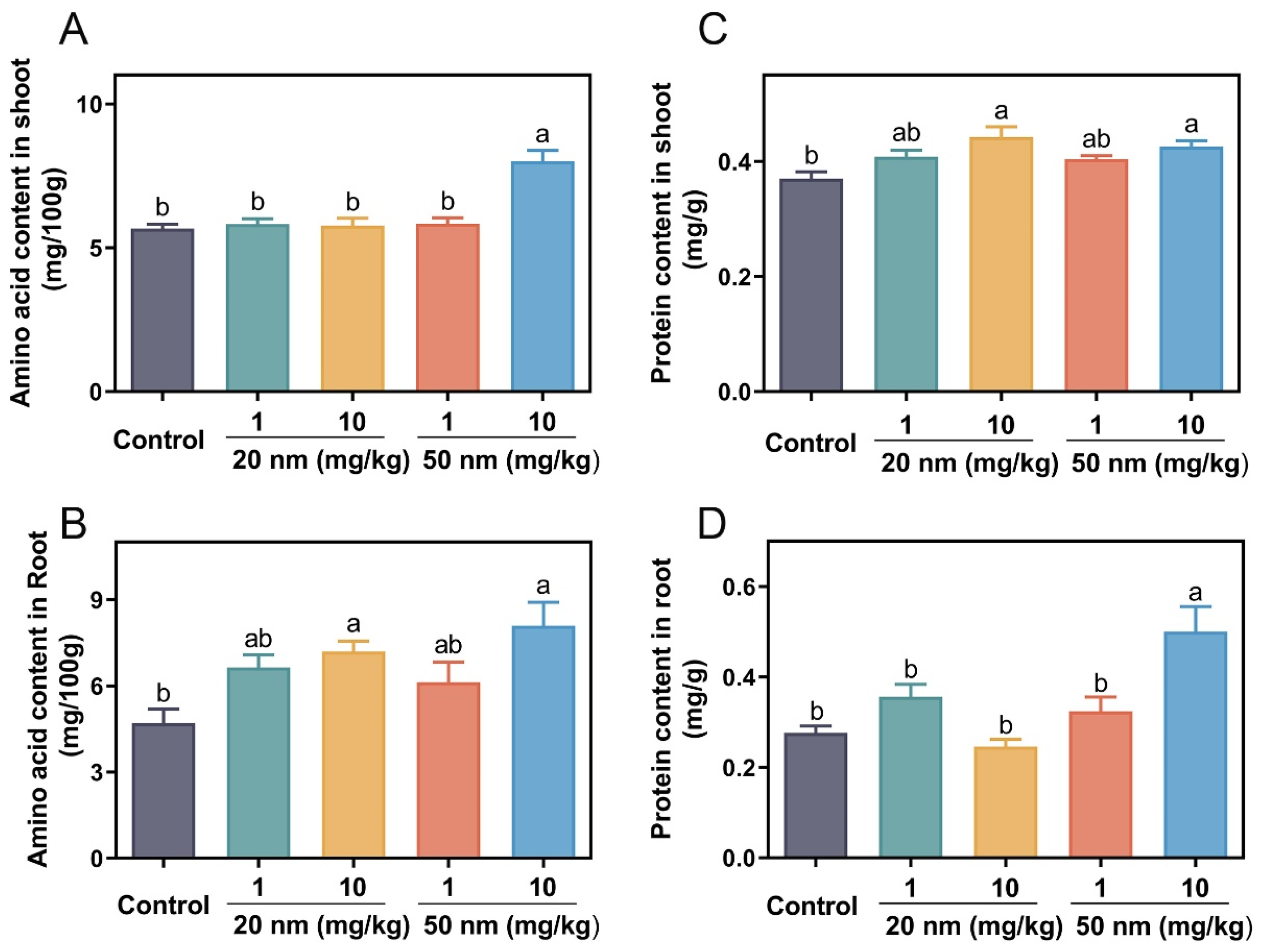
3.4. Amino Acid and Soluble Protein Content
3.5. Soil Enzyme Activity
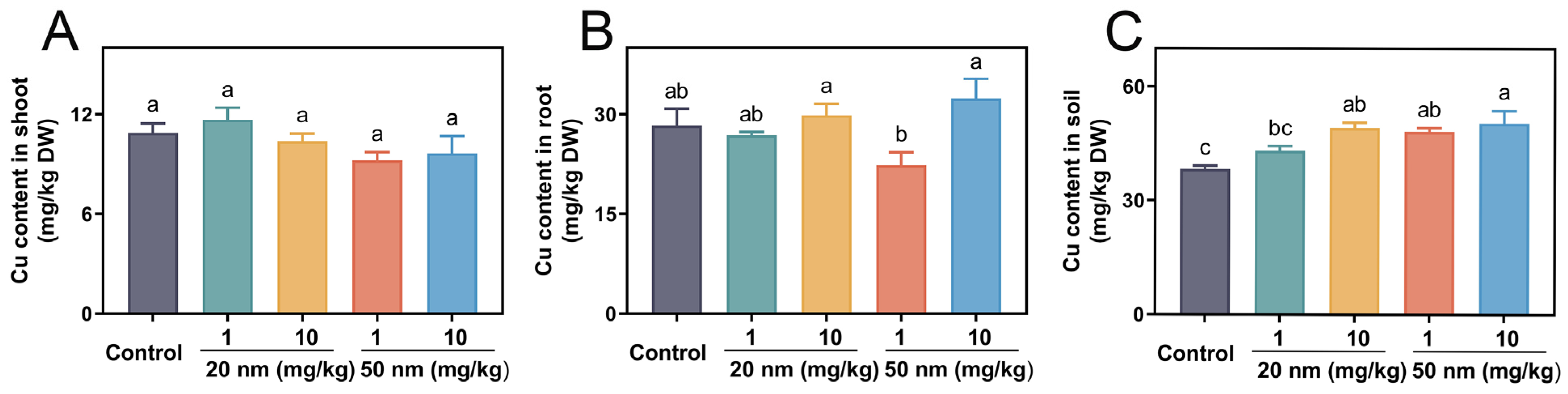
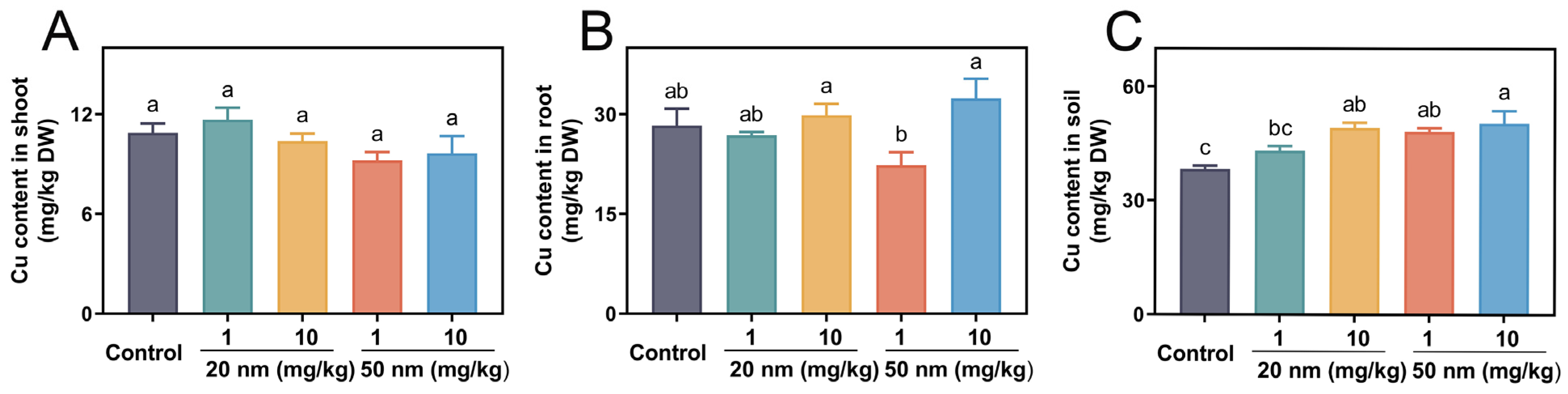
3.6. Cu Uptake and Accumulation in Soybeans
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, R.; Lal, R. Potentials of engineered nanoparticles as fertilizers for increasing agronomic productions. Sci. Total Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, R.; Gulati, D.; Sethi, G. Biofilms and nanoparticles: Applications in agriculture. Folia Microbiol. 2021, 66, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Raghupathi, K.R.; Koodali, R.T.; Manna, A.C. Size-Dependent Bacterial Growth Inhibition and Mechanism of Antibacterial Activity of Zinc Oxide Nanoparticles. Langmuir 2011, 27, 4020–4028. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Wu, D.; Zhang, J.; Tong, X.; Yin, Y.; Sun, Y.; Guo, H. Size-dependent biological effect of copper oxide nanoparticles exposure on cucumber (Cucumis sativus). Environ. Sci. Pollut. Res. 2022, 29, 69517–69526. [Google Scholar] [CrossRef]
- Chhipa, H. Chapter 6—Applications of nanotechnology in agriculture. In Methods in Microbiology; Gurtler, V., Ball, A.S., Soni, S., Eds.; Academic Press: Jhalawar, India, 2019; Volume 46, pp. 115–142. [Google Scholar] [CrossRef]
- Singh, R.P.; Handa, R.; Manchanda, G. Nanoparticles in sustainable agriculture: An emerging opportunity. J. Control. Release 2021, 329, 1234–1248. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Kumar, A.; Tomar, P.K. Synthesis and characterization of copper nanoparticles by reducing agent. J. Saudi Chem. Soc. 2014, 18, 149–153. [Google Scholar] [CrossRef]
- Rastogi, A.; Zivcak, M.; Sytar, O.; Kalaji, H.M.; He, X.; Mbarki, S.; Brestic, M. Impact of Metal and Metal Oxide Nanoparticles on Plant: A Critical Review. Front. Chem. 2017, 5, 78. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Ali, G.A.M.; Hassanein, A.; Attia, A.M.; Marzouk, E.R. Toxicity and Uptake of CuO Nanoparticles: Evaluation of an Emerging Nanofertilizer on Wheat (Triticum aestivum L.) Plant. Sustainability 2022, 14, 4914. [Google Scholar] [CrossRef]
- Lee, W.M.; An, Y.J.; Yoon, H.; Kweon, H.S. Toxicity and bioavailability of copper nanoparticles to the terrestrial plants Mung Bean (Phaseolus radiatus) and Wheat (Triticum aestivum): Plant agar test for water-insoluble nanoparticles. Environ. Toxicol. Chem. 2008, 27, 1915–1921. [Google Scholar] [CrossRef]
