Abstract
The extent of similarity between E. faecium strains found in healthy feedlot beef cattle and those causing extraintestinal infections in humans is not yet fully understood. This study used whole-genome sequencing to analyse the antimicrobial resistance profile of E. faecium isolated from beef cattle (n = 59) at a single feedlot and compared them to previously reported Australian isolates obtained from pig (n = 60) and meat chicken caecal samples (n = 8), as well as human sepsis cases (n = 302). The E. faecium isolated from beef cattle and other food animal sources neither carried vanA/vanB responsible for vancomycin nor possessed gyrA/parC and liaR/liaS gene mutations associated with high-level fluoroquinolone and daptomycin resistance, respectively. A small proportion (7.6%) of human isolates clustered with beef cattle and pig isolates, including a few isolates belonging to the same sequence types ST22 (one beef cattle, one pig, and two human isolates), ST32 (eight beef cattle and one human isolate), and ST327 (two beef cattle and one human isolate), suggesting common origins. This provides further evidence that these clonal lineages may have broader host range but are unrelated to the typical hospital-adapted human strains belonging to clonal complex 17, significant proportions of which contain vanA/vanB and liaR/liaS. Additionally, none of the human isolates belonging to these STs contained resistance genes to WHO critically important antimicrobials. The results confirm that most E. faecium isolated from beef cattle in this study do not pose a significant risk for resistance to critically important antimicrobials and are not associated with current human septic infections.
1. Introduction
Enterococci are ubiquitous Gram-positive commensal bacteria that encompass more than 50 species found in various environments, such as the gastrointestinal tract of animals and humans, as well as the hospital environment [1]. Some Enterococcus spp. such as Enterococcus faecalis strain Symbioflor 1 are used as probiotics to treat diarrhoea, improve immunity, and provide other health benefits [2,3]. However, other Enterococcus spp. such as Enterococcus faecium are among the most important causes of hospital-acquired infections in humans, including endocarditis, sepsis, urinary tract, and central nervous system infection [1,4]. More than 90% of enterococcal infections identified in patients with bacteraemia are caused by either E. faecium or Enterococcus faecalis [5]. In additional to their ability to form biofilms, E. faecium and E. faecalis possess additional virulence factors that have contributed to their pathogenesis and clinical significance [6]. Some of these virulence factors include the production of enzymes and toxins which allow the bacteria to adhere to and colonize host tissues, modulate host immunity, and increase the severity of infection [7].
E. faecium are also intrinsically resistant to various antimicrobial classes, including aminoglycosides, cephalosporins, and trimethoprim-sulfamethoxazole [1,8,9]. E. faecium is known for its efficiency in recruiting and exchanging AMR determinants through mutation or horizontal transfer of ARGs located on mobile genetic elements (MGEs), which can be readily transferred between E. faecium lineages [10]. E. faecium is also capable of acquiring plasmid-mediated resistance determinants from other Enterococcus species [11]. Moreover, the transfer of vancomycin resistance from E. faecium to methicillin-resistant Staphylococcus aureus (MRSA) via broad host range plasmids is a major concern [12]. Plasmids found in enterococci are categorized based on their rep (replication) families, which have either narrow or broad host ranges. Rep-family plasmids such as rep2, rep3, rep5, rep8, rep9, rep11, rep12, rep14, rep15, rep16, rep18, and rep19 are narrow host range, whereas others, like rep1, rep4, rep6, rep7, rep10, and rep13 are considered broad host range [13]. The presence of plasmids in enterococci is extensive, as demonstrated by the detection of at least one and up to seven plasmids in 94.6% of E. faecium isolates [14]. Multidrug-resistant E. faecium is the most common cause of hospital-acquired infections compared to other species of enterococci [15]. In addition, many human E. faecium isolates have developed resistance to last-resort antimicrobials, such as daptomycin, linezolid, tigecycline, and vancomycin, which could have dire consequences for public health in the future [8,9,16,17,18]. Among clinical E. faecium isolates worldwide, the vanA and vanB vancomycin resistance genotypes are the most prevalent ARG variants [19].
Whilst some genetic similarity has been found among enterococci isolated from animals and those causing human infections [20], other studies have reported that animal-derived and human-derived enterococci are genetically distinct populations [21,22]. However, there is limited information on the host-species specificity of individual E. faecium sequence types (STs), together with their correlation with specific hospital-infection-associated AMR determinants and virulence factors. Australia represents a unique study site to explore the host specificity of animal-origin E. faecium isolates given its isolation, quarantine restrictions including a ban on importation of live food-producing animals, and conservative regulation of antimicrobial use in both human and animal health [23]. As a case in point, the use of antimicrobials in livestock production is highly regulated in Australia. For example, Australian legislation prohibits the use of fourth-generation cephalosporins, colistin, fluoroquinolones, and gentamicin in food-producing animals [24,25]. As a result, a low prevalence of AMR has been reported in enterococci isolated from different food-producing animals in Australia compared to other countries [26,27,28].
In our previous study, we investigated the frequency of AMR in enterococci from faecal samples collected at the entrance to and exit from a beef feedlot in South Australia [29]. While the prevalence of AMR to individual antimicrobials remained largely static between entry and exit, there was a dramatic shift in Enterococcus spp. prevalence, with E. faecium infrequently isolated from entry samples (9/104; 8.7%), but then becoming the most prevalent species in exit samples (117/144; 81.3%). The interplay between diet, management practices, and microbial ecology is a potential factor that could contribute to the observed shift in Enterococcus spp. [30,31]. This shift in Enterococcus prevalence is noteworthy as it highlights the importance of monitoring alterations in microbial communities, where these changes can have implications for both animal health and food safety. Further genomic interrogation is now required to determine the genetic similarity of these cattle-origin E. faecium isolates compared to those detected in other food animals and humans in Australia as well as to examine capacity for cross-host species transmission and potential spread via the food chain or the environment. Therefore, this study aimed to conduct further genetic analysis on the beef cattle E. faecium isolates through comparing them to isolates obtained from healthy meat chickens and pigs at slaughter as well as human clinical blood sepsis isolates in Australia described in previous studies [5,29].
2. Results
2.1. Distribution of MLST Genotypes and Virulence Factors
Overall, 59 strains of E. faecium from beef cattle were analysed via WGS and compared with commensal isolates from healthy slaughter age pigs (n = 60), meat chickens (n = 8), and a large collection of human clinical sepsis isolates (n = 302). SNP analysis of the beef cattle, chicken, and pig isolates confirmed that they formed two main clades (Clade 1 and Clade 2). Clade 2 was subsequently divided into two subclades (Clade 2a and Clade 2b). Clade 2b contained a notable monophyletic subclade (Clade 2ba) that was composed entirely of more closely related human isolates (Figure 1). The predominant animal isolate clades also contained a small proportion of human isolates (23/302; 7.3%) including five isolates each located within the beef-cattle-isolate-dominated Clade 1 and the pig-isolate-dominated Clade 2a (Table S1). Among beef cattle isolates, 15 distinct sequence types (STs) were identified via MLST, the most common being ST798 in Clade 1 (9/59; 15.2%), followed by ST32 in Clade 2b and ST1036 in Clade 1 (8/59; 13.5% each). By comparison, the most common STs among pig and human isolates were ST5 in Clade 2a (6/60; 10.0%) and ST796 in Clade 2ba (47/302; 15.6%), respectively. Three E. faecium STs contained both animal and human isolates. These included ST32, which comprised eight beef cattle isolates and one human isolate; ST22, which comprised one beef cattle isolate, one pig isolate, and two human isolates; and ST327, which comprised two beef cattle isolates and one human isolate.
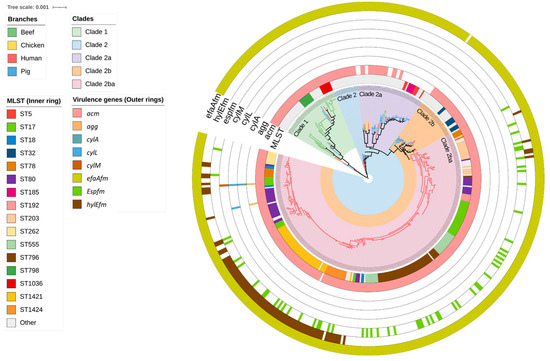
Figure 1.
