
Insights into the Virulence and Antimicrobial Resistance of Staphylococcus hyicus Isolates from Spanish Swine Farms

 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. Isolation and Characterization of Staphylococcus hyicus
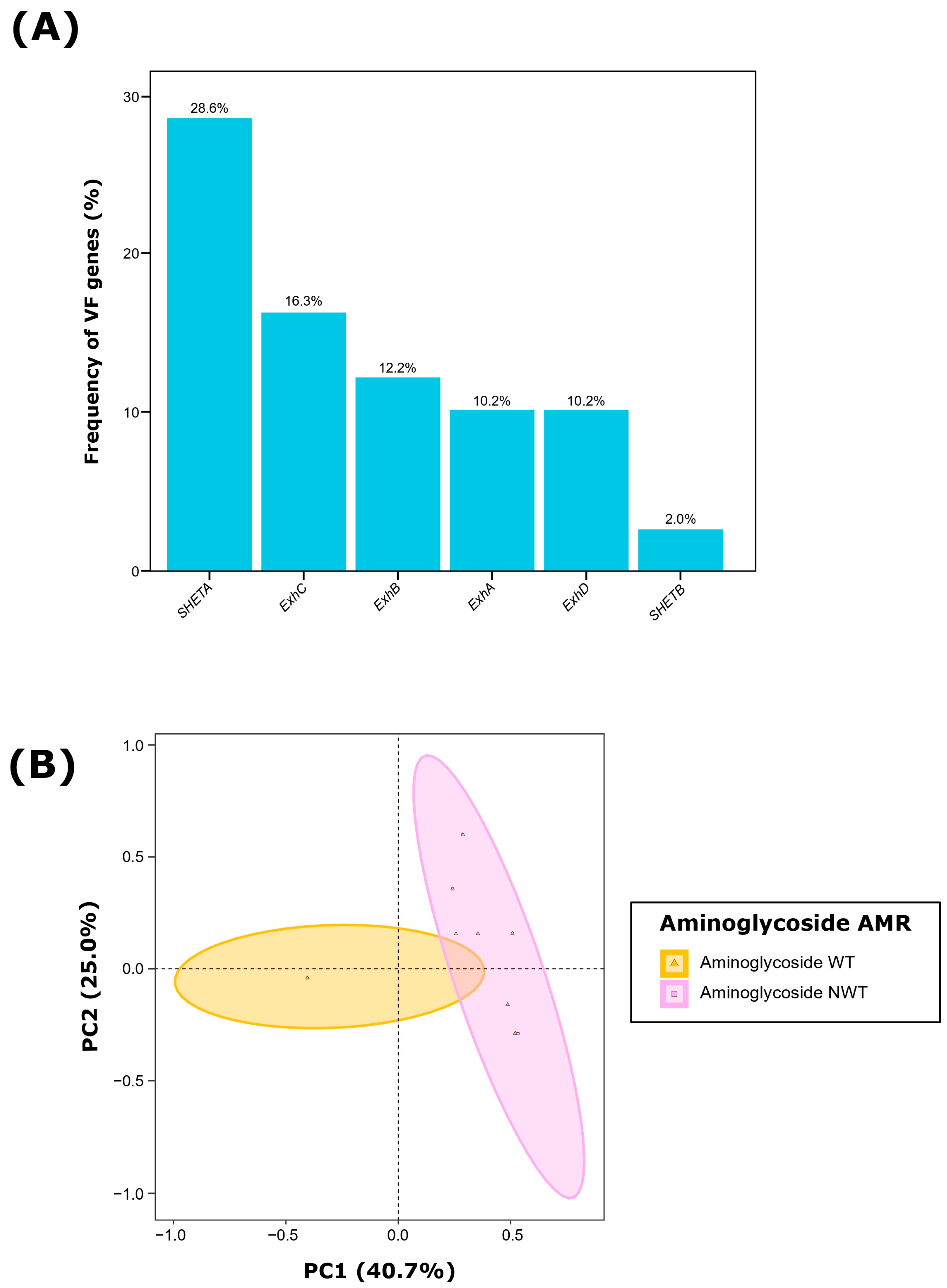
2.2. Characterization of Virulence Factors
2.3. Antimicrobial Susceptibility Profiling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial | Nº of Isolates with MIC (µg/mL) | MIC | WT | NWT | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | MIC50 | MIC90 | n | % | n | % | |
| c Penicillin | 12 | 3 | 6 | 2 | 2 | 1 | 2 | 23 | 4 | >8 | 12 | 24.5 | 37 | 75.5 | |||||
| b Ampicillin | 17 | 7 | 7 | 6 | 8 | 3 | 0 | 1 | 1 | 4 | 17 | 39.7 | 32 | 30.3 | |||||
| c Ceftiofur | 2 | 11 | 31 | 3 | 1 | 1 | 1 | 2 | 47 | 95.9 | 2 | 4.1 | |||||||
| b Gentamicin | 38 | 1 | 1 | 2 | 3 | 4 | ≤1 | 16 | 39 | 79.6 | 10 | 20.4 | |||||||
| c Spectinomycin | 0 | 0 | 5 | 9 | 35 | >64 | >64 | 14 | 28.6 | - | - | ||||||||
| b Neomycin | 42 | 6 | 0 | 1 | ≤4 | 8 | - | - | 7 | 14.3 | |||||||||
| b Tulathromycin | 1 | 8 | 10 | 2 | 1 | 0 | 0 | 27 | >64 | >64 | 21 | 42.9 | 28 | 51.1 | |||||
| b Tilmicosin | 20 | 0 | 0 | 1 | 0 | 28 | ≤4 | ≤4 | 20 | 40.8 | 29 | 59.2 | |||||||
| b Tylosin | 2 | 10 | 8 | 0 | 0 | 0 | 1 | 28 | >32 | >32 | 20 | 40.8 | 29 | 59.2 | |||||
| b Chlortetracycline | 6 | 7 | 3 | 0 | 1 | 32 | >8 | >8 | 13 | 26.5 | 36 | 73.5 | |||||||
| b Oxytetracycline | 12 | 4 | 0 | 0 | 0 | 33 | >8 | >8 | 16 | 32.7 | 33 | 67.3 | |||||||
| d Danofloxacin | 7 | 23 | 2 | 3 | 14 | 0.25 | >1 | - | - | - | - | ||||||||
| b Enrofloxacin | 19 | 10 | 2 | 3 | 2 | 13 | 0.25 | >2 | 29 | 59.2 | 20 | 40.8 | |||||||
| c Clindamycin | 8 | 0 | 2 | 0 | 2 | 3 | 0 | 36 | >16 | >16 | 8 | 16.3 | 41 | 83.7 | |||||
| a SXT | 47 | 2 | ≤2/38 | ≤2/38 | 47 | 95.9 | 2 | 4.1 | |||||||||||
| d Sulfadimethoxine | 43 | 6 | ≤256 | >256 | - | - | - | - | |||||||||||
| b Tiamulin | 1 | 7 | 1 | 1 | 0 | 1 | 1 | 37 | >32 | >32 | 9 | 18.4 | 40 | 81.6 | |||||
| b Florfenicol | 0 | 0 | 0 | 8 | 17 | 15 | 9 | 8 | >8 | 25 | 51 | 24 | 49 | ||||||

