Simple Summary
The lycopsid Sublepidodendron is cosmopolitan in the Late Devonian and Early Carboniferous. Sublepidodendron grabaui is known from several localities of the Upper Devonian Wutong Formation in South China, with its overall morphology and male reproductive organs (i.e., the microsporangiate strobili and microspores) studied previously. Here we describe new specimens of S. grabaui from Guangde City, Anhui Province, to get further knowledge on this plant, especially its female reproductive organs (i.e., megasporangiate strobili and megaspores). The megasporangiate strobili are borne terminally on fertile axes and occasionally dichotomized, bearing at least eight Lagenicula-type megaspores with a small gula in each sporangium. Based on the previous and present study, this plant is considered as a tree lycopsid exhibiting multiple dichotomized stems, occasionally produced lateral branches and monosporangiate strobili.
Abstract
South China displays Devonian strata with well-exposed outcrops and is regarded as a diversity hotspot of Late Devonian lycopsids. The heterosporous lycopsid Sublepidodendron grabaui has been studied for over ten years, with its general morphology, aerial stem anatomy, microsporangiate strobili, and growth architecture reported. Based on new specimens from Guangde City, Anhui Province, this study provides further knowledge about the megasporangiate strobili and megaspores of S. grabaui. Its slender megasporangiate strobili occur singly or in pairs and occasionally bifurcate in the middle-upper portion. Each megasporophyll consists of a flattened pedicel and an adaxially curved lamina. The lamina forms a downturned heel at the base. Each sessile megasporangium contains at least eight Lagenicula-type megaspores with a small gula. The other observed characteristics of S. grabaui in this study conform to those previously known and are compared to relative coeval taxa.
1. Introduction
The lycopsids dominated the Paleozoic wetland landscape and possessed a long evolutionary history, evidenced by the earliest fossil traced back to the late Silurian [1,2,3]. Middle Devonian lycopsids evolved many key characteristics, e.g., the heterospory [4,5] and the tree habit [6,7]. Heterosporous lycopsids blossomed in the Late Devonian (e.g., [8,9,10,11,12,13,14,15,16]), which is regarded as a major step in the full colonization of land surfaces by plants [17,18]. In the past 20 years, over ten species of Late Devonian heterosporous lycopsids have been reported or restudied all around the world (e.g., [9,10,11,12,13,14,15,16,19,20,21,22]); especially, the South China Plate held eight species with monosporangiate strobili (e.g., [9,10,11,12,13,14,15,16]), suggesting a probable origin center and diversity hotspot of this group.
Sublepidodendron grabaui is known as the combination of the former morphological species S. wusihense and Lepidostrobus grabaui and is described as an arborescent, heterosporous lycopsid from several localities of the Upper Devonian Wutong Formation in South China [12,23]. The previous studies revealed the general morphology, aerial stem anatomy, microsporangiate strobili, and growth architecture of S. grabaui. In this article, its megaporangiate strobili and megaspores are investigated based on new materials excavated from the Upper Devonian of Guangde City, Anhui Province, China.
2. Materials and Methods
The specimens were collected from the Yongchuan mine, Xinhang town, Guangde City, Anhui Province, China (Figure 1). The fossil plant in this study is restricted to the bottom portion of the Leigutai Member, i.e., the upper member of the Upper Devonian (Famennian) Wutong Formation (Figure 2), and is preserved as compressions and impressions in yellow, brown, or grayish mudstones. Steel needles were used to expose the morphology of some dichotomized axes and strobili. The limonitized stele shows the anatomy in the transverse section, which was examined under a stereomicroscope. The specimens were photographed using a Nikon Z6 digital camera and a Leica S9 stereomicroscope. All figures were organized using Adobe Photoshop CC 2018 (ver. 19.0, San Jose, CA, USA) and Corel Draw 2018 (ver. 20.1, Ottawa, ON, Canada). All specimens are housed at the Department of Geology, Peking University, Beijing, China.
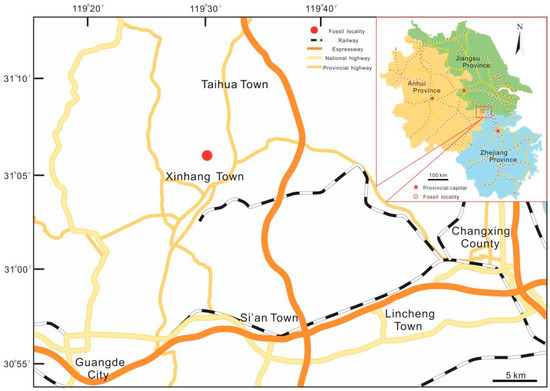
Figure 1.
Map showing the fossil locality of Sublepidodendron grabaui in this study.
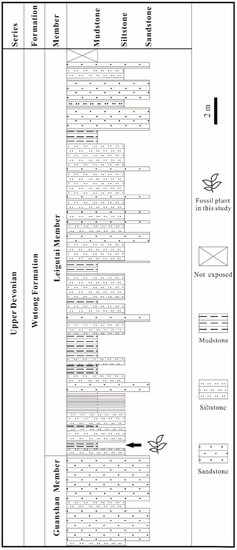
Figure 2.
Stratigraphic column of the Yongchuan Section in Xinhang Town, Guangde City, Anhui Province, China, showing the fossil-bearing bed at the bottom part of Leigutai Member, Wutong Formation.
Systematic Palaeobotany
Class: Lycopsida Pichi-Sermolli 1958.
Order: Isoëtales sensu lato Meyen 1987.
Suborder: Dichostrobiles DiMichele and Bateman 1996.
Family: Sublepidodendraceae Kräusel and Weyland 1949.
Genus: Sublepidodendron (Nathorst) Hirmer 1927.
Type species: Sublepidodendron mirabile (Nathorst) Hirmer 1927.
Sublepidodendron grabaui (Sze) Wang and Xu 2005 [23] emend. Meng et al., 2016 [12] emend.
Neotype: PB 2505 (first designated by Wang and Xu in Bot. J. Linn. Soc., 2005 [23], and deposited at NIGP-CAS).