A mid-point rooted, maximum-likelihood phylogenetic tree constructed based on analysis of single-nucleotide polymorphisms (SNPs) of the core SNPs of 429 Enterococcus faecium genomes isolated from beef cattle (n = 59), meat chicken (n = 8), pig (n = 60), and human (n = 302) sources. The branch colour indicates the source of the isolates. Clades are coloured according to the legend. The sequence type of the isolates (inner ring) and type of virulence gene (outer rings) are annotated according to the legend.
Out of the eight E. faecium virulence genes identified in this study, acm (collagen-binding) and efaAfm (adherence) were most prevalent across all sources of isolates. Among the beef cattle isolates, acm was found in 56/59 (95.0%) and efaAfm in 58/59 (98.3%) (Figure 1). The remaining E. faecium virulence genes were exclusively found in human-source isolates in Clade 2ba. These included agg (adherence, 2/302; 0.7%), cylA (cytolysin, 1/302; 0.3%), cylL (cytolysin, 2/302; 0.7%), cylM (cytolysin, 1/302; 0.3%), espfm (surface protein, 55/302; 18.2%), and hylEfm (hyaluronidase 82/302; 27.1%). The human E. faecium isolates located in STs 32, 22, and 372 also did not contain espfm and hylEfm markers typically exclusive to hospital-associated strains, but carried both acm and efaAfm.
2.2. Antimicrobial Resistance Genes Including Those Encoding Resistance to Critically Important Antimicrobials
Heat maps were generated based on the identification of genes from the ResFinder and CARD databases (Figure 2). Overall, 38 ARGs responsible for encoding resistance to aminoglycosides (aadA5, aadD, aph(2”)-Ia, aph(2”)-Ie, aph(3’)-III, aac(6’)-aph(2”), aac(6’)-Ii, ant(6)-Ia, ant(9)-Ia), ß-lactams (blaZ, pbp5), fluoroquinolones (efmA, gyrA, parC), folate pathway inhibitor (dfrG), glycopeptides (vanA, vanB), lipopeptides (cls, liaR, liaS), macrolides/lincosamides, and/or streptogramins (MLS) (eatAv, erm(A), erm(B), erm(T), lnu(B), lnu(G), lsa(E), msr(C), vatE), phenicols (cat, fexB, poxtA), and tetracyclines (tet(45), tet(L), tet(M), tet(O), tet(S), tet(W)) were identified in the entire collection of 429 isolates (Figure S1). Among these, eatAv, efmA, msrC, tetL, and tet(45) are associated with multidrug efflux pumps and other transporters, which may potentially lead to resistance against multiple antimicrobials. The number of ARGs identified in each isolate ranged from one to ten. Human E. faecium isolates possessed the highest mean number of ARGs per isolate (n = 8) compared to pig and chicken (n = 5, each) and beef cattle isolates (n = 3).
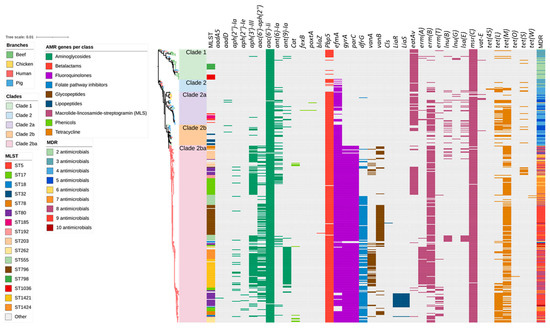
Figure 2.
Antimicrobial resistance gene (ARG) profiles clustered on the basis of Figure 1’s SNP-based phylogenetic tree composed of 429 Enterococcus faecium genomes isolated from beef cattle (n = 59), chicken (n = 8), pig (n = 60), and human (n = 302) sources. Branches are coloured based on source followed by clade according to the legend. The remaining columns indicate (1) isolate sequence type, (2) ARGs detected in the isolate, and (3) multidrug resistance profile of the isolate. The detected ARGs are clustered according to their respective antimicrobial classes as follows: aminoglycoside (9 ARGs shown in dark green), amphenicols (3 ARGs shown in light green), β-lactams (2 ARGs shown in red), fluoroquinolones (3 ARGs shown in purple), folate synthesis inhibitors (1 ARG shown in light blue), glycopeptides (2 ARGs shown in dark brown), lipopeptides (3 ARGs shown in dark blue), macrolide/lincosamide and/or streptogramins (8 ARGs shown in magenta), and tetracyclines (6 ARGs shown in light brown).
A total of 13 ARGs were identified in the beef cattle E. faecium isolate collection, including those imparting resistance to aminoglycosides (aac(6′)Ii, and ant(6)-Ia), ß-lactams (pbp5), fluoroquinolones (efmA), MLS (eatAv, lnu(G), vat(E), msr(C), and erm(B)), and tetracyclines (tet(45), tet(L), tet(M), and tet(S)) (Figure 2). The most common ARGs observed in beef cattle isolates were plasmid-mediated aac (6’)-Ii (extremely high 59/59; 100.0%) and the chromosomally encoded msr(C) (extremely high 58/59; 98.3%), eatAv (extremely high 45/59; 76.3%), pbp5 (very high 30/59; 50.8%), and efmA (high 21/59; 35.6%), which are responsible for resistance to aminoglycosides, MLS, β-lactams, and fluoroquinolones, respectively.
Similar to the results obtained for the pig- and chicken-origin E. faecium isolates, none of the E. faecium isolates from beef cattle were found to possess gyrA or parC mutations associated with high-level fluoroquinolone resistance. However, some did possess the ABC transporter gene efmA, which potentially confers low-level resistance as a fluoroquinolone efflux pump (Figure 2 and Figure 3, Table 1). Similarly, none of the cattle isolates possessed vanA/vanB genes encoding resistance to vancomycin (Figure 4) or liaR/liaS genes mutations to daptomycin (Figure 5). By comparison, significant proportions of the human sepsis isolates carried a chromosomal quinolone resistance determining region (QRDR) and daptomycin resistance mutations together with vancomycin ARGs (Table 1).
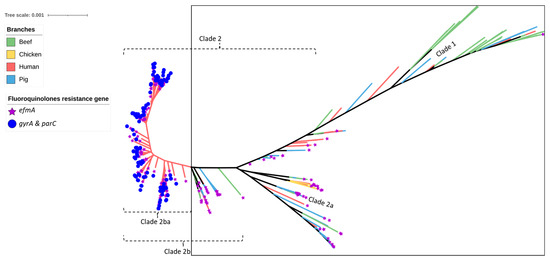
Figure 3.
SNP-based phylogeny of 429 Enterococcus faecium isolates isolated from beef cattle (green branches, n = 59), meat chickens (orange branches, n = 8), pigs (blue branches, n = 60), and humans (red branches, n = 302). Purple stars indicate the isolates shown to contain the efmA efflux pump gene (imparting resistance to fluoroquinolones), whereas the blue circles indicate the isolates shown to contain gyrA and parC point mutations (imparting high-level resistance to fluoroquinolones). All beef, meat chicken, and pig isolates (encompassed within the square box) contained the efflux pump gene only.

Table 1.
Prevalence of antimicrobial resistance genes identified in E. faecium isolated from beef cattle, meat chickens, pigs, and humans in Australia.
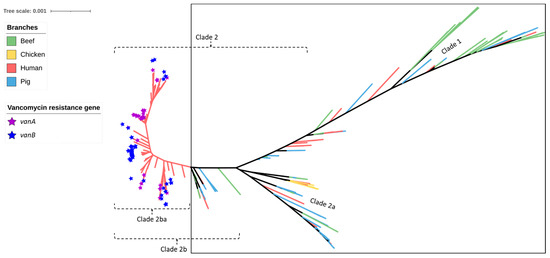
Figure 4.
SNP-based phylogeny of 429 Enterococcus faecium isolates obtained from beef cattle (green branches, n = 59), meat chickens (orange branches, n = 8), pigs (blue branches, n = 60), and humans (red branches, n = 302). Purple stars indicate the isolates shown to contain vanA and blue stars indicate the isolates shown to contain vanB (both genes imparting resistance to vancomycin). None of the beef cattle, meat chicken, and pig isolates (encompassed within the square box) possessed vanA or vanB vancomycin resistance genes.