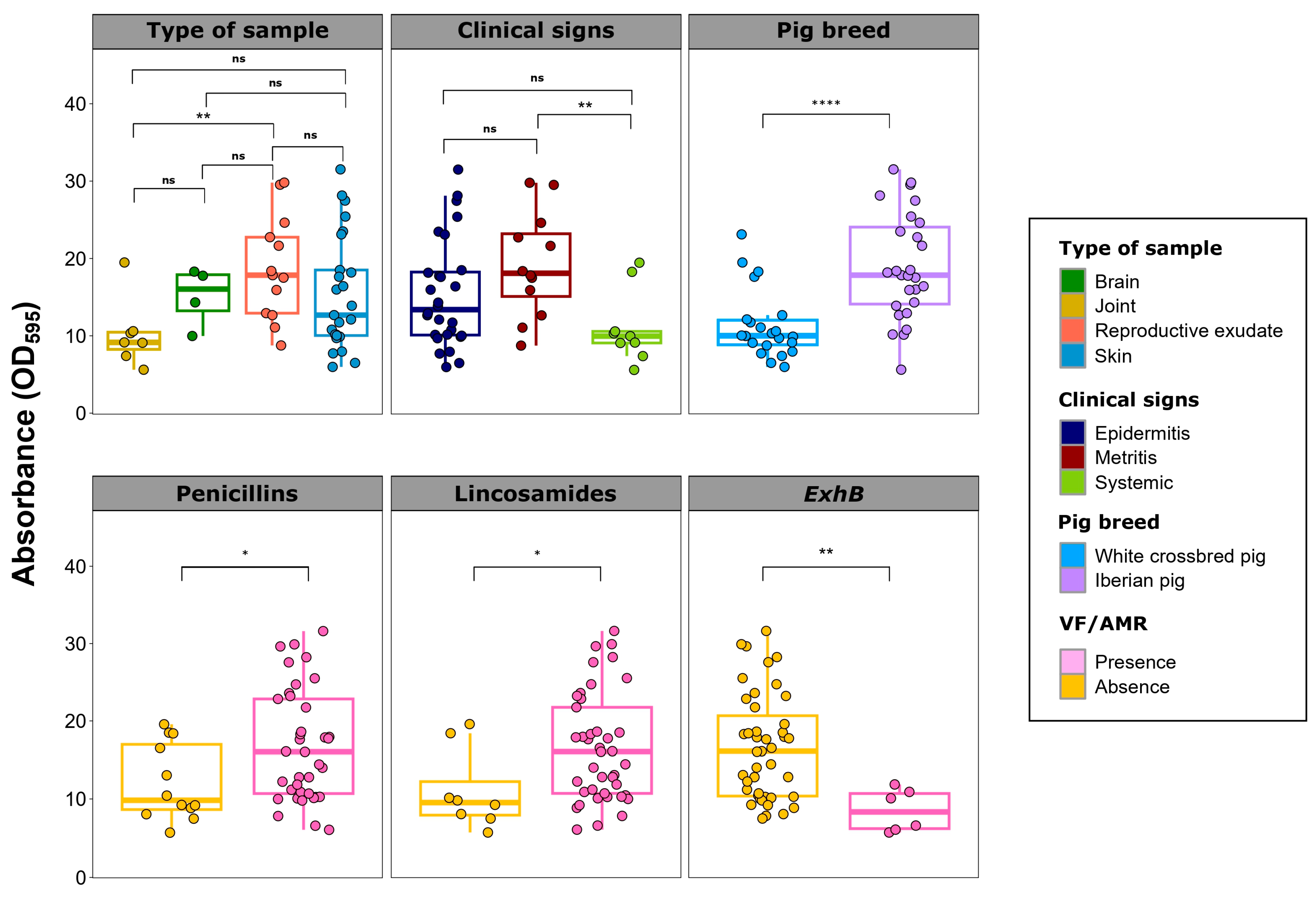
2.4. Cooccurrence among Virulence Factors and Antimicrobial Susceptibility Profiling
2.5. Biofilm Formation
3. Discussion
4. Materials and Methods
4.1. Sampling and Bacterial Isolation and Characterization
4.2. Molecular Characterization of Virulence Factors
4.3. Antimicrobial Susceptibility Testing
4.4. Biofilm Formation Assay
4.5. Data Analysis and Results Visualization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casanova, C.; Iselin, L.; Von Steiger, N.; Droz, S.; Sendi, P. Staphylococcus hyicus Bacteremia in a Farmer. J. Clin. Microbiol. 2011, 49, 4377–4378. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Friendship, R.M.; Poljak, Z.; Weese, J.S.; Dewey, C.E. An Investigation of Exudative Epidermitis (Greasy Pig Disease) and Antimicrobial Resistance Patterns of Staphylococcus hyicus and Staphylococcus aureus Isolated from Clinical Cases. Can. Vet. J. 2013, 54, 139. [Google Scholar] [PubMed]
- Wegener, H.C.; Skov-Jensen, E.W. Exudative epidermitis. In Diseases of Swine, 9th ed.; Straw, B.E., Zimmerman, J.J., D’Allaire, S., Taylor, D.J., Eds.; Blackwell Pub: Ames, IA, USA, 2006; pp. 675–679. [Google Scholar]
- de Winter, P.J.J.; Verdonck, M.; de Kruif, A.; Devriese, L.A.; Haesebrouck, F. Bacterial Endometritis and Vaginal Discharge in the Sow: Prevalence of Different Bacterial Species and Experimental Reproduction of the Syndrome. Anim. Reprod. Sci. 1995, 37, 325–335. [Google Scholar] [CrossRef]
- Hill, B.D.; Corney, B.G.; Wagner, T.M. Importance of Staphylococcus hyicus as a Cause of Arthritis in Pigs up to 12 Weeks of Age. Aust. Vet. J. 1996, 73, 179–181. [Google Scholar] [CrossRef]
- Tanabe, T.; Sato, H.; Sato, H.; Watanabe, K.; Hirano, M.; Hirose, K.; Kurokawa, S.; Nakano, K.; Saito, H.; Maehara, N. Correlation between Occurrence of Exudative Epidermitis and Exfoliative Toxin-Producing Ability of Staphylococcus hyicus. Vet. Microbiol. 1996, 48, 9–17. [Google Scholar] [CrossRef]
