
Mobile Antimicrobial Resistance Genes in Probiotics

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data
2.2. DNA Extraction and Metagenomics Library Preparation for PRJNA644361
2.3. Bioinformatic Analysis
3. Results
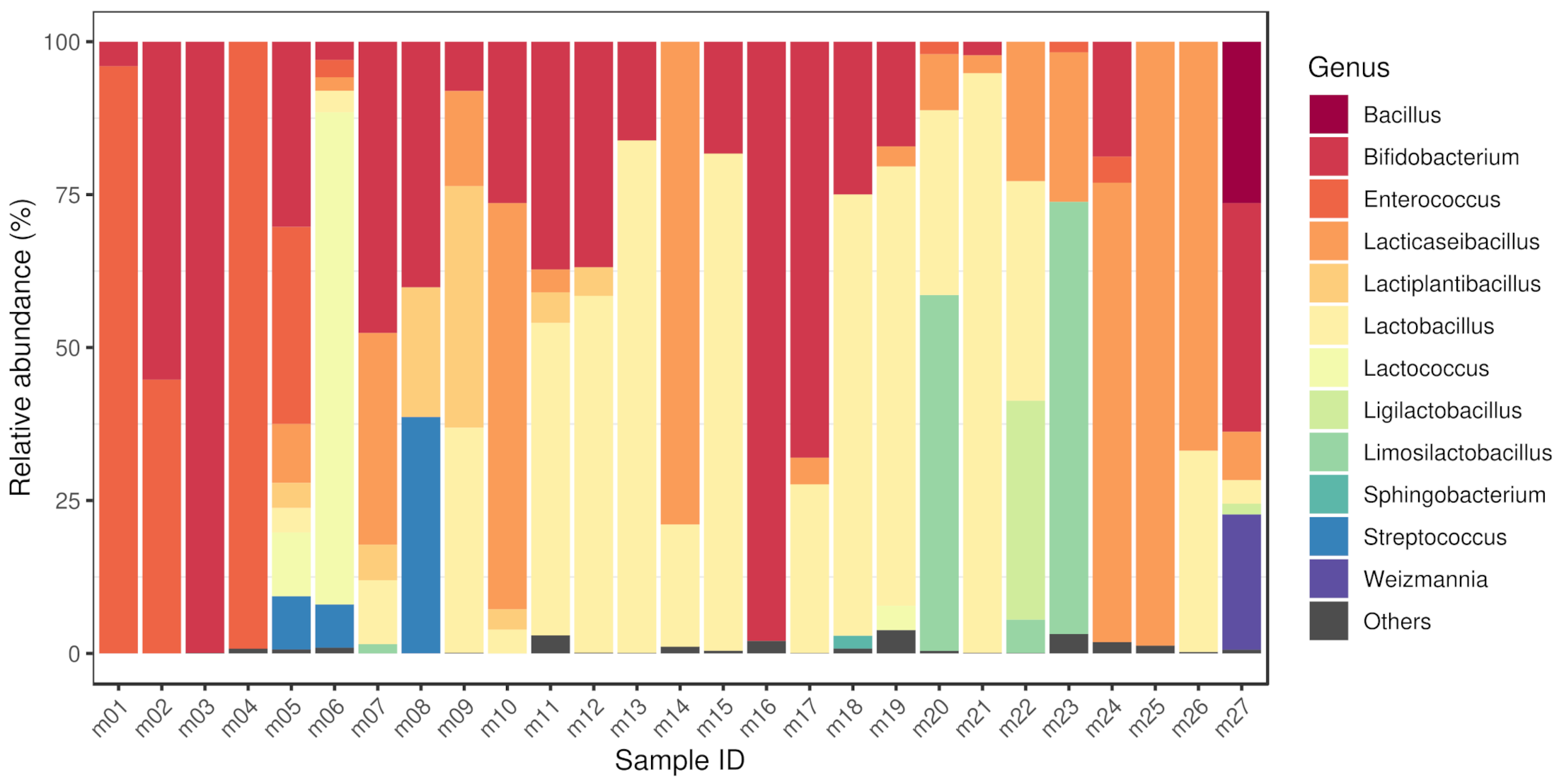
3.1. Bacteriome
3.2. Resistome
3.3. Mobilome
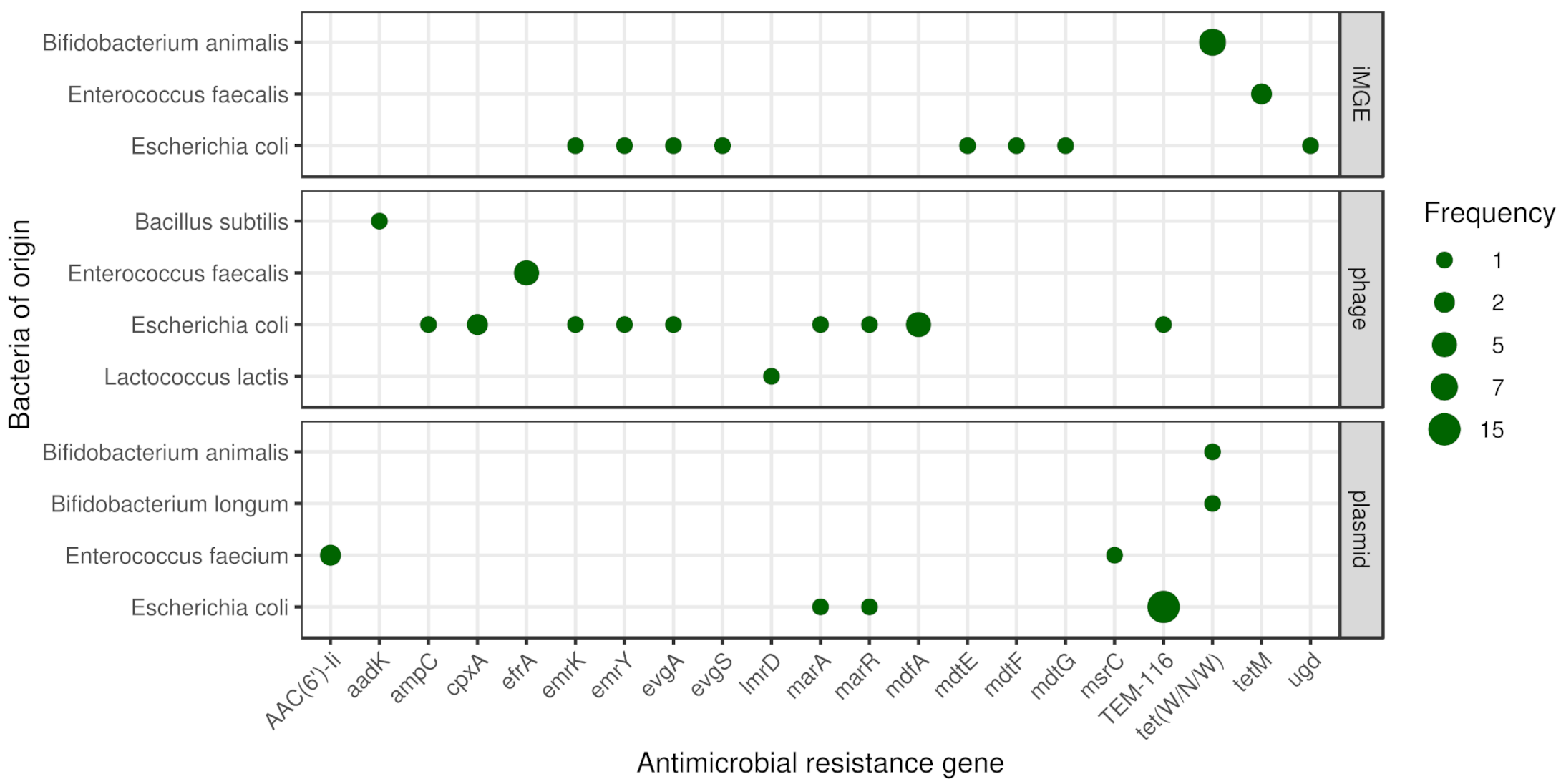
3.4. Coexistence of ARGs and iMGEs
3.5. Plasmids
3.6. Phages
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanders, M.; Merenstein, D.; Merrifield, C.; Hutkins, R. Probiotics for human use. Nutr. Bull. 2018, 43, 212–225. [Google Scholar] [CrossRef]
- Zmora, N.; Zilberman-Schapira, G.; Suez, J.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Kotler, E.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.Z.; et al. Personalized gut mucosal colonization resistance to empiric probiotics is associated with unique host and microbiome features. Cell 2018, 174, 1388–1405. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.; Karpinets, T.; Prieto, P.; Vicente, D.; Hoffman, K.; Wei, S.; et al. Gut microbiome modulates response to anti–PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Zhang, R.; Tian, X.; Zhou, X.; Pan, X.; Wong, A. Assessing the risk of probiotic dietary supplements in the context of antibiotic resistance. Front. Microbiol. 2017, 8, 908. [Google Scholar] [CrossRef]
- Berreta, A.; Baumgardner, R.M.; Kopper, J.J. Evaluation of commercial veterinary probiotics containing enterococci for transferrable vancomycin resistance genes. BMC Res. Notes 2020, 13, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rozman, V.; Lorbeg, P.M.; Accetto, T.; Matijašić, B.B. Characterization of antimicrobial resistance in lactobacilli and bifidobacteria used as probiotics or starter cultures based on integration of phenotypic and in silico data. Int. J. Food Microbiol. 2020, 314, 108388. [Google Scholar] [CrossRef] [PubMed]
- Selvin, J.; Maity, D.; Sajayan, A.; Kiran, G.S. Revealing antibiotic resistance in therapeutic and dietary probiotic supplements. J. Glob. Antimicrob. Resist. 2020, 22, 202–205. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.d.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G. Guidance on the characterisation of microorganisms used as feed additives or as production organisms. EFSA J. 2018, 16, e05206. [Google Scholar]
- European Food Safety Authority. EFSA statement on the requirements for whole genome sequence analysis of microorganisms intentionally used in the food chain. EFSA J. 2021, 19, e06506. [Google Scholar]
- Cabezón, E.; Ripoll-Rozada, J.; Peña, A.; De La Cruz, F.; Arechaga, I. Towards an integrated model of bacterial conjugation. FEMS Microbiol. Rev. 2015, 39, 81–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, S. Phage transduction. In Clostridium Difficile; Humana Press: New York, NY, USA, 2016; pp. 177–185. [Google Scholar]
- Aires, J.; Doucet-Populaire, F.; Butel, M. Tetracycline resistance mediated by tet (W), tet (M), and tet (O) genes of Bifidobacterium isolates from humans. Appl. Environ. Microbiol. 2007, 73, 2751–2754. [Google Scholar] [CrossRef] [Green Version]