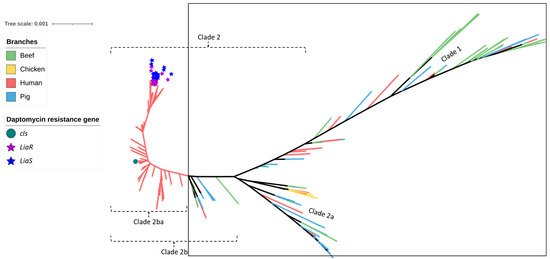
Figure 5.
SNP-based phylogeny of 429 Enterococcus faecium isolates obtained from beef cattle (green branches, n = 59), meat chickens (orange branches, n = 8), pigs (blue branches, n = 60), and humans (red branches, n = 302). The green circle indicates the isolate shown to contain cls, the purple stars indicate the isolates shown to contain liaR, and the blue stars indicate the isolates shown to contain liaS mutations imparting resistance to daptomycin. None of the beef cattle, meat chicken, and pig isolates (encompassed within the square box) possessed these gene mutations.
2.3. Plasmid Replicons
Overall, 21 plasmid replicon families were observed in the isolate collection. The number of plasmids observed in each isolate ranged from zero to eight, with the highest number of plasmids found in three human isolates (1.0%). Overall, 43 (72.9%) of beef cattle isolates contained at least one plasmid, and 2 isolates harboured six plasmids. A significant number of beef cattle isolates (16/59; 27.1%) did not possess any plasmids compared to human (7/302, 2.3%) and pig isolates (1/60, 1.7%). Plasmid replicon rep1 (broad host range) was predominantly found in beef cattle (33/59; 55.9%) and pig isolates (30/60; 50.0%), but of comparatively lower prevalence in human isolates (15/302; 5.0%), while rep2 (narrow host range) was more commonly found in human (238/302 78.8%) and pig isolates (24/60; 40%) but was present in a much smaller proportion (5/59; 8.5%) of beef cattle isolates. Highly diverse sets of plasmids were identified among the E. faecium isolates from human sepsis cases (Figure 6 and Figure S2).
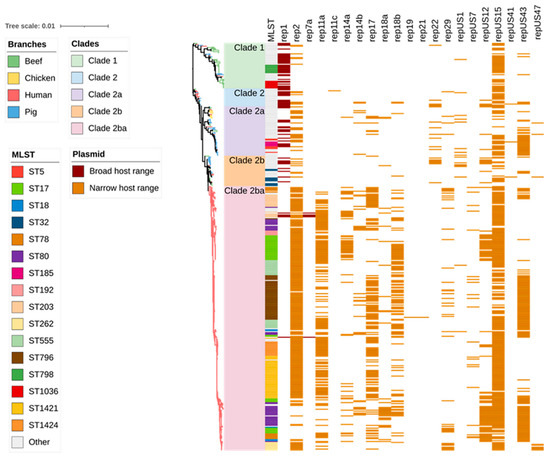
Figure 6.
Plasmid replicon profiles clustered on the basis of Figure 1’s SNP-based phylogenetic tree composed of 429 Enterococcus faecium genomes isolated from beef cattle (n = 59), meat chickens (n = 8), pigs (n = 60), and humans (n = 302). Branches are coloured based on source followed by clade according to the legend. The remaining columns indicate isolate sequence type and plasmid replicon detected in the isolate. The detected plasmids are clustered according to their respective types as follows: broad host range (2 plasmid replicons shown in dark brown) and narrow host range (19 plasmid replicons shown in light brown).
2.4. Agreement between Plasmid Replicons and ARG Content
The presence of vanA (found only in isolates from humans) was mostly linked to three plasmid replicons: rep2 (98.5%; 64/65), repUS15 (95.4%; 62/65), and rep11a (81.5%; 53/65). Most E. faecium isolates carrying the streptogramin resistance gene vatE (found only in isolates from animals) were associated with the plasmid replicons rep2 (80%; 4/5) and rep17 (60%; 3/5) (Figure 7). When plasmid replicons and E. faecium ARGs known to be plasmid-mediated were cross tabulated and compared according to isolate source (beef cattle vs. human), clear differences in matches were apparent between the two host sources, apart from repUS15, which was highly prevalent in both groups of isolates and often associated with aac(6’)-Ii, erm(B), and tet(M) ARGs (Figure 8).
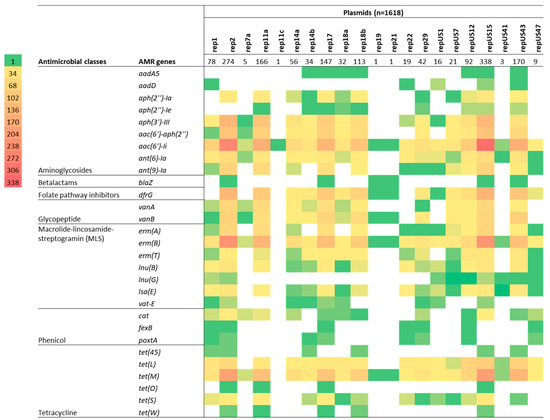
Figure 7.
Cross-tabulation heat map showing the degree of correlation between plasmid replicon types and the presence of an antimicrobial resistance gene (ARG) for 429 isolates of Enterococcus faecium isolated from beef cattle (n = 59), meat chickens (n = 8), pigs (n = 60), and humans (n = 302). Plasmid replicons are listed horizontally (total number identified in the 429 isolates is also indicated), whereas the ARGs are listed vertically in their classes. The colour strips indicate the number of isolates (in multiples of 34) exhibiting each particular plasmid/ARG match.
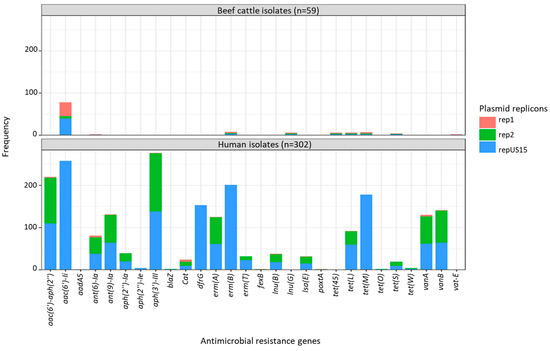
Figure 8.
Cross-tabulation of plasmid-mediated antimicrobial resistance genes and plasmid replicons in Enterococcus faecium isolated from beef cattle (n = 59) and humans (n = 302). rep1—broad host range shown in pink; rep2 and repUS15—narrow host range shown in green and blue. The ST22 and ST32 isolates, which were detected in beef cattle, pigs, and humans, were clustered together in the same clade. The analysis of ARGs showed that the ST22 and ST32 isolates had similar chromosomal genes for antimicrobial resistance, such as efmA, msr(C), and pbp5, in addition to aac(6’)-I, which is more likely to be plasmid-mediated. The pig isolate possessed additional ARGs linked to macrolide and tetracycline resistance. The ST327 isolate obtained from humans and beef cattle were clustered within Clade 1 and carried the chromosomal eatAv gene, conferring resistance to lincosamides, streptogramin As, and pleuromutilins in addition to msr(C). Regardless of the presence of the plasmid replicon repUS15, the ARG content in the beef isolates of ST22 and ST32 remained unchanged (Table 2).
3. Discussion
Enterococci are commonly found as commensals in the gastrointestinal tract of animals and humans, but some species, such as E. faecium, can cause serious nosocomial infections, including endocarditis, sepsis, and urinary tract infections in humans [20,32]. Regular monitoring and testing of AMR in E. faecium isolated from both animal production and human hospital settings can provide important information about the evolution and spread of resistance and the relative impact of food and/or environmental transmission compared to direct human-to-human contact. Through detailed whole bacterial genome bioinformatic analyses, this study focused on AMR, plasmid, and virulence factor profiles identified in E. faecium isolated from the gastrointestinal tracts of healthy slaughter-age beef cattle from a single feedlot in Australia and compared these with previous data established for similar pig [33] and meat chicken isolates [28] together with a large collection of clinical isolates from cases of sepsis in humans in Australia [5].
