

Increasing Fluroquinolone Susceptibility and Genetic Diversity of ESBL-Producing E. coli from the Lower Respiratory Tract during the COVID-19 Pandemic

Abstract
:1. Introduction
2. Results
2.1. LRT ESBL-EC Isolates from Both Study Periods
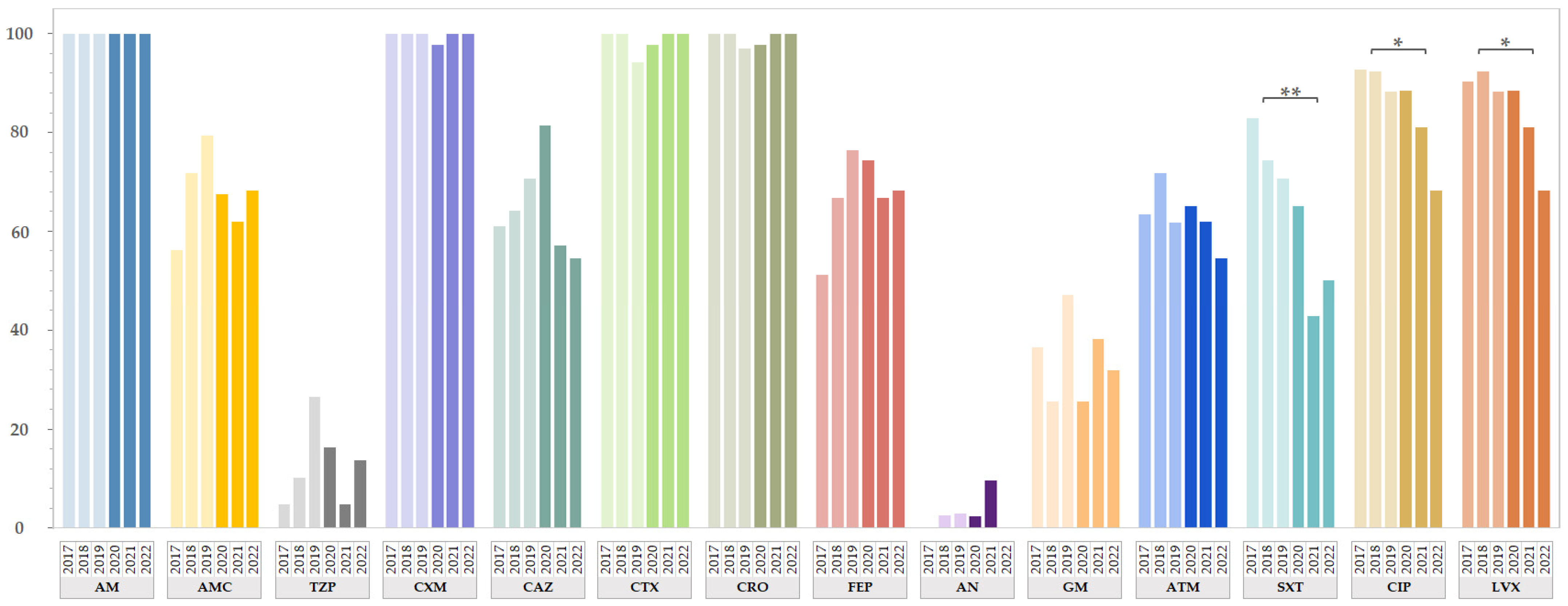
2.2. Antimicrobial Resistance of ESBL-EC from Both Study Periods
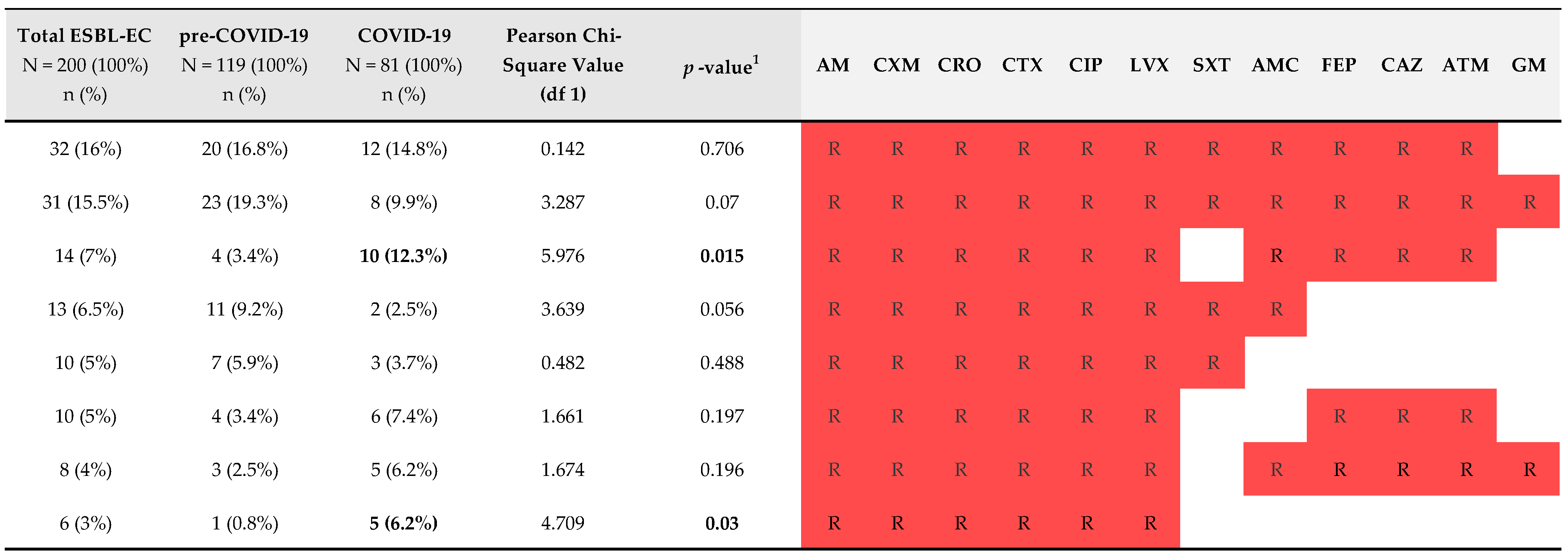
2.3. Comparison of Genotype Data of ESBL-EC Isolated from LRTs from Both Study Periods
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Patients
4.2. Antimicrobial Susceptibility Testing
4.3. Molecular Characterization of ESBL-EC Isolates
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- WHO. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 5 April 2024).
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. The Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Massella, E.; Giacometti, F.; Bonilauri, P.; Reid, C.J.; Djordjevic, S.P.; Merialdi, G.; Bacci, C.; Fiorentini, L.; Massi, P.; Bardasi, L.; et al. Antimicrobial Resistance Profile and ExPEC Virulence Potential in Commensal Escherichia coli of Multiple Sources. Antibiotics 2021, 10, 351. [Google Scholar] [CrossRef] [PubMed]
- Noster, J.; Thelen, P.; Hamprecht, A. Detection of Multidrug-Resistant Enterobacterales—From ESBLs to Carbapenemases. Antibiotics 2021, 10, 1140. [Google Scholar] [CrossRef] [PubMed]