- Ammor, M.S.; Flórez, A.B.; Van Hoek, A.H.; Clara, G.; Aarts, H.J.; Margolles, A.; Mayo, B. Molecular characterization of intrinsic and acquired antibiotic resistance in lactic acid bacteria and bifidobacteria. J. Mol. Microbiol. Biotechnol. 2008, 14, 6–15. [Google Scholar] [CrossRef]
- Bennedsen, M.; Stuer-Lauridsen, B.; Danielsen, M.; Johansen, E. Screening for antimicrobial resistance genes and virulence factors via genome sequencing. Appl. Environ. Microbiol. 2011, 77, 2785–2787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gueimonde, M.; Sánchez, B.; de Los Reyes-Gavilán, C.G.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef] [Green Version]
- Mancino, W.; Lugli, G.A.; van Sinderen, D.; Ventura, M.; Turroni, F. Mobilome and resistome reconstruction from genomes belonging to members of the Bifidobacterium genus. Microorganisms 2019, 7, 638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO/WHO. Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria; Food and Agriculture Organization of the United Nations and World Health Organization Expert Consultation Report; FAO: Cordoba, Argentina, 2001. [Google Scholar]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar]
- Lahti, L.; Shetty, S. Microbiome R package, 2012–2019. Available online: http://microbiome.github.io (accessed on 22 March 2021).
- Li, D.; Liu, C.M.; Luo, R.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyatt, D.; Chen, G.L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [Green Version]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The comprehensive antibiotic resistance database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Johansson, M.H.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, P.S.; Lipinski, L.; Dziembowski, A. PlasFlow: Predicting plasmid sequences in metagenomic data using genome signatures. Nucleic Acids Res. 2018, 46, e35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Bolduc, B.; Zayed, A.A.; Varsani, A.; Dominguez-Huerta, G.; Delmont, T.O.; Pratama, A.A.; Gazitúa, M.C.; Vik, D.; Sullivan, M.B.; et al. VirSorter2: A multi-classifier, expert-guided approach to detect diverse DNA and RNA viruses. Microbiome 2021, 9, 1–13. [Google Scholar] [CrossRef]
- Duc, L.H.; Hong, H.A.; Barbosa, T.M.; Henriques, A.O.; Cutting, S.M. Characterization of Bacillus probiotics available for human use. Appl. Environ. Microbiol. 2004, 70, 2161–2171. [Google Scholar] [CrossRef] [Green Version]
- Adel, M.; El-Sayed, A.F.M.; Yeganeh, S.; Dadar, M.; Giri, S.S. Effect of potential probiotic Lactococcus lactis subsp. lactis on growth performance, intestinal microbiota, digestive enzyme activities, and disease resistance of Litopenaeus vannamei. Probiotics Antimicrob. Proteins 2017, 9, 150–156. [Google Scholar] [CrossRef]
- Baccouri, O.; Boukerb, A.M.; Farhat, L.B.; Zébré, A.; Zimmermann, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I.; et al. Probiotic potential and safety evaluation of Enterococcus faecalis OB14 and OB15, isolated from traditional tunisian testouri cheese and rigouta, using physiological and genomic analysis. Front. Microbiol. 2019, 10, 881. [Google Scholar] [CrossRef] [PubMed]