The study had three major findings. First, phylogenetic analysis confirmed that the E. faecium isolates from healthy slaughter-age beef cattle, pig, and meat chicken sources (found in Clades 1–2b) were genetically distinct from most human clinical isolates, which formed a monophyletic clade (Clade 2ba). Second, none of the animal-origin isolates from beef cattle, pig, and meat chicken sources possessed ARGs or mutations responsible for high-MIC-level resistance to three critically important antimicrobials (ciprofloxacin, vancomycin, and daptomycin) typically present in human isolates. Third, a small proportion (23/302; 7.6%) of human clinical isolates were found clustered with some of the animal isolates (including some belonging to the same ST) within Clades 1–2b, but none of these human isolates possessed ARGs to critically important antimicrobials. These results confirm that, as previously determined for Australian pig and poultry E. faecium isolates [33], beef cattle from this particular feedlot also represent an extremely low risk of introducing resistant E. faecium (or their ARGs) to humans via the food chain or through the environment with corresponding minimal impact to human health. Evidence for this conclusion can be found in the limited crossover of animal vs. human STs (the majority of beef cattle isolates were located in Clade 1) and the differences in virulence genes and ARG arrays observed, particularly to the critically important antimicrobials (fluoroquinolones, daptomycin, vancomycin). Furthermore, far more plasmid replicons were observed in the E. faecium isolated from humans compared to beef cattle isolates.
The MLST analysis of E. faecium isolates from beef cattle in this study revealed significant diversity in their STs with most located in Clade 1. Only three primarily beef-cattle-origin STs (ST22, ST32, and ST327) contained either one or two E. faecium isolates obtained from human sepsis cases. This is in agreement with a previous report that clinical E. faecium isolates causing infections in humans tend to belong to different sub-lineages than those found in animals, food, and the environment [20]. For example, E. faecium ST17, a clonal lineage responsible for hospital-acquired infections throughout the globe, was only identified among human isolates in the present study (where it represented 12.6% of the Australian human E. faecium isolate collection). Compared to other strains in the collection, ST17 E. faecium were more likely to exhibit high-level resistance to aminoglycosides, beta-lactams, fluoroquinolones, and tetracycline, which correlated with the possession of aac (6’)-Ii, tet (L), and tet (M) genes, as well as mutations in gyrA, parC, and pbp5, as documented in a previous international study [34]. Additionally, the vancomycin resistance gene vanB was detected in the hospital-derived ST17 human isolates (3/38; 7.9%). This study found that the human isolates carrying the van operon were not clustered in the same evolutionary clade as the beef, chicken, and pig isolates. However, the ST22 and ST32 isolates shared the same ARG types, including plasmid-mediated genes responsible for resistance to aminoglycosides (aac(6’)-I); the chromosomally encoded ampicillin resistance mutation (pbp5); low-level fluoroquinolone resistance (efmA); and macrolide, lincosamide, and streptogramin ARGs (msr(C) and eatAv in ST327 isolates). It has been previously reported that ST32 and ST22 share the same ARG profiles, yet they are not typical clones associated with vancomycin resistance nor do they have point mutations in gyrA and parC responsible for high-level fluoroquinolone resistance, indicating a limited human health risk [35,36,37]. Both ST22 and ST32 are categorized as commensal isolates and are part of CC328, which is not associated to any of the established clonal complexes of colonizing or hospital-adapted strains [37].
Enterococci are rapidly evolving to become resistant to various classes of antimicrobials, including fluoroquinolones, glycopeptides, lipopeptides, oxazolidinones, and streptogramins [38,39,40]. Within Australia, the streptogramin pristinamycin is also rated of critical importance and grouped with the aforementioned antimicrobial classes in the ASTAG importance ratings [41]. The use of virginiamycin as a feed additive in food-producing animals is believed to contribute to the development of quinupristin/dalfopristin resistance, which are sometimes used as a last resort treatment for vancomycin-resistant E. faecium infections in humans [42]. The current study showed that resistance to streptogramins in beef cattle E. faecium isolates was comparatively lower compared to human and pig isolates. Some beef E. faecium isolates carried the ermB gene (3/59; 5.1%), which encodes an enzyme responsible for methylating the 23S rRNA component of the bacterial ribosome, whilst nearly all contained the msr(C) gene (58/59; 98.3%), which encodes an efflux pump, resulting in resistance to macrolides and streptogramin B (i.e., quinupristin), and most contained the eatAV gene which encodes an ABC transporter and encodes resistance to lincosamide, pleuromutlins, and streptogramin A [43]. Furthermore, one isolate (1.7%) from beef and pig samples and four isolates from meat chickens (50%) were found to harbor the vatE gene responsible for streptogramin A (i.e., dalfopristin) resistance. Possession of either ermB or eatAV with msr(C) genes is associated with resistance to quinupristin–dalfopristin, a combination of streptogramin A and B [44]. Whilst a recent survey found that 19.5% of Australian feedlots reported using virginiamycin [45], the present study found that chicken and pig isolates had higher levels of streptogramin resistance genes compared to beef cattle isolates. High prevalence of msr(C) in the beef cattle isolates may also be related to the common use of macrolides as first-line treatments for bovine respiratory disease in Australian feedlots [46]. As virginiamycin has not been used in the Australian pig and poultry industries for many years, the higher prevalence of resistance could be related to the co-selection of enterococci resistant to quinupristin–dalfopristin as a result of the historic use of virginiamycin in poultry and current use of tylosin in pigs [33,47].
In recent decades, the vancomycin resistance gene vanA has been reported in E. faecium isolated from animals and animal-derived food products [48]. VanA-positive enterococci were isolated from food animals in England in 1993 [49]. Since then, it has been found among food animals worldwide, causing significant public health concerns [50,51,52]. The detection of vancomycin-resistant E. faecium has been shown to be associated with the use of avoparcin for growth promotion in food animals [53,54]. However, after vancomycin-resistant enterococci emerged, avoparcin was withdrawn from the market in various countries [55]. In this study, the vancomycin resistance genes vanA and vanB were found in 21.5% and 28.5% of human isolates, respectively. However, no vancomycin ARGs were found in any of the beef, chicken, and pig isolates. In another study in Australia using phenotypic methods, enterococci isolated from cattle were sensitive to a number of medically important antimicrobials, such as daptomycin, linezolid, tigecycline, and vancomycin [27]. The findings of this study provide evidence that E. faecium isolated from Australian healthy slaughter-age beef cattle, pigs, and meat chickens are mutually exclusive from the source of the circulating vancomycin-resistant strains of clinical significance in the public health sector [5].
Daptomycin is another antimicrobial used as a last-resort antimicrobial to treat vancomycin-resistant enterococci [40]. However, there have been increasing reports of vancomycin-resistant enterococci (VRE) developing resistance to daptomycin [56]. In this study, none of the E. faecium isolated from food-producing animals harboured known resistance genes for daptomycin. However, 23 (7.6%) of the human isolates showed mutations in the genes responsible for daptomycin resistance (liaR and liaS). Contrary to this, some studies have shown that daptomycin resistance does not always result from liaFSR mutations. Mutations in chromosomal genes cls and gdpD, which encode cardiolipin synthase and glycerophosphoryl diester phosphodiesterase, respectively, have been linked to daptomycin resistance [40]. In this study, it was found that only one human isolate had a mutation in the cls gene. In our previously published study, we detected daptomycin resistance in E. faecium isolates obtained from both entry (1/9; 11.1%) and exit (21/117; 17.9%) beef cattle faecal samples using a phenotypic method, but the underlying mechanism behind this resistance could not be determined [29]. These findings suggest that there may be other, as yet unknown, pathways responsible for daptomycin resistance in animal-origin enterococci [57,58], including within Australia [28].
