
Antitrypanosomal Activity of Anthriscus Nemorosa Essential Oils and Combinations of Their Main Constituents



 ,
, 
 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
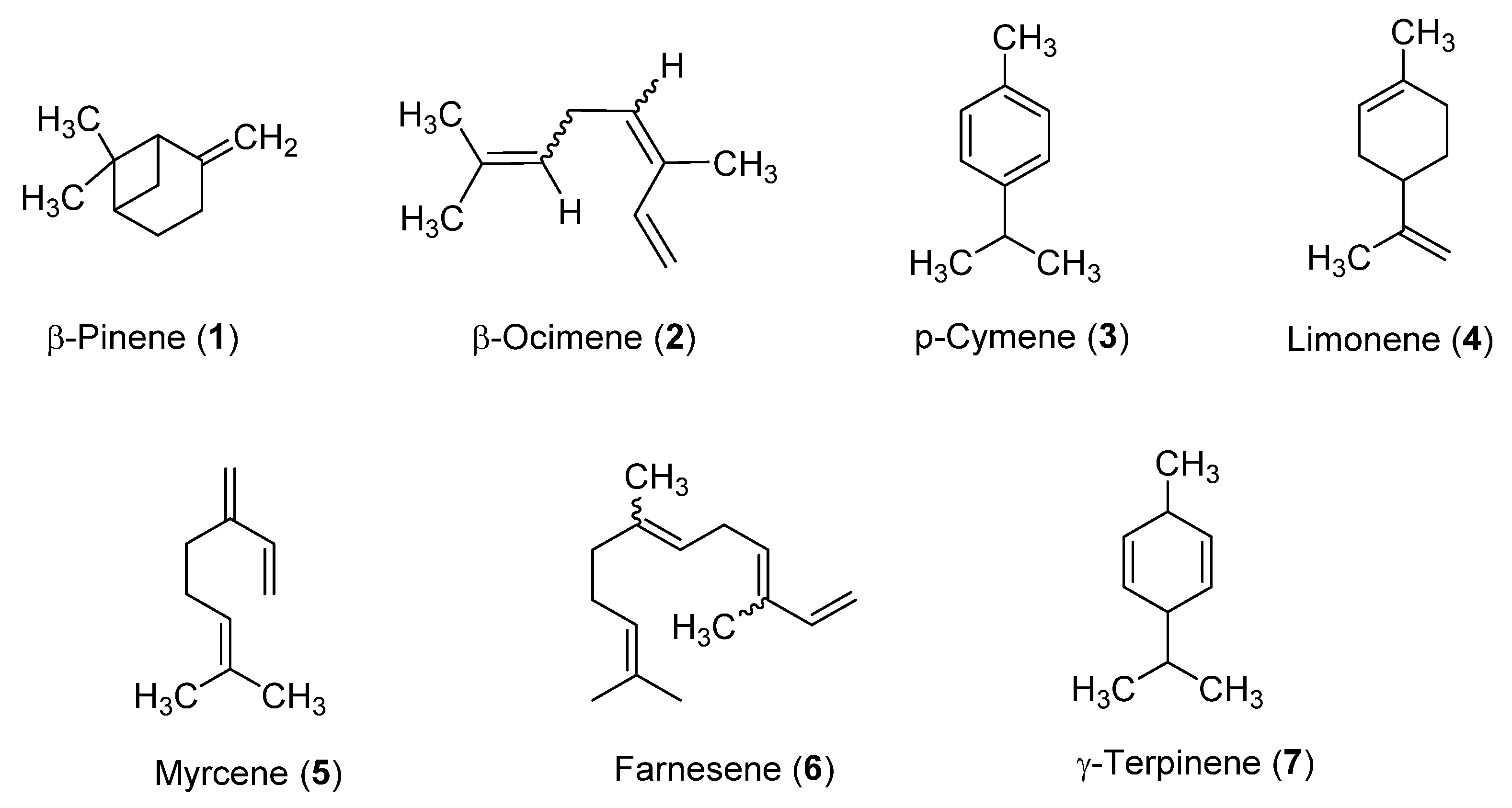
2.1. EO Compositions
2.2. Antitrypanosomal Activity
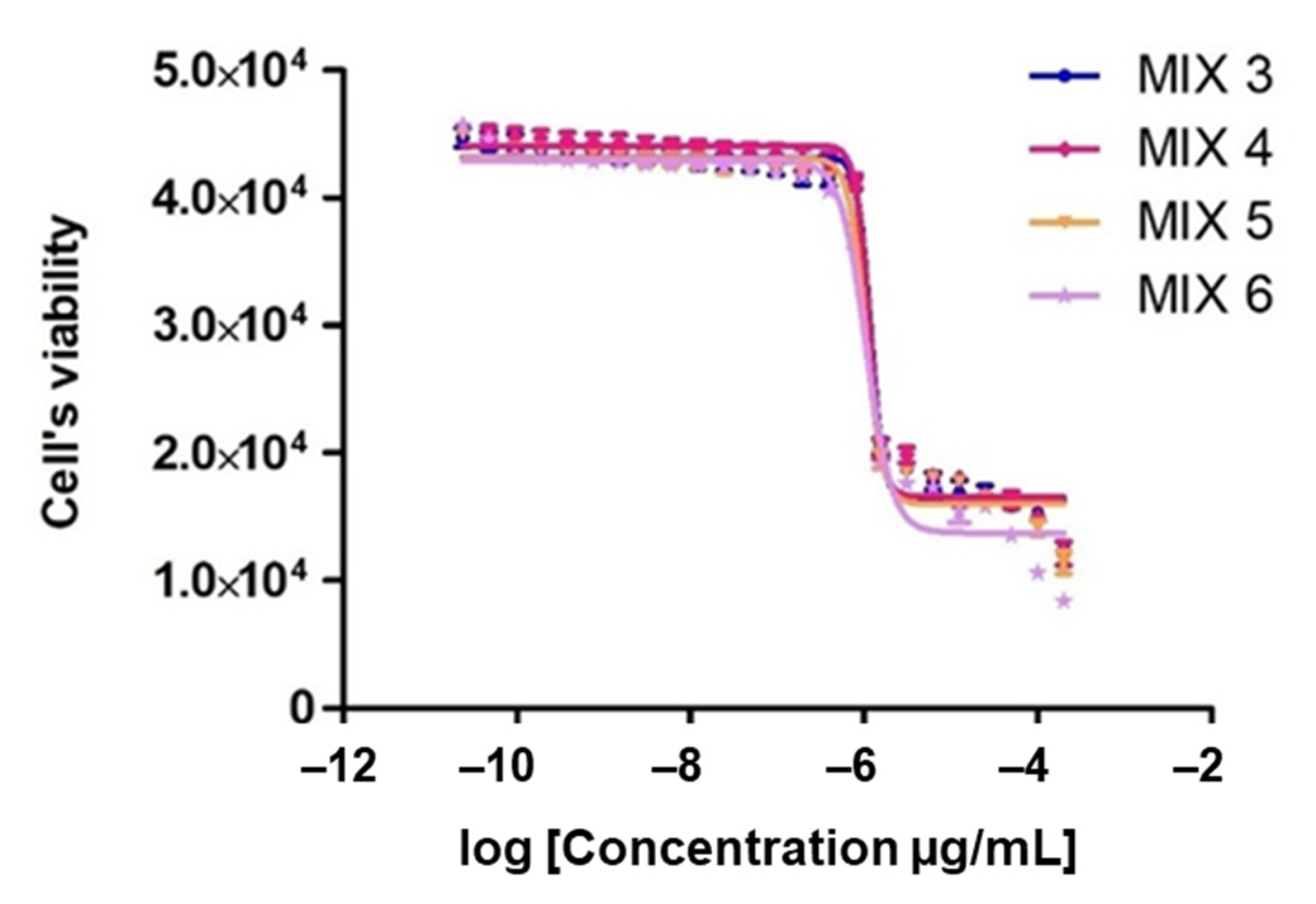
2.3. Contribution of Individual Compounds to the Overall Antitrypanosomal Activity and Synergistic Effects
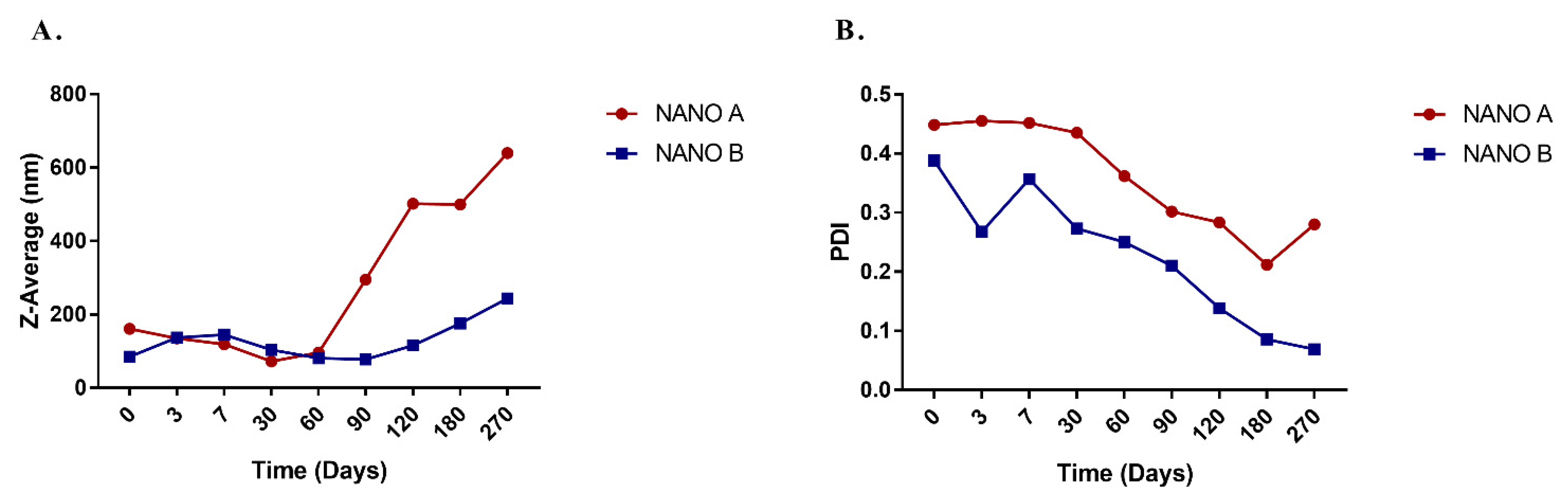
2.4. Preparation and Characterization of A. nemorosa EO-Based Nanoemulsions
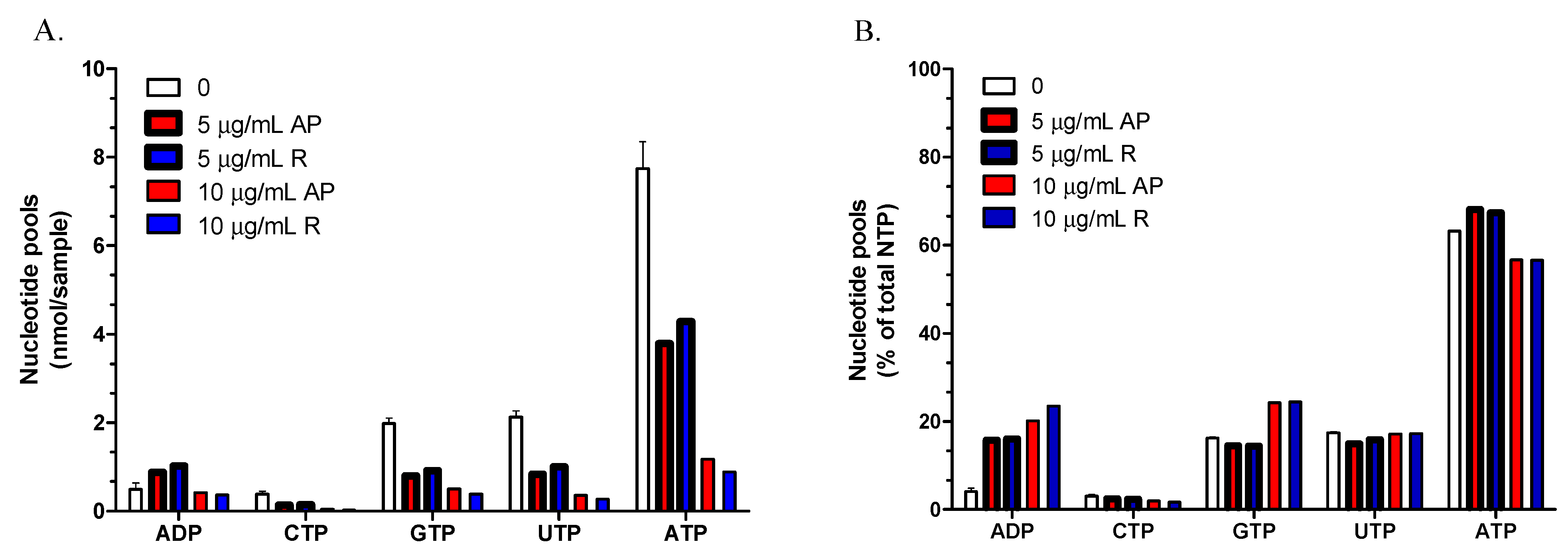
2.5. Determination of NTP and ADP Pools in T. brucei
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Chemicals
4.3. Hydrodistillation
4.4. GC-MS Analysis
4.5. T. brucei and Mammalian Cell Culture
4.6. Growth Inhibition Assay on T. brucei and Balb/3T3 Cells
4.7. Comparative Activities
4.8. Synergistic Interactions among Four Major Constituents
4.9. Preparation of Anthriscus nemorosa EO-Based Nanoemulsions
4.10. Nanoemulsion Characterization
4.11. Determination of NTP and ADP Pools in T. brucei
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Barrett, M.P.; Burchmore, R.J.; Stich, A.; Lazzari, J.O.; Frasch, A.C.; Cazzulo, J.J.; Krishna, S. The trypanosomiases. Lancet 2003, 362, 1469–1480. [Google Scholar] [CrossRef]
- Simarro, P.P.; Cecchi, G.; Paone, M.; Franco, J.R.; Diarra, A.; Ruiz, J.A.; Fèvre, E.M.; Courtin, F.; Mattioli, R.C.; Jannin, J.G. The Atlas of Human African Trypanosomiasis: A Contribution to Global Mapping of Neglected Tropical Diseases. Int. J. Health Geogr. 2010, 9, 57–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzoe, F.A. Current situation of African trypanosomiasis. Acta Trop. 1993, 5, 153–162. [Google Scholar] [CrossRef]
- World Health Organization: African Trypanosomiasis (Sleeping Sickness); World Health Organization Fact Sheet; World Health Organization: Geneva, Switzerland, 2006; Volume 259.