- Poorni, S.; Srinivasan, M.R.; Nivedhitha, M.S. Probiotic Streptococcus strains in caries prevention: A systematic review. J. Conserv. Dent. JCD 2019, 22, 123. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, H.S.; Quigley, E.M. The probiotic Bifidobacterium in the management of Coronavirus: A theoretical basis. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420961304. [Google Scholar] [CrossRef]
- Drago, L.; Meroni, G.; Chiaretti, A.; Laforgia, N.; Cucchiara, S.; Baldassarre, M.E. Effect of Limosilactobacillus reuteri LRE02–Lacticaseibacillus rhamnosus LR04 Combination on Antibiotic-Associated Diarrhea in a Pediatric Population: A National Survey. J. Clin. Med. 2020, 9, 3080. [Google Scholar] [CrossRef]
- Minj, J.; Chandra, P.; Paul, C.; Sharma, R.K. Bio-functional properties of probiotic Lactobacillus: Current applications and research perspectives. Crit. Rev. Food Sci. Nutr. 2020, 61, 2207–2264. [Google Scholar] [CrossRef] [PubMed]
- Dec, M.; Stępień-Pyśniak, D.; Puchalski, A.; Hauschild, T.; Pietras-Ożga, D.; Ignaciuk, S.; Urban-Chmiel, R. Biodiversity of Ligilactobacillus salivarius Strains from Poultry and Domestic Pigeons. Animals 2021, 11, 972. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, N.; Battista, N.; Prete, R.; Corsetti, A. Health-Promoting Role of Lactiplantibacillus plantarum Isolated from Fermented Foods. Microorganisms 2021, 9, 349. [Google Scholar] [CrossRef] [PubMed]
- Hussain, N.; Li, R.; Takala, T.M.; Tariq, M.; Zaidi, A.H.; Saris, P.E. Generation of Lactose-and Protease-Positive Probiotic Lacticaseibacillus rhamnosus GG by Conjugation with Lactococcus lactis NCDO 712. Appl. Environ. Microbiol. 2021, 87, e02957-20. [Google Scholar] [CrossRef] [PubMed]
- Kunchala, R.; Banerjee, R.; Mazumdar, S.D.; Durgalla, P.; Srinivas, V.; Gopalakrishnan, S. Characterization of potential probiotic bacteria isolated from sorghum and pearl millet of the semi-arid tropics. Afr. J. Biotechnol. 2016, 15, 613–621. [Google Scholar]
- Ezekiel, C.N.; Ayeni, K.I.; Ezeokoli, O.T.; Sulyok, M.; van Wyk, D.A.; Oyedele, O.A.; Akinyemi, O.M.; Chibuzor-Onyema, I.E.; Adeleke, R.A.; Nwangburuka, C.C.; et al. High-throughput sequence analyses of bacterial communities and multi-mycotoxin profiling during processing of different formulations of Kunu, a traditional fermented beverage. Front. Microbiol. 2019, 9, 3282. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Patel, S.; Saini, N.; Chen, S. Robust demarcation of 17 distinct Bacillus species clades, proposed as novel Bacillaceae genera, by phylogenomics and comparative genomic analyses: Description of Robertmurraya kyonggiensis sp. nov. and proposal for an emended genus Bacillus limiting it only to the members of the Subtilis and Cereus clades of species. Int. J. Syst. Evol. Microbiol. 2020, 70, 5753–5798. [Google Scholar] [PubMed]
- Cao, J.; Yu, Z.; Liu, W.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Probiotic characteristics of Bacillus coagulans and associated implications for human health and diseases. J. Funct. Foods 2020, 64, 103643. [Google Scholar] [CrossRef]
- Keiichi, O.; Takaaki, T.; Norihisa, N.; Megumi, K. Nucleotide sequence of the chromosomal gene coding for the aminoglycoside 6-adenylyltransferase from Bacillus subtilis Marburg 168. Gene 1989, 78, 377–378. [Google Scholar] [CrossRef]