- Leekitcharoenphon, P.; Pamp, S.J.; Andresen, L.O.; Aarestrup, F.M. Comparative Genomics of Toxigenic and Non-Toxigenic Staphylococcus hyicus. Vet. Microbiol. 2016, 185, 34–40. [Google Scholar] [CrossRef]
- Andresen, L.O.; Ahrens, P. A Multiplex PCR for Detection of Genes Encoding Exfoliative Toxins from Staphylococcus hyicus. J. Appl. Microbiol. 2004, 96, 1265–1270. [Google Scholar] [CrossRef]
- Sato, H.; Watanabe, T.; Higuchi, K.; Teruya, K.; Ohtake, A.; Murata, Y.; Saito, H.; Aizawa, C.; Danbara, H.; Maehara, N. Chromosomal and Extrachromosomal Synthesis of Exfoliative Toxin from Staphylococcus hyicus. J. Bacteriol. 2000, 182, 4096. [Google Scholar] [CrossRef]
- Watanabe, T.; Sato, H.; Hatakeyama, Y.; Matsuzawa, T.; Kawai, M.; Aizawa, C.; Danbara, H.; Maehara, N. Cloning of the Gene Coding for Staphylococcus hyicus Exfoliative Toxin B and Its Expression in Escherichia coli. J. Bacteriol. 2000, 182, 4101. [Google Scholar] [CrossRef]
- Argemi, X.; Hansmann, Y.; Prola, K.; Prévost, G. Coagulase-Negative Staphylococci Pathogenomics. Int. J. Mol. Sci. 2019, 20, 1215. [Google Scholar] [CrossRef]
- Silva, V.; Correia, E.; Pereira, J.E.; González-Machado, C.; Capita, R.; Alonso-Calleja, C.; Igrejas, G.; Poeta, P. Exploring the Biofilm Formation Capacity in S. pseudintermedius and Coagulase-Negative Staphylococci Species. Pathogens 2022, 11, 689. [Google Scholar] [CrossRef] [PubMed]
- Regulation—2019/6—EN—EUR-Lex. Available online: https://eur-lex.europa.eu/eli/reg/2019/6/oj (accessed on 20 July 2024).
- Moreno, A.M.; Moreno, L.Z.; Poor, A.P.; Matajira, C.E.C.; Moreno, M.; Gomes, V.T.d.M.; da Silva, G.F.R.; Takeuti, K.L.; Barcellos, D.E. Antimicrobial Resistance Profile of Staphylococcus hyicus Strains Isolated from Brazilian Swine Herds. Antibiotics 2022, 11, 205. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Hu, J.; Zhu, L.; Guo, C.; Lu, H.; Guo, C.; Li, X.; Wang, X. A Fatal Suppurative Pneumonia in Piglets Caused by a Pathogenic Coagulase-Positive Strain of Staphylococcus hyicus. Vet. Res. Commun. 2017, 41, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Foissac, M.; Lekaditi, M.; Loutfi, B.; Ehrhart, A.; Dauchy, F.A. Spondylodiscitis and Bacteremia Due to Staphylococcus hyicus in an Immunocompetent Man. Germs 2016, 6, 106. [Google Scholar] [CrossRef] [PubMed]