- Lindner, A.K.; Lejon, V.; Chappuis, F.; Seixas, J.; Kazumba, L.; Barrett, M.P.; Mwamba, E.; Erphas, O.; Akl, E.A.; Villanueva, G.; et al. New WHO guidelines for treatment of gambiense human African trypanosomiasis including fexinidazole: Substantial changes for clinical practice. Lancet Infect. Dis. 2020, 20, e38–e46. [Google Scholar] [CrossRef]
- Kamte, S.L.N.; Ranjbarian, F.; Cianfaglione, K.; Sut, S.; Dall’Acqua, S.; Bruno, M.; Afshar, F.H.; Iannarelli, R.; Benelli, G.; Cappellacci, L.; et al. Identification of highly effective antitrypanosomal compounds in essential oils from the Apiaceae family. Ecotoxicol. Environ. Saf. 2018, 156, 154–165. [Google Scholar] [CrossRef]
- Petrelli, R.; Ranjbarian, F.; Dall’Acqua, S.; Papa, F.; Iannarelli, R.; Kamte, S.L.N.; Vittori, S.; Benelli, G.; Maggi, F.; Hofer, A.; et al. An overlooked horticultural crop, Smyrnium olusatrum, as a potential source of compounds effective against African trypanosomiasis. Parasitol. Int. 2017, 66, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Altundag, E.; Ozturk, M. Ethnomedicinal studies on the plant resources of east Anatolia, Turkey. Procedia Soc. Behav. Sci. 2011, 19, 756–777. [Google Scholar] [CrossRef] [Green Version]
- Nikolić, V. Anthriscus Pers. In Flora SR Srbije; Josifović, M., Ed.; Srpska Akademija Nauka i Umetnosti: Beograd, Serbia, 1973; Volume 5, pp. 334–338. [Google Scholar]
- Navchoo, I.A.; Buth, G.M. Medicinal system of Ladakh, India. J. Ethnopharmacol. 1989, 26, 137–146. [Google Scholar] [CrossRef]
- Gairola, S.; Sharma, J.; Bedi, Y.S. A cross-cultural analysis of Jammu, Kashmir and Ladakh (India) medicinal plant use. J. Ethnopharmacol. 2014, 155, 925–986. [Google Scholar] [CrossRef]
- Karakaya, S.; Yılmaz, S.V.; Koca, M.; Demirci, B.; Sytar, O. Screening of non-alkaloid acetylcholinesterase inhibitors from extracts and essential oils of Anthriscus nemorosa (M. Bieb.) Spreng.(Apiaceae). S. Afr. J. Bot. 2019, 125, 261–269. [Google Scholar] [CrossRef]