- Klyachko, K.A.; Schuldiner, S.; Neyfakh, A.A. Mutations affecting substrate specificity of the Bacillus subtilis multidrug transporter Bmr. J. Bacteriol. 1997, 179, 2189–2193. [Google Scholar] [CrossRef] [Green Version]
- Jack, D.L.; Storms, M.L.; Tchieu, J.H.; Paulsen, I.T.; Saier, M.H. A broad-specificity multidrug efflux pump requiring a pair of homologous SMR-type proteins. J. Bacteriol. 2000, 182, 2311–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, K.I.; Ohki, Y.H.; Murata, M.; Kinehara, M.; Matsuoka, H.; Satomura, T.; Ohki, R.; Kumano, M.; Yamane, K.; Fujita, Y. Bacillus subtilis LmrA is a repressor of the lmrAB and yxaGH operons: Identification of its binding site and functional analysis of lmrB and yxaGH. J. Bacteriol. 2004, 186, 5640–5648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hachmann, A.B.; Sevim, E.; Gaballa, A.; Popham, D.L.; Antelmann, H.; Helmann, J.D. Reduction in membrane phosphatidylglycerol content leads to daptomycin resistance in Bacillus subtilis. Antimicrob. Agents Chemother. 2011, 55, 4326–4337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowe-McAuliffe, C.; Graf, M.; Huter, P.; Takada, H.; Abdelshahid, M.; Nováček, J.; Murina, V.; Atkinson, G.C.; Hauryliuk, V.; Wilson, D.N. Structural basis for antibiotic resistance mediated by the Bacillus subtilis ABCF ATPase VmlR. Proc. Natl. Acad. Sci. USA 2018, 115, 8978–8983. [Google Scholar] [CrossRef] [Green Version]
- Pawlowski, A.C.; Stogios, P.J.; Koteva, K.; Skarina, T.; Evdokimova, E.; Savchenko, A.; Wright, G.D. The evolution of substrate discrimination in macrolide antibiotic resistance enzymes. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Kim, B.J.; Kim, H.Y.; Yun, Y.J.; Kim, B.J.; Kook, Y.H. Differentiation of Bifidobacterium species using partial RNA polymerase β-subunit (rpoB) gene sequences. Int. J. Syst. Evol. Microbiol. 2010, 60, 2697–2704. [Google Scholar] [CrossRef] [Green Version]
- Lokesh, D.; Parkesh, R. Bifidobacterium adolescentis is intrinsically resistant to antitubercular drugs. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef]
- Polit, A.; Yang, H.; Amund, D. Investigating the transmissibility of tet (W) in bifidobacteria exposed to acid and bile stress. Biosci. Microbiota Food Health 2018, 37, 39–43. [Google Scholar] [CrossRef] [Green Version]
- Tóth, A.G.; Csabai, I.; Maróti, G.; Jerzsele, Á.; Dubecz, A.; Patai, Á.V.; Judge, M.F.; Nagy, S.Á.; Makrai, L.; Bányai, K.; et al. A glimpse of antimicrobial resistance gene diversity in kefir and yoghurt. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Serafini, F.; Bottacini, F.; Viappiani, A.; Baruffini, E.; Turroni, F.; Foroni, E.; Lodi, T.; van Sinderen, D.; Ventura, M. Insights into physiological and genetic mupirocin susceptibility in bifidobacteria. Appl. Environ. Microbiol. 2011, 77, 3141–3146. [Google Scholar] [CrossRef] [Green Version]
- Coque, T.M.; Singh, K.V.; Weinstock, G.M.; Murray, B.E. Characterization of Dihydrofolate Reductase Genes from Trimethoprim-Susceptible and Trimethoprim-Resistant Strains ofEnterococcus faecalis. Antimicrob. Agents Chemother. 1999, 43, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Sanz, E.; Haro-Moreno, J.M.; Jensen, S.O.; Roda-Garcia, J.J.; Lopez-Perez, M. Staphylococcus sciuri C2865 from a distinct subspecies cluster as reservoir of the novel transferable trimethoprim resistance gene, dfrE, and adaptation driving mobile elements. bioRxiv 2020. [Google Scholar] [CrossRef]