- Kirk, F.; Mashicharan, M.; Braddick, M.; Saxena, P. Staphylococcus hyicus, a Novel Pathogen Causing Destructive Infective Endocarditis Requiring Mitral Annular Reconstruction. JTCVS Tech. 2022, 13, 70. [Google Scholar] [CrossRef]
- Andresen, L.O. Production of Exfoliative Toxin by Isolates of Staphylococcus hyicus from Different Countries. Vet. Rec. 2005, 157, 376–378. [Google Scholar] [CrossRef]
- Kanbar, T.; Voytenko, A.V.; Alber, J.; Lämmler, C.; Weiss, R.; Skvortzov, V.N. Distribution of the Putative Virulence Factor Encoding Gene Sheta in Staphylococcus hyicus Strains of Various Origins. J. Vet. Sci. 2008, 9, 327. [Google Scholar] [CrossRef]
- Futagawa-Saito, K.; Ba-Thein, W.; Higuchi, T.; Sakurai, N.; Fukuyasu, T. Nationwide Molecular Surveillance of Exfoliative Toxigenic Staphylococcus hyicus on Pig Farms across Japan. Vet. Microbiol. 2007, 124, 370–374. [Google Scholar] [CrossRef]
- Onuma, K.; Uoya, Y.; Koide, T.; Shibata, A.; Tanabe, T.; Sato, H. Detection of Staphylococcus hyicus Exfoliative Toxin Genes by Dot Blot Hybridization and Multiplex Polymerase Chain Reaction. Microbiol. Immunol. 2011, 55, 168–173. [Google Scholar] [CrossRef]
- Fudaba, Y.; Nishifuji, K.; Andresen, L.O.; Yamaguchi, T.; Komatsuzawa, H.; Amagai, M.; Sugai, M. Staphylococcus hyicus Exfoliative Toxins Selectively Digest Porcine Desmoglein 1. Microb. Pathog. 2005, 39, 171–176. [Google Scholar] [CrossRef]
- Király, J.; Hajdučková, V.; Gregová, G.; Szabóová, T.; Pilipčinec, E. Resistant S. aureus Isolates Capable of Producing Biofilm from the Milk of Dairy Cows with Subclinical Mastitis in Slovakia. Agriculture 2024, 14, 571. [Google Scholar] [CrossRef]
- Araújo, D.; Silva, A.R.; Fernandes, R.; Serra, P.; Barros, M.M.; Campos, A.M.; Oliveira, R.; Silva, S.; Almeida, C.; Castro, J. Emerging Approaches for Mitigating Biofilm-Formation-Associated Infections in Farm, Wild, and Companion Animals. Pathogens 2024, 13, 320. [Google Scholar] [CrossRef]
- Dastghey, S.; Parvizi, J.; Shapiro, I.M.; Hickok, N.J.; Otto, M. Effect of Biofilms on Recalcitrance of Staphylococcal Joint Infection to Antibiotic Treatment. J. Infect. Dis. 2015, 211, 641. [Google Scholar] [CrossRef] [PubMed]
- Hagstrand Aldman, M.; Thompson, O.; Påhlman, L.I. Biofilm Formation Is Associated with Poor Outcome in Prosthetic Joint Infections Caused by Staphylococcus lugdunensis. Infect. Dis. 2023, 55, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Miguelez-Perez, R.; Mencia-Ares, O.; Gutierrez-Martin, C.B.; Gonzalez-Fernandez, A.; Petrocchi-Rilo, M.; Delgado-Garcia, M.; Martinez-Martinez, S. Biofilm Formation in Streptococcus suis: In Vitro Impact of Serovar and Assessment of Coinfections with Other Porcine Respiratory Disease Complex Bacterial Pathogens. bioRxiv 2024. [Google Scholar] [CrossRef]
- Silva, V.; Correia, E.; Pereira, J.E.; González-Machado, C.; Capita, R.; Alonso-Calleja, C.; Igrejas, G.; Poeta, P. Biofilm Formation of Staphylococcus aureus from Pets, Livestock, and Wild Animals: Relationship with Clonal Lineages and Antimicrobial Resistance. Antibiotics 2022, 11, 772. [Google Scholar] [CrossRef]
- Arsenakis, I.; Boyen, F.; Haesebrouck, F.; Maes, D.G.D. Autogenous Vaccination Reduces Antimicrobial Usage and Mortality Rates in a Herd Facing Severe Exudative Epidermitis Outbreaks in Weaned Pigs. Vet. Rec. 2018, 182, 744. [Google Scholar] [CrossRef]
- Futagawa-Saito, K.; Ba-Thein, W.; Fukuyasu, T. Antimicrobial Susceptibilities of Exfoliative Toxigenic and Non-Toxigenic Staphylococcus hyicus Strains in Japan. J. Vet. Med. Sci. 2009, 71, 681–684. [Google Scholar] [CrossRef]
- European Medicines Agency (EMA). Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2022—Trends from 2010 to 2022—Thirteenth ESVAC Report. 2023. Available online: https://op.europa.eu/en/publication-detail/-/publication/59eda847-b429-11ee-b164-01aa75ed71a1/language-en (accessed on 23 July 2024).
- Larsson, D.G.J.; Flach, C.F. Antibiotic Resistance in the Environment. Nat. Rev. Microbiol. 2021, 20, 257–269. [Google Scholar] [CrossRef]
- Murray, L.M.; Hayes, A.; Snape, J.; Kasprzyk-Hordern, B.; Gaze, W.H.; Murray, A.K. Co-Selection for Antibiotic Resistance by Environmental Contaminants. NPJ Antimicrob. Resist. 2024, 2, 9. [Google Scholar] [CrossRef]
- Mencía-Ares, O.; Borowiak, M.; Argüello, H.; Cobo-Díaz, J.F.; Malorny, B.; Álvarez-Ordóñez, A.; Carvajal, A.; Deneke, C. Genomic Insights into the Mobilome and Resistome of Sentinel Microorganisms Originating from Farms of Two Different Swine Production Systems. Microbiol. Spectr. 2022, 10, e02896-22. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, P.; Andresen, L.O. Cloning and Sequence Analysis of Genes Encoding Staphylococcus hyicus Exfoliative Toxin Types A, B, C, and D. J. Bacteriol. 2004, 186, 1833–1837. [Google Scholar] [CrossRef] [PubMed]
- Howden, B.P.; Giulieri, S.G.; Wong Fok Lung, T.; Baines, S.L.; Sharkey, L.K.; Lee, J.Y.H.; Hachani, A.; Monk, I.R.; Stinear, T.P. Staphylococcus aureus Host Interactions and Adaptation. Nat. Rev. Microbiol. 2023, 21, 380–395. [Google Scholar] [CrossRef] [PubMed]
- EUCAST The European Committee on Antimicrobial Susceptibility Testing—EUCAST. Available online: http://www.eucast.org/ (accessed on 15 May 2024).
- EUCAST MIC and Inhibition Zone Diameter Distributions of Microorganisms without and with Phenotypically Evident Resistance Mechanisms. Available online: https://mic.eucast.org/ (accessed on 17 May 2024).
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Croes, S.; Deurenberg, R.H.; Boumans, M.L.L.; Beisser, P.S.; Neef, C.; Stobberingh, E.E. Staphylococcus aureus Biofilm Formation at the Physiologic Glucose Concentration Depends on the S. aureus Lineage. BMC Microbiol. 2009, 9, 229. [Google Scholar] [CrossRef]
- Dong, C.L.; Che, R.X.; Wu, T.; Qu, Q.W.; Chen, M.; Zheng, S.D.; Cai, X.H.; Wang, G.; Li, Y.H. New Characterization of Multi-Drug Resistance of Streptococcus suis and Biofilm Formation from Swine in Heilongjiang Province of China. Antibiotics 2023, 12, 132. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Wickham, H. Ggplot2. 2016. Available online: https://link.springer.com/book/10.1007/978-3-319-24277-4 (accessed on 17 June 2024).
- Network Analysis and Visualization [R Package Igraph Version 2.0.3]. 2024. Available online: https://cran.r-project.org/web/packages/igraph/index.html (accessed on 17 June 2024).
- Pedersen, T.L. An Implementation of Grammar of Graphics for Graphs and Networks [R Package Ggraph Version 2.2.1]. 2024. Available online: https://cran.r-project.org/web/packages/ggraph/index.html (accessed on 17 June 2024).
- CRAN: Package Pheatmap. Available online: https://cran.r-project.org/web/packages/pheatmap/index.html (accessed on 24 July 2024).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]

| Frequence (n) | Percentage (%) | ||
|---|---|---|---|
| Geographic origin | Castile and León | 9 | 18.4 |
| Catalonia | 10 | 20.4 | |
| Extremadura | 11 | 22.4 | |
| Galicia | 6 | 12.2 | |
| Region of Murcia | 13 | 26.5 | |
| Type of pig | Iberian pig | 27 | 55.1 |
| White crossbred pigs | 22 | 44.9 | |
| Production phase | Suckling | 12 | 24.5 |
| Weaning | 24 | 49.0 | |
| Sow/Boar | 13 | 26.5 | |
| Type of sample | Brain | 7 | 14.3 |
| Joint | 4 | 8.2 | |
| Reproductive exudate | 13 | 26.5 | |
| Skin | 25 | 51.0 | |
| Clinical signs | Epidermitis | 28 | 57.1 |
| Metritis | 12 | 24.5 | |
| Systemic | 9 | 18.4 |
| Pairwise Association | Φ Coefficient | Φ Categorization | p-Value |
|---|---|---|---|
| Aminoglycosides—ExhC | 0.45 | Very strong | 0.001 |
| Phenicols—ExhC | 0.40 | Very strong | 0.002 |
| Aminoglycosides—SHETA | 0.32 | Very strong | 0.023 |
| Macrolides—ExhC | 0.31 | Very strong | 0.015 |
| Pleuromutilins—ExhD | 0.28 | Very strong | 0.037 |
| SHETA—ExhC | 0.39 | Very strong | 0.004 |
| Macrolides—Phenicols | 0.61 | Very strong | <0.001 |
| Lincosamides—Pleuromutilins | 0.57 | Very strong | <0.001 |
| Macrolides—Tetracyclines | 0.49 | Very strong | 0.001 |
| Lincosamides—Macrolides | 0.48 | Very strong | <0.001 |
| Penicillins—Tetracyclines | 0.46 | Very strong | 0.001 |
| Lincosamides—Penicillins | 0.45 | Very strong | 0.001 |
| Lincosamides—Phenicols | 0.38 | Very strong | 0.004 |
| Macrolides—Penicillins | 0.35 | Very strong | 0.008 |
| Macrolides—Aminoglycosides | 0.33 | Very strong | 0.016 |
| Penicillins—Phenicols | 0.32 | Very strong | 0.018 |
| Lincosamides—Tetracyclines | 0.30 | Very strong | 0.023 |
| Aminoglycosides—Tetracyclines | 0.29 | Very strong | 0.021 |
| Penicillins—Pleuromutilins | 0.28 | Very strong | 0.029 |
| Aminoglycosides—Penicillins | 0.27 | Very strong | 0.024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mencía-Ares, O.; Ramos-Calvo, E.; González-Fernández, A.; Aguarón-Turrientes, Á.; Pastor-Calonge, A.I.; Miguélez-Pérez, R.; Gutiérrez-Martín, C.B.; Martínez-Martínez, S. Insights into the Virulence and Antimicrobial Resistance of Staphylococcus hyicus Isolates from Spanish Swine Farms. Antibiotics 2024, 13, 871. https://doi.org/10.3390/antibiotics13090871
Mencía-Ares O, Ramos-Calvo E, González-Fernández A, Aguarón-Turrientes Á, Pastor-Calonge AI, Miguélez-Pérez R, Gutiérrez-Martín CB, Martínez-Martínez S. Insights into the Virulence and Antimicrobial Resistance of Staphylococcus hyicus Isolates from Spanish Swine Farms. Antibiotics. 2024; 13(9):871. https://doi.org/10.3390/antibiotics13090871
Chicago/Turabian StyleMencía-Ares, Oscar, Eva Ramos-Calvo, Alba González-Fernández, Álvaro Aguarón-Turrientes, Ana Isabel Pastor-Calonge, Rubén Miguélez-Pérez, César B. Gutiérrez-Martín, and Sonia Martínez-Martínez. 2024. "Insights into the Virulence and Antimicrobial Resistance of Staphylococcus hyicus Isolates from Spanish Swine Farms" Antibiotics 13, no. 9: 871. https://doi.org/10.3390/antibiotics13090871
APA StyleMencía-Ares, O., Ramos-Calvo, E., González-Fernández, A., Aguarón-Turrientes, Á., Pastor-Calonge, A. I., Miguélez-Pérez, R., Gutiérrez-Martín, C. B., & Martínez-Martínez, S. (2024). Insights into the Virulence and Antimicrobial Resistance of Staphylococcus hyicus Isolates from Spanish Swine Farms. Antibiotics, 13(9), 871. https://doi.org/10.3390/antibiotics13090871