
Amplectobeluid Radiodont Guanshancaris gen. nov. from the Lower Cambrian (Stage 4) Guanshan Lagerstätte of South China: Biostratigraphic and Paleobiogeographic Implications

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
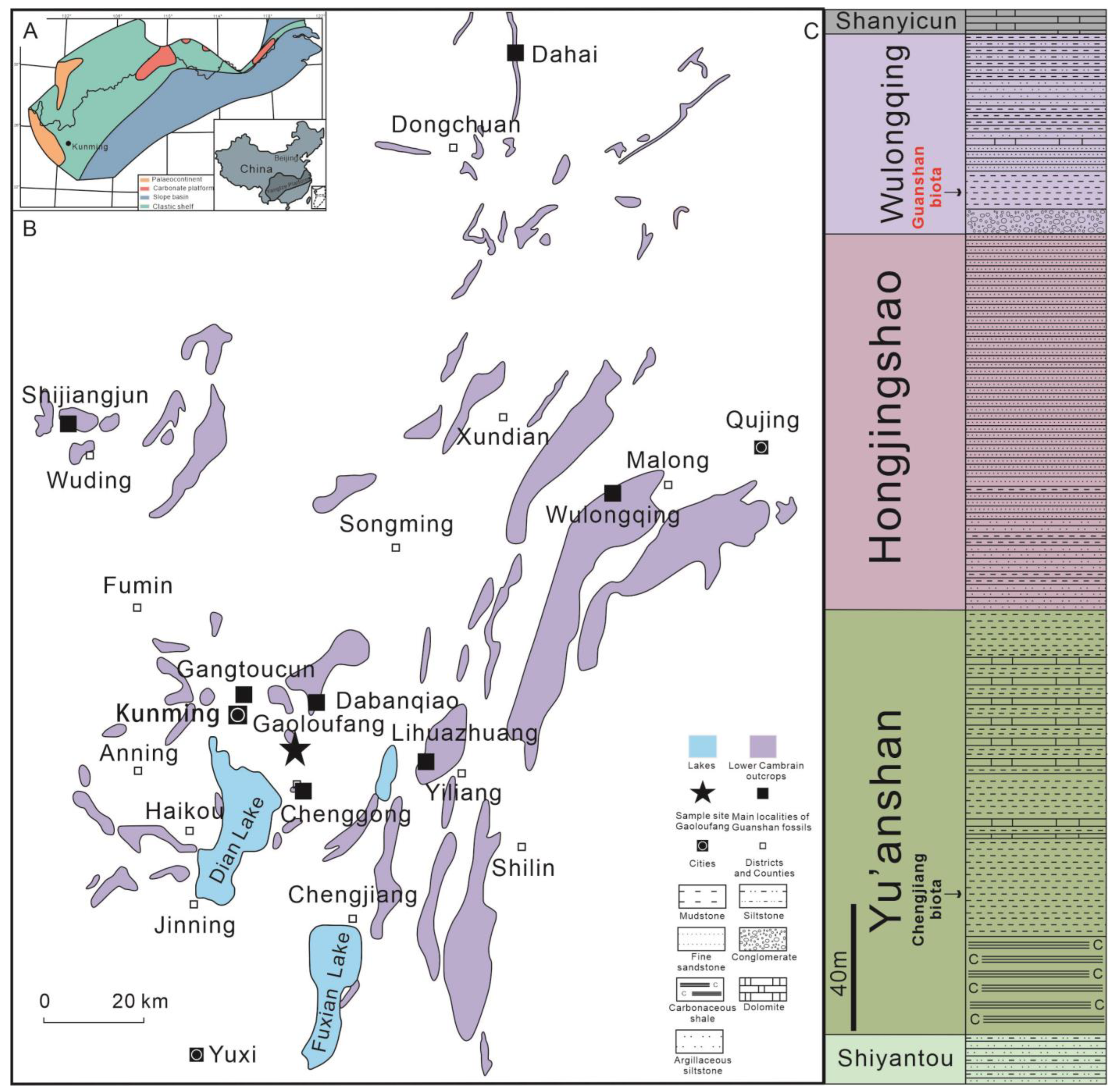
2. Materials and Methods
3. Results
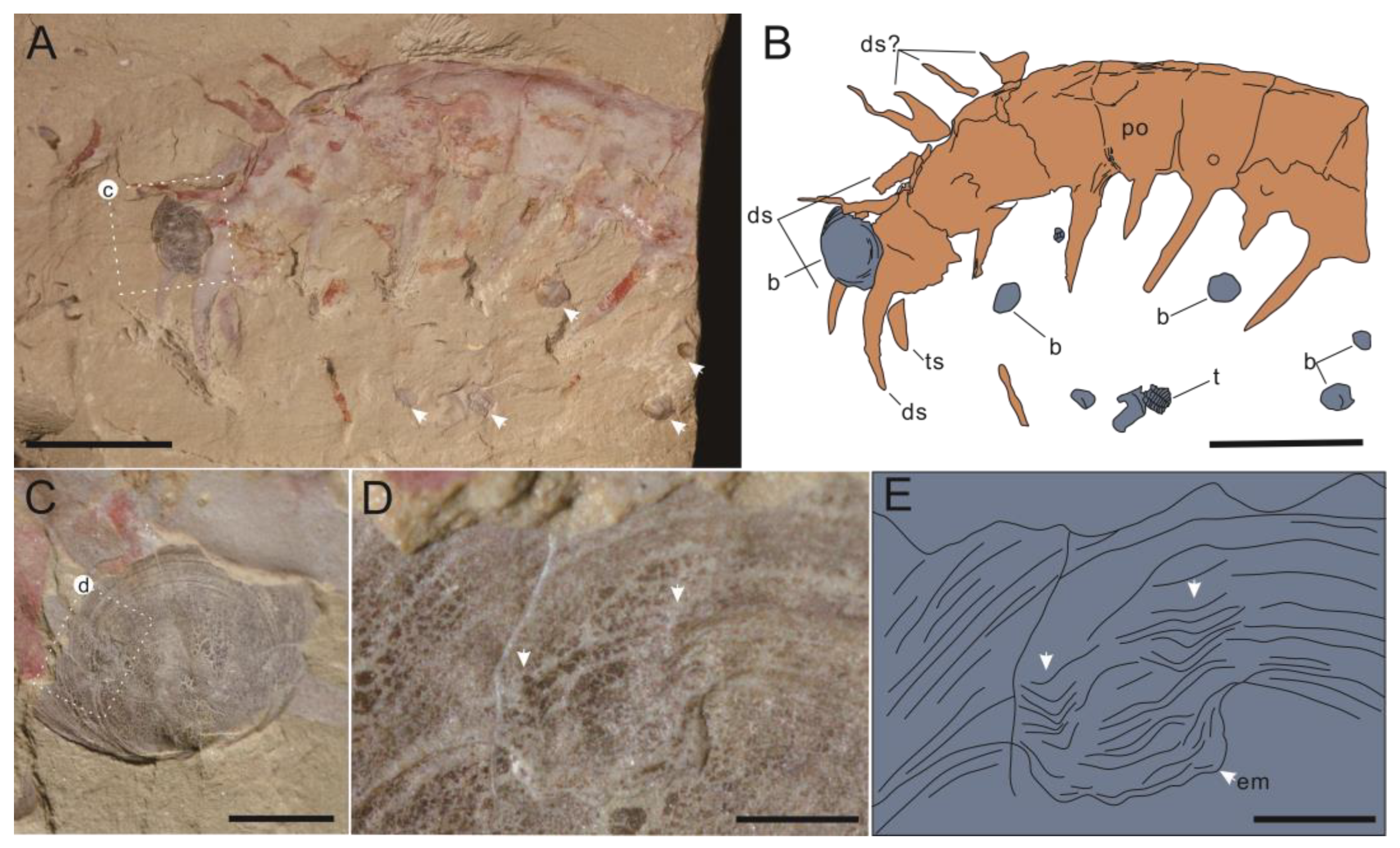
Systematic Paleontology
- Superphylum PANARTHROPODA Nielsen, 1995 [50].