- Lee, E.W.; Chen, J.; Huda, M.N.; Kuroda, T.; Mizushima, T.; Tsuchiya, T. Functional cloning and expression of emeA, and characterization of EmeA, a multidrug efflux pump from Enterococcus faecalis. Biol. Pharm. Bull. 2003, 26, 266–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerma, L.L.; Benomar, N.; Valenzuela, A.S.; Muñoz, M.d.C.C.; Gálvez, A.; Abriouel, H. Role of EfrAB efflux pump in biocide tolerance and antibiotic resistance of Enterococcus faecalis and Enterococcus faecium isolated from traditional fermented foods and the effect of EDTA as EfrAB inhibitor. Food Microbiol. 2014, 44, 249–257. [Google Scholar] [CrossRef]
- Singh, K.V.; Weinstock, G.M.; Murray, B.E. An Enterococcus faecalis ABC homologue (Lsa) is required for the resistance of this species to clindamycin and quinupristin-dalfopristin. Antimicrob. Agents Chemother. 2002, 46, 1845–1850. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, M.; Hirt, H.; Zurek, L. Horizontal transfer of the tetracycline resistance gene tetM mediated by pCF10 among Enterococcus faecalis in the house fly (Musca domestica L.) alimentary canal. Microb. Ecol. 2009, 58, 509–518. [Google Scholar] [CrossRef]
- Malbruny, B.; Werno, A.M.; Murdoch, D.R.; Leclercq, R.; Cattoir, V. Cross-resistance to lincosamides, streptogramins A, and pleuromutilins due to the lsa (C) gene in Streptococcus agalactiae UCN70. Antimicrob. Agents Chemother. 2011, 55, 1470–1474. [Google Scholar] [CrossRef] [Green Version]
- Haubert, L.; da Cunha, C.E.P.; Lopes, G.V.; da Silva, W.P. Food isolate Listeria monocytogenes harboring tetM gene plasmid-mediated exchangeable to Enterococcus faecalis on the surface of processed cheese. Food Res. Int. 2018, 107, 503–508. [Google Scholar] [CrossRef]
- Gabashvili, E.; Osepashvili, M.; Koulouris, S.; Ujmajuridze, L.; Tskhitishvili, Z.; Kotetishvili, M. Phage Transduction is Involved in the Intergeneric Spread of Antibiotic Resistance-Associated bla CTX-M, mel, and tetM Loci in Natural Populations of Some Human and Animal Bacterial Pathogens. Curr. Microbiol. 2020, 77, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Costa, Y.; Galimand, M.; Leclercq, R.; Duval, J.; Courvalin, P. Characterization of the chromosomal aac (6’)-Ii gene specific for Enterococcus faecium. Antimicrob. Agents Chemother. 1993, 37, 1896–1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isnard, C.; Malbruny, B.; Leclercq, R.; Cattoir, V. Genetic basis for in vitro and in vivo resistance to lincosamides, streptogramins A, and pleuromutilins (LSAP phenotype) in Enterococcus faecium. Antimicrob. Agents Chemother. 2013, 57, 4463–4469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.V.; Malathum, K.; Murray, B.E. Disruption of an Enterococcus faecium species-specific gene, a homologue of acquired macrolide resistance genes of staphylococci, is associated with an increase in macrolide susceptibility. Antimicrob. Agents Chemother. 2001, 45, 263–266. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, E.D.; Cove, J.H. Resistance to telithromycin is conferred by msr (A), msrC and msr (D) in Staphylococcus aureus. J. Antimicrob. Chemother. 2005, 56, 1179–1180. [Google Scholar] [CrossRef]