- Bagci, E.; Aydin, E.; Ungureanu, E.; Hritcu, L. Anthriscus nemorosa essential oil inhalation prevents memory impairment, anxiety and depression in scopolamine-treated rats. Biomed. Pharmacother. 2016, 84, 1313–1320. [Google Scholar] [CrossRef] [PubMed]
- Menemen, Y. Anthriscus Pers. In Türkiye Bitkileri Listesi (Damarlı Bitkiler); Güner, A., Aslan, S., Ekim, T., Vural, M., Babaç, M.T., Eds.; Nezahat Gökyiğit Botanic Garden: İstanbul, Turkey, 2012; pp. 62–64. [Google Scholar]
- Karimi, S.; Farzaneh, F.; Asnaashari, S.; Parina, P.; Sarvari, Y.; Hazrati, S. Phytochemical Analysis and Anti-microbial Activity of Some Important Medicinal Plants from North-west of Iran. Iranian J. Pharm. Res. IJPR 2019, 18, 1871. [Google Scholar]
- Van Den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Petrelli, R.; Orsomando, G.; Sorci, L.; Maggi, F.; Ranjbarian, F.; Biapa Nya, P.C.; Petrelli, D.; Vitali, L.A.; Lupidi, G.; Quassinti, L.; et al. Biological Activities of the Essential Oil from Erigeron floribundus. Molecules 2016, 21, 1065. [Google Scholar] [CrossRef]
- Ngahang Kamte, S.L.; Ranjbarian, F.; Campagnaro, G.D.; Biapa Nya, P.C.; Mbuntcha, H.; Woguem, V.; Womeni, H.M.; Azefack Tapondjou, L.; Giordani, C.; Barboni, L.; et al. Trypanosoma brucei inhibition by essential oils from medicinal aromatic plants traditionally used in Cameroon (Azadirachta indica, Aframomum melegueta, Aframomum daniellii, Clausena anisata, Dichrostachys cinerea and Echinops giganteus). Int. J. Environ. Res. Public Health 2017, 14, 737. [Google Scholar] [CrossRef]
- Sadeghi, R.; Etemad, S.G.; Keshavarzi, E.; Haghshenasfard, M. Investigation of alumina nanofluid stability by UV–vis spectrum. Microfluid. Nanofluid. 2015, 18, 1023–1030. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Nickavar, B.; Mojab, F.; Mojahedi, A. Composition of the essential oil from Anthriscus nemorosa. Chem. Nat. Compd. 2009, 45, 443–444. [Google Scholar] [CrossRef]