The prevalence of fluoroquinolone resistance among E. faecium isolates is increasing globally, posing a significant threat to public health. In recent years, efflux pumps have played a crucial role in the MDR status of various bacteria [59]. In this study, the efmA gene efflux pump, which encodes resistance to fluoroquinolones and macrolides, was observed in E. faecium isolated from beef (35.6%), chicken (100%), human (92.4%), and pig (56.7%) samples. Efflux pumps are known to contribute to intrinsic and acquired resistance to antimicrobials used to treat infectious diseases [60]. However, fluoroquinolone resistance characterized by high MIC values is commonly associated with mutations in topoisomerase IV (parC) and DNA gyrase (gyrA), as previously reported [61]. In this study, no fluoroquinolone-resistance-causing mutations were identified in E. faecium isolates obtained from beef cattle.
The potential for AMR to be transferred to other bacteria exists through several means, with plasmids serving as one of the primary mechanisms for this transfer to occur [62]. Plasmids that contain AMR determinants play an important role in horizontal gene transfer as they often harbour other mobile genetic elements such a transposons, integrons and insertional sequences within their structures [63]. In this study, a previously published plasmid classification scheme was utilized to investigate the possible contribution of specific plasmid families to AMR in Enterococcus spp. [13]. Among the human sepsis isolates, vanA and vatE genes, responsible for resistance to vancomycin and quinupristin-dalfopristin, respectively, were mostly associated with narrow host range rep-2 plasmid replicons. By comparison, the most commonly observed plasmids in E. faecium isolates from beef cattle were repUS15 (67.8%) and broad host range rep1 (55.9%), with only a relatively minor proportion carrying rep2 (8.5%). Other international studies have also shown the strong association between carriage of rep2 plasmid replicons and resistance to glycopeptides in enterococci isolated from both humans and food-producing animals [64,65]. In summary, plasmid replicon identification and their associations with critically important ARGs has also shown that Australian beef cattle E. faecium isolates (together with pig and meat chicken isolates) pose limited risk to human health from horizontal gene transfer.
Enterococci can carry various virulence genes, including those involved in biofilm formation, adhesion, and invasion of host cells [66]. This study identified a number of virulence genes in the genomes of sequenced E. faecium isolates, with the acm and efaAfm genes being the most commonly detected among all isolates regardless of source and espfm only detected in human isolates. Both acm and efaAfm were prevalent in over 95% of E. faecium isolates from Australian beef cattle, similar to the results obtained for chicken and pig isolates. In human E. faecium isolates, the virulence genes espfm (18.2%) and hylEfm (27.1%) were also commonly observed in addition to acm and efaAfm. The hylEfm gene, which is responsible for intestinal colonization, has been detected in ampicillin-resistant E. faecium ST17 and vancomycin-resistant E. faecium (VREF) strains in hospitals worldwide [67,68,69]. This study shows that the type, resistance, and virulence characteristics of E. faecium found in food-producing animals differ from those found in human isolates.
This study has some limitations that should be taken into account. One major limitation is that the E. faecium isolates from meat chickens, pigs, and humans were obtained from a universal database as secondary data. Second, the beef cattle isolates were obtained as part of longitudinal study comparing entry and exit samples from a single feedlot, which may not be sufficient to generalize the results to the wider population. Ideally, this study could now be replicated in a wider range of feedlots. Third, the E. faecium isolates from food animals obtained during surveillance were compared with pathogenic isolates from human sepsis cases, and future studies could also examine human carriage isolates. Finally, due to the use of short-read sequencing data, it was not possible to determine the AMR profile and virulence factor composition of each plasmid replicon. In the future, the scope of this study could be expanded to include a range of farms and geographical locations.
4. Materials and Methods
4.1. Genomic Analysis
The E. faecium isolates from beef cattle were whole-genome sequenced and assembled using a previously described method (BioProject PRJNA879912) [29]. The obtained data were subsequently compared to three previous studies that reported WGS of gastrointestinal E. faecium isolates from healthy pigs (accession number PRJNA639902) [33] and meat chickens (accession number PRJNA524396) [28] from slaughterhouses as well as isolates from human sepsis cases (accession number PRJNA562414) [5] from Australia. Assembled sequences with less than 30× coverage and less than 25,000 SNPs were excluded from further analysis. The isolates that met the criteria were from beef cattle (n = 59), pigs (n = 60), meat chickens (n = 8), and human sepsis cases (n = 302). The genetic relationships between isolates were examined using single-nucleotide polymorphisms (SNPs) from cleaned WGS reads that were mapped to an E. faecium complete genome (NCBI Assembly Accession: CP003583.1). The software Snippy v4.6.0 (https://github.com/tseemann/snippy, (accessed on 21 September 2022) was used to call core SNPs, i.e., SNPs that can be determined in all isolates. A maximum likelihood (ML) tree was constructed with RAxML v8.2.10 using the model GTRCAT, and a rapid bootstrap analysis with 100 bootstraps for the best-scoring ML tree was undertaken [70]. This was followed by recombination removal using ClonalFrameML v1.12 [71]. Multilocus sequence typing (MLST) was undertaken using MLST 2.0 [72]. The final phylogenetic tree and heat map were manipulated with iTOL (https://itol.embl.de/, accessed on 23 October 2022) for display [73]. A heat map illustrating the presence or absence of each trait for each isolate was created to assess all data elements for all isolates.
4.2. Comparative Analysis of Virulence Genes, AMR Genes and Plasmids
The detection of virulence genes was carried out using VirulenceFinder 2.0 [74,75]. To identify antimicrobial resistance genes (ARGs), we used ResFinder 4.0 [76] and, to further pinpoint the chromosomal point mutations associated with AMR, we used PointFinder [77]. The ResFinder web server (www.genomicepidemiology.org, accessed on 12 October 2022) was used to identify acquired ARGs in the WGS data, using a threshold of 98.0% identity (ID). ARGs were also predicted using the Antibiotic Resistance Genes Database (ARDB) and the Comprehensive Antibiotic Resistance Database (CARD) in addition to ResFinder [78]. To detect plasmid replicons, PlasmidFinder was used, with minimum identity and coverage of 95% and 60% parameters, respectively [79].
4.3. Statistical Analysis
Categorical measured traits including ARGs, the presence of plasmid replicons, and virulence factors were converted into a numerical code, with 1 indicating presence and 0 indicating absence. Proportions of AMR, MDR, ARGs, plasmid replicons or virulence factors were calculated using R Statistical Package version 4.0.0. Resistance profiles were categorised as MDR if the isolate exhibited resistance to one or more antimicrobials in three or more antimicrobial classes [80]. AMR and ARG frequencies were described as rare: <0.1%; very low: 0.1 to 1.0%; low: >1 to 10.0%; moderate: >10.0 to 20.0%; high: >20.0 to 50.0%; very high: >50.0 to 70.0%; and extremely high: >70.0% [81].
5. Conclusions
The continued use of antimicrobial agents in health care and agriculture settings inevitably leads to the emergence of AMR. This study compared the prevalence of virulence genes, ARGs, and plasmid replicon types identified among E. faecium isolates obtained from beef cattle at a single feedlot with those obtained from different animal hosts (pigs and meat chickens) and humans within Australia. Beef cattle, meat chicken, and pig-source E. faecium isolates were found to be of limited risk and largely unrelated to human-source isolates. The strict regulation of antimicrobials used in food animals is likely the reason for the low level of AMR found in food animals in Australia. The higher levels of AMR observed in human isolates may be because they are clinical isolates exposed to more antimicrobials than the surveillance isolates from food animals included in this study. This research approach helps understand the general trend and spread of AMR E. faecium in different sectors.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/antibiotics12071122/s1, Figure S1. The frequency of antimicrobial resistance gene in E. faecium isolated from beef (n = 59), pig (n = 60), chicken (n = 8) and human (n = 302); Figure S2. The frequency of plasmid replicons in E. faecium isolated from beef (n = 59), pig (n = 60), chicken (n = 8) and human (n = 302); Table S1. Prevalence of antimicrobial resistance genes, plasmid replicons, and virulence genes in E. faecium (n = 23) human Isolates mixed in animal isolate-dominated clades.