- Flórez, A.B.; de Los Reyes-Gavilán, C.G.; Wind, A.; Mayo, B.; Margolles, A. Ubiquity and diversity of multidrug resistance genes in Lactococcus lactis strains isolated between 1936 and 1995. FEMS Microbiol. Lett. 2006, 263, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Esaiassen, E.; Hjerde, E.; Cavanagh, J.P.; Simonsen, G.S.; Klingenberg, C. Bifidobacterium bacteremia: Clinical characteristics and a genomic approach to assess pathogenicity. J. Clin. Microbiol. 2017, 55, 2234–2248. [Google Scholar] [CrossRef] [Green Version]
- Heuer, H.; Krögerrecklenfort, E.; Wellington, E.; Egan, S.; Van Elsas, J.; Van Overbeek, L.; Collard, J.M.; Guillaume, G.; Karagouni, A.; Nikolakopoulou, T.; et al. Gentamicin resistance genes in environmental bacteria: Prevalence and transfer. FEMS Microbiol. Ecol. 2002, 42, 289–302. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Ding, L.J.; Fan, M.Z. Resistance patterns and detection of aac (3)-IV gene in apramycin-resistant Escherichia coli isolated from farm animals and farm workers in northeastern of China. Res. Vet. Sci. 2009, 87, 449–454. [Google Scholar] [CrossRef]
- Paterson, D.L.; Bonomo, R.A. Extended-spectrum β-lactamases: A clinical update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [Green Version]
- Rawat, D.; Nair, D. Extended-spectrum β-lactamases in Gram Negative Bacteria. J. Glob. Infect. Dis. 2010, 2, 263. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Bae, I.K.; Lee, J.H.; Sohn, S.G.; Kang, G.H.; Jeon, G.J.; Kim, Y.H.; Jeong, B.C.; Lee, S.H. Molecular characterization of extended-spectrum beta-lactamases produced by clinical isolates of Klebsiella pneumoniae and Escherichia coli from a Korean nationwide survey. J. Clin. Microbiol. 2004, 42, 2902–2906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahlaoui, H.; Dahmen, S.; Moussa, M.; Omrane, B. First detection of TEM-116 extended-spectrum β-lactamase in a Providencia stuartii isolate from a Tunisian hospital. Indian J. Med Microbiol. 2011, 29, 258. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.L.; Tang, S.I.; Fang, C.T.; Hsueh, P.R.; Chang, S.C.; Wang, J.T. Extended-spectrum β-lactamase genes of Klebsiella pneumoniae strains in Taiwan: Recharacterization of shv-27, shv-41, and tem-116. Microb. Drug Resist. 2006, 12, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.; Messing, J. The pUC plasmids, an M13mp7-derived system for insertion mutagenesis and sequencing with synthetic universal primers. Gene 1982, 19, 259–268. [Google Scholar] [CrossRef]
- Koncan, R.; Valverde, A.; Morosini, M.I.; García-Castillo, M.; Cantón, R.; Cornaglia, G.; Baquero, F.; del Campo, R. Learning from mistakes: Taq polymerase contaminated with β-lactamase sequences results in false emergence of Streptococcus pneumoniae containing TEM. J. Antimicrob. Chemother. 2007, 60, 702–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, J.L.; Coque, T.M.; Baquero, F. What is a resistance gene? Ranking risk in resistomes. Nat. Rev. Microbiol. 2015, 13, 116–123. [Google Scholar] [CrossRef] [PubMed]
- CDC. Outpatient Antibiotic Prescriptions; CDC: Atlanta, GA, USA, 2018.