- Kiliç, Ö. Essential oil composition of aerial parts of two Anthriscus Pers. species from Turkey. TEOP 2017, 20, 591–596. [Google Scholar] [CrossRef]
- Wagner, H.; Ulrich-Merzenich, G. Synergy research: Approaching a new generation of phytopharmaceuticals. Phytomedicine 2009, 16, 97–110. [Google Scholar] [CrossRef]
- Wink, M. Evolutionary advantage and molecular modes of action of multi-component mixtures used in phytomedicine. Curr. Drug Metab. 2008, 10, 996–1009. [Google Scholar] [CrossRef]
- Monzote, L.; Alarcon, O.; Setzer, W.N. Antiprotozoal activity of essential oils. Agric. Conspec. Sci. 2012, 77, 167–175. [Google Scholar]
- Cascone, P.; Iodice, L.; Maffei, M.E.; Bossi, S.; Arimura, G.-I.; Guerrieri, E. Tobacco overexpressing β-ocimene induces direct and indirect responses against aphids in receiver tomato plants. J. Plant Physiol. 2015, 173, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Bouyahya, A.; Et-Touys, A.; Bakri, Y.; Talbaui, A.; Fellah, H.; Abrini, J.; Dakka, N. Chemical composition of Mentha pulegium and Rosmarinus officinalis essential oils and their antileishmanial, antibacterial and antioxidant activities. Microb. Pathog. 2017, 111, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Quassinti, L.; Bramucci, M.; Lupidi, G.; Barboni, L.; Ricciutelli, M.; Sagratini, G.; Papa, F.; Caprioli, G.; Petrelli, D.; Vitali, L.A.; et al. In vitro biological activity of essential oils and isolated furanosesquiterpenes from the neglected vegetable Smyrnium olusatrum L. (Apiaceae). Food Chem. 2013, 138, 808–813. [Google Scholar] [CrossRef]
- Miresmailli, S.; Bradbury, R.; Isman, M.B. Comparative toxicity of Rosmarinus officinalis L. essential oil and blends of its major constituents against Tetranychus urticae Koch (Acari: Tetranychidae) on two different host plants. Pest Manag. Sci. 2006, 62, 366–371. [Google Scholar] [CrossRef]