- Baggio, D.; Ananda-Rajah, M.R. Fluoroquinolone antibiotics and adverse events. Aust. Prescr. 2021, 44, 161–164. [Google Scholar] [CrossRef]
- Dalhoff, A. Global fluoroquinolone resistance epidemiology and implictions for clinical use. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 976273. [Google Scholar] [CrossRef]
- Karampela, I.; Dalamaga, M. Could Respiratory Fluoroquinolones, Levofloxacin and Moxifloxacin, Prove to be Beneficial as an Adjunct Treatment in COVID-19? Arch. Med. Res. 2020, 51, 741–742. [Google Scholar] [CrossRef] [PubMed]
- Marciniec, K.; Beberok, A.; Pęcak, P.; Boryczka, S.; Wrześniok, D. Ciprofloxacin and moxifloxacin could interact with SARS-CoV-2 protease: Preliminary in silico analysis. Pharmacol. Rep. PR 2020, 72, 1553–1561. [Google Scholar] [CrossRef]
- Olivieri, R.; Vannini, P.; Corzani, A.; Bianco, M.T.; Franchi, F.; Cusi, M.G.; Scolletta, S.; Arena, F.; Basagni, C.; Gusinu, R.; et al. Rapid Decrease in Fluoroquinolones Consumption following Implementation of a Simple Antimicrobial Stewardship Bundled Intervention in a University Hospital during the COVID-19 Pandemic. Antibiotics 2023, 12, 694. [Google Scholar] [CrossRef]
- WHO. Global Action Plan on Antimicrobial Resistance. Available online: https://www.who.int/publications/i/item/9789241509763 (accessed on 10 June 2024).
- Kocsis, B.; Gulyás, D.; Szabó, D. Emergence and Dissemination of Extraintestinal Pathogenic High-Risk International Clones of Escherichia coli. Life 2022, 12, 2077. [Google Scholar] [CrossRef]
- Masters, P.A.; O’Bryan, T.A.; Zurlo, J.; Miller, D.Q.; Joshi, N. Trimethoprim-sulfamethoxazole revisited. Arch. Intern. Med. 2003, 163, 402–410. [Google Scholar] [CrossRef]
- Fukushige, M.; Ngo, N.H.; Lukmanto, D.; Fukuda, S.; Ohneda, O. Effect of the COVID-19 pandemic on antibiotic consumption: A systematic review comparing 2019 and 2020 data. Front. Public Health 2022, 10, 946077. [Google Scholar] [CrossRef] [PubMed]
- Romaszko-Wojtowicz, A.; Tokarczyk-Malesa, K.; Doboszyńska, A.; Glińska-Lewczuk, K. Impact of COVID-19 on antibiotic usage in primary care: A retrospective analysis. Sci. Rep. 2024, 14, 4798. [Google Scholar] [CrossRef] [PubMed]
- Siewierska, M.; Gajda, M.; Opalska, A.; Brudło, M.; Krzyściak, P.; Gryglewska, B.; Różańska, A.; Wójkowska-Mach, J. Hospital antibiotic consumption-an interrupted time series analysis of the early and late phases of the COVID-19 pandemic in Poland, a retrospective study. Pharmacol. Rep. PR 2023, 75, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Hrovat, K.; Molan, K.; Seme, K.; Ambrožič Avguštin, J. Molecular characterization of extended-spectrum β-lactamase-producing Escherichia coli isolated from lower respiratory tract samples between 2002 and 2019 in the Central Slovenia region. Ann. Clin. Microbiol. Antimicrob. 2024, 23, 6. [Google Scholar] [CrossRef] [PubMed]
- CDC. COVID-19: U.S. Impact on Antimicrobial Resistance, Special Report 2022. Available online: https://www.cdc.gov/drugresistance/covid19.html (accessed on 16 August 2024).
- Petrakis, V.; Panopoulou, M.; Rafailidis, P.; Lemonakis, N.; Lazaridis, G.; Terzi, I.; Papazoglou, D.; Panagopoulos, P. The Impact of the COVID-19 Pandemic on Antimicrobial Resistance and Management of Bloodstream Infections. Pathogens 2023, 12, 780. [Google Scholar] [CrossRef]
- Khouja, T.; Mitsantisuk, K.; Tadrous, M.; Suda, K.J. Global consumption of antimicrobials: Impact of the WHO Global Action Plan on Antimicrobial Resistance and 2019 coronavirus pandemic (COVID-19). J. Antimicrob. Chemother. 2022, 77, 1491–1499. [Google Scholar] [CrossRef]
- Knight, G.M.; Glover, R.E.; McQuaid, C.F.; Olaru, I.D.; Gallandat, K.; Leclerc, Q.J.; Fuller, N.M.; Willcocks, S.J.; Hasan, R.; van Kleef, E.; et al. Antimicrobial resistance and COVID-19: Intersections and implications. eLife 2021, 10, e64139. [Google Scholar] [CrossRef]
- Rusic, D.; Vilovic, M.; Bukic, J.; Leskur, D.; Seselja Perisin, A.; Kumric, M.; Martinovic, D.; Petric, A.; Modun, D.; Bozic, J. Implications of COVID-19 Pandemic on the Emergence of Antimicrobial Resistance: Adjusting the Response to Future Outbreaks. Life 2021, 11, 220. [Google Scholar] [CrossRef]
- ECDC. Antimicrobial Resistance in the EU/EEA (EARS-Net)—Annual Epidemiological Report for 2022. Available online: https://www.ecdc.europa.eu/en/publications-data/surveillance-antimicrobial-resistance-europe-2022 (accessed on 12 June 2024).