Author Contributions
Conceptualization: D.J.T. and K.R.P.; data curation, Y.E.M., M.F.H., K.R.P., W.Y.L. and T.V.; formal analysis, Y.E.M., W.Y.L. and K.R.P.; Funding acquisition, K.R.P., D.J.T. and J.P.M.; investigation, Y.E.M., M.F.H. and T.V.; project administration, K.R.P.; methodology, Y.E.M., M.F.H., T.V., W.Y.L., D.J.T. and K.R.P.; resources, T.V., W.Y.L., S.P.K., D.J.T. and K.R.P.; software, Y.E.M., T.V., K.R.P. and W.Y.L.; supervision T.V., D.J.T., J.P.M., S.P.K., W.Y.L. and K.R.P.; validation Y.E.M., M.F.H., T.V. and K.R.P.; visualization, Y.E.M., D.J.T., K.R.P. and W.Y.L.; writing—original draft, Y.E.M.; writing—review editing, Y.E.M., J.P.M., S.P.K., W.Y.L., K.R.P. and D.J.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Meat and Livestock Australia (MLA), grant number B.FLT.3003.
Institutional Review Board Statement
The sample collection protocols and procedures were approved by the University of Adelaide Animal Ethics Committee (AEC number S2019-072).
Informed Consent Statement
Not applicable.
Data Availability Statement
WGS reads are available in the SRA under BioProject beef (PRJNA879912), pig (PRJNA639902), meat chicken (PRJNA524396) and human (PRJNA562414) isolates.
Acknowledgments
We thank Meat and Livestock Australia for the funding of this project. We would like to thank the farm and abattoir staff for their participation in the study. We are also grateful for the contribution made by ACARE and the Davies Livestock Research Centre laboratory members for assisting during the bench work. We also thank Mandi Carr and Manouchehr Khazandi for their initial contribution to the study design.
Conflicts of Interest
The authors declare that there are no conflict of interest.
References
- García-Solache, M.; Rice, L.B. The Enterococcus: A Model of Adaptability to Its Environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [PubMed]
- Franz, C.M.; Huch, M.; Abriouel, H.; Holzapfel, W.; Gálvez, A. Enterococci as probiotics and their implications in food safety. Int. J. Food Microbiol. 2011, 151, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Fritzenwanker, M.; Kuenne, C.; Billion, A.; Hain, T.; Zimmermann, K.; Goesmann, A.; Chakraborty, T.; Domann, E. Complete Genome Sequence of the Probiotic Enterococcus faecalis Symbioflor 1 Clone DSM 16431. Genome Announc. 2013, 1, e00165-12. [Google Scholar] [CrossRef]
- O’Driscoll, T.; Crank, C.W. Vancomycin-resistant enterococcal infections: Epidemiology, clinical manifestations, and optimal management. Infect. Drug Resist. 2015, 8, 217. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Pang, S.; Stegger, M.; Sahibzada, S.; Abraham, S.; Daley, D.; Coombs, G. A three-year whole genome sequencing perspective of Enterococcus faecium sepsis in Australia. PLoS ONE 2020, 15, e0228781. [Google Scholar] [CrossRef]
- Sava, I.G.; Heikens, E.; Huebner, J. Pathogenesis and immunity in enterococcal infections. Clin. Microbiol. Infect. 2010, 16, 533–540. [Google Scholar] [CrossRef]
- Jett, B.D.; Huycke, M.M.; Gilmore, M.S. Virulence of enterococci. Clin. Microbiol. Rev. 1994, 7, 462–478. [Google Scholar] [CrossRef]
- Kristich, C.J.; Rice, L.B.; Arias, C.A. Enterococcal Infection—Treatment and Antibiotic Resistance. Available online: https://pubmed.ncbi.nlm.nih.gov/24649502/2014 (accessed on 12 March 2023).
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–569. [Google Scholar] [CrossRef]
- Sanderson, H.; Gray, K.L.; Manuele, A.; Maguire, F.; Khan, A.; Liu, C.; Navanekere Rudrappa, C.; Nash, J.H.E.; Robertson, J.; Bessonov, K.; et al. Exploring the mobilome and resistome of Enterococcus faecium in a One Health context across two continents. Microb. Genom. 2022, 8, 000880. [Google Scholar] [CrossRef]
- Weaver, K.E.; Kwong, S.M.; Firth, N.; Francia, M.V. The RepA_N replicons of Gram-positive bacteria: A family of broadly distributed but narrow host range plasmids. Plasmid 2009, 61, 94–109. [Google Scholar] [CrossRef]
- Palmer, K.L.; Kos, V.N.; Gilmore, M.S. Horizontal gene transfer and the genomics of enterococcal antibiotic resistance. Curr. Opin. Microbiol. 2010, 13, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.B.; Garcia-Migura, L.; Valenzuela, A.J.S.; Løhr, M.; Hasman, H.; Aarestrup, F. A classification system for plasmids from enterococci and other Gram-positive bacteria. J. Microbiol. Methods 2010, 80, 25–43. [Google Scholar] [CrossRef] [PubMed]
- Rosvoll, T.C.; Pedersen, T.; Sletvold, H.; Johnsen, P.J.; Sollid, J.E.; Simonsen, G.S.; Jensen, L.B.; Nielsen, K.M.; Sundsfjord, A. PCR-based plasmid typing in Enterococcus faecium strains reveals widely distributed pRE25-, pRUM-, pIP501-and pHTβ-related replicons associated with glycopeptide resistance and stabilizing toxin–antitoxin systems. FEMS Immunol. Med. Microbiol. 2010, 58, 254–268. [Google Scholar] [CrossRef]
- Qin, X.; Galloway-Peña, J.R.; Sillanpaa, J.; Roh, J.H.; Nallapareddy, S.R.; Chowdhury, S.; Bourgogne, A.; Choudhury, T.; Muzny, D.M.; Buhay, C.J.; et al. Complete genome sequence of Enterococcus faecium strain TX16 and comparative genomic analysis of Enterococcus faecium genomes. BMC Microbiol. 2012, 12, 135. [Google Scholar] [CrossRef]
- Markwart, R.; Willrich, N.; Eckmanns, T.; Werner, G.; Ayobami, O. Low Proportion of Linezolid and Daptomycin Resistance Among Bloodborne Vancomycin-Resistant Enterococcus faecium and Methicillin-Resistant Staphylococcus aureus Infections in Europe. Front. Microbiol. 2021, 12, 664199. [Google Scholar] [CrossRef]
- Yadav, G.; Thakuria, B.; Madan, M.; Agwan, V.; Pandey, A. Linezolid and Vancomycin Resistant Enterococci: A Therapeutic Problem. J. Clin. Diagn. Res. 2017, 11, GC07–GC11. [Google Scholar] [CrossRef]
- Fiedler, S.; Bender, J.K.; Klare, I.; Halbedel, S.; Grohmann, E.; Szewzyk, U.; Werner, G. Tigecycline resistance in clinical isolates of Enterococcus faecium is mediated by an upregulation of plasmid-encoded tetracycline determinants tet(L) and tet(M). J. Antimicrob. Chemother. 2015, 71, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.K.; Cattoir, V.; Hegstad, K.; Sadowy, E.; Coque, T.M.; Westh, H.; Hammerum, A.M.; Schaffer, K.; Burns, K.; Murchan, S.; et al. Update on prevalence and mechanisms of resistance to linezolid, tigecycline and daptomycin in enterococci in Europe: Towards a common nomenclature. Drug Resist. Updates 2018, 40, 25–39. [Google Scholar] [CrossRef]