- Don-Pedro, K.N. Investigation of single and joint fumigant insecticidal action of citrus peel oil components. Pest Sci. 1996, 46, 79–84. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Skourti, A.; Nika, E.P.; Ntalaka, C.T.; Boukouvala, M.C.; Bonacucina, G.; Cespi, M.; Petrelli, R.; Cappellacci, L.; Maggi, F.; et al. Isofuranodiene-based nanoemulsion: Larvicidal and adulticidal activity against tenebrionid beetles attacking stored wheat. J. Stored Prod. Res. 2021, 93, 101859. [Google Scholar] [CrossRef]
- Hofer, A.; Ekanem, J.T.E.; Thelander, L. Allosteric regulation of Trypanosoma brucei ribonucleotide reductase studied in vitro and in vivo. J. Biol. Chem. 1998, 273, 34098–34104. [Google Scholar] [CrossRef] [Green Version]
- Ranjbarian, F.; Sharma, S.; Falappa, G.; Taruschio, W.; Chabes, A.; Hofer, A. Isocratic HPLC analysis for the simultaneous determination of dNTPs, rNTPs, and ADP in biological samples. Nucleic Acids Res. 2021. accepted. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Component a | RI b | RI Lit. c | % A. nemorosa EO-AP | % A. nemorosa EO-R | ID d |
|---|---|---|---|---|---|---|
| 1 | n-nonane | 901 | 900 | 1.7 | 9.5 | RI, MS |
| 2 | α-thujene | 921 | 924 | Tr e | tr | RI, MS |
| 3 | α-pinene | 927 | 932 | 0.4 | 1.7 | Std |
| 4 | sabinene | 964 | 969 | 0.4 | 0.4 | Std |
| 5 | β-pinene | 969 | 974 | 3.0 | 22.0 | Std |
| 6 | myrcene | 990 | 988 | 40.1 | 19.9 | Std |
| 7 | n-decane | 1001 | 1000 | 0.1 | 0.8 | RI, MS |
| 8 | α-phellandrene | 1004 | 1002 | 0.2 | Std | |
| 9 | α-terpinene | 1015 | 1014 | 0.1 | tr | Std |
| 10 | p-cymene | 1022 | 1020 | 2.2 | 2.8 | Std |
| 11 | limonene | 1024 | 1024 | 5.7 | 11.5 | Std |
| 12 | β-phellandrene | 1026 | 1025 | 3.0 | 2.7 | Std |
| 13 | (Z)-β-ocimene | 1038 | 1032 | 4.4 | 8.9 | Std |
| 14 | (E)-β-ocimene | 1048 | 1044 | 2.8 | 1.8 | Std |
| 15 | γ-terpinene | 1056 | 1055 | 11.6 | 9.2 | Std |