- Velicogna, J.R.; Schwertfeger, D.M.; Beer, C.; Jesmer, A.H.; Kuo, J.; Chen, H.; Scroggins, R.P.; Princz, J.I. Phytotoxicity of copper oxide nanoparticles in soil with and without biosolid amendment. NanoImpact 2020, 17, 100196. [Google Scholar] [CrossRef]
- Yue, L.; Zhao, J.; Yu, X.; Lv, K.; Wang, Z.; Xing, B. Interaction of CuO nanoparticles with duckweed (Lemna minor. L): Uptake, distribution and ROS production sites. Environ. Pollut. 2018, 243, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Rawat, S.; Pullagurala, V.L.R.; Hernandez-Molina, M.; Sun, Y.; Niu, G.; Hernandez-Viezcas, J.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Impacts of copper oxide nanoparticles on bell pepper (Capsicum annum L.) plants: A full life cycle study. Environ. Sci. Nano 2018, 5, 83–95. [Google Scholar] [CrossRef]
- Pelegrino, M.T.; Kohatsu, M.Y.; Seabra, A.B.; Monteiro, L.R.; Gomes, D.G.; Oliveira, H.C.; Rolim, W.R.; de Jesus, T.A.; Batista, B.L.; Lange, C.N. Effects of copper oxide nanoparticles on growth of lettuce (Lactuca sativa L.) seedlings and possible implications of nitric oxide in their antioxidative defense. Environ. Monit. Assess. 2020, 192, 232. [Google Scholar] [CrossRef] [PubMed]
- Kohatsu, M.Y.; Pelegrino, M.T.; Monteiro, L.R.; Freire, B.M.; Pereira, R.M.; Fincheira, P.; Rubilar, O.; Tortella, G.; Batista, B.L.; de Jesus, T.A.; et al. Comparison of foliar spray and soil irrigation of biogenic CuO nanoparticles (NPs) on elemental uptake and accumulation in lettuce. Environ. Sci. Pollut. Res. 2021, 28, 16350–16367. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Adhikari, S.; Adhikari, A.; Ghosh, S.; Azahar, I.; Basuli, D.; Hossain, Z. Impact of CuO nanoparticles on maize: Comparison with CuO bulk particles with special reference to oxidative stress damages and antioxidant defense status. Chemosphere 2022, 287, 131911. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.J.; Kwak, J.I.; Lee, W.M.; Holden, P.A.; An, Y.J. Zinc oxide nanoparticles delay soybean development: A standard soil microcosm study. Ecotoxicol. Environ. Saf. 2014, 100, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Sá, A.G.A.; Moreno, Y.M.F.; Carciofi, B.A.M. Plant proteins as high-quality nutritional source for human diet. Trends Food Sci. Technol. 2020, 97, 170–184. [Google Scholar] [CrossRef]
- Priester, J.H.; Ge, Y.; Mielke, R.E.; Horst, A.M.; Moritz, S.C.; Espinosa, K.; Gelb, J.; Walker, S.L.; Nisbet, R.M.; An, Y.-J.; et al. Soybean susceptibility to manufactured nanomaterials with evidence for food quality and soil fertility interruption. Proc. Natl. Acad. Sci. USA 2012, 109, E2451–E2456. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Stowers, C.; Rossi, L.; Zhang, W.; Lombardini, L.; Ma, X. Physiological effects of cerium oxide nanoparticles on the photosynthesis and water use efficiency of soybean (Glycine max (L.) Merr.). Environ. Sci. Nano 2017, 4, 1086–1094. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Cassman, K.G.; Specht, J.E.; Walters, D.T.; Weiss, A.; Dobermann, A. Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crop. Res. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- Mejias, J.H.; Salazar, F.; Pérez Amaro, L.; Hube, S.; Rodriguez, M.; Alfaro, M. Nanofertilizers: A Cutting-Edge Approach to Increase Nitrogen Use Efficiency in Grasslands. Front. Environ. Sci. 2021, 9, 635114. [Google Scholar] [CrossRef]
- Xu, H.; Liu, C.; Lu, R.; Guo, G.; Chen, Z.; He, T.; Gao, R.; Li, Y.; Huang, J. The difference in responses to nitrogen deprivation and re-supply at seedling stage between two barley genotypes differing nitrogen use efficiency. Plant Growth Regul. 2016, 79, 119–126. [Google Scholar] [CrossRef]
- Wang, M.; Shen, Q.; Xu, G.; Guo, S. New insight into the strategy for nitrogen metabolism in plant cells. Int. Rev. Cell Mol. Biol. 2014, 310, 1–37. [Google Scholar] [CrossRef]
- Guan, X.; Gao, X.; Avellan, A.; Spielman-Sun, E.; Xu, J.; Laughton, S.; Yun, J.; Zhang, Y.; Bland, G.D.; Zhang, Y.; et al. CuO Nanoparticles Alter the Rhizospheric Bacterial Community and Local Nitrogen Cycling for Wheat Grown in a Calcareous Soil. Environ. Sci. Technol. 2020, 54, 8699–8709. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Xie, F. Effect of Drought Stress at Reproductive Stages on Growth and Nitrogen Metabolism in Soybean. Agronomy 2020, 10, 302. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, R.; Chen, Z.; Pu, J.; Wang, J.; Zhang, H.; Yang, Y. Nanoscale molybdenum oxide improves plant growth and increases nitrate utilisation in rice (Oryza sativa L.). Food Energy Secur. 2022, 11, e383. [Google Scholar] [CrossRef]
- Wang, Y. Effect of NPK on NR and NiR activity of sugar beet. J. Nucl. Agric. Sci. 2012, 26, 0803–0808. [Google Scholar]
- Zhou, P.; Jiang, Y.; Adeel, M.; Shakoor, N.; Zhao, W.; Liu, Y.; Li, Y.; Li, M.; Azeem, I.; Rui, Y.; et al. Nickel Oxide Nanoparticles Improve Soybean Yield and Enhance Nitrogen Assimilation. Environ. Sci. Technol. 2023, 57, 7547–7558. [Google Scholar] [CrossRef]
- Ponce García, C.O.; Soto Parra, J.M.; Sánchez, E.; Muñoz-Márquez, E.; Piña-Ramírez, F.J.; Flores-Córdova, M.A.; Pérez-Leal, R.; Yáñez Muñoz, R.M. Efficiency of Nanoparticle, Sulfate, and Zinc-Chelate Use on Biomass, Yield, and Nitrogen Assimilation in Green Beans. Agronomy 2019, 9, 128. [Google Scholar] [CrossRef]
- Pradhan, S.; Patra, P.; Mitra, S.; Dey, K.K.; Jain, S.; Sarkar, S.; Roy, S.; Palit, P.; Goswami, A. Manganese Nanoparticles: Impact on Non-nodulated Plant as a Potent Enhancer in Nitrogen Metabolism and Toxicity Study both In Vivo and In Vitro. J. Agric. Food Chem. 2014, 62, 8777–8785. [Google Scholar] [CrossRef]
- Li, M.; Zhang, P.; Guo, Z.; Zhao, W.; Li, Y.; Yi, T.; Cao, W.; Gao, L.; Tian, C.F.; Chen, Q.; et al. Dynamic Transformation of Nano-MoS2 in a Soil–Plant System Empowers Its Multifunctionality on Soybean Growth. Environ. Sci. Technol. 2024, 58, 1211–1222. [Google Scholar] [CrossRef] [PubMed]
- Bagale, S.; Abdelhamid, M. Nutrient Management for Soybean Crops. Int. J. Agron. 2021, 2021, 3304634. [Google Scholar] [CrossRef]
- Miflin, B.J. Distribution of Nitrate and Nitrite Reductase in Barley. Nature 1967, 214, 1133–1134. [Google Scholar] [CrossRef]
- Wang, X.; Xiong, S.; Ma, X.; Zhang, J.; Wang, Z. Effects of different nitrogen forms on key enzyme activity involved in nitrogen metabolism and grain protein content in speciality wheat cultivars. Acta Ecol. Sin. 2005, 25, 802–807. [Google Scholar]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 2008, 6, 71–80. [Google Scholar] [CrossRef]
- Weatherburn, M.W. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Xu, X.; Zhao, C.; Qian, K.; Sun, M.; Hao, Y.; Han, L.; Wang, C.; Ma, C.; White, J.C.; Xing, B. Physiological responses of pumpkin to zinc oxide quantum dots and nanoparticles. Environ. Pollut. 2022, 296, 118723. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guo, Y.Z.; Liang, A.Q.; Xu, X.X.; Hao, Y.; Shang, H.P.; Li, C.Y.; Cai, Z.Y.; Han, L.F.; Zhao, J.; et al. Commonly Used Engineered Nanomaterials Improve Soil Health via Suppressing Soil-Borne Fusarium and Positively Altering Soil Microbiome. Acs EST Eng. 2024, 4, 915–927. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Srivastava, A.K.; El-sadek, M.S.A.; Kordrostami, M.; Tran, L.-S.P. Titanium Dioxide Nanoparticles Improve Growth and Enhance Tolerance of Broad Bean Plants under Saline Soil Conditions. Land Degrad. Dev. 2018, 29, 1065–1073. [Google Scholar] [CrossRef]
- Aslam, M.; Huffaker, R.C. Role of Nitrate and Nitrite in the Induction of Nitrite Reductase in Leaves of Barley Seedlings. Plant Physiol. Biochem. 1989, 91, 1152–1156. [Google Scholar] [CrossRef]
- Liu, X.X.; Zhang, H.H.; Zhu, Q.Y.; Ye, J.Y.; Zhu, Y.X.; Jing, X.T.; Du, W.X.; Zhou, M.; Lin, X.Y.; Zheng, S.J.; et al. Phloem iron remodels root development in response to ammonium as the major nitrogen source. Nat. Commun. 2022, 13, 561. [Google Scholar] [CrossRef]
- Li, Y.; Ma, C.; Gong, Z. Study on glutamine synthetase activity in spring soybean. J. Northeast. Agric. Univ. 2006, 6, 729–732. [Google Scholar] [CrossRef]
- Li, X.; Yao, S.; Bian, Y.; Jiang, X.; Song, Y. The combination of biochar and plant roots improves soil bacterial adaptation to PAH stress: Insights from soil enzymes, microbiome, and metabolome. J. Hazard. Mater. 2020, 400, 123227. [Google Scholar] [CrossRef]
- Wang, C.; Ji, Y.; Cao, X.; Yue, L.; Chen, F.; Li, J.; Yang, H.; Wang, Z.; Xing, B. Carbon Dots Improve Nitrogen Bioavailability to Promote the Growth and Nutritional Quality of Soybeans under Drought Stress. ACS Nano 2022, 16, 12415–12424. [Google Scholar] [CrossRef]
- Yu, Q. Study on Soil Phosphat ase Activity and their Influenced Factors. J. Anhui Tech. Teach. Coll. 2001, 15, 5–8. [Google Scholar]
- Zhao, Y.; Li, X.; Han, S.; Hu, Y. Soil enzyme activities under two forest types as affected by different levels of nitrogen deposition. Chin. J. Appl. Ecol. 2008, 19, 2769–2773. [Google Scholar]
- Wang, J.; Li, G.; Xiu, M.; Song, X.; Zhao, J.; Yang, D. Effects of Nitrogen and Water on Soil Enzyme Activity and Soil Microbial Biomass in Stipa baicalensis Steppe, Inner Mongolia of North China. J. Agric. Resour. Environ. 2014, 31, 237–245. [Google Scholar] [CrossRef]
- Liu, C.; Yu, Y.C.; Liu, H.Z.; Xin, H. Effect of different copper oxide particles on cell division and related genes of soybean roots. Plant Physiol. Biochem. 2021, 163, 205–214. [Google Scholar] [CrossRef]
- Wang, S.; Liu, H.; Zhang, Y.; Xin, H. The effect of CuO NPs on reactive oxygen species and cell cycle gene expression in roots of rice. Environ. Toxicol. Chem. 2015, 34, 554–561. [Google Scholar] [CrossRef]
- Dietz, K.J.; Herth, S. Plant nanotoxicology. Trends Plant Sci. 2011, 16, 582–589. [Google Scholar] [CrossRef]
- Navarro, E.; Piccapietra, F.; Wagner, B.; Marconi, F.; Kaegi, R.; Odzak, N.; Sigg, L.; Behra, R. Toxicity of Silver Nanoparticles to Chlamydomonas reinhardtii. Environ. Sci. Technol. 2008, 42, 8959–8964. [Google Scholar] [CrossRef] [PubMed]
- Kurepa, J.; Paunesku, T.; Vogt, S.; Arora, H.; Rabatic, B.M.; Lu, J.; Wanzer, M.B.; Woloschak, G.E.; Smalle, J.A. Uptake and Distribution of Ultrasmall Anatase TiO2 Alizarin Red S Nanoconjugates in Arabidopsis thaliana. Nano Lett. 2010, 10, 2296–2302. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.K.; Ghosh, S.; Kalaji, H.M.; Bosa, K.; Brestic, M.; Zivcak, M.; Hossain, Z. Nano-CuO stress induced modulation of antioxidative defense and photosynthetic performance of Syrian barley (Hordeum vulgare L.). Environ. Exp. Bot. 2014, 102, 37–47. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Sako, K.; Matsui, A.; Suzuki, Y.; Mostofa, M.G.; Ha, C.V.; Tanaka, M.; Tran, L.P.; Habu, Y.; Seki, M. Ethanol Enhances High-Salinity Stress Tolerance by Detoxifying Reactive Oxygen Species in Arabidopsis thaliana and Rice. Front. Plant Sci. 2017, 8, 1001. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Li, H.; Hao, Y.; Shang, H.; Jia, W.; Liang, A.; Xu, X.; Li, C.; Ma, C. Size Effects of Copper Oxide Nanoparticles on Boosting Soybean Growth via Differentially Modulating Nitrogen Assimilation. Nanomaterials 2024, 14, 746. https://doi.org/10.3390/nano14090746
Guo Y, Li H, Hao Y, Shang H, Jia W, Liang A, Xu X, Li C, Ma C. Size Effects of Copper Oxide Nanoparticles on Boosting Soybean Growth via Differentially Modulating Nitrogen Assimilation. Nanomaterials. 2024; 14(9):746. https://doi.org/10.3390/nano14090746
Chicago/Turabian StyleGuo, Yaozu, Hao Li, Yi Hao, Heping Shang, Weili Jia, Anqi Liang, Xinxin Xu, Chunyang Li, and Chuanxin Ma. 2024. "Size Effects of Copper Oxide Nanoparticles on Boosting Soybean Growth via Differentially Modulating Nitrogen Assimilation" Nanomaterials 14, no. 9: 746. https://doi.org/10.3390/nano14090746
APA StyleGuo, Y., Li, H., Hao, Y., Shang, H., Jia, W., Liang, A., Xu, X., Li, C., & Ma, C. (2024). Size Effects of Copper Oxide Nanoparticles on Boosting Soybean Growth via Differentially Modulating Nitrogen Assimilation. Nanomaterials, 14(9), 746. https://doi.org/10.3390/nano14090746