- Hammerum, A.M. Enterococci of animal origin and their significance for public health. J. Anal. Methods Chem. 2012, 18, 619–625. [Google Scholar] [CrossRef]
- Zaheer, R.; Cook, S.R.; Barbieri, R.; Goji, N.; Cameron, A.; Petkau, A.; Polo, R.O.; Tymensen, L.; Stamm, C.; Song, J.; et al. Surveillance of Enterococcus spp. reveals distinct species and antimicrobial resistance diversity across a One-Health continuum. Sci. Rep. 2020, 10, 3937. [Google Scholar] [CrossRef]
- Willems, R.J.L.; Top, J.; van Den Braak, N.; Van Belkum, A.; Endtz, H.; Mevius, D.; Stobberingh, E.; van den Bogaard, A.; Van Embden, J.D.A. Host Specificity of Vancomycin-Resistant Enterococcus faecium. J. Infect. Dis. 2000, 182, 816–823. [Google Scholar] [CrossRef]
- Trott, D.J.; Turnidge, J.; Kovac, J.H.; Simjee, S.; Wilson, D.; Watts, J. Comparative macrolide use in humans and animals: Should macrolides be moved off the World Health Organisation’s critically important antimicrobial list? J. Antimicrob. Chemother. 2021, 76, 1955–1961. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.D.; Pratt, R.; Hart, W.S. Antibiotic resistance in animals. Commun. Dis. Intell. 2003, 27, S121–S126. [Google Scholar]
- Barton, M.D. Antibiotic use in animal feed and its impact on human health. Nutr. Res. Rev. 2000, 13, 279–299. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.; O’Dea, M.; Page, S.W.; Trott, D.J. Current and future antimicrobial resistance issues for the Australian pig industry. Anim. Prod. Sci. 2017, 57, 2398–2407. [Google Scholar] [CrossRef]
- Barlow, R.S.; McMillan, K.E.; Duffy, L.L.; Fegan, N.; Jordan, D.; Mellor, G.E. Antimicrobial resistance status of Enterococcus from Australian cattle populations at slaughter. PLoS ONE 2017, 12, e0177728. [Google Scholar] [CrossRef]
- O’dea, M.; Sahibzada, S.; Jordan, D.; Laird, T.; Lee, T.; Hewson, K.; Pang, S.; Abraham, R.; Coombs, G.W.; Harris, T.; et al. Genomic, Antimicrobial Resistance, and Public Health Insights into Enterococcus spp. from Australian Chickens. J. Clin. Microbiol. 2019, 57, e00319-19. [Google Scholar] [CrossRef]
- Messele, Y.E.; Hasoon, M.F.; Trott, D.J.; Veltman, T.; McMeniman, J.P.; Kidd, S.P.; Low, W.Y.; Petrovski, K.R. Longitudinal Analysis of Antimicrobial Resistance among Enterococcus Species Isolated from Australian Beef Cattle Faeces at Feedlot Entry and Exit. Animals 2022, 12, 2690. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Abecia, L.; Angarita, E.; Aravena, P.; Nora Arenas, G.; Ariza, C.; et al. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef]
- Myer, P.R.; Freetly, H.C.; Wells, J.E.; Smith, T.P.L.; Kuehn, L.A. Analysis of the gut bacterial communities in beef cattle and their association with feed intake, growth, and efficiency. J. Anim. Sci. 2017, 95, 3215–3224. [Google Scholar] [CrossRef]
- Ramos, S.; Silva, V.; Dapkevicius, M.d.L.E.; Igrejas, G.; Poeta, P. Enterococci, from Harmless Bacteria to a Pathogen. Microorganisms 2020, 8, 1118. [Google Scholar] [CrossRef]
- Lee, T.; Jordan, D.; Sahibzada, S.; Abraham, R.; Pang, S.; Coombs, G.W.; O’dea, M.; Abraham, S. Antimicrobial Resistance in Porcine Enterococci in Australia and the Ramifications for Human Health. Appl. Environ. Microbiol. 2021, 87, e03037-20. [Google Scholar] [CrossRef]
- Trościańczyk, A.; Nowakiewicz, A.; Osińska, M.; Łagowski, D.; Gnat, S.; Chudzik-Rząd, B. Comparative characteristics of sequence types, genotypes and virulence of multidrug-resistant Enterococcus faecium isolated from various hosts in eastern Poland. Spread of clonal complex 17 in humans and animals. Res. Microbiol. 2022, 173, 103925. [Google Scholar] [CrossRef]
- Lee, R.S.; Gonçalves da Silva, A.; Baines, S.L.; Strachan, J.; Ballard, S.; Carter, G.P.; Kwong, J.C.; Schultz, M.B.; Bulach, D.M.; Seemann, T. The changing landscape of vancomycin-resistant Enterococcus faecium in Australia: A population-level genomic study. J. Antimicrob. Chemother. 2018, 73, 3268–3278. [Google Scholar] [CrossRef]
- Freitas, A.R.; Tedim, A.P.; Duarte, B.; Elghaieb, H.; Abbassi, M.S.; Hassen, A.; Read, A.; Alves, V.; Novais, C.; Peixe, L. Linezolid-resistant (Tn6246::fexB-poxtA) Enterococcus faecium strains colonizing humans and bovines on different continents: Similarity without epidemiological link. J. Antimicrob. Chemother. 2020, 75, 2416–2423. [Google Scholar] [CrossRef]
- Werner, G.; Fleige, C.; Feßler, A.T.; Timke, M.; Kostrzewa, M.; Zischka, M.; Peters, T.; Kaspar, H.; Schwarz, S. Improved identification including MALDI-TOF mass spectrometry analysis of group D streptococci from bovine mastitis and subsequent molecular characterization of corresponding Enterococcus faecalis and Enterococcus faecium isolates. Vet. Microbiol. 2012, 160, 162–169. [Google Scholar] [CrossRef]
- Rajendiran, S.; Veloo, Y.; Thahir, S.S.A.; Shaharudin, R. Resistance towards Critically Important Antimicrobials among Enterococcus faecalis and E. faecium in Poultry Farm Environments in Selangor, Malaysia. Antibiotics 2022, 11, 1118. [Google Scholar] [CrossRef]
- Jung, Y.-H.; Shin, E.S.; Kim, O.; Yoo, J.S.; Lee, K.M.; Yoo, J.I.; Chung, G.T.; Lee, Y.S. Characterization of Two Newly Identified Genes, vgaD and vatG, Conferring Resistance to Streptogramin A in Enterococcus faecium. Antimicrob. Agents Chemother. 2010, 54, 4744–4749. [Google Scholar] [CrossRef]
- Li, W.; Hu, J.; Li, L.; Zhang, M.; Cui, Q.; Ma, Y.; Su, H.; Zhang, X.; Xu, H.; Wang, M. New Mutations in cls Lead to Daptomycin Resistance in a Clinical Vancomycin- and Daptomycin-Resistant Enterococcus faecium Strain. Front. Microbiol. 2022, 13, 896916. [Google Scholar] [CrossRef]
- ASTAG. Importance Ratings and Summary of Antibacterial Uses in Human and Animal Health in Australia. Available online: https://www.amr.gov.au/resources/importance-ratings-and-summary-antibacterial-uses-human-and-animal-health-australia2018 (accessed on 12 April 2023).
- De Graef, E.; Decostere, A.; De Leener, E.; Goossens, H.; Baele, M.; Haesebrouck, F. Prevalence and Mechanism of Resistance against Macrolides, Lincosamides, and Streptogramins among Enterococcus faecium Isolates from Food-Producing Animals and Hospital Patients in Belgium. Microb. Drug Resist. 2007, 13, 135–141. [Google Scholar] [CrossRef]
- Isnard, C.M.; Malbruny, B.; Leclercq, R.; Cattoir, V. Genetic Basis for In Vitro and In Vivo Resistance to Lincosamides, Streptogramins A, and Pleuromutilins (LS A P Phenotype) in Enterococcus faecium. Antimicrob. Agents Chemother. 2013, 57, 4463–4469. [Google Scholar] [CrossRef]
- Wang, S.; Guo, Y.; Lv, J.; Qi, X.; Li, D.; Chen, Z.; Zhang, X.; Wang, L.; Yu, F. Characteristic of Enterococcus faecium clinical isolates with quinupristin/dalfopristin resistance in China. BMC Microbiol. 2016, 16, 246. [Google Scholar] [CrossRef]
- Badger, S.; Sullivan, K.; Jordan, D.; Caraguel, C.; Page, S.; Cusack, P.; Frith, D.; Trott, D. Antimicrobial use and stewardship practices on Australian beef feedlots. Aust. Vet. J. 2020, 98, 37–47. [Google Scholar] [CrossRef]
- Lean, I.; Page, S.; Rabiee, A.; Willams, S. A Survey of Antibacterial Product Use in the Australian Cattle Industry; Meat and Livestock Australia Report; Meat & Livestock Australia Limited: North Sydney, Australia, 2013. [Google Scholar]
- APVMA. Findings of the Reconsideration of the Registration of Products Containing Virginiamycin, and Their Labels; APVMA: Canberra, Australia, 2004. [Google Scholar]
- Sparo, M.; Delpech, G.; García Allende, N. Impact on Public Health of the Spread of High-Level Resistance to Gentamicin and Vancomycin in Enterococci. Front. Microbiol. 2018, 9, 3073. [Google Scholar] [CrossRef]
- Bates, J.; Jordens, J.Z.; Griffiths, D.T. Farm animals as a putative reservoir for vancomycin-resistant enterococcal infection in man. J. Antimicrob. Chemother. 1994, 34, 507–514. [Google Scholar] [CrossRef]
- Descheemaeker, P.R.; Chapelle, S.; Devriese, L.A.; Butaye, P.; Vandamme, P.; Goossens, H. Comparison of Glycopeptide-Resistant Enterococcus faecium Isolates and Glycopeptide Resistance Genes of Human and Animal Origins. Antimicrob. Agents Chemother. 1999, 43, 2032–2037. [Google Scholar] [CrossRef]
- Klare, I.; Heier, H.; Claus, H.; Reissbrodt, R.; Witte, W. vanA-mediated high-level glycopeptide resistance in Enterococcus faecium from animal husbandry. FEMS Microbiol. Lett. 1995, 125, 165–171. [Google Scholar] [CrossRef]
- Yoshimura, H.; Ishimaru, M.; Endoh, Y.S.; Suginaka, M.; Yamatani, S. Isolation of Glycopeptide-Resistant Enterococci from Chickens in Japan. Antimicrob. Agents Chemother. 1998, 42, 3333. [Google Scholar] [CrossRef]
- Aarestrup, F.M. Occurrence of Glycopeptide Resistance among Enterococcus faecium Isolates from Conventional and Ecological Poultry Farms. Microb. Drug Resist. 1995, 1, 255–257. [Google Scholar] [CrossRef]
- Bager, F.; Madsen, M.; Christensen, J.; Aarestrup, F.M. Avoparcin used as a growth promoter is associated with the occurrence of vancomycin-resistant on Danish poultry and pig farms. Prev. Vet. Med. 1997, 31, 95–112. [Google Scholar] [CrossRef]
- Althaus, F.R. Ban on antimicrobial growth promoters: A safety advantage for consumers? Schweiz. Arch. Tierheilkd. 1999, 141, 103–107. [Google Scholar] [PubMed]
- Sabol, K.; Patterson, J.E.; Lewis, J.S.; Owens, A.; Cadena, J.; Jorgensen, J.H. Emergence of Daptomycin Resistance in Enterococcus faecium during Daptomycin Therapy. Antimicrob. Agents Chemother. 2005, 49, 1664–1665. [Google Scholar] [CrossRef] [PubMed]
- Werth, B.J.; Steed, M.E.; Ireland, C.E.; Tran, T.T.; Nonejuie, P.; Murray, B.E.; Rose, W.E.; Sakoulas, G.; Pogliano, J.; Arias, C.A.; et al. Defining Daptomycin Resistance Prevention Exposures in Vancomycin-Resistant Enterococcus faecium and E. faecalis. Antimicrob. Agents Chemother. 2014, 58, 5253–5261. [Google Scholar] [CrossRef] [PubMed]
- Lellek, H.; Franke, G.C.; Ruckert, C.; Wolters, M.; Wolschke, C.; Christner, M.; Büttner, H.; Alawi, M.; Kröger, N.; Rohde, H. Emergence of daptomycin non-susceptibility in colonizing vancomycin-resistant Enterococcus faecium isolates during daptomycin therapy. Int. J. Med. Microbiol. 2015, 305, 902–909. [Google Scholar] [CrossRef]
- Li, J.; Zhang, H.; Ning, J.; Sajid, A.; Cheng, G.; Yuan, Z.; Hao, H. The nature and epidemiology of OqxAB, a multidrug efflux pump. Antimicrob. Resist. Infect. Control 2019, 8, 44. [Google Scholar] [CrossRef]
- Hernando-Amado, S.; Blanco, P.; Alcalde-Rico, M.; Corona, F.; Reales-Calderón, J.A.; Sánchez, M.B.; Martínez, J.L. Multidrug efflux pumps as main players in intrinsic and acquired resistance to antimicrobials. Drug Resist. Updates 2016, 28, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Amin, N.E.; Jalal, S.; Wretlind, B. Alterations in GyrA and ParC Associated with Fluoroquinolone Resistance in Enterococcus faecium. Antimicrob. Agents Chemother. 1999, 43, 947–949. [Google Scholar] [CrossRef] [PubMed]
- Bennett, P.M. Plasmid encoded antibiotic resistance: Acquisition and transfer of antibiotic resistance genes in bacteria. Br. J. Pharmacol. 2008, 153, S347–S357. [Google Scholar] [CrossRef]
- Vrancianu, C.O.; Popa, L.I.; Bleotu, C.; Chifiriuc, M.C. Targeting Plasmids to Limit Acquisition and Transmission of Antimicrobial Resistance. Front. Microbiol. 2020, 11, 761. [Google Scholar] [CrossRef]
- Freitas, A.R.; Coque, T.M.; Novais, C.; Hammerum, A.M.; Lester, C.H.; Zervos, M.J.; Donabedian, S.; Jensen, L.B.; Francia, M.V.; Baquero, F.; et al. Human and Swine Hosts Share Vancomycin-Resistant Enterococcus faecium CC17 and CC5 and Enterococcus faecalis CC2 Clonal Clusters Harboring Tn on Indistinguishable Plasmids. J. Clin. Microbiol. 2011, 49, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Tedim, A.P.; Lanza, V.F.; Rodríguez, C.M.; Freitas, A.R.; Novais, C.; Peixe, L.; Baquero, F.; Coque, T.M. Fitness cost of vancomycin-resistant Enterococcus faecium plasmids associated with hospital infection outbreaks. J. Antimicrob. Chemother. 2021, 76, 2757–2764. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C.; Jonas, D.; Huber, I.; Karygianni, L.; Wölber, J.; Hellwig, E.; Arweiler, N.; Vach, K.; Wittmer, A.; Al-Ahmad, A. Enterococcus faecalis from Food, Clinical Specimens, and Oral Sites: Prevalence of Virulence Factors in Association with Biofilm Formation. Front. Microbiol. 2016, 6, 1534. [Google Scholar] [CrossRef] [PubMed]
- Coque, T.M.; Willems, R.J.; Fortún, J.; Top, J.; Diz, S.; Loza, E.; Cantón, R.; Baquero, F. Population Structure of Enterococcus faecium Causing Bacteremia in a Spanish University Hospital: Setting the Scene for a Future Increase in Vancomycin Resistance? Antimicrob. Agents Chemother. 2005, 49, 2693–2700. [Google Scholar] [CrossRef] [PubMed]
- Galloway-Peña, J.R.; Nallapareddy, S.R.; Arias, C.A.; Eliopoulos, G.M.; Murray, B.E. Analysis of Clonality and Antibiotic Resistance among Early Clinical Isolates of Enterococcus faecium in the United States. J. Infect. Dis. 2009, 200, 1566–1573. [Google Scholar] [CrossRef]
- Werner, G.; Coque, T.M.; Hammerum, A.M.; Hope, R.; Hryniewicz, W.; Johnson, A.; Klare, I.; Kristinsson, K.G.; Leclercq, R.; Lester, C.H.; et al. Emergence and spread of vancomycin resistance among enterococci in Europe. Eurosurveillance 2008, 13, 19046. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Didelot, X.; Wilson, D.J. ClonalFrameML: Efficient Inference of Recombination in Whole Bacterial Genomes. PLoS Comput. Biol. 2015, 11, e1004041. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontéen, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-Time Whole-Genome Sequencing for Routine Typing, Surveillance, and Outbreak Detection of Verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef]
- Tetzschner, A.M.M.; Johnson, J.R.; Johnston, B.D.; Lund, O.; Scheutz, F. In Silico Genotyping of Escherichia coli Isolates for Extraintestinal Virulence Genes by Use of Whole-Genome Sequencing Data. J. Clin. Microbiol. 2020, 58, e01269-20. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcìa-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Voldby Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids. Antimicrob using PlasmidFinder and plasmid multilocus sequence typing. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- EFSA. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2016. EFSA J. 2018, 16, e05182. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).