| 16 | n-undecane | 1101 | 1100 | 0.4 | 0.9 | RI, MS |
| 17 | allo-ocimene | 1129 | 1128 | 0.2 | 0.5 | RI, MS |
| 18 | β-elemene | 1384 | 1389 | 0.4 | tr | RI, MS |
| 19 | (E)-caryophyllene | 1387 | 1417 | 0.6 | 0.1 | Std |
| 20 | β-copaene | 1420 | 1430 | 0.5 | RI, MS | |
| 21 | (E)-β-farnesene | 1457 | 1440 | 0.7 | Std | |
| 22 | germacrene D | 1472 | 1484 | 10.4 | RI, MS | |
| 23 | 1-dodecanol | 1476 | 1469 | 0.5 | RI, MS | |
| 24 | epi-cubebol | 1487 | 1493 | 0.3 | RI, MS | |
| 25 | (E,E)-α-farnesene | 1508 | 1505 | 4.0 | RI, MS | |
| 26 | δ-cadinene | 1517 | 1522 | 2.3 | 0.9 | RI, MS |
| 27 | elemol | 1543 | 1534 | 0.5 | RI, MS | |
| 28 | guaiol | 1591 | 1600 | 0.8 | RI, MS | |
| 29 | 10-epi-γ-eudesmol | 1608 | 1622 | 0.5 | RI, MS | |
| 30 | epi-α-muurolol + epi-α-cadinol | 1633 | 1640/1638 | 1.0 | 0.5 | RI, MS |
| 31 | β-eudesmol | 1639 | 1649 | 0.2 | 0.4 | RI, MS |
| 32 | valerianol | 1643 | 1656 | 0.1 | 0.4 | RI, MS |
| 33 | α-cadinol | 1646 | 1652 | 1.7 | 0.4 | RI, MS |
| 34 | bulnesol | 1659 | 1670 | 0.7 | RI, MS | |
| Oil yield (%, w/w) | 0.06 | 0.08 | ||||
| Total identified (%) | 98.1 | 98.6 | ||||
| Grouped compunds (%) | ||||||
| Monoterpene hydrocarbons | 74.0 | 81.5 | ||||
| Oxygenated monoterpenes | ||||||
| Sesquiterpenes hydrocarbons | 18.8 | 1.0 | ||||
| Oxygenated sesquiterpenes | 3.0 | 4.4 | ||||
| Alkanes | 2.3 | 11.2 | ||||
| Others | 0.5 |
| Samples | EC50 | Selectivity Index (SI) | |
|---|---|---|---|
| T. brucei (s427) | Balb3T3 | ||
| Essential oils | μg/mL | μg/mL | |
| A. nemorosa aerial parts | 1.17 ± 0.09 | 5.48 ± 0.61 | 4.65 |
| A. nemorosa roots | 2.36 ± 0.09 | 5.33 ± 0.49 | 2.25 |
| Pure compounds | μg/mL(μM) | μg/mL(μM) | |
| β-Pinene (1): 3% AP, 22% R | 11.4 ± 2.6 (83.7) c | >100 | >8.77 |
| β-Ocimene (2): 7,2% AP, 10.7% R | 1.1 ± 0.5 (8) b | >100 | >91 |
| p-Cymene (3): 2.2% AP, 2.8% R | 4.5 ± 1.0 (33) b | 28 ± 7 | 6.2 |