- WHO. WHO Report on Surveillance of Antibiotic Consumption: 2016–2018 Early Implementation; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Nishino, K.; Yamaguchi, A. Role of histone-like protein H-NS in multidrug resistance of Escherichia coli. J. Bacteriol. 2004, 186, 1423–1429. [Google Scholar] [CrossRef] [Green Version]
- Founou, L.L.; Allam, M.; Ismail, A.; Essack, S.Y. Enterococcus faecalis ST21 harbouring Tn6009 isolated from a carriage sample in South Africa. S. Afr. Med J. 2021, 111, 98–99. [Google Scholar] [CrossRef]
- Ammor, M.S.; Florez, A.B.; Álvarez-Martín, P.; Margolles, A.; Mayo, B. Analysis of tetracycline resistance tet (W) genes and their flanking sequences in intestinal Bifidobacterium species. J. Antimicrob. Chemother. 2008, 62, 688–693. [Google Scholar] [CrossRef] [Green Version]
- Duranti, S.; Lugli, G.A.; Mancabelli, L.; Turroni, F.; Milani, C.; Mangifesta, M.; Ferrario, C.; Anzalone, R.; Viappiani, A.; van Sinderen, D.; et al. Prevalence of antibiotic resistance genes among human gut-derived bifidobacteria. Appl. Environ. Microbiol. 2017, 83, e02894-16. [Google Scholar] [CrossRef] [Green Version]
- Kazimierczak, K.A.; Flint, H.J.; Scott, K.P. Comparative analysis of sequences flanking tet (W) resistance genes in multiple species of gut bacteria. Antimicrob. Agents Chemother. 2006, 50, 2632–2639. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Tolmasky, M.E. Amikacin: Uses, resistance, and prospects for inhibition. Molecules 2017, 22, 2267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, G.; Hildebrandt, B.; Witte, W. The newly described msrC gene is not equally distributed among all isolates of Enterococcus faecium. Antimicrob. Agents Chemother. 2001, 45, 3672–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overdevest, I.; Willemsen, I.; Rijnsburger, M.; Eustace, A.; Xu, L.; Hawkey, P.; Heck, M.; Savelkoul, P.; Vandenbroucke-Grauls, C.; van der Zwaluw, K.; et al. Extended-spectrum β-lactamase genes of Escherichia coli in chicken meat and humans, The Netherlands. Emerg. Infect. Dis. 2011, 17, 1216. [Google Scholar] [CrossRef]
- Kamaruzzaman, E.A.; Abdul Aziz, S.; Bitrus, A.A.; Zakaria, Z.; Hassan, L. Occurrence and Characteristics of Extended-Spectrum β-Lactamase-Producing Escherichia coli from Dairy Cattle, Milk, and Farm Environments in Peninsular Malaysia. Pathogens 2020, 9, 1007. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, Y.; Valverde, A.; Cason, E.D.; Pierneef, R.E.; Cowan, D.A. A clinically important, plasmid-borne antibiotic resistance gene (β-lactamase TEM-116) present in desert soils. Sci. Total. Environ. 2020, 719, 137497. [Google Scholar] [CrossRef]
- Sharma, P.; Haycocks, J.R.; Middlemiss, A.D.; Kettles, R.A.; Sellars, L.E.; Ricci, V.; Piddock, L.J.; Grainger, D.C. The multiple antibiotic resistance operon of enteric bacteria controls DNA repair and outer membrane integrity. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panthee, S.; Paudel, A.; Hamamoto, H.; Ogasawara, A.A.; Iwasa, T.; Blom, J.; Sekimizu, K. Complete genome sequence and comparative genomic analysis of Enterococcus faecalis EF-2001, a probiotic bacterium. Genomics 2021, 113, 1534–1542. [Google Scholar] [CrossRef]
- Bozdogan, B.; Galopin, S.; Gerbaud, G.; Courvalin, P.; Leclercq, R. Chromosomal aadD2 encodes an aminoglycoside nucleotidyltransferase in Bacillus clausii. Antimicrob. Agents Chemother. 2003, 47, 1343–1346. [Google Scholar] [CrossRef] [Green Version]
- Courvalin, P. Antibiotic resistance: The pros and cons of probiotics. Dig. Liver Dis. 2006, 38, S261–S265. [Google Scholar] [CrossRef]
- Gaeta, N.C.; Bean, E.; Miles, A.M.; de Carvalho, D.U.O.G.; Alemán, M.A.R.; Carvalho, J.S.; Gregory, L.; Ganda, E. A cross-sectional study of dairy cattle metagenomes reveals increased antimicrobial resistance in animals farmed in a heavy metal contaminated environment. Front. Microbiol. 2020, 11, 590325. [Google Scholar] [CrossRef]
- Shi, L.D.; Xu, Q.J.; Liu, J.Y.; Han, Z.X.; Zhu, Y.G.; Zhao, H.P. Will a Non-antibiotic Metalloid Enhance the Spread of Antibiotic Resistance Genes: The Selenate Story. Environ. Sci. Technol. 2020, 55, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Seol, D.; Jhang, S.Y.; Kim, H.; Kim, S.Y.; Kwak, H.S.; Kim, S.H.; Lee, W.; Park, S.; Kim, H.; Cho, S.; et al. Accurate and strict identification of probiotic species based on coverage of whole-metagenome shotgun sequencing data. Front. Microbiol. 2019, 10, 1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sample ID | BioProject | Run | Description |