- ECDC. Antimicrobial Consumption in the EU/EEA (ESAC-Net)—Annual Epidemiological Report for 2022. Available online: https://www.ecdc.europa.eu/en/publications-data/surveillance-antimicrobial-consumption-europe-2022 (accessed on 12 June 2024).
- EMA. Quinolone- and Fluoroquinolone-Containing Medicinal Products-Referral. Available online: https://www.ema.europa.eu/en/medicines/human/referrals/quinolone-fluoroquinolone-containing-medicinal-products (accessed on 12 June 2024).
- Abdelaziz Abdelmoneim, S.; Mohamed Ghazy, R.; Anwar Sultan, E.; Hassaan, M.A.; Anwar Mahgoub, M. Antimicrobial resistance burden pre and post-COVID-19 pandemic with mapping the multidrug resistance in Egypt: A comparative cross-sectional study. Sci. Rep. 2024, 14, 7176. [Google Scholar] [CrossRef]
- Araújo, M.R.B.; Sant’Anna, L.O.; Santos, N.; Seabra, L.F.; Santos, L.S.D. Monitoring fluoroquinolone resistance among ESBL-positive and ESBL-negative Escherichia coli strains isolated from urinary tract infections: An alert for empirical treatment. Rev. Soc. Bras. Med. Trop. 2023, 56, e0513. [Google Scholar] [CrossRef]
- Kettani Halabi, M.; Lahlou, F.A.; Diawara, I.; El Adouzi, Y.; Marnaoui, R.; Benmessaoud, R.; Smyej, I. Antibiotic Resistance Pattern of Extended Spectrum Beta Lactamase Producing Escherichia coli Isolated From Patients with Urinary Tract Infection in Morocco. Front. Cell Infect. Microbiol. 2021, 11, 720701. [Google Scholar] [CrossRef] [PubMed]
- Pitout, J.D.; DeVinney, R. Escherichia coli ST131: A multidrug-resistant clone primed for global domination. F1000Research 2017, 6. [Google Scholar] [CrossRef]
- Pitout, J.D.D.; Peirano, G.; Chen, L.; DeVinney, R.; Matsumura, Y. Escherichia coli ST1193: Following in the Footsteps of E. coli ST131. Antimicrob. Agents Chemother. 2022, 66, e0051122. [Google Scholar] [CrossRef] [PubMed]
- Tomassetti, S.; Ciani, L.; Luzzi, V.; Gori, L.; Trigiani, M.; Giuntoli, L.; Lavorini, F.; Poletti, V.; Ravaglia, C.; Torrego, A.; et al. Utility of bronchoalveolar lavage for COVID-19: A perspective from the Dragon consortium. Front. Med. 2024, 11, 1259570. [Google Scholar] [CrossRef] [PubMed]
- Ružić Gorenjec, N.; Kejžar, N.; Manevski, D.; Pohar Perme, M.; Vratanar, B.; Blagus, R. COVID-19 in Slovenia, from a Success Story to Disaster: What Lessons Can Be Learned? Life 2021, 11, 1045. [Google Scholar] [CrossRef]
- Zagar, T.; Tomsic, S.; Zadnik, V.; Bric, N.; Birk, M.; Vurzer, B.; Mihor, A.; Lokar, K.; Oblak, I. Impact of the COVID-19 epidemic on cancer burden and cancer care in Slovenia: A follow-up study. Radiol. Oncol. 2022, 56, 488–500. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19—27 July 2020. Available online: https://www.who.int/director-general/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---27-july-2020 (accessed on 10 June 2024).
- Hrovat, K.; Zupančič, J.; Seme, K.; Avguštin, J.A. QAC Resistance Genes in ESBL-Producing E. coli Isolated from Patients with Lower Respiratory Tract Infections in the Central Slovenia Region-A 21-Year Survey. Trop. Med. Infect. Dis. 2023, 8, 273. [Google Scholar] [CrossRef]
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Versions 4.0 to 9.0. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 5 April 2024).