| Limonene (4): 5.7% AP, 11.5% R | 5.6 ± 1.6 (41) a | >100 | >18 |
| Myrcene (5): 40.1% AP, 19.9% R | 26.2 ± 0.44 (193) | 49.6 ± 0.64 | 1.9 |
| Farnesene (6): 4.7% AP | 0.84 ± 0.04 (4.11) | 4.09 ± 0.39 | 4.85 |
| γ-Terpinene (7): 11.6% AP, 9.2% R | >100 | >100 | - |
| Reference drug | μg/mL(μM) | μg/mL(μM) | |
| Suramin | 0.025 ± 0.001 (0.0147) | >5 | >262 |
| Ratio% w/w | Observed T.b. brucei EC50 (μg/mL) | Observed Balb3T3 EC50 (μg/mL) | Selectivity Index (SI) | Expected EC50 a (μg/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Myrcene | β-Pinene | Limonene | β-Ocimene | p-Cymene | Farnesene | Carrier (Acetone) | Wadley b | R c | S d | ||||
| Mix 1 | 63.7 | 4.7 | 9.1 | 11.4 | 3.7 | 7.4 | x | 7.25 ± 0.26 | 28.3 ± 1.06 | 3.91 | 4.09 | 0.56 | Add |
| Mix 2 | x | 4.7 | 9.1 | 11.4 | 3.7 | 7.4 | 63.7 | 2.25 ± 0.18 | 9.22 ± 0.67 | 4.11 | 4.54 | 2.01 | Syn |
| Mix 3 | x | x | 9.1 | 11.4 | 3.7 | 7.4 | 68.4 | 1.43 ± 0.16 | 5.14 ± 0.36 | 3.63 | 4.63 | 3.23 | Syn |
| Mix 4 | x | x | x | 11.4 | 3.7 | 7.4 | 77.5 | 1.58 ± 0.14 | 5.48 ± 0.31 | 3.47 | 5.00 | 3.17 | Syn |
| Mix 5 | x | x | 9.1 | 11.4 | 3.7 | x | 75.8 | 1.27 ± 0.02 | 6.98 ± 0.85 | 5.46 | 7.80 | 6.14 | Syn |
| Mix 6 | x | x | 9.1 | 11.4 | x | 7.4 | 72.1 | 1.06 ± 0.04 | 3.83 ± 0.19 | 3.62 | 4.81 | 4.53 | Syn |
| Mix 7 | x | x | 9.1 | x | 3.7 | 7.4 | 79.8 | 1.40 ± 0.11 | 5.77 ± 0.15 | 4.11 | 8.88 | 6.34 | Syn |
| Mix 8 | 63.7 | 4.7 | x | x | x | 7.4 | 24.2 | 15.3 ± 0.33 | 29.3 ± 3.42 | 1.91 | 8.58 | 0.56 | Add |
| Mix 9 | 63.7 | 4.7 | x | x | 3.7 | x | 27.9 | 49.2 ± 3.71 | 66.8 ± 3.19 | 1.36 | 27.28 | 0.55 | Add |
| Mix 10 | 63.7 | 4.7 | x | 11.4 | x | x | 20.2 | 27.1 ± 0.51 | 25.6 ± 1.21 | 0.94 | 7.57 | 0.28 | Ant |
| Mix 11 | 63.7 | 4.7 | 9.1 | x | x | x | 22.5 | 24.3 ± 0.45 | 60.9 ± 2.87 | 2.51 | 22.4 | 0.92 | Add |
| Samples | Composition |
|---|---|
| Control | Polysorbate 80 at 2% (w/w) in water |
| NANO A | 2% (w/w) polysorbate 80 + 6% (w/w) A. nemorosa EO from aerial parts |