|---|---|---|---|
| Isolates | |||
| s01 | PRJEB14693 | ERR1554589 | Lactiplantibacillus plantarum |
| s02 | PRJEB14693 | ERR1554590 | Lactiplantibacillus plantarum |
| s03 | PRJEB14693 | ERR1554591 | Lactiplantibacillus plantarum |
| s04 | PRJEB38007 | ERR4421718 | Pseudomonas sp. RGM2144 |
| s05 | PRJNA312743 | SRR3205957 | Limosilactobacillus fermentum |
| s06 | PRJNA347617 | SRR4417252 | Limosilactobacillus fermentum |
| s07 | PRJNA635872 | SRR11966381 | Lactiplantibacillus plantarum |
| s08 | PRJNA639653 | SRR12037315 | Lactobacillus delbrueckii subsp. bulgaricus |
| s09 | PRJNA639653 | SRR12037316 | Lactobacillus delbrueckii subsp. bulgaricus |
| s10 | PRJNA639653 | SRR12037890 | Streptococcus thermophilus |
| s11 | PRJNA649814 | SRR12375795 | Enterococcus faecalis |
| s12 | PRJNA649814 | SRR12375796 | Enterococcus faecalis |
| s13 | PRJNA649814 | SRR12375797 | Enterococcus faecalis |
| s14 | PRJNA650131 | SRR12376423 | Escherichia coli |
| s15 | PRJNA650131 | SRR12376425 | Escherichia coli |
| s16 | PRJNA650131 | SRR12376427 | Escherichia coli |
| s17 | PRJNA650131 | SRR12376429 | Escherichia coli |
| s18 | PRJNA650131 | SRR12376431 | Escherichia coli |
| s19 | PRJNA650131 | SRR12376433 | Escherichia coli |
| s20 | PRJNA639653 | SRR12412204 | Lacticaseibacillus rhamnosus |
| Microbiota | |||
| m01 | PRJNA474998 | SRR8132838 | probiotic powder (FC13678) |
| m02 | PRJNA475000 | SRR8138827 | probiotic powder (FC13669) |
| m03 | PRJNA474989 | SRR8140233 | probiotic powder (FC13655) |
| m04 | PRJNA474995 | SRR8140386 | probiotic powder (FC13628) |
| m05 * | PRJNA508569 | SRR8289759 | probiotic product (2) |
| m06 | PRJNA508569 | SRR8289760 | probiotic product (1) |
| m07 * | PRJNA508569 | SRR8289761 | probiotic product (4) |
| m08 * | PRJNA508569 | SRR8289762 | probiotic product (3) |
| m09 * | PRJNA508569 | SRR8289763 | probiotic product (6) |
| m10 * | PRJNA508569 | SRR8289764 | probiotic product (5) |
| m11 | PRJNA542229 | SRR9040978 | dietary supplement (PB4) |
| m12 | PRJNA542229 | SRR9040979 | dietary supplement (PB10) |
| m13 | PRJNA542229 | SRR9040980 | dietary supplement (PB11) |
| m14 | PRJNA542229 | SRR9040981 | dietary supplement (PB2) |
| m15 | PRJNA542229 | SRR9040982 | dietary supplement (PB14) |
| m16 | PRJNA542229 | SRR9040983 | dietary supplement (PB13) |
| m17 | PRJNA542229 | SRR9040984 | dietary supplement (PB16) |
| m18 | PRJNA542229 | SRR9040986 | dietary supplement (PB18) |
| m19 | PRJNA542229 | SRR9040987 | dietary supplement (PB17) |
| m20 | PRJNA542229 | SRR9040988 | dietary supplement (PB8) |
| m21 | PRJNA542229 | SRR9040989 | dietary supplement (PB19) |
| m22 | PRJNA542229 | SRR9040990 | dietary supplement (PB12) |
| m23 | PRJNA542229 | SRR9040991 | dietary supplement (PB9) |
| m24 | PRJNA542229 | SRR9040992 | dietary supplement (PB6) |
| m25 | PRJNA542229 | SRR9040993 | dietary supplement (PB5) |
| m26 | PRJNA542229 | SRR9040994 | dietary supplement (PB7) |
| m27 | PRJNA644361 | SRR12153424 | probiotic capsule |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tóth, A.G.; Csabai, I.; Judge, M.F.; Maróti, G.; Becsei, Á.; Spisák, S.; Solymosi, N. Mobile Antimicrobial Resistance Genes in Probiotics. Antibiotics 2021, 10, 1287. https://doi.org/10.3390/antibiotics10111287
Tóth AG, Csabai I, Judge MF, Maróti G, Becsei Á, Spisák S, Solymosi N. Mobile Antimicrobial Resistance Genes in Probiotics. Antibiotics. 2021; 10(11):1287. https://doi.org/10.3390/antibiotics10111287
Chicago/Turabian StyleTóth, Adrienn Gréta, István Csabai, Maura Fiona Judge, Gergely Maróti, Ágnes Becsei, Sándor Spisák, and Norbert Solymosi. 2021. "Mobile Antimicrobial Resistance Genes in Probiotics" Antibiotics 10, no. 11: 1287. https://doi.org/10.3390/antibiotics10111287
APA StyleTóth, A. G., Csabai, I., Judge, M. F., Maróti, G., Becsei, Á., Spisák, S., & Solymosi, N. (2021). Mobile Antimicrobial Resistance Genes in Probiotics. Antibiotics, 10(11), 1287. https://doi.org/10.3390/antibiotics10111287