- EUCAST. Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance. Version 2.0. pp. 1–43. Available online: http://www.eucast.org/resistance_mechanisms/ (accessed on 5 April 2024).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Wolfensberger, A.; Kuster, S.P.; Marchesi, M.; Zbinden, R.; Hombach, M. The effect of varying multidrug-resistence (MDR) definitions on rates of MDR gram-negative rods. Antimicrob. Resist. Infect. Control. 2019, 8, 193. [Google Scholar] [CrossRef] [PubMed]
- Le Bouguenec, C.; Archambaud, M.; Labigne, A. Rapid and specific detection of the pap, afa, and sfa adhesin-encoding operons in uropathogenic Escherichia coli strains by polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 1189–1193. [Google Scholar] [CrossRef]
- Woodford, N.; Fagan, E.J.; Ellington, M.J. Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum (beta)-lactamases. J. Antimicrob. Chemother. 2006, 57, 154–155. [Google Scholar] [CrossRef] [PubMed]
- Dallenne, C.; Da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Cattoir, V.; Poirel, L.; Rotimi, V.; Soussy, C.J.; Nordmann, P. Multiplex PCR for detection of plasmid-mediated quinolone resistance qnr genes in ESBL-producing enterobacterial isolates. J. Antimicrob. Chemother. 2007, 60, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Cattoir, V.; Weill, F.X.; Poirel, L.; Fabre, L.; Soussy, C.J.; Nordmann, P. Prevalence of qnr genes in Salmonella in France. J. Antimicrob. Chemother. 2007, 59, 751–754. [Google Scholar] [CrossRef] [PubMed]
- Kraychete, G.B.; Botelho, L.A.; Campana, E.H.; Picão, R.C.; Bonelli, R.R. Updated Multiplex PCR for Detection of All Six Plasmid-Mediated qnr Gene Families. Antimicrob. Agents Chemother. 2016, 60, 7524–7526. [Google Scholar] [CrossRef]
- Wareham, D.W.; Umoren, I.; Khanna, P.; Gordon, N.C. Allele-specific polymerase chain reaction (PCR) for rapid detection of the aac(6′)-Ib-cr quinolone resistance gene. Int. J. Antimicrob. Agents 2010, 36, 476–477. [Google Scholar] [CrossRef]
- Cattoir, V.; Poirel, L.; Nordmann, P. Plasmid-mediated quinolone resistance pump QepA2 in an Escherichia coli isolate from France. Antimicrob. Agents Chemother. 2008, 52, 3801–3804. [Google Scholar] [CrossRef]
- Zou, L.; Meng, J.; McDermott, P.F.; Wang, F.; Yang, Q.; Cao, G.; Hoffmann, M.; Zhao, S. Presence of disinfectant resistance genes in Escherichia coli isolated from retail meats in the USA. J. Antimicrob. Chemother. 2014, 69, 2644–2649. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.H.; Sørensen, S.J.; Jørgensen, H.S.; Jensen, L.B. The prevalence of the OqxAB multidrug efflux pump amongst olaquindox-resistant Escherichia coli in pigs. Microb. Drug Resist. 2005, 11, 378–382. [Google Scholar] [CrossRef]
- Ni, Q.; Tian, Y.; Zhang, L.; Jiang, C.; Dong, D.; Li, Z.; Mao, E.; Peng, Y. Prevalence and quinolone resistance of fecal carriage of extended-spectrum β-lactamase-producing Escherichia coli in 6 communities and 2 physical examination center populations in Shanghai, China. Diagn. Microbiol. Infect. Dis. 2016, 86, 428–433. [Google Scholar] [CrossRef]
- Versalovic, J.; Koeuth, T.; Lupski, J.R. Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed]
- Doumith, M.; Day, M.; Ciesielczuk, H.; Hope, R.; Underwood, A.; Reynolds, R.; Wain, J.; Livermore, D.M.; Woodford, N. Rapid identification of major Escherichia coli sequence types causing urinary tract and bloodstream infections. J. Clin. Microbiol. 2015, 53, 160–166. [Google Scholar] [CrossRef]
- Clermont, O.; Dhanji, H.; Upton, M.; Gibreel, T.; Fox, A.; Boyd, D.; Mulvey, M.R.; Nordmann, P.; Ruppé, E.; Sarthou, J.L.; et al. Rapid detection of the O25b-ST131 clone of Escherichia coli encompassing the CTX-M-15-producing strains. J. Antimicrob. Chemother. 2009, 64, 274–277. [Google Scholar] [CrossRef]
- Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Starčič Erjavec, M.; Palandačić, A.; Žgur-Bertok, D.; Ambrožič Avguštin, J. Genetic background of uropathogenic Escherichia coli isolates from Slovenia in relation to fluoroquinolone and sulfamethoxazole/trimethoprim resistance. Acta Biol. Slov. 2011, 54, 5–13. [Google Scholar] [CrossRef]
- Johnson, J.R.; Russo, T.A.; Tarr, P.I.; Carlino, U.; Bilge, S.S.; Vary, J.C., Jr.; Stell, A.L. Molecular epidemiological and phylogenetic associations of two novel putative virulence genes, iha and iroN (E. coli), among Escherichia coli isolates from patients with urosepsis. Infect. Immun. 2000, 68, 304–307. [Google Scholar] [CrossRef]
- Janben, T.; Schwarz, C.; Preikschat, P.; Voss, M.; Philipp, H.C.; Wieler, L.H. Virulence-associated genes in avian pathogenic Escherichia coli (APEC) isolated from internal organs of poultry having died from colibacillosis. Int. J. Med. Microbiol. 2001, 291, 371–378. [Google Scholar] [CrossRef]
- Johnson, J.R.; Brown, J.J. A novel multiply primed polymerase chain reaction assay for identification of variant papG genes encoding the Gal(alpha 1-4)Gal-binding PapG adhesins of Escherichia coli. J. Infect. Dis. 1996, 173, 920–926. [Google Scholar] [CrossRef]
- Ruiz, J.; Navia, M.M.; Vila, J.; Gascón, J. Prevalence of the sat gene among clinical isolates of Shigella spp. causing travelers’ diarrhea: Geographical and specific differences. J. Clin. Microbiol. 2002, 40, 1565–1566. [Google Scholar] [CrossRef] [PubMed]
- Parham, N.J.; Pollard, S.J.; Desvaux, M.; Scott-Tucker, A.; Liu, C.; Fivian, A.; Henderson, I.R. Distribution of the serine protease autotransporters of the Enterobacteriaceae among extraintestinal clinical isolates of Escherichia coli. J. Clin. Microbiol. 2005, 43, 4076–4082. [Google Scholar] [CrossRef] [PubMed]
- Schubert, S.; Rakin, A.; Karch, H.; Carniel, E.; Heesemann, J. Prevalence of the “high-pathogenicity island” of Yersinia species among Escherichia coli strains that are pathogenic to humans. Infect. Immun. 1998, 66, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Terai, A.; Yuri, K.; Kurazono, H.; Takeda, Y.; Yoshida, O. Detection of urovirulence factors in Escherichia coli by multiplex polymerase chain reaction. FEMS Immunol. Med. Microbiol. 1995, 12, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Stapleton, A.E.; Russo, T.A.; Scheutz, F.; Brown, J.J.; Maslow, J.N. Characteristics and prevalence within serogroup O4 of a J96-like clonal group of uropathogenic Escherichia coli O4:H5 containing the class I and class III alleles of papG. Infect. Immun. 1997, 65, 2153–2159. [Google Scholar] [CrossRef] [PubMed]
- Vadnov, M.; Barbič, D.; Žgur-Bertok, D.; Erjavec, M.S. Escherichia coli isolated from feces of brown bears (Ursus arctos) have a lower prevalence of human extraintestinal pathogenic E. coli virulence-associated genes. Can. J. Vet. Res. = Rev. Can. De Rech. Vet. 2017, 81, 59–63. [Google Scholar]
- Trkov, M.; Dovečar, D.; Paragi, M.; Ambrožič Avguštin, J. Phylogenetic grouping of Escherichia coli isolates from patients’ stool samples with diarrhoea. Clin. Microbiol. Infect. 2008, 14, 157. [Google Scholar]

{kind=link}
{kind=link}
| Pre-COVID-19 N = 119 (100%) n (%) | COVID-19 N = 81 (100%) n (%) | Pearson’s Chi-Squared Value (df 1) | p Value 1 | |
|---|---|---|---|---|
| Average age | 67.3 | 70.8 | / | / |
| 0–10 years | 11 (9.2%) | 2 (2.5%) | 3.639 | 0.056 |
| 11–20 years | 1 (0.8%) | 0 (0%) | 0.684 | 0.408 |
| 21–30 years | 1 (0.8%) | 1 (1.2%) | 0.076 | 0.783 |
| 31–40 years | 0 (0%) | 1 (1.2%) | 1.477 | 0.224 |
| 41–50 years | 7 (5.9%) | 5 (6.2%) | 0.007 | 0.932 |
| 51–60 years | 7 (5.9%) | 5 (6.2%) | 0.007 | 0.932 |
| 61–70 years | 20 (16.8%) | 18 (22.2%) | 0.918 | 0.338 |
| 71–80 years | 28 (23.5%) | 20 (24.7%) | 0.036 | 0.85 |
| 81–90 years | 36 (30.3%) | 27 (33.3%) | 0.212 | 0.645 |
| 91+ years | 8 (6.7%) | 2 (2.5%) | 1.836 | 0.175 |
| Male | 70 (58.8%) | 57 (70.4%) | 2.772 | 0.096 |
| Female | 49 (41.2%) | 24 (29.6%) | 2.772 | 0.096 |
| Tracheal aspirate | 87 (73.1%) | 50 (61.7%) | 2.893 | 0.089 |
| Sputum | 30 (25.2%) | 21 (25.9%) | 0.013 | 0.909 |
| Bronchoalveolar lavage | 2 (1.7%) | 10 (12.3%) | 9.72 | 0.002 |
| Pre-COVID-19 N = 119 (100%) n (%) | COVID-19 N = 81 (100%) n (%) | Pearson’s Chi-Squared Value (df 1) | p Value 1 | |
|---|---|---|---|---|
| Plasmid-mediated quinolone resistance genes | ||||
| qnrA | 0 (0%) | 0 (0%) | / | / |
| qnrB | 0 (0%) | 0 (0%) | / | / |
| qnrS | 3 (2.5%) | 4 (4.9%) | 0.834 | 0.361 |
| qnrC | 0 (0%) | 0 (0%) | / | / |
| qnrD | 0 (0%) | 0 (0%) | / | / |
| qnrVC | 0 (0%) | 0 (0%) | / | / |
| qepA | 0 (0%) | 0 (0%) | / | / |
| aac(6′)-I-cr | 46 (38.7%) | 20 (24.7%) | 4.25 | 0.039 |
| β-lactamase group genes | ||||
| blaCTX-M-1 | 101 (84.9%) | 61 (75.3%) | 2.865 | 0.091 |
| blaCTX-M-2 | 0 (0%) | 2 (2.5%) | 2.968 | 0.085 |
| blaCTX-M-9 | 15 (12.6%) | 15 (18.5%) | 1.322 | 0.25 |
| blaCTX-M-8 | 0 (0%) | 0 (0%) | / | / |
| blaCTX-M-25 | 0 (0%) | 0 (0%) | / | / |
| blaTEM | 63 (52.9%) | 26 (32.1%) | 8.477 | 0.004 |
| blaSHV | 1 (0.8%) | 0 (0%) | 0.684 | 0.408 |
| blaOXA | 47 (39.5%) | 24 (29.6%) | 2.049 | 0.152 |
| Biocide resistance genes encoded on mobile genetic elements | ||||
| qacEΔ1 | 62 (52.1%) | 24 (29.6%) | 9.929 | 0.002 |
| qacE | 0 (0%) | 0 (0%) | / | / |
| qacF/H/I | 0 (0%) | 1 (1.2%) | 1.477 | 0.224 |
| qacG | 0 (0%) | 0 (0%) | / | / |
| sugE (p) | 0 (0%) | 0 (0%) | / | / |
| Chromosome-encoded biocide resistance genes | ||||
| emrE | 92 (77.3%) | 65 (80.2%) | 0.246 | 0.62 |
| mdfA | 119 (100%) | 81 (100%) | / | / |
| sugE (c) | 119 (100%) | 81 (100%) | / | / |
| ydgE | 119 (100%) | 81 (100%) | / | / |
| ydgF | 119 (100%) | 81 (100%) | / | / |
| Integrons | ||||
| int1 | 33 (27.7%) | 37 (45.7%) | 6.824 | 0.009 |
| int2 | 0 (0%) | 1 (1.2%) | 1.477 | 0.224 |
| int3 | 0 (0%) | 0 (0%) | / | / |
| Pre-COVID-19 N = 119 (100%) n (%) | COVID-19 N = 81 (100%) n (%) | Pearson’s Chi-Squared Value (df 1) | p Value 1 | |
|---|---|---|---|---|
| EP1 | 54 (45.4%) | 18 (22.2%) | 11.216 | 0.001 |
| EP2 | 8 (6.7%) | 0 (0%) | 5.672 | 0.017 |
| EP3 | 21 (17.6%) | 23 (28.4%) | 3.244 | 0.072 |
| EP4 | 3 (2.5%) | 9 (11.1%) | 6.306 | 0.012 |
| EP5 | 1 (0.8%) | 1 (1.2%) | 0.076 | 0.783 |
| EP6 | 0 (0%) | 2 (2.5%) | 2.968 | 0.085 |
| EPx | 32 (26.9%) | 28 (34.6%) | 1.353 | 0.245 |
| Pre-COVID-19 N = 119 (100%) n (%) | COVID-19 N = 81 (100%) n (%) | Pearson’s Chi-Squared Value (df 1) | p Value 1 | |
|---|---|---|---|---|
| Adhesins | ||||
| afa/dra | 31 (26.1%) | 13 (16%) | 2.809 | 0.094 |
| fimH | 115 (96.6%) | 77 (95.1%) | 0.312 | 0.576 |
| iha | 86 (72.3%) | 50 (61.7%) | 2.461 | 0.117 |
| papC | 15 (12.6%) | 14 (17.3%) | 0.851 | 0.356 |
| papGII | 12 (10.1%) | 11 (13.6%) | 0.579 | 0.447 |
| Autotransporters | ||||
| fluA | 88 (73.9%) | 64 (79%) | 0.677 | 0.411 |
| sat | 26 (21.8%) | 53 (65.4%) | 38.308 | <0.001 |
| tsh | 0 (0%) | 4 (4.9%) | 5.996 | 0.014 |
| vat | 4 (3.4%) | 5 (6.2%) | 0.886 | 0.346 |
| Protectins | ||||
| iss | 12 (10.1%) | 12 (14.8%) | 1.021 | 0.312 |
| kpsMTII | 81 (68.1%) | 55 (67.9%) | 0.001 | 0.98 |
| ompTAPEC | 14 (11.8%) | 13 (16%) | 0.758 | 0.384 |
| traT | 98 (82.4%) | 69 (85.2%) | 0.281 | 0.596 |
| Iron acquisition systems | ||||
| fyuA | 105 (88.2%) | 68 (84%) | 0.758 | 0.384 |
| iroN | 15 (12.6%) | 13 (16%) | 0.475 | 0.491 |
| irp2 | 107 (89.9%) | 69 (85.2%) | 1.021 | 0.312 |
| iucD | 97 (81.5%) | 70 (86.4%) | 0.842 | 0.359 |
| iutA | 98 (82.4%) | 70 (86.4%) | 0.593 | 0.441 |
| Toxins | ||||
| ehxA | 0 (0%) | 0 (0%) | / | / |
| hlyA | 14 (11.8%) | 9 (11.1%) | 0.02 | 0.887 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hrovat, K.; Seme, K.; Ambrožič Avguštin, J. Increasing Fluroquinolone Susceptibility and Genetic Diversity of ESBL-Producing E. coli from the Lower Respiratory Tract during the COVID-19 Pandemic. Antibiotics 2024, 13, 797. https://doi.org/10.3390/antibiotics13090797
Hrovat K, Seme K, Ambrožič Avguštin J. Increasing Fluroquinolone Susceptibility and Genetic Diversity of ESBL-Producing E. coli from the Lower Respiratory Tract during the COVID-19 Pandemic. Antibiotics. 2024; 13(9):797. https://doi.org/10.3390/antibiotics13090797
Chicago/Turabian StyleHrovat, Katja, Katja Seme, and Jerneja Ambrožič Avguštin. 2024. "Increasing Fluroquinolone Susceptibility and Genetic Diversity of ESBL-Producing E. coli from the Lower Respiratory Tract during the COVID-19 Pandemic" Antibiotics 13, no. 9: 797. https://doi.org/10.3390/antibiotics13090797