
The Puzzle of Coccoid Forms of Helicobacter pylori: Beyond Basic Science

 ,
,  ,
,
Abstract
:1. Introduction
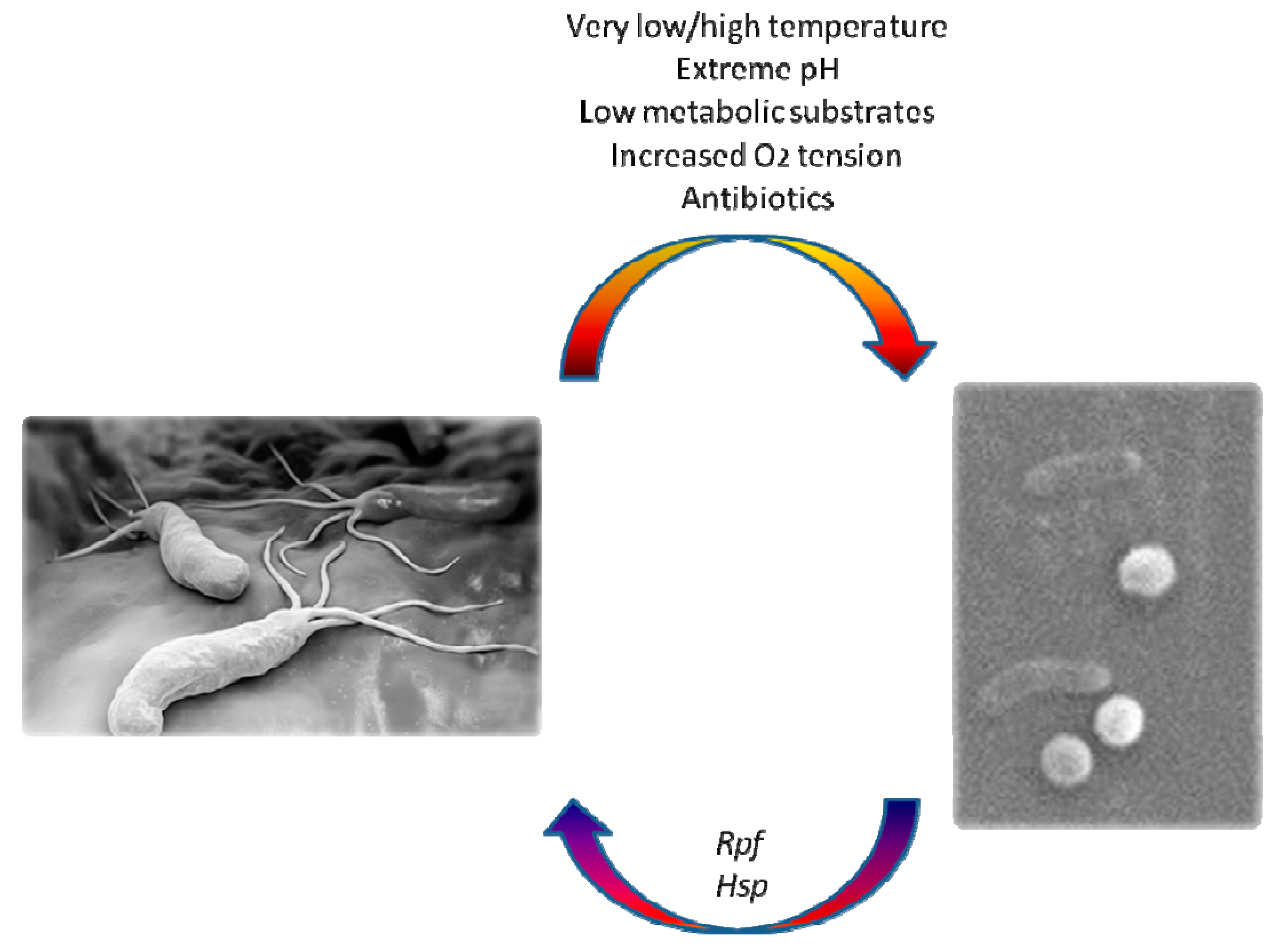
2. Coccoid Forms of H. pylori
3. Biology and Immunology
4. Diagnostic Potential
5. Clinical Relevance
- “Relapse”: the bacterium responsible for the recurrence is genetically the same as that identified before the eradication and, relapse usually occurs within the first year following the eradication;
- “Re-infection”: the bacterium causing the recurrence is different from that identified before the eradication and develops a long time after the first eradication.
6. Possible Estimation of Problem Dimension: Preliminary Experience
7. Therapeutic Prospective Options
- Increase of acid inhibition: this option may be carried out in three different ways:
- -
- High-dose dual therapy consisting of the combination of amoxicillin (e.g, 1 g tid or 750 mg qid) and a proton pump inhibitor (standard-dose tid or qid or double standard-dose bid) for 14 days. This approach is based on previous studies showing that this regimen has achieved a satisfactory therapeutic outcome as empirical first-line or rescue therapy for H. pylori infection [91].
- -
- Increase of proton pump inhibitor (PPI) dose. This assumption fits with the fact that the reproduction cycle of H. pylori is dependent on gastric pH, since it enters the replicative cycle at neutral pH below the gastric mucus layer and turns into coccoid forms at acidic pH. In this setting, not only the dose, but also the choice of PPI might have relevance. In fact, it is presumable that second-generation PPIs (esomeprazole, rabeprazole) are more effective than first-generation PPIs (omeprazole, lansoprazole). The explanation may be that second-generation drugs have a metabolism less dependent on CYP2C19 allelic variants and, therefore, active metabolite serum levels are more stable over time [92].
- -
- Use of Vonoprazan (VPZ). VPZ is a new, powerful acid-inhibitory drug. It is a competitive inhibitor of H+/K+ ATPase pump located on the apical membrane of gastric parietal cells. VPZ became clinically available in Japan in 2015. Compared to conventional PPIs, gastric acid was inhibited by VPZ more rapidly, more strongly and for a longer duration. Moreover, VPZ does not require pharmacological activation and has a protracted half-life. A study reported that VPZ (20 mg bid) potently suppressed acid for 24 h [93]. The Tokyo Helicobacter pylori Study Group showed that the H. pylori eradication rate in the third-line treatment was higher in VPZ-based therapy than in PPI-based therapy [93,94]. Moreover, it has been shown that VPZ could increase the eradication rate of triple therapy even in clarithromycin-resistant strains [94]; therefore, it is presumable that VPZ could help the transition of H. pylori from coccoid to spiral shape. However, VPZ is currently available only on the Japanese market, and other confirmatory studies are necessary to assess its potential in Western countries [95].
- Free fatty acids. A very recent study has shown that free fatty acids, such as linolenic acid and liposomal linolenic acid, have a bactericidal effect “in vitro” on both bacillary and coccoid forms, regardless of the resistance to antibiotics [96]. Linolenic acid may be effective because it enhances membrane permeability, therefore facilitates cell lysis and antibiotic access into the cells [97].
- Mucolytic substances. Similar to what happens for other bacteria, such as Pseudomonas aeruginosa, Staphylococcus epidermidis and E. coli, the production of biofilm by H. pylori is known to facilitate antibiotic resistance. Biofilm is a complex composed by bacteria and extracellular matrix of polyanionic polysaccharides. H. pylori coccoid forms may be hidden within biofilm. Some studies demonstrated, in this context, the beneficial role of N-acetylcysteine for its mucolytic and also bacteriostatic properties. On these bases, N-acetylcysteine could be used as a pre-treatment followed by antibiotic therapy [98,99,100].
8. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Ierardi, E.; Goni, E.; Losurdo, G.; Di Mario, F. Helicobacter pylori and nonmalignant diseases. Helicobacter 2014, 19 (Suppl. 1), 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasbarrini, G.; Genta, R.M.; Anti, M.; Fox, J.; Caselli, M.; Doglioni, C.; Ierardi, E.; Masala, G.; Palli, D.; Testoni, P.A.; et al. Update on Helicobacter pylori research. Malignancies. Eur. J. Gastroenterol. Hepatol. 1997, 9, 621–623. [Google Scholar] [CrossRef] [PubMed]
- Zullo, A.; Hassan, C.; Cristofari, F.; Andriani, A.; De Francesco, V.; Ierardi, E.; Tomao, S.; Stolte, M.; Morini, S.; Vaira, D. Effects of Helicobacter pylori eradication on early stage gastric mucosa-associated lymphoid tissue lymphoma. Clin. Gastroenterol. Hepatol. 2010, 8, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Zagari, R.M.; Romano, M.; Ojetti, V.; Stockbrugger, R.; Gullini, S.; Annibale, B.; Farinati, F.; Ierardi, E.; Maconi, G.; Rugge, M.; et al. Guidelines for the management of Helicobacter pylori infection in Italy: The III Working Group Consensus Report 2015. Dig. Liver Dis. 2015, 47, 903–912. [Google Scholar] [CrossRef]
- Ierardi, E.; Giorgio, F.; Losurdo, G.; Di Leo, A.; Principi, M. How antibiotic resistances could change Helicobacter pylori treatment: A matter of geography? World J. Gastroenterol. 2013, 19, 8168–8180. [Google Scholar] [CrossRef]
- Zagari, R.M.; Romiti, A.; Ierardi, E.; Gravina, A.G.; Panarese, A.; Grande, G.; Savarino, E.; Maconi, G.; Stasi, E.; Euseby, L.H.; et al. The “three-in-one” formulation of bismuth quadruple therapy for Helicobacter pylori eradication with or without probiotics supplementation: Efficacy and safety in daily clinical practice. Helicobacter 2018, 23, e12502. [Google Scholar] [CrossRef]
- Ierardi, E.; Giorgio, F.; Iannone, A.; Losurdo, G.; Principi, M.; Barone, M.; Pisani, A.; Di Leo, A. Noninvasive molecular analysis of Helicobacter pylori: Is it time for tailored first-line therapy? World J. Gastroenterol. 2017, 23, 2453–2458. [Google Scholar] [CrossRef]
- Cabrera, C.E.; Gòmez, R.F.; Zuniga, A.D. La resistencia de bacterias a antibióticos, antisépticos y desinfectantes una manifestación de los mecanismos de supervivencia y adaptación. Colomb. Med. 2007, 38, 149–158. [Google Scholar]
- Li, L.; Mendis, N.; Trigui, H.; Oliver, J.D.; Faucher, S.P. The importance of the viable but non-culturable state in human bacterial pathogens. Front. Microbiol. 2014, 5, 258. [Google Scholar] [CrossRef] [Green Version]
- Citterio, B.; Casaroli, A.; Pierfelici, L.; Battistelli, M.; Falcieri, E.; Baffone, W. Morphological changes and outer membrane protein patterns in Helicobacter pylori during conversion from bacillary to coccoid form. New Microbiol. 2004, 27, 353–360. [Google Scholar]
- Kusters, J.G.; Gerrits, M.M.; Van Strijp, J.A.; Vandenbroucke-Grauls, C.M. Coccoid forms of Helicobacter pylori are the morphologic manifestation of cell death. Infect. Immun. 1997, 65, 3672–3679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilius, M.; Strohle, A.; Bode, G.; Malfertheiner, P. Coccoid like forms (CLF) of Helicobacter pylori. Enzyme activity and antigenicity. Zentralbl. Bakteriol. 1993, 280, 259–272. [Google Scholar] [CrossRef]
- Hirukawa, S.; Sagara, H.; Kaneto, S.; Kondo, T.; Kiga, K.; Sanada, T.; Kiyono, H.; Mimuro, H. Characterization of morphological conversion of Helicobacter pylori under anaerobic conditions. Microbiol. Immunol. 2018, 62, 221–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Percival, S.L.; Suleman, L. Biofilms and Helicobacter pylori: Dissemination and persistence within the environment and host. World J. Gastrointest. Pathophysiol. 2014, 5, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Sato, F. Helicobacter pylori in culture: An ultrastructural study. Hokkaido Igaku Zasshi. 2000, 75, 187–196. [Google Scholar]
- O’Rourke, J.; Bode, G. Morphology and ultrastructure. In Helicobacter Pylori: Physiology and Genetics; Mobley, H.L.T., Mendz, G.L., Hazell, S.L., Eds.; ASM Press: Washington, DC, USA, 2001; Chapter 6. [Google Scholar]
- Kaprelyants, A.S.; Gottschal, J.C.; Kell, D.B. Dormancy in non-sporulating bacteria. FEMS Microbiol. Rev. 1993, 10, 271–285. [Google Scholar] [CrossRef]
- Anuchin, A.M.; Mulyukin, A.L.; Suzina, N.E.; Duda, V.I.; El-Registan, G.I.; Kaprelyants, A.S. Dormant forms of Mycobacterium smegmatis with distinct morphology. Microbiology 2009, 155, 1071–1079. [Google Scholar] [CrossRef] [Green Version]
- Kudykina, Y.K.; Shleeva, M.O.; Artsabanov, V.Y.; Suzina, N.E.; Kaprel’iants, A.S. Generation of dormant forms by Mycobacterium smegmatis in the poststationary phase during gradual acidification of the medium. Mikrobiologiia 2011, 80, 625–636. [Google Scholar] [CrossRef]
- Dubois, A. Intracellular Helicobacter pylori and gastric carcinogenesis: An “old” frontier worth revisiting. Gastroenterology 2007, 132, 1177–1180. [Google Scholar] [CrossRef] [Green Version]
- Can, F.; Karahan, C.; Dolapei, I.; Demirbilek, M.; Tekeli, A.; Arslan, H. Urease activity and urea gene sequencing of coccoid forms of H. pylori induced by different factors. Curr. Microbiol. 2008, 56, 150–155. [Google Scholar] [CrossRef]
- Hua, J.; Ho, B. Is the coccoid form of Helicobacter pylori viable? Microbios 1996, 87, 103–112. [Google Scholar] [PubMed]
- Catrenich, C.E.; Makin, K.M. Characterization of the morphologic conversion of Helicobacter pylori from bacillary to coccoid forms. Scand. J. Gastroenterol. Suppl. 1991, 181, 54–64. [Google Scholar]
- Moshkowitz, M.; Gorea, A.; Arber, N.; Konikoff, F.; Berger, S.; Gilat, T. Morphological transformation of Helicobacter pylori during prolonged incubation: Association with decreased acid resistance. J. Clin. Pathol. 1994, 47, 172–174. [Google Scholar] [CrossRef] [PubMed]
- Benaissa, M.; Babin, P.; Quellard, N.; Pezennec, L.; Cenatiempo, Y.; Fauchère, J.L. Changes in Helicobacter pylori ultrastructure and antigens during conversion from the bacillary to the coccoid form. Infect. Immun. 1996, 64, 2331–2335. [Google Scholar] [CrossRef] [Green Version]
- Faghri, J.; Poursina, F.; Moghim, S.; Zarkesh Esfahani, H.; Nasr Esfahani, B.; Fazeli, H.; Mirzaei, N.; Jamshidian, A.; Ghasemian Safaei, H. Morphological and Bactericidal Effects of Different Antibiotics on Helicobacter pylori. Jundishapur J. Microbiol. 2014, 7, e8704. [Google Scholar] [CrossRef] [Green Version]
- Catrenich, C.E.; Chestnut, M.H. Character and origin of vacuoles induced in mammalian cells by the cytotoxin of Helicobacter pylori. J. Med. Microbiol. 1992, 37, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Sörberg, M.; Nilsson, M.; Hanberger, H.; Nilsson, L.E. Morphologic conversion of Helicobacter pylori from bacillary to coccoid form. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 216–219. [Google Scholar] [CrossRef]
- Mouery, K.; Rader, B.A.; Gaynor, E.C.; Guillemin, K. The stringent response is required for Helicobacter pylori survival of stationary phase, exposure to acid, and aerobic shock. J. Bacteriol. 2006, 188, 5494–5500. [Google Scholar] [CrossRef] [Green Version]
- Khomeriki, S.G.; Morozov, I.A. The role of coccoid forms of Helicobacter pylori in pathogenetic mechanisms and persistence of Helicobacter infection. Russ. J. Gastroenterol. Hepatol. Coloproctol. 2001, 13, 99–102. [Google Scholar]
- Mukamolova, G.V.; Turapov, O.A.; Kazarian, K.; Telkov, M.; Kaprelyants, A.S.; Kell, D.B.; Young, M. The Rpf gene of Micrococcus luteus encodes an essential secreted growth factor. Mol. Microbiol. 2002, 46, 611–621. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, N.S. Chronic inactive gastritis and coccoid Helicobacter pylori in patients treated for gastroesophageal reflux disease or with H pylori eradication therapy. Am. J. Clin. Pathol. 2002, 118, 719–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kansau, I.; Labigne, A. Heat shock proteins of Helicobacter pylori. Aliment. Pharmacol. Ther. 1996, 1, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Noach, L.A.; Rolf, T.M.; Tytgat, G.N. Electron microscopic study of association between Helicobacter pylori and gastric and duodenal mucosa. J. Clin. Pathol. 1994, 47, 699–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, G.; Malfertheiner, P.; Ditschuneit, H. Pathogenetic implications of ultrastructural findings in Campylobacter pylori related gastroduodenal disease. Scand. J. Gastroenterol. 1988, 23, 25–39. [Google Scholar] [CrossRef]
- Tominaga, K.; Hamasaki, N.; Watanabe, T.; Uchida, T.; Fujiwara, Y.; Takaishi, O. Effect of culture conditions on morphological changes of Helicobacter pylori. J. Gastroenterol. 1999, 34, 28–31. [Google Scholar]
- Park, A.M.; Li, Q.; Nagata, K.; Tamura, T.; Shimono, K.; Sato, E.F.; Inoue, M. Oxygen tension regulates reactive oxygen generation and mutation of Helicobacter pylori. Free Radic. Biol. Med. 2004, 36, 1126–1133. [Google Scholar] [CrossRef]
- Nilsson, H.O.; Blom, J.; Abu-Al-Soud, W.; Ljungh, A.A.; Andersen, L.P.; Wadström, T. Effect of cold starvation, acid stress, and nutrients on metabolic activity of Helicobacter pylori. Appl. Environ. Microbiol. 2002, 68, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Berry, V.; Jennings, K.; Woodnutt, G. Bactericidal and morphological effects of amoxicillin on Helicobacter pylori. Antimicrob. Agents Chemother. 1995, 39, 1859–1861. [Google Scholar] [CrossRef] [Green Version]
- Kadkhodaei, S.; Siavoshi, F.; Akbari Noghabi, K. Mucoid and coccoid Helicobacter pylori with fast growth and antibiotic resistance. Helicobacter 2020, 25, e12678. [Google Scholar] [CrossRef]
- Sisto, F.; Brenciaglia, M.I.; Scaltrito, M.M.; Dubini, F. Helicobacter pylori: ureA, cagA and vacA expression during conversion to the coccoid form. Int. J. Antimicrob. Agents 2000, 15, 277–282. [Google Scholar] [CrossRef]
- Amieva, M.R.; Salama, N.R.; Tompkins, L.S.; Falkow, S. Helicobacter pylori enter and survive within multivesicular vacuoles of epithelial cells. Cell Microbiol. 2002, 4, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Ozbek, A.; Ozbek, E.; Dursun, H.; Kalkan, Y.; Demirci, T. Can Helicobacter pylori invade human gastric mucosa? An in vivo study using electron microscopy, immunohistochemical methods, and real-time polymerase chain reaction. J. Clin. Gastroenterol. 2010, 44, 416–422. [Google Scholar]
- Chu, Y.T.; Wang, Y.H.; Wu, J.J.; Lei, H.Y. Invasion and multiplication of Helicobacter pylori in gastric epithelial cells and implications for antibiotic resistance. Infect. Immun. 2010, 78, 4157–4165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, I.; Utt, M.; Nilsson, H.O.; Ljungh, A.; Wadström, T. Two-dimensional electrophoretic and immunoblot analysis of cell surface proteins of spiral-shaped and coccoid forms of Helicobacter pylori. Electrophoresis 2000, 21, 2670–2677. [Google Scholar] [CrossRef]
- Roe, I.H.; Son, S.H.; Oh, H.T.; Choi, J.; Shin, J.H.; Lee, J.H.; Hah, Y.C. Changes in the evolution of the antigenic profiles and morphology during coccoid conversion of Helicobacter pylori. Korean J. Intern. Med. 1999, 14, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, G.; Faùndez, G.; Troncozso, M.; Navarrete, P.; Toledo, M.S. Immunoglobulin G antibody response to infection with coccoid forms of Helicobacter pylori. Clin. Diagn. Lab. Immunol. 2002, 9, 1067–1071. [Google Scholar] [CrossRef] [Green Version]
- Ng, B.L.; Quak, S.H.; Aw, M.; Goh, K.T.; Ho, B. Immune responses to differentiated forms of Helicobacter pylori in children with epigastric pain. Clin. Diagn. Lab. Immunol. 2003, 10, 866–869. [Google Scholar] [CrossRef] [Green Version]
- Cellini, L.; Allocati, N.; Angelucci, D.; Lezzi, T.; Di Campli, E.; Marzio, L.; Dainelli, B. Coccoid Helicobacter pylori not culturable in vitro reverts in mice. Microbiol. Immunol. 1994, 38, 843–850. [Google Scholar] [CrossRef]
- She, F.F.; Su, D.H.; Lin, J.Y.; Zhou, L.Y. Virulence and potential pathogenicity of coccoid helicobacter pylori induced by antibiotics. World J. Gastroenterol. 2001, 7, 254–258. [Google Scholar] [CrossRef]
- Rabelo-Goncalves, E.M.; Nishimura, N.F.; Zeitune, J.M. Acute inflammatory response in the stomach of BALB/c mice challenged with coccoidal Helicobacter pylori. Mem. Inst. Oswaldo Cruz. 2002, 97, 1201–1206. [Google Scholar] [CrossRef] [Green Version]
- Sarem, M.; Macìas, M.; Amendola, R.; Bori, J.; Corti, R. Transmisión de la bacteria Helicobacter pylori a partir del agua. Acta Gastroenterol. Latinoam. 2011, 41, 17. [Google Scholar]
- Andersen, L.P.; Rasmussen, L. Helicobacter pylori-coccoid forms and biofilm formation. FEMS Immunol. Med. Microbiol. 2009, 56, 112–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizoguchi, H.; Fujioka, T.; Nasu, M. Evidence for viability of coccoid forms of Helicobacter pylori. J. Gastroenterol. 1999, 34 (Suppl. 11), 32–36. [Google Scholar]
- Bode, G.; Mauch, F.; Malfertheiner, P. The coccoid forms of Helicobacter pylori. Criteria for their viability. Epidemiol. Infect. 1993, 111, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saniee, P.; Shahreza, S.; Siavoshi, F. Negative Effect of Proton-pump Inhibitors (PPIs) on Helicobacter pylori Growth, Morphology, and Urease Test and Recovery after PPI Removal—An In vitro Study. Helicobacter 2016, 21, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Shahamat, M.; Mai, U.; Paszko-Kolva, C.; Kessel, M.; Colwell, R.R. Use of autoradiography to assess viability of Helicobacter pylori in water. Appl. Environ. Microbiol. 1993, 59, 1231–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willén, R.; Carlén, B.; Wang, X.; Papadogiannakis, N.; Odselius, R.; Wadström, T. Morphologic conversion of Helicobacter pylori from spiral to coccoid form. Scanning (SEM) and transmission electron microscopy (TEM) suggest viability. Upsala J. Med. Sci. 2000, 105, 31–40. [Google Scholar] [CrossRef]
- Moreno, Y.; Ferrús, M.A.; Alonso, J.L.; Jiménez, A.; Hernández, J. Use of fluorescent in situ hybridization to evidence the presence of Helicobacter pylori in water. Water Res. 2003, 37, 2251–2256. [Google Scholar] [CrossRef] [Green Version]
- Angelidis, A.S.; Tirodimos, I.; Bobos, M.; Kalamaki, M.S.; Papageorgiou, D.K.; Arvanitidou, M. Detection of Helicobacter pylori in raw bovine milk by fluorescence in situ hybridization (FISH). Int. J. Food Microbiol. 2011, 151, 252–256. [Google Scholar] [CrossRef]
- Kabir, S. Detection of Helicobacter pylori in faeces by culture, PCR and enzyme immunoassay. J. Med. Microbiol. 2001, 50, 1021–1029. [Google Scholar] [CrossRef]
- Bahrami, A.R.; Rahimi, E.; Ghasemian Safaei, H. Detection of Helicobacter pylori in city water, dental units’ water, and bottled mineral water in Isfahan, Iran. Sci. World J. 2013, 2013, 280510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.J.; Perng, C.L.; Shyu, R.Y.; Chen, C.H.; Lou, Q.; Chong, S.K.; Lee, C.H. Comparison of five PCR methods for detection of Helicobacter pylori DNA in gastric tissues. J. Clin. Microbiol. 1999, 37, 772–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunn, J.E.; MacKay, W.G.; Thomas, J.E.; Reid, D.C.; Weaver, L.T. Detection of Helicobacter pylori DNA in drinking water biofilms: Implications for transmission in early life. Lett. Appl. Microbiol. 2002, 34, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Janzon, A.; Sjöling, A.; Lothigius, A.; Ahmed, D.; Qadri, F.; Svennerholm, A.M. Failure to detect Helicobacter pylori DNA in drinking and environmental water in Dhaka, Bangladesh, using highly sensitive real-time PCR assays. Appl. Environ. Microbiol. 2009, 75, 3039–3044. [Google Scholar] [CrossRef] [Green Version]
- Sen, K.; Schable, N.A.; Lye, D.J. Development of an internal control for evaluation and standardization of a quantitative PCR assay for detection of Helicobacter pylori in drinking water. Appl. Environ. Microbiol. 2007, 73, 7380–7387. [Google Scholar] [CrossRef] [Green Version]
- McDaniels, A.E.; Wymer, L.; Rankin, C.; Haugland, R. Evaluation of quantitative real time PCR for the measurement of Helicobacter pylori at low concentrations in drinking water. Water Res. 2005, 39, 4808–4816. [Google Scholar] [CrossRef]
- Minami, M.; Ohta, M.; Ohkura, T.; Ando, T.; Torii, K.; Hasegawa, T.; Goto, H. Use of a combination of brushing technique and the loop-mediated isothermal amplification method as a novel, rapid, and safe system for detection of Helicobacter pylori. J. Clin. Microbiol. 2006, 44, 4032–4037. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y.; Notomi, T. Loop-mediated isothermal amplification (LAMP): A rapid, accurate, and cost-effective diagnostic method for infectious diseases. J. Infect. Chemother. 2009, 15, 62–69. [Google Scholar] [CrossRef]
- Chamanrokh, P.; Shahhosseiny, M.H.; Mazaheri Assadi, M.; Nejadsattari, T.; Esmaili, D. A comparison of loop-mediated isothermal amplification (LAMP) With PCR and rapid urease test(RUT) to detect Helicobacter pylori in biopsy samples. J. Pure Appl. Microbiol. 2014, 8, 1079–1086. [Google Scholar]
- Khan, M.M.; Pyle, B.H.; Camper, A.K. Specific and rapid enumeration of viable but nonculturable and viable-culturable gram-negative bacteria by using flow cytometry. Appl. Environ. Microbiol. 2010, 76, 5088–5096. [Google Scholar] [CrossRef] [Green Version]
- Handelsman, J. Metagenomics: Application of genomics to uncultured microorganisms. Microbiol. Mol. Biol. Rev. 2004, 68, 669–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.; Andersson, A.F.; Ye, W.; Nyrén, O.; Normark, S.; Engstrand, L. A method for metagenomics of Helicobacter pylori from archived formalin-fixed gastric biopsies permitting longitudinal studies of carcinogenic risk. PLoS ONE 2011, 6, e26442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Mesonero, L.; Hortelano, I.; Moreno, Y.; Ferrús, M.A. Evidence of viable Helicobacter pylori and other bacteria of public health interest associated with free-living amoebae in lettuce samples by next generation sequencing and other molecular techniques. Int. J. Food Microbiol. 2019, 318, 108477. [Google Scholar] [CrossRef] [PubMed]
- Monstein, H.J.; Jonasson, J. Differential virulence-gene mRNA expression in coccoid forms of Helicobacter pylori. Biochem. Biophys. Res. Commun. 2001, 285, 530–536. [Google Scholar] [CrossRef]
- Wang, X.F.; Wang, K.X. Cloning and expression of vacA gene fragment of Helicobacter pylori with coccoid form. J. Chin. Med. Assoc. 2004, 67, 549–556. [Google Scholar]
- Wen, M.; Zhang, Y.; Yamada, N.; Matsuhisa, T.; Matsukura, N.; Sugisaki, Y. An evaluative system for the response of antibacterial therapy: Based on the morphological change of Helicobacter pylori and mucosal inflammation. Pathol. Int. 1999, 49, 332–337. [Google Scholar] [CrossRef]
- Raymond, J.; Thiberge, J.M.; Dauga, C. Diagnosis of Helicobacter pylori recurrence: Relapse or reinfection? Usefulness of molecular tools. Scand. J. Gastroenterol. 2016, 51, 672–678. [Google Scholar] [CrossRef]
- Ierardi, E.; Francavilla, R.; Balzano, T.; Negrini, R.; Francavilla, A. Autoantibodies reacting with gastric antigens in Helicobacter pylori associated body gastritis of dyspeptic children. Ital. J. Gastroenterol. Hepatol. 1998, 30, 478–480. [Google Scholar]
- Negrini, R.; Lisato, L.; Zanella, I.; Cavazzini, L.; Gullini, S.; Villanacci, V.; Poiesi, C.; Albertini, A.; Ghielmi, S. Helicobacter pylori infection induces antibodies cross-reacting with human gastric mucosa. Gastroenterology 1991, 101, 437–445. [Google Scholar] [CrossRef]
- Van der Ende, A.; van der Hulst, R.W.; Dankert, J.; Tytgat, G.N. Reinfection versus recrudescence in Helicobacter pylori infection. Aliment. Pharmacol. Ther. 1997, 11, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.X.; Talley, N.J.; Keane, C.T.; O’Morain, C.A. Recurrence of Helicobacter pylori infection after successful eradication: Nature and possible causes. Dig. Dis. Sci. 1997, 42, 1821–1834. [Google Scholar] [CrossRef] [PubMed]
- Figura, N.; Moretti, E.; Vaglio, L.; Vernillo, R.; Vindigni, C.; Giordano, N. Factors modulating the outcome of treatment for the eradication of Helicobacter pylori infection. New Microbiol. 2012, 35, 335–340. [Google Scholar] [PubMed]
- De Francesco, V.; Zullo, A.; Giorgio, F.; Saracino, I.; Zaccaro, C.; Hassan, C.; Ierardi, E.; Di Leo, A.; Fiorini, G.; Castelli, V.; et al. Change of point mutations in Helicobacter pylori rRNA associated with clarithromycin resistance in Italy. J. Med. Microbiol. 2014, 63Pt 3, 453–457. [Google Scholar] [CrossRef]
- Giorgio, F.; Ierardi, E.; Sorrentino, C.; Principi, M.; Barone, M.; Losurdo, G.; Iannone, A.; Giangaspero, A.; Monno, R.; di Leo, A. Helicobacter pylori DNA isolation in the stool: An essential pre-requisite for bacterial noninvasive molecular analysis. Scand. J. Gastroenterol. 2016, 51, 1429–1432. [Google Scholar] [CrossRef] [PubMed]
- Ierardi, E.; Giorgio, F.; Losurdo, G.; Sorrentino, C.; Principi, M.; Di Leo, A. Detection of Helicobacter pylori DNA sequences in gastric biopsy samples to refine the diagnosis and therapy. J. Med. Microbiol. 2015, 64, 788–789. [Google Scholar] [CrossRef] [Green Version]
- Iannone, A.; Giorgio, F.; Russo, F.; Riezzo, G.; Girardi, B.; Pricci, M.; Palmer, S.C.; Barone, M.; Principi, M.; Strippoli, G.F.; et al. New fecal test for non-invasive Helicobacter pylori detection: A diagnostic accuracy study. World J. Gastroenterol. 2018, 24, 3021–3029. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Megraud, F.; O’Morain, C.A.; Gisbert, J.P.; Kuipers, E.J.; Axon, A.T.; Bazzoli, F.; Gasbarrini, A.; Atherton, J.; Graham, D.Y.; et al. Management of Helicobacter pylori infection-the Maastricht V/Florence Consensus Report. Gut 2017, 66, 6–30. [Google Scholar] [CrossRef] [Green Version]
- Sarem, M.; Corti, R. Role of helicobacter pylori coccoid forms in infection and recrudescence. Gastroenterologìa Hepatologìa. 2016, 39, 28–35. [Google Scholar] [CrossRef]
- Russo, F.; Berloco, P.; Cuomo, R.; Caruso, M.L.; Di Matteo, G.; Giorgio, P.; De Francesco, V.; Di Leo, A.; Ierardi, E. Helicobacter pylori strains and histologically-related lesions affect the outcome of triple eradication therapy: A study from southern Italy. Aliment. Pharmacol. Ther. 2003, 17, 421–428. [Google Scholar] [CrossRef]
- Gao, C.P.; Zhou, Z.; Wang, J.Z.; Han, S.X.; Li, L.P.; Lu, H. Efficacy and safety of high-dose dual therapy for Helicobacter pylori rescue therapy: A systematic review and meta-analysis. J. Dig. Dis. 2016, 17, 811–819. [Google Scholar] [CrossRef]
- Ierardi, E.; Losurdo, G.; La Fortezza, R.F.; Principi, M.; Barone, M.; Di Leo, A. Optimizing proton pump inhibitors in Helicobacter pylori treatment: Old and new tricks to improve effectiveness. World J. Gastroenterol. 2019, 25, 5097–5104. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Yamaoka, Y. Role of Vonoprazan in Helicobacter pylori Eradication Therapy in Japan. Front. Pharmacol. 2019, 15, 9–1560. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Oshima, T.; Horikawa, T.; Tozawa, K.; Tomita, T.; Fukui, H.; Watari, J.; Miwa, H. Systematic review with meta-analysis: Vonoprazan, a potent acid blocker, is superior to proton-pump inhibitors for eradication of clarithromycin-resistant strains of Helicobacter pylori. Helicobacter 2018, 23, e12495. [Google Scholar] [CrossRef] [PubMed]
- Abadi, A.T.B.; Ierardi, E. Vonoprazan and Helicobacter pylori Treatment: A Lesson From Japan or a Limited Geographic Phenomenon? Front. Pharmacol. 2019, 10, 316. [Google Scholar] [CrossRef] [Green Version]
- Obonyo, M.; Zhang, L.; Thamphiwatana, S.; Pornpattananangkul, D.; Fu, V.; Zhang, L. Antibacterial activities of liposomal linolenic acids against antibiotic-resistant Helicobacter pylori. Mol. Pharm. 2012, 9, 2677–2685. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.W.; Thamphiwatana, S.; Zhang, L.; Obonyo, M. Mechanism of antibacterial activity of liposomal linolenic acid against Helicobacter pylori. PLoS ONE 2015, 10, e0116519. [Google Scholar] [CrossRef] [Green Version]
- Makipour, K.; Friedenberg, F.K. The potential Role of N-Acetylcysteine for the treatment of Helicobacter pylori. J. Clin. Gastroenterol. 2011, 45, 841–853. [Google Scholar] [CrossRef] [Green Version]
- Cammarota, G.; Branca, G.; Ardito, F.; Sanguinetti, M.; Ianiro, G.; Cianci, R.; Torelli, R.; Masala, G.; Gasbarrini, A.; Fadda, G.; et al. Biofilm demolition and antibiotic treatment to eradicate resistant Helicobacter pylori: A clinical trial. Clin. Gastroenterol. Hepatol. 2010, 8, 817–820. [Google Scholar] [CrossRef]
- Ianiro, G.; Cammarota, G.; Gasbarrini, A.; Gasbarrini, G. Role of N-acetyl-cisteine in the treatment of H. pylori. Minerva Gastroenterol. Dietol. 2014, 60, 101. [Google Scholar]


{kind=link}
| No. | % | |
|---|---|---|
| Total patients | 185 | 100.0 |
| Positivity of at least two conventional tests and DNA | 179 | 96.7 |
| Negative conventional tests with positive DNA (coccoid?) | 6 | 3.3 |
| Overall clarithromycin resistance | 45 | 24.3 |
| Clarithromycin resistance in negative conventional tests/positive DNA | 2 | 1.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ierardi, E.; Losurdo, G.; Mileti, A.; Paolillo, R.; Giorgio, F.; Principi, M.; Di Leo, A. The Puzzle of Coccoid Forms of Helicobacter pylori: Beyond Basic Science. Antibiotics 2020, 9, 293. https://doi.org/10.3390/antibiotics9060293
Ierardi E, Losurdo G, Mileti A, Paolillo R, Giorgio F, Principi M, Di Leo A. The Puzzle of Coccoid Forms of Helicobacter pylori: Beyond Basic Science. Antibiotics. 2020; 9(6):293. https://doi.org/10.3390/antibiotics9060293
Chicago/Turabian StyleIerardi, Enzo, Giuseppe Losurdo, Alessia Mileti, Rosa Paolillo, Floriana Giorgio, Mariabeatrice Principi, and Alfredo Di Leo. 2020. "The Puzzle of Coccoid Forms of Helicobacter pylori: Beyond Basic Science" Antibiotics 9, no. 6: 293. https://doi.org/10.3390/antibiotics9060293
APA StyleIerardi, E., Losurdo, G., Mileti, A., Paolillo, R., Giorgio, F., Principi, M., & Di Leo, A. (2020). The Puzzle of Coccoid Forms of Helicobacter pylori: Beyond Basic Science. Antibiotics, 9(6), 293. https://doi.org/10.3390/antibiotics9060293