Epitype: PB 19,255 (first designated by Wang and Xu in Bot. J. Linn. Soc., 2005 [23], and deposited at NIGP-CAS).
Specimens examined here: PKUB22000–22002, 22003a, 22003b, 22004–22009, 22010a, 22010b, 22011–22015 (Deposited at Department of Geology, Peking University, Beijing, China).
Emended specific diagnosis: (emended and additional generic characters are in brackets) Arborescent and heterosporous lycopsid plant with a trunk, branches, and monosporangiate strobili. Trunk, 32–100 mm wide. Branches, 1.5–24 mm wide terminate trunk, dichotomized at least four times or pseudomonopodially branched. Leaves, simple, linear, [inserted at 30–90° on the axis], 0.4–[2.0] mm wide at the proximal portion, 18–60 mm long, with a single midvein. Leaf bases, elongate fusiform in outline, 0.3–1.3 mm at the widest point, 1.6–20 mm long, with an obtusely rounded ridge at the widest point, and one vascular bundle and keel. Each branch terminated by a single cigar-shaped strobilus, [6.0–11] mm wide (excluding the distal part of sporophylls), up to 160 mm in total length. The stalk of sporophyll 0.2 mm wide and 3.0–4.0 mm long. Sporophyll laminae with smooth margins, rhomboid in outline, with pointed apex. Sporangia adaxial, elongate elliptical in outline, 0.8–[1.3] × [2.7]–4.0 mm, without obvious pedicels. [Megasporangiate strobili single, paired, or occasionally once-dichotomized. Lagenicula type] megaspores with circular amb, [a gula and spiny ornamentation, 661]–1200 μm in diameter. [Gula 289–440 μm high.] Cingulate Lycospora-type microspores, 18–35 μm in diameter, proximal surface with finely microgranulate ornamentation and distal surface with densely microspinate ornamentation. Trunk stele with a pith, exarch primary xylem, and secondary xylem; stele of large branches with a small pith, exarch primary xylem; a solid exarch primary xylem strand present in small branches.
3. Results
The specimens of Sublepidodendron grabaui described here include vegetative organs of stems and leaves (Figure 3 and Figure 4), fertile structures (Figure 5 and Figure 6), and anatomy of the axis (Figure 7). Stems are up to 55 cm long and 5.5–33.5 mm wide (Figure 3A–G), displaying at least three dichotomies (Figure 3A–D). Each dichotomy produces two daughter axes of half-width, and the angles between the daughter axes are 30–60° (Figure 3A–F). 15.0–33.5 mm wide stems display elongated fusiform leaf bases (Figure 3G–L) that are up to 11.8 mm long and ca. 1.0 mm wide. All these leaf bases have a similar length/width ratio of ca. 12:1. Linear ornamentations occur on the interspaces between adjacent leaf bases (Figure 3G–J). Each leaf base displays a median fissure-like false leaf scar (i.e., the vertical “keel”), which occasionally shows an oval vascular bundle on the top (Figure 3K–M). Distal slender axes bear rhomboidal or fusiform leaf bases closely arranged in helices (Figure 4A–D). They are 3.4–5.5 mm long and 0.8–1.2 mm wide. These leaf bases have a similar length/width ratio (about 6:1), and each of them also bears a false leaf scar (Figure 4D). Vegetative leaves with entire margins are up to 12.4 mm long and inserted at 30–90° on the axis (Figure 4E–G). Each leaf has an obvious single vein extending from base to apex (Figure 4H,I).

Figure 3.
Stems and elongate-fusiform leaf bases of Sublepidodendron grabaui. (A) Stem dichotomizing several times. White arrows 1, 2, and 3 indicating the bifurcate portions. Rectangle indicating the counterpart enlarged in Figure 3B. The black arrow in the lower part indicating a limonitized stele in the stem. Scale bar = 5 cm. PKUB22010a. (B) Enlargement of the counterpart of Figure 3A (rectangle), showing stem dichotomizing two times. The arrow indicating the bifurcate portion. Scale bar = 5 cm. PKUB22010b. (C) Enlargement of Figure 3B (arrow 1), showing axes with a dichotomy. Scale bar = 2 cm. (D) Line illustration of the Sublepidodendron grabaui stems shown in Figure 3A–C. Stem with three times of dichotomy and dense leaves. The dotted lines indicating the missing portion of the stem. Scale bar = 5 cm. (E) A once-dichotomized slender axis. Scale bar = 2 cm. PKUB22008. (F) A once-dichotomized axis. Scale bar = 2 cm. PKUB22011. (G) Two stems with helically arranged leaf bases. The rectangle indicating the portion enlarged in Figure 3H. the arrows indicating the portions enlarged in Figure 3K (upper) and Figure 3L (lower), respectively. Scale bar = 2 cm. PKUB22006. (H) Enlargement of Figure 3G (rectangle), indicating elongate-fusiform leaf bases. Scale bar = 5 mm. (I) Stem with helically arranged leaf bases. The rectangle indicating the portion enlarged in Figure 3J. Scale bar = 1 cm. PKUB22014. (J) Enlargement of Figure 3I (rectangle), indicating elongate-fusiform leaf cushions. Scale bar = 5 mm. (K,L) Enlargement of Figure 3G (upper and lower arrows, respectively), showing vascular bundle scar and keel in leaf base. Scale bar = 2 mm. (M) Line illustration of a leaf base based on Figure 3H–L. Scale bar = 2 mm.
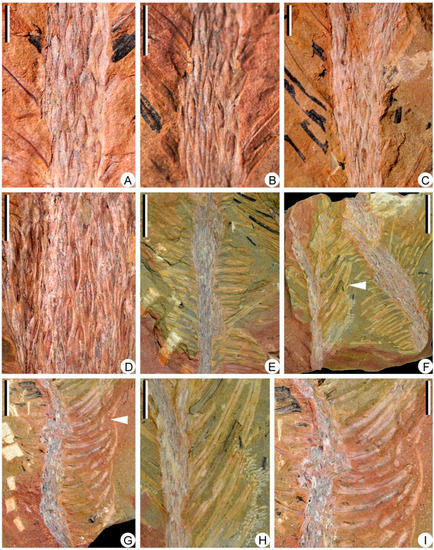
Figure 4.
Leaf bases and vegetative leaves along slender axes of Sublepidodendron grabaui. (A–C) Different stems with helically arranged leaf bases. Scale bar = 5 mm. (D) Helically arranged leaf bases, each showing a vertical groove in the middle. Scale bar = 5 mm. (E) A vegetative axis with persistent dense leaves. Scale bar = 1 cm. (F) Two once-dichotomized vegetative axes with persistent linear leaves. The arrows indicating the portions enlarged in Figure 4H. Scale bar = 2 cm. PKUB22012. (G) A vegetative axis with persistent linear leaves. The arrows indicating the portions enlarged in Figure 4I. Scale bar = 2 cm. PKUB22001. (H,I) Enlargement of arrowed portion in Figure 4F and Figure 4G, respectively, showing vegetative leaves, each with a single midvein. Scale bar = 1 cm.

Figure 5.
Fertile axes and strobili of Sublepidodendron grabaui. (A) Two strobili with slender sporophylls. Scale bar = 2 cm. PKUB22002. (B) A strobilus with basal fertile axis preserved. Scale bar = 2 cm. PKUB22004. (C) A once-dichotomized axis (arrow) bearing two strobili. Scale bar = 2 cm. PKUB22001. (D) A once-dichotomized axis bearing two strobili. Scale bar = 2 cm. PKUB22000. (E,F) Part and counterpart of a bifurcated strobilus. Scale bar = 1 cm. PKUB22003a, 22003b. (G) A once-dichotomized strobilus. The arrow indicating the bifurcate portion. Scale bar = 2 cm. PKUB22007. (H) Three broken slender strobili. Black arrow indicating the gap between two rock layers. Arrows 1, 2, and 3 indicating the three strobili, respectively. The rectangle indicating the portion enlarged in Figure 6A. Scale bar = 2 cm. (I) The counterpart of Figure 5H. Black arrow indicating the gap between two rock layers. Arrows 1, 2, and 3 indicating the three strobili shown in Figure 5H. Scale bar = 2 cm. (J) Three strobili preserved in the same direction. Scale bar = 2 cm. PKUB22015. (K) Five strobili (arrows 1–5) preserved in two different directions (arrows 1–3 and arrows 4 and 5, respectively). Scale bar = 1 cm. PKUB22005.
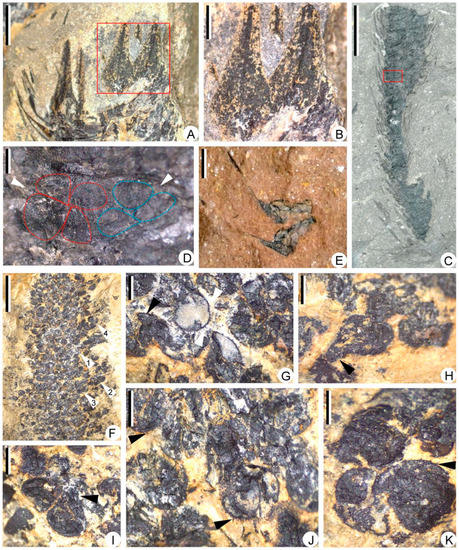
Figure 6.
Megasporophylls and megaspores of Sublepidodendron grabaui. (A) Enlargement of portion in Figure 5H (rectangle), showing sporophylls in helices. Scale bar = 2 mm. (B) Enlargement of portion in Figure 6A (rectangle), showing front view of sporophyll laminae, each with a midvein. Scale bar = 1 mm. (C) A partially preserved strobilus. The rectangle indicating the portion enlarged in Figure 6D. Scale bar = 1 cm. PKUB22009. (D) Enlargement of Figure 6C (rectangle), showing contents of a sporangium. The arrows indicating two possible tetrads. The dotted line shows the outline of megaspores in tetrads. (E) Two isolated megasporophyll–sporangium complexes with lateral view of sporophylls. Scale bar = 2 mm. (F) Large numbers of megaspores. Arrows 1–4 indicating portions enlarged in Figure 6G,I–K, respectively. Scale bar = 5 mm. PKUB22013. (G) Enlargement of Figure 6F (arrow 1), showing megaspores with gula and spiny ornamentations. The black and white arrows indicating gula and spiny ornamentation, respectively. Scale bar = 500 μm. (H) A pear-shaped megaspore. The arrows indicating the position of the gula. Scale bar = 500 μm. (I) Enlargement of Figure 6F (arrow 2), showing two megaspores with gula. The arrow indicating the position of the gula. Scale bar = 500 μm. (J,K) Enlargement of Figure 6F (arrows 3 and 4). The arrow indicating the spiny ornamentations on the megaspore surface. Scale bar = 500 μm.
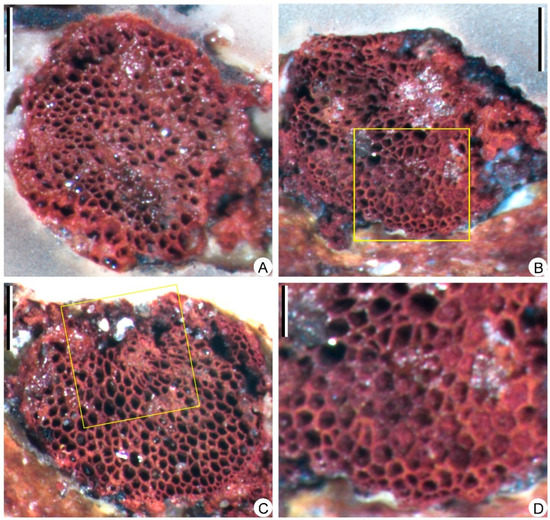
Figure 7.
Transverse sections of the stele, showing the anatomy of Sublepidodendron grabaui. (A) Solid protostele. Scale bar = 1 mm. (B) Partially preserved stele. The rectangle indicating the portions enlarged in Figure 7D. Scale bar = 1 mm. (C) Partially preserved stele. The rectangle indicating the portions where tracheids are torn. Scale bar = 1 mm. (D) Enlargement of Figure 7B (rectangle) displaying the periphery of the stele with protoxylem tracheids. Scale bar = 400 μm.
The strobili are slender and cylindrical, 60–150 mm long and 6–11 mm wide, excluding sporophyll laminae (Figure 5A–J). Some strobili are fully preserved (Figure 5A), while the others are broken at distal or basal ends (Figure 5B–J). Most strobili are singly borne on the fertile axes (Figure 5B–D), while two specimens display paired strobili on dichotomy axes (Figure 5C,D), and two strobili are once-dichotomized (Figure 5E–G). The longest preserved strobilus is ca. 8 mm wide, broken at basal parts (Figure 5H). Three strobili are in the same direction (Figure 5I). Five strobili (arrows 1–5 in Figure 5J) are preserved in two directions (arrows 1–3 and arrows 4 and 5, respectively).
The central strobilar axes are up to 1.5 mm in width, bearing sporophylls in helices. The sporophylls are long-triangular in front view, ca. 2.0 mm at the widest part, showing a single vein in the middle (Figure 6A,B). Each sporophyll consists of a horizontal pedicel and an adaxially curved lamina (Figure 6C,E). All strobili were found to contain megaspores and are thus megasporangiate. Each sporophyll bears one sessile sporangium on the adaxial surface of the pedicel. However, only a few specimens show unbroken sporangial walls to recognize the contour of sporangia (Figure 6C–E). The sporangia are long-ellipsoidal in shape, 2.7–3.3 mm long, and 1.1–1.3 mm high (Figure 6D,E). One specimen shows two adjacent megaspore tetrads that are probably in one sporangium (Figure 6D). Isolated megasporophyll-sporangium complexes are ca. 3.2 mm long, showing exposed megaspores (Figure 6E). The megaspores are Lagenicula-type and 661–943 μm in diameter. The spores are round to pear-shaped in outline (Figure 6G–K). Each megaspore is composed of a spherical body and a distinct gula, with a gula height-to-body diameter ratio of 0.34 (Figure 6H,I). Spiny ornamentations, 18–77 μm long, are clear on some megaspores (Figure 6J,K).
The limonitized stele in Figure 3A (black arrow) is sliced and examined under the microscope. The primary xylem cylinder is exarch, ca. 3.8 mm in diameter (Figure 7A), without ridges of protoxylem. Protoxylem and metaxylem tracheids are circular or polygonal in transverse section, 20–30 μm and 40–120 μm in diameter, respectively (Figure 7B–D). One section displays a partially preserved metaxylem with torn tracheids (Figure 7C).
4. Comparisons
4.1. Comparison among Specimens of Sublepidodendron grabaui in China (Table 1)
Sublepidodendron [9,12,23,24,25] is cosmopolitan in the Late Devonian and Early Carboniferous. We consider our material represents the Sublepidodendron grabaui on the similarities of the following characteristics (Table 1): (1) correlation of horizon and age; (2) repeatedly bifurcated axes; (3) elongate fusiform leaf bases with evident fissure-shaped vertical false leaf scar (the length/width ratio of leaf bases, 6–12:1 in this study vs. 4–20: 1 in [23]); (4) wrinkles or longitudinal ornamentations between adjacent leaf bases; (5) paired slender strobili (60–150 mm × 6–11 mm in this study vs. 160 mm × 8.0 mm in [23]); (6) similar-shaped megasporophylls; (7) anatomy of the stele (solid exarch primary xylem). In this paper, we further provide the bifurcating nature of some S. grabaui megasporangiate strobili and illustrate the Lagenicula megaspores in them.

Table 1.
Comparison among specimens of Sublepidodendron grabaui in China.
Table 1.
Comparison among specimens of Sublepidodendron grabaui in China.
| S. grabaui (Wang and Xu [23]) | S. grabaui (Meng et al. [12]) | Specimens in this Study | |
|---|---|---|---|
| Vegetative axis | |||
| Width (mm) | 1.5–100 | 3.0–43 | 3.0–33.5 |
| Branching | at least four times-dichotomized | isotomously dichotomized or pseudomonopodially branched | at least thrice-dichotomized |
| Anatomy | The trunk: exarch primary xylem with a pith and secondary xylem; the branch: from the primary xylem with a pith to a solid primary xylem strand | A medullated stele or solid exarch primary xylem. | A solid exarch primary xylem. |
| Leaf base | |||
| Shape | elongated fusiform | elongated fusiform | fusiform or elongated fusiform |
| Size (L × W, mm) | (1.6–20) × (0.3–1.2) | (3.0–8.5) × (0.5–1.3) | (3.4–12) × (0.8–1.2) |
| Long-width ratio | (4–20):1 | 7:1 | (6–12):1 |
| Vascular bundle scar | Present | – | Present |
| False leaf scar/“keel” | Present | Present | Present |
| Strobilus | |||
| Shape | Cigar-shaped; singly or occasionally in pairs | Cigar-shaped | Cigar-shaped; singly, in pairs or occasionally once-dichotomized |
| Dimension (L × W, mm) | 160 × 8.0 | 90 × (8.0–10) | (60–150) × (6–11) |
| Megasporangium | Elongate elliptical | Elliptical | Elongate ovoid |
| Megaspore | |||
| Type | – | – | Lagenicula |
| Number per sporangium | – | – | 8 or more |
| Diameter (μm) | 1200 | – | 661–943 (gula/body = 0.34) |
| Ornamentation | – | – | spiny |
| Microspore | |||
| Type | – | Lycospora | – |
| Diameter (μm) | – | 18–35 | – |
| Ornamentation | – | Finely microgranulate on surface | – |
Note: –, lack of information.
4.2. Comparison with Lycopsids Bearing Bisporangiate Strobili (Table 2)
During the Middle and Late Devonian, lycopsids with bisporangiate strobili are widely distributed in the world (e.g., [5,20,21,22,26]). All these taxa display megasporophylls and microsporophylls in the basal and terminal parts of one strobilus (Table 2), respectively, and thus differ from Sublepidodendron grabaui, which possesses megasporangiate strobili.
Table 2.
Comparison among S. grabaui and Devonian bisporangiate lycopsid representatives.
Table 2.
Comparison among S. grabaui and Devonian bisporangiate lycopsid representatives.
| Sublepidodendron grabaui | Yuguangia ordinata | Kossoviella timanica | Cymastrobus irvingii | Clevelandodendron ohioensis | Kowieria alveoformis | |
|---|---|---|---|---|---|---|
| Reference | Wang and Xu [23]; Meng et al. [12]; this study | Hao et al. [5] | Orlova et al. [22] | Evreïnoff et al. [20] | Chitaley and Pigg [26] | Gess and Prestianni [21] |
| Locality | South China | South China | Russia | Australia | America | South Africa |
| Dimension of strobilus | 60–160 mm long, 6–11 mm wide (L/W = 12–19) | up to 160 mm long, 8.6 mm wide (L/W = 18.6) | 50–160 mm long, 2–8 mm wide (L/W = 20–25) | up to 80 mm long, 50 mm wide (L/W = 1.6) | 90 mm long, 60 mm wide (L/W = 1.5) | up to 15 mm long |
| Attachement of terminal strobilus | singly, in pairs or occasionally once-dichotomized | singly or occasionally once-dichotomized | singly or occasionally once-dichotomized | singly | singly | singly |
| Megasporangium | Elongate ovoid | ovoid–elongate-ovoid | ovoid | oblate | oblate | ovoid |
| Megaspore diameter | 661–943 | 450–832 | 450–1180 | 420–490 | 320–360 | 580–720 |
| Megaspore number | 8 or more | 4 or more | 8 | multiple | multiple | Up to 4 |
| Megaspore type | Lagenicula | Triletes | Triletes | – | Triletes | Lagenicula |
Note: –, lack of information.
The Middle Devonian (Givetian) Yuguangia ordinata [5] and the Late Devonian (Frasnian) Kossoviella timanica [22] are bisporangiate lycopsids reported from South China and Northern Russia, respectively. These two plants bear slender and occasionally dichotomized strobili that are morphologically similar to Sublepidodendron grabaui, while they differ in the type of spores and shape of the sporangium.
Cymastrobus irvingii [20] and Clevelandodendron ohioensis [26] bear stubby strobili (length/width = 1.5–1.6) with oblate megasporangium. Each sporangium contains a large number of megaspores. Clevelandodendron ohioensis produces Trileites-type megaspores, while the megaspores of Cymastrobus irvingii show rows of papillae surrounding the trilete mark. In contrast, Sublepidodendron grabaui exhibits slender strobili (length/width = 11.7–13.8) and Lagenicula-type megaspores.
Kowieria alveoformis [21] from the Famennian of South Africa produces Lagenicula megaspores as in Sublepidodendron grabaui, while the numbers of megaspores per each sporangium in these two plants are different. Furthermore, K. alveoformis bears falcate sporophylls homomorphic to vegetative leaves that are dissimilar to those of S. grabaui.
4.3. Comparison with Lycopsids Bearing Monosporangiate Strobili from China (Table 3)
Sublepidodendron songziense occurs in the Upper Devonian Xiejingsi (previously known as Hsiehchingssu) Formation of the Hubei Province and the Wutong Formation of the Anhui Province, China. S. songziense and S. grabaui can be basically distinguished from each other by the different shapes of leaf bases and the ornamentations between them [24]. Detailed studies on these two plants’ Lycospora-type microspores show that the distal surface of the S. songziense microspore is granulated [24], while that of S. grabaui is microspinate [12]. Meanwhile, S. songziense is characterized by a complex branching system consisting of many lateral branches along its trunk, whereas S. grabaui probably displays bifurcations in stems but occasional pseudomonopodial branchings in slender axes (Table 3) and thus may exhibit a different architecture with S. songziense.
Table 3.
Comparison among S. grabaui and related Devonian lycopsids bearing monosporangiate strobili from China.
Table 3.
Comparison among S. grabaui and related Devonian lycopsids bearing monosporangiate strobili from China.
| Sublepidodendron grabaui | Sublepidodendron songziensis | Longostachys latisporophyllum | Minostrobus chaohuensis | Changxingia longifolia | Guangdedendron micrum | |
|---|---|---|---|---|---|---|
| Reference | Wang and Xu [22]; Meng et al. [12]; this study | Wang et al. [24,25]; Meng et al. [9] | Cai and Chen [6] | Meng et al. [8,11] | Wang et al. [10] | Wang et al. [27]; Gao et al. [14] |
| Branching system | at least thrice-dichotomized; isotomously dichotomized or pseudomonopodially branched | multiple dichotomized lateral branches | possible four times-dichotomized or more | eight times dichotomized at least | twice-dichotomized at least | twice-dichotomized at least |
| Leaf (length) | 18–60 mm | 10–15 mm | 20–70 mm | 5–7 mm | 18–25 mm | 20–92 mm |
| Leaf base /cushion (mm) | 1.6–20 in length, 0.3–1.3 mm in width | 1.0–3.5 in length, 0.8–1.5 in width | 9.0–10.0 in length, 1.3–1.5 in width | 6.0–9.0 in length, 1.0–1.6 in width | 10.7–12.5 in length, 2.0–2.3 in width | 15.6–22.7 in length, 2.6–4.3 in width |
| Shape of leaf base/cushion | fusiform or elongated fusiform | Fusiform, rhomboid or oval | narrow-fusiform | narrow-fusiform | rhomboid or fusiform | ovoid or narrow-fusiform |
| Anatomy | primary xylem with a pith and secondary xylem; a solid primary xylem strand | primary xylem, secondary xylem | primary xylem, secondary xylem | primary xylem | – | – |
| Megasporangia strobilus (mm) | 60–160 in length, 6–11 in diameter; singly, in pairs or once-dichotomized | 100–150 in length, 6.0–9.0 in diameter | 30–225 in length, 7.0–10.0 in diameter; singly, occasionally in pairs | up to 125 in length, 4.5–6.0 in diameter | 20–50 in length, 6.0–9.6 in diameter; singly, occasionally in pairs | 50–234 in length, 9.0–24.0 in diameter; singly, in pairs or once-dichotomized |
| Sporangium | elongate ovoid | elongate-elliptical | round or ovoid | spherical to ellipsoidal | elliptical | long-ellipsoidal |
| Megaspore | 661–943 (gula/body = 0.34) | gula/body = 0.51 | 1188–1425 μm in diameter | gula/body = 0.47 | gula/body = 0.52 | gula/body = 0.89 |
| Megaspore number | 8 or more | at least 20 | 4 | 4 | probable 4 | multiple |
| Megaspore type | Lagenicula | Lagenicula | Laevigatisporites? | Lagenicula | Lagenicula | Lagenicula |
Note: –, lack of information.
Longostachys latisporophyllus [6] is a small arborescent lycopsid with megasporangiate strobili from the Middle Devonian (Givetian) of Hunan Province. Its stems are possibly at least four times dichotomized, and its leaves show spiny appendages. Slender strobili consist of spoon-shaped sporophylls and contain Laevigatisporites?-type megaspores. The megasporophyll of Sublepidodendron grabaui shows horizontal pedicel and upturned lamina and produces Lagenicula megaspores.
Minostrobus chaohuensis [8,11,28] is another arborescent lycopsid with monosporangiate strobili found in the Wutong formation from South China. Its aerial axes display multiple dichotomies and fusiform leaf bases/cushions. Protoxylem confined to ridges at the periphery of primary xylem. Each megasporangium of M. chaohuensis contains a single tetrad of Lagenicula megaspores. Sublepidodendron grabaui differs from M. chaohuensis in the number of megaspores in each sporangium and the shape of leaf bases/cushions.
Changxingia longifolia is also reported from the Upper Devonian (Famennian) of South China and is interpreted as a small-sized lycopsid with monosporangiate strobili [10,15]. Its rhomboidal leaf cushions and oblanceolate leaf bases are helically arranged on the stems. Both C. longifolia and Sublepidodendron grabaui produce dichotomized stem and terminal megasporangiate strobili in pairs, while the former bears shorter strobili (20–50 mm in length), and each sporangium contains four Lagenicula-type megaspores.
Guangdedendron micrum shares the same fossil locality with Sublepidodendron grabaui and is regarded as the major tree lycopsid that made up the Xinhang fossil forest [14,27]. In the Leigutai Member of Yongchuan Section, G. micrum is widely distributed, while S. grabaui is restricted to the bottom portion. In G. micrum, the strobili are borne singly, in pairs, and occasionally once-dichotomized, resembling some specimens of S. grabaui; however, the S. grabaui stems dichotomize several times, while G. micrum stems rarely branch. Furthermore, S. grabaui shows a much smaller overall architecture than G. micrum: the axes, strobili and leaves of the former are roughly half the size of those of the latter. The strobili of G. micrum are 70–240 mm in length and 13–23 mm in diameter, while those of S. grabaui are 60–150 mm and 6–11 mm (Figure 8). Along the thick stems or the main trunks, G. micrum displays a fusiform leaf cushion (the length/width ratio = 6:1) with an elliptical leaf scar in the middle, while S. grabaui shows slenderer, elongated fusiform leaf bases (the length/width ratio = 12:1) with a fissure-like false leaf scar. In addition, the Lagenicula-type megaspores of the two plants can be clearly distinguished: megaspores of S. grabaui show smaller gula (gula/body = 0.34), whereas those of G. micrum possess larger gula (gula/body = 0.89).
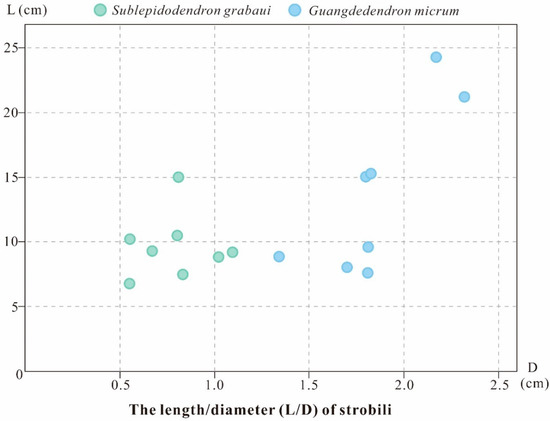
Figure 8.
Comparison plot of the dimensions of Sublepidodendron grabaui and Guangdedendron micrum strobili. Data from Wang et al. [27], Gao et al. [14], and this study.
5. Discussion
Heterosporous lycopsids occurred in the Middle Devonian [4,5,6] and underwent their first evolutionary radiation during the Late Devonian [3,5]. The previous phylogenic result shows that heterosporous lycopsids with monosporangiate strobili originated in those with bisporangiate strobili and are proposed to be the most derived clade [29,30,31]. Wang et al. [32] considered that the reproductive diversification of arborescent lycopsids occurred in the Late Devonian according to Lepidostrobus specimens. Paired or bifurcated strobili occurred in Famennian Sublepidodendron grabaui, as well as Guangdedendron micrum and several other taxa, and such a trait was interpreted as a reproductive strategy to produce more sporangia [14].
Lagenicula-type megaspores are characterized by a prominent gula. Till now, almost all Late Devonian lycopsids with monosporangiate strobili found in South China produce Lagenicula-type megaspores (e.g., Minostrobus chaohuensis, Sublepidodendron songziensis, and Guangdedendron micrum). However, diversification also exists among these Lagenicula megaspores, i.e., the gula height-to-body diameter ratio of these Late Devonian lycopsids varies. Such ratio is 0.34–0.52 in S. grabaui, M. chaohuensis, S. songziense and Changxingia longifolia, while 0.89 in G. micrum. The varied dimensions of gula and body among different species could result in a difference in adaptability to the environment, as some evidence suggests that megaspores with a prominent gula could adapt to the dispersal from a highly dense canopy [33]. In this regard, G. micrum is better adapted to the high density of communities than S. grabaui, which is consistent with the quantitative difference we observed between them in the Xinhang Forest.
The reduced number of megaspores per megasporangium is considered as a derived trait in the Suborder Dichostrobiles of Order Isoëtales sensu lato [8]. The Late Devonian Minostrobus and Changxingia show four megaspores in each megasporangium, while the Carboniferous Sigillaria shows numerous ones [34] and may thus hint a mosaic pattern of characteristic evolution of heterosporous lycopsids [30]. Moreover, some other lycopsids, including Kowieria with bisporangiate strobili, and Wuxia with intercalary megasporangiate fertile zones, also contain four megaspores in each of their megasporangium [16,21]. Therefore, the reduced number of megaspores per megasporangium in lycopsids might be a trait with multiple independent origins beyond the Suborder Dichostrobiles.
6. Conclusions
We excavated new specimens of Sublepidodendron grabaui from Guangde City, Anhui Province, China, to further investigate its characteristics. This plant is now considered a tree lycopsid exhibiting multiply dichotomized stems and occasional lateral branches, and its terminal fertile axes may bear single, paired, or bifurcated megasporangiate strobili. Each megasporangium produces at least eight Lagenicula megaspores. Despite their relatively small quantity, S. grabaui was involved in the formation of the Xinhang Forest for some time.
Author Contributions
P.X. performed the experiments and prepared the figures. P.X. and L.L. wrote the manuscript. D.W. collected the fossils and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported by the National Natural Science Foundation of China (No. 42072016, 41802015) and the Fundamental Research Funds for the Central Universities (No. 2022YQDC04).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The specimen is deposited in the Department of Geology, Peking University, Beijing, China.
Acknowledgments
The authors thank Min Qin (Linyi University), Yi Zhou (Sun Yat-sen University), Xue Gao and Jiang-Nan Yang (Peking University) for field assistance.
Conflicts of Interest
The authors declare that there’s no conflict of interest.
References
- Gensel, P.G.; Berry, C.M. Early lycophyte evolution. Am. Fern J. 2001, 91, 74–98. [Google Scholar] [CrossRef]
- Taylor, T.N.; Taylor, E.L.; Krings, M. Paleobotany: The Biology and Evolution of Fossil Plants, 2nd ed.; Academic Press: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Xu, H.-H.; Wang, Y. The earliest cormose rhizomorph of putative lycopsid affinity from the Middle Devonian of West Junggar, Xinjiang, China. Rev. Palaeobot. Palynol. 2016, 226, 54–57. [Google Scholar] [CrossRef]
- Senkevitsch, M.A.; Jurina, A.L.; Arkhangelskaya, A.D. On fructifications, morphology and anatomy of Givetian Lepidophytes in Kazakhstan (USSR). Palaeontogr. Abt. B Palaophytol. 1993, 230, 43–58. [Google Scholar]
- Hao, S.-G.; Xue, J.-Z.; Wang, Q.; Liu, Z. Yuguangia ordinata gen. et sp. nov., a New Lycopsid from the Middle Devonian (Late Givetian) of Yunnan, China, and Its Phylogenetic Implications. Int. J. Plant Sci. 2007, 168, 1161–1175. [Google Scholar] [CrossRef]
- Cai, C.-Y.; Chen, L.-Z. On a Chinese Givetian lycopod, Longostachys latisporophyllus Zhu, Hu and Feng, emend.: Its morphology, anatomy and reconstruction. Palaeontogr. Abt. B 1996, 238, 1–43. [Google Scholar]
- Xu, H.-H.; Wang, Y.; Wang, Q. A new homosporous, arborescent lycopsid from the Middle Devonian of Xinjiang, Northwest China. Palaeontology 2012, 55, 957–966. [Google Scholar] [CrossRef]
- Meng, M.-C.; Wang, D.-M.; Xue, J.-Z.; Zhu, X. New insights and evolutionary significance of the megasporangiate strobilus of Minostrobus chaohuensis (Lycopsida) from the Upper Devonian of South China. Rev. Palaeobot. Palynol. 2013, 190, 20–40. [Google Scholar] [CrossRef]
- Meng, M.-C.; Wang, D.-M.; Tian, T. New insights on the megasporangiate strobilus of Sublepidodendron songziense from the Late Devonian of Hubei Province. Acta Palaeontol. Sin. 2014, 53, 180–190. [Google Scholar]
- Wang, D.-M.; Meng, M.-C.; Xue, J.-Z.; Basinger, J.F.; Guo, Y.; Liu, L. Changxingia longifolia gen. et sp. nov., a new lycopsid from the Late Devonian of Zhejiang Province, South China. Rev. Palaeobot. Palynol. 2014, 203, 35–47. [Google Scholar] [CrossRef]
- Meng, M.-C.; Wang, D.-M.; Yao, J.-X. Vegetative characters, growth habit and microsporangiate strobilus of lycopsid Minostrobus chaohuensis. PLoS ONE 2015, 10, e0122167. [Google Scholar] [CrossRef]
- Meng, M.-C.; Liu, L.; Wang, D.-M.; Yao, J.-X. Growth architecture and microsporangiate strobilus of Sublepidodendron grabaui (Lycopsida) from the Late Devonian of South China. Rev. Palaeobot. Palynol. 2016, 224, 83–93. [Google Scholar] [CrossRef]
- Wang, D.-M.; Qin, M.; Meng, M.-C.; Liu, L.; Ferguson, D.K. New insights into the heterosporous lycopsid Changxingia from the Upper Devonian Wutong Formation of Zhejiang Province, China. Plant Syst. Evol. 2017, 303, 11–21. [Google Scholar] [CrossRef]
- Gao, X.; Liu, L.; Qin, M.; Zhou, Y.; Mao, L.; Wang, D.M. Re-study of Guangdedendron micrum from the Late Devonian Xinhang forest. BMC Ecol. Evol. 2022, 22, 1–16. [Google Scholar] [CrossRef]
- Gerrienne, P.; Cascales-Minana, B.; Prestianni, C.; Steemans, P.; Li, C.S. Lilingostrobus chaloneri gen. et sp. nov., a Late Devonian woody lycopsid from Hunan, China. PLoS ONE 2018, 13, e0198287. [Google Scholar] [CrossRef]
- Berry, C.M.; Wang, Y.; Cai, C.-Y. A Lycopsid with Novel Reproductive Structures from the Upper Devonian of Jiangsu, China. Int. J. Plant Sci. 2003, 164, 263–273. [Google Scholar] [CrossRef]
- DiMichele, W.A.; Bateman, R.M. Diaphorodenranceae, fam. nov. (lycopsida:Carboniferous): Systematics and evolutionary relationships of Diaphorodendron and Synchysidendron, gen. nov. Am. J. Bot. 1992, 79, 605–617. [Google Scholar] [CrossRef]
- Bateman, R.M. Evolutionary-developmental change in the growth architecture of fossil rhizomorphic lycopsids: Scenarios constructed on cladistic foundations. Biol. Rev. 1994, 69, 527–597. [Google Scholar] [CrossRef]
- Chaloner, W.G. The cone of Cyclostigma kiltorkense Haughton, from the Upper Devonian of Ireland. J. Linn. Soc. Lond. Bot. 1968, 61, 25–36. [Google Scholar] [CrossRef]
- Evreïnoff, M.; Meyer-Berthaud, B.; Decombeix, A.L.; Lebrun, R.; Tafforeau, P. A new Late Devonian isoetalean lycopsid from New South Wales, Australia: Cymastrobus irvingensis gen. et sp. nov. Palaeontol. Electron. 2017, 20, 1–16. [Google Scholar] [CrossRef]
- Gess, R.W.; Prestianni, C. Kowieria alveoformis gen. nov. sp. nov., a new heterosporous lycophyte from the Latest Devonian of Southern Africa. Rev. Palaeobot. Palynol. 2018, 249, 1–8. [Google Scholar] [CrossRef]
- Orlova, O.A.; Zavialova, N.; Snigirevsky, S.; Jurina, A.; Lidskaya, A. Kossoviella timanica Petrosjan emend. from the Upper Devonian of North Timan: Morphology and spore ultrastructure. Earth Environ. Sci. Trans. R. Soc. Edinb. 2018, 108, 355–372. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, H.-H. Sublepidodendron grabaui comb. nov., a lycopsid from the Upper Devonian of China. Bot. J. Linn. Soc. 2005, 149, 299–311. [Google Scholar] [CrossRef]
- Wang, Q.; Hao, S.-G.; Wang, D.-M.; Wang, Y.; Denk, T. A Late Devonian arborescent lycopsid Sublepidodendron songziense Chen emend. (Sublepidodendraceae Kräusel et Weyland 1949) from China, with a revision of the genus Sublepidodendron (Nathorst) Hirmer 1927. Rev. Palaeobot. Palynol. 2003, 127, 269–305. [Google Scholar] [CrossRef]
- Wang, Q.; Hao, S.-G.; Wang, D.-M.; Dilcher, D.L. An anatomically preserved arborescent lycopsid, Sublepidodendron songziense (Sublepidodendraceae), from the Late Devonian of Hubei, China. Am. J. Bot. 2002, 89, 1468–1477. [Google Scholar] [CrossRef] [PubMed]
- Chitaley, S.; Pigg, K. Clevelandodendron ohioensis, gen. et sp. nov., a slender upright lycopsid from the Late Devonian cleveland shale of Ohio. Am. J. Bot. 1996, 83, 781–789. [Google Scholar] [CrossRef]
- Wang, D.-M.; Qin, M.; Liu, L.; Liu, L.; Zhou, Y.; Zhang, Y.; Huang, P.; Xue, J.Z.; Zhang, S.H.; Meng, M.C. The Most extensive Devonian fossil forest with small lycopsid trees bearing the earliest stigmarian roots. Curr. Biol. 2019, 29, 2604–2615.e2602. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, H.-H.; Wang, Q. Re-study of Minostrobus chaohuensis Wang (Lycopsida) from the Upper Devonian of Anhui, South China. Palaeoworld 2012, 21, 20–28. [Google Scholar] [CrossRef]
- Bateman, R.M.; DiMichele, W.A.; Willard, D.A. Experimental Cladistic Analysis of Anatomically Preserved Arborescent Lycopsids from the Carboniferous of Euramerica: An Essay on Paleobotanical Phylogenetics. Ann. Mo. Bot. Gard. 1992, 79, 500–559. [Google Scholar] [CrossRef]
- Dimichele, W.A.; Bateman, R.M. Plant paleoecology and evolutionary inference: Two examples from the Paleozoic. Rev. Palaeobot. Palynol. 1996, 90, 223–247. [Google Scholar] [CrossRef]
- Xue, J.-Z. Phylogeny of Devonian Lycopsids Inferred from Bayesian Phylogenetic Analyses. Acta Geol. Sin.-Engl. Ed. 2011, 85, 569–580. [Google Scholar] [CrossRef]
- Wang, Q.; Li, C.-S.; Geng, B.-Y.; Chitaley, S. A new species of Lepidostrobus from the Upper Devonian of Xinjiang, China and its bearing on the phylogenetic significance of the order Isoëtales. Bot. J. Linn. Soc. 2003, 143, 55–67. [Google Scholar] [CrossRef]
- Hemsley, A.R.; Scott, A.; Collinson, M.E. The architecture and functional biology of freely-dispersed megaspores. In The Evo-lintion of Plant Architecture; Kurmann, M.H., Hemsley, A.R., Eds.; Royal Botanic Gardens, Kew: London, UK, 1999; pp. 253–277. [Google Scholar]
- Pigg, K.B. The Morphology and Reproductive Biology of the Sigillarian Cone Mazocarpon. Bot. Gaz. 1983, 144, 600–613. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).