- (Stem–group) ARTHROPODA Von Siebold, 1848 [51].
- Order RADIODONTA Collins, 1996 [26].
- Family AMPLECTOBELUIDAE Pates, Daley, Edgecombe, Cong, Lieberman, Zhang, 2019 [22].
- Genus Guanshancaris gen. nov.
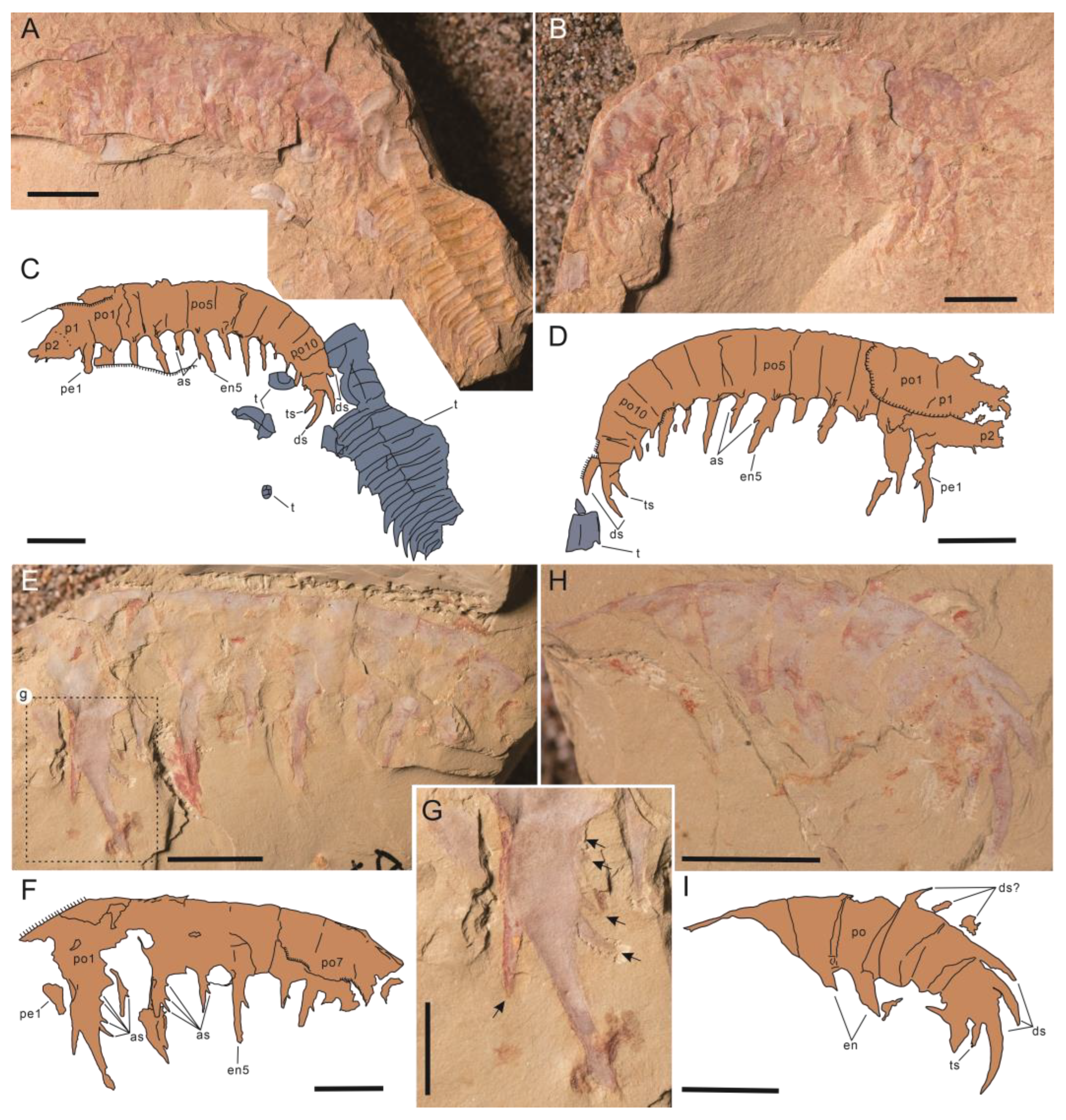
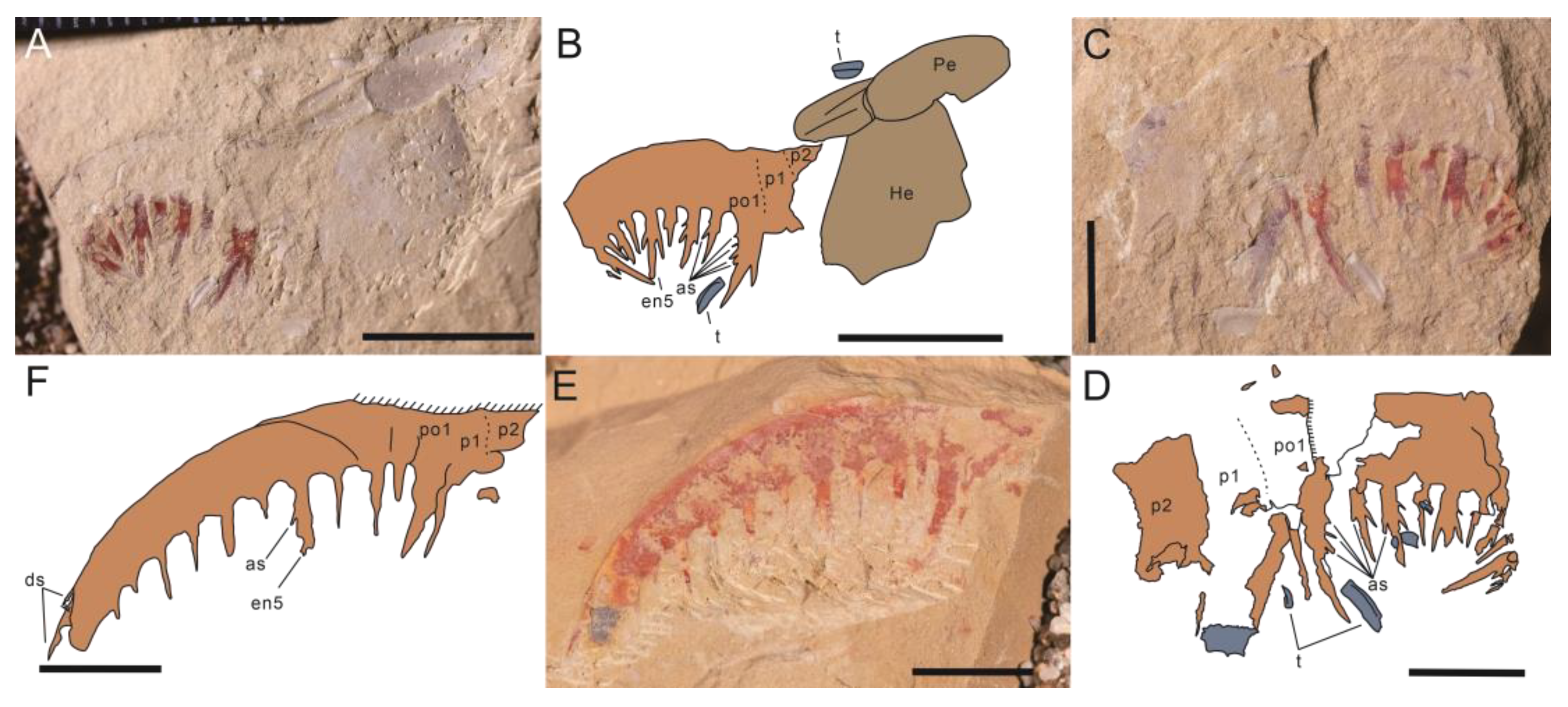
- Guanshancaris kunmingensis (Wang et al., 2013) comb. nov.