| NANO B | 2% (w/w) polysorbate 80 + 4% (w/w) A. nemorosa EO from aerial parts + 2% (w/w) ethyl oleate |
| EC50 | Selectivity Index (SI) | ||
|---|---|---|---|
| T. brucei (s427) | Balb3T3 | ||
| Nanoemulsions | μg/mL | μg/mL | |
| Control | >40,000 | >40,000 | |
| NANO A | 167.4 ± 12.8 (normalized 9.89 ± 0.797) | 1399 ± 464 | 8.35 |
| NANO B | 322.3 ± 43.5 (normalized 12.7 ± 1.72) | 1745 ± 347 | 5.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldassarri, C.; Falappa, G.; Mazzara, E.; Acquaticci, L.; Ossoli, E.; Perinelli, D.R.; Bonacucina, G.; Dall’Acqua, S.; Cappellacci, L.; Maggi, F.; et al. Antitrypanosomal Activity of Anthriscus Nemorosa Essential Oils and Combinations of Their Main Constituents. Antibiotics 2021, 10, 1413. https://doi.org/10.3390/antibiotics10111413
Baldassarri C, Falappa G, Mazzara E, Acquaticci L, Ossoli E, Perinelli DR, Bonacucina G, Dall’Acqua S, Cappellacci L, Maggi F, et al. Antitrypanosomal Activity of Anthriscus Nemorosa Essential Oils and Combinations of Their Main Constituents. Antibiotics. 2021; 10(11):1413. https://doi.org/10.3390/antibiotics10111413
Chicago/Turabian StyleBaldassarri, Cecilia, Giulia Falappa, Eugenia Mazzara, Laura Acquaticci, Elena Ossoli, Diego Romano Perinelli, Giulia Bonacucina, Stefano Dall’Acqua, Loredana Cappellacci, Filippo Maggi, and et al. 2021. "Antitrypanosomal Activity of Anthriscus Nemorosa Essential Oils and Combinations of Their Main Constituents" Antibiotics 10, no. 11: 1413. https://doi.org/10.3390/antibiotics10111413
APA StyleBaldassarri, C., Falappa, G., Mazzara, E., Acquaticci, L., Ossoli, E., Perinelli, D. R., Bonacucina, G., Dall’Acqua, S., Cappellacci, L., Maggi, F., Ranjbarian, F., Hofer, A., & Petrelli, R. (2021). Antitrypanosomal Activity of Anthriscus Nemorosa Essential Oils and Combinations of Their Main Constituents. Antibiotics, 10(11), 1413. https://doi.org/10.3390/antibiotics10111413