- 2013 Anomalocaris kunmingensis sp. nov.; Wang et al., 2013, pp. 3938–3939, Figure 1 [7].
- 2013 Anomalocaris kunmingensis; Hu et al., 2013, Figures 178–180 [28].
- 2014 Anomalocaris kunmingensis; Cong et al., 2014, ext. data Figure 4 [53].
- 2014 Amplectobelua kunmingensis; Vinther et al., 2014, Figure 3 [16].
- 2015 Anomalocaris kunmingensis; Van Roy et al., 2015, ext. data Figure 10 [17].
- 2018 Anomalocaris kunmingensis; Lerosey-Aubril and Pates 2018, Figure 2, sup. Figures 4 and 5 [23].
- 2018 “Anomalocaris” kunmingensis; Liu et al., 2018, sup. Figures 2 and 3C [44].
- 2017 Radiodontan gen. indet. sp. indet.; Zeng et al., 2017, Figure 2 [43].
- 2021 “Anomalocaris” kunmingensis; Jiao et al., 2021, pp. 260–262, Figures 2–4 [10].
4. Discussion
4.1. Guanshancaris as a Possible Durophagous Predator
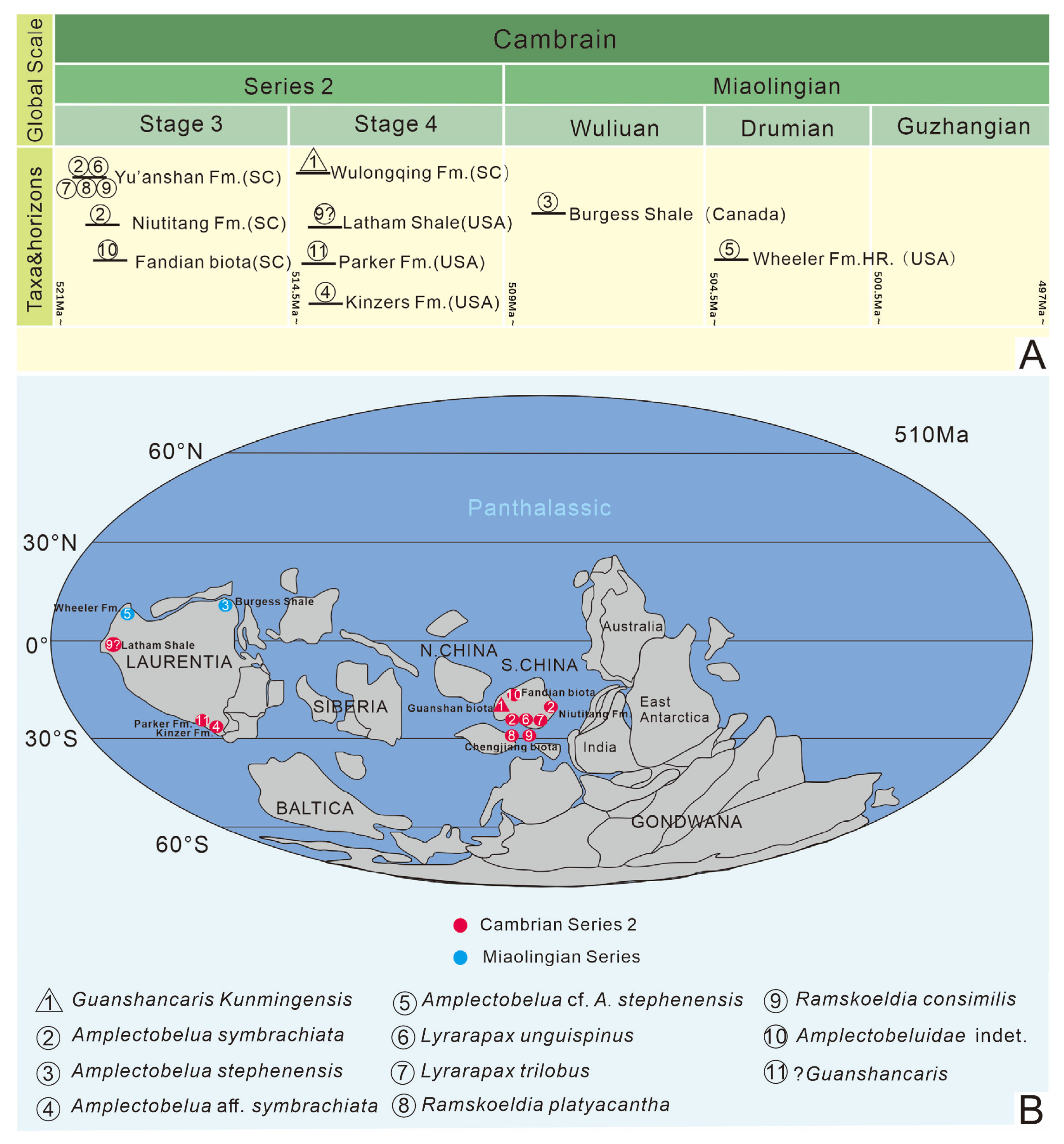
4.2. Paleoenvironmental Distribution
4.3. Spatio–Temporal Distribution of Amplectobeluidae
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daley, A.C.; Budd, G.E.; Caron, J.-B. Morphology and systematics of the anomalocaridid arthropod Hurdia from the Middle Cambrian of British Columbia and Utah. J. Syst. Palaeontol. 2013, 11, 743–787. [Google Scholar] [CrossRef]
- Ortega-Hernández, J. Making sense of ‘lower’ and ‘upper’ stem–group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848. Biol. Rev. Camb. Philos. Soc. 2016, 91, 255–273. [Google Scholar] [CrossRef]
- Whittington, H.; Briggs, D. The Largest Cambrian Animal, Anomalocaris, Burgess Shale, British Columbia. Philos. Trans. R. Soc. B Biol. Sci. 1985, 309, 569–609. [Google Scholar] [CrossRef]
- Chen, J.-Y.; Ramskold, L.; Zhou, G.-Q. Evidence for Monophyly and Arthropod Affinity of Cambrian Giant Predators. Science 1994, 264, 1304–1308. [Google Scholar] [CrossRef]
- Nedin, C. The Emu Bay Shale, a Lower Cambrian fossil Lagerstätte, Kangaroo Island, South Australia. Mem. Assoc. Australas. Palaeontol. 1995, 18, 31–40. [Google Scholar]
- Liu, Q. The first discovery of anomalocaridid appendages from the Balang Formation (Cambrian Series 2) in Hunan, China. Alcheringa Australas. J. Palaeontol. 2013, 37, 338–343. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, D.; Hu, S. New anomalocardid frontal appendages from the Guanshan biota, eastern Yunnan. Chin. Sci. Bull. 2013, 58, 3937–3942. [Google Scholar] [CrossRef] [Green Version]
- Pates, S.; Daley, A.C. The Kinzers Formation (Pennsylvania, USA): The most diverse assemblage of Cambrian Stage 4 radiodonts. Geol. Mag. 2018, 156, 1233–1246. [Google Scholar] [CrossRef] [Green Version]
- Pates, S.; Daley, A. Caryosyntrips: A radiodontan from the Cambrian of Spain, USA and Canada. Pap. Palaeontol. 2017, 3, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Jiao, D.-G.; Pates, S.; Lerosey-Aubril, R.; Ortega-Hernández, J.; Yang, J.; Lan, T.; Zhang, X.-G. The endemic radiodonts of the Cambrian Stage 4 Guanshan biota of South China. Acta Palaeontol. Pol. 2021, 66, 255–274. [Google Scholar] [CrossRef]
- Briggs, D. Giant Predators from the Cambrian of China. Science 1994, 264, 1283–1284. [Google Scholar] [CrossRef]
- Daley, A.C.; Budd, G.E. New anomalocaridid appendages from the Burgess Shale, Canada. Palaeontology 2010, 53, 721–738. [Google Scholar] [CrossRef]
- Daley, A.C.; Bergstrom, J. The oral cone of Anomalocaris is not a classic “peytoia”. Naturwissenschaften 2012, 99, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Daley, A.C.; Paterson, J.R.; Edgecombe, G.D.; García-Bellido, D.C.; Jago, J.B.; Donoghue, P. New anatomical information on Anomalocaris from the Cambrian Emu Bay Shale of South Australia and a reassessment of its inferred predatory habits. Palaeontology 2013, 56, 971–990. [Google Scholar] [CrossRef]
- Daley, A.C.; Edgecombe, G.D. Morphology of Anomalocaris canadensis from the Burgess Shale. J. Paleontol. 2015, 88, 68–91. [Google Scholar] [CrossRef] [PubMed]
- Vinther, J.; Stein, M.; Longrich, N.R.; Harper, D.A. A suspension–feeding anomalocarid from the Early Cambrian. Nature 2014, 507, 496–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Roy, P.; Daley, A.C.; Briggs, D.E. Anomalocaridid trunk limb homology revealed by a giant filter–feeder with paired flaps. Nature 2015, 522, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Moysiuk, J.; Caron, J.B. A new hurdiid radiodont from the Burgess Shale evinces the exploitation of Cambrian infaunal food sources. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191079. [Google Scholar] [CrossRef] [Green Version]
- De Vivo, G.; Lautenschlager, S.; Vinther, J. Three–dimensional modelling, disparity and ecology of the first Cambrian apex predators. Proc. R. Soc. B Biol. Sci. 2021, 288, 20211176. [Google Scholar] [CrossRef]
- Zeng, H.; Zhao, F.; Zhu, M. Innovatiocaris, a complete radiodont from the early Cambrian Chengjiang Lagerstätte and its implications for the phylogeny of Radiodonta. J. Geol. Soc. 2022, 180, jgs2021-164. [Google Scholar] [CrossRef]
- Raymond, P.E. Leanchoilia and other Mid–Cambrian Arthropoda. Bulletion Mus. Comp. Zool. Harv. Univ. 1935, 46, 202–230. [Google Scholar]
- Pates, S.; Daley, A.C.; Edgecombe, G.D.; Cong, P.; Lieberman, B.S.; Zhang, X.G. Systematics, preservation and biogeography of radiodonts from the southern Great Basin, USA, during the upper Dyeran (Cambrian Series 2, Stage 4). Pap. Palaeontol. 2019, 7, 235–262. [Google Scholar] [CrossRef]
- Lerosey-Aubril, R.; Pates, S. New suspension–feeding radiodont suggests evolution of microplanktivory in Cambrian macronekton. Nat. Commun. 2018, 9, 3774. [Google Scholar] [CrossRef] [PubMed]
- Cong, P.; Daley, A.C.; Edgecombe, G.D.; Hou, X. The functional head of the Cambrian radiodontan (stem–group Euarthropoda) Amplectobelua symbrachiata. BMC Evol. Biol. 2017, 17, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cong, P.-Y.; Edgecombe, G.D.; Daley, A.C.; Guo, J.; Pates, S.; Hou, X.-G.; Zhang, X.-G. New radiodonts with gnathobase–like structures from the Cambrian Chengjiang biota and implications for the systematics of Radiodonta. Pap. Palaeontol. 2018, 4, 605–621. [Google Scholar] [CrossRef]
- Collins, D.H. The “evolution” of Anomalocaris and its classification in the arthropod class Dinocarida (nov.) and order Radiodonta (nov.). J. Paleontol. 1996, 70, 280–293. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, G.Q. Biology of the Chengjiang Fauna. Bull. Natl. Mus. Nat. Sci. 1997, 10, 11–106. [Google Scholar]
- Hu, S.X.; Zhu, M.Y.; Luo, H.L.; Steiner, M.; Zhao, F.C.; Li, G.X.; Liu, Q.; Zhang, Z.F. The Guanshan Biota [in Chinese with English Summary]; Yunnan Science and Technology Press: Kunming, China, 2013; 204p. [Google Scholar]
- Paterson, J.R.; García-Bellido, D.C.; Jago, J.B.; Gehling, J.G.; Lee, M.S.Y.; Edgecombe, G.D. The Emu Bay Shale Konservat-Lagerstätte: A view of Cambrian life from East Gondwana. J. Geol. Soc. 2015, 173, 1. [Google Scholar] [CrossRef]
- Skinner, E.S. Taphonomy and depositional circumstances of exceptionally preserved fossils from the Kinzers Formation (Cambrian), southeastern Pennsylvania. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 220, 167–192. [Google Scholar] [CrossRef]
- Ivantsov, A.Y.; Zhuravlev, A.Y.; Leguta, A.V.; Krassilov, V.A.; Melnikova, L.M.; Ushatinskaya, G.T. Palaeoecology of the Early Cambrian Sinsk biota from the Siberian Platform. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 220, 69–88. [Google Scholar] [CrossRef]
- Yuanlong, Z.; Maoyan, Z.H.U.; Babcock, L.E.; Jinliang, Y.; Parsley, R.L.; Jin, P.; Xinglian, Y.; Yue, W. Kaili Biota: A Taphonomic Window on Diversification of Metazoans from the Basal Middle Cambrian: Guizhou, China. Acta Geol. Sin. Engl. Ed. 2005, 79, 751–765. [Google Scholar] [CrossRef]
- Hu, S.; Zhu, M.; Steiner, M.; Luo, H.; Zhao, F.; Liu, Q. Biodiversity and taphonomy of the Early Cambrian Guanshan biota, eastern Yunnan. Sci. China Earth Sci. 2010, 53, 1765–1773. [Google Scholar] [CrossRef]
- Hu, S.; Zhang, Z.; Holmer, L.E.; Skovsted, C.B. Soft–Part Preservation in a Linguliform Brachiopod from the Lower Cambrian Wulongqing Formation (Guanshan Fauna) of Yunnan, South China. Acta Palaeontol. Pol. 2010, 55, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Yong, L.I.; Huilin, L.U.O.; Xiaoping, F.U.; Ting, Y.O.U.; Jiyuan, P.; Qi, L.; Steiner, M. New Record of Palaeoscolecids from the Early Cambrian of Yunnan, China. Acta Geol. Sin. Engl. Ed. 2010, 82, 244–248. [Google Scholar] [CrossRef]
- Hu, S.; Luo, H.; Hou, S.; Erdtmann, B.-D. Eocrinoid echinoderms from the Lower Cambrian Guanshan Fauna in Wuding, Yunnan, China. Chin. Sci. Bull. 2007, 52, 717–719. [Google Scholar] [CrossRef]
- Yang, J.; Hou, X.G.; Cong, P.; Dong, W.; Zhang, Y.X.; Luo, M.B. A new vetulicoliid from lower Cambrian, Kunming, Yunnan. Acta Palaeontol. Sin. 2010, 49, 54–63. [Google Scholar]
- Huilin, L.; Xiaoping, F.; Hu, S.; Yong, L.; Shuguang, H.; Ting, Y.; Jiyuan, P.; Qi, L. A new arthropod, Guangweicaris gen. nov from the Early Cambrian Guanshan Fauna, Kunming, China. Acta Geol. Sin. 2007, 81, 1–7. [Google Scholar] [CrossRef]
- Luo, H.L.; Fu, X.P.; Hu, S.; Li, Y.; Chen, L.Z.; You, T.; Liu, Q. New bivalved arthropods from the Early Cambrian Guanshan fauna in the Kunming and Wuding area. Acta Palaeontol. Sin. 2006, 45, 460–472. [Google Scholar]
- Guang, J.; Pates, S.; Lerosey-Aubril, R.; Ortega-Hernández, J.; Yang, J.; Tian, L.; Zhang, X.-G. New multipodomerous appendages of stem–group euarthropods from the Cambrian (Stage 4) Guanshan Konservat–Lagerstätte. R. Soc. Open Sci. 2021, 8, 211134. [Google Scholar] [CrossRef]
- Guang, J.; Du, K.; Zhang, X.-G.; Yang, J.; Eggink, D. A new small soft–bodied non–trilobite artiopod from the Cambrian Stage 4 Guanshan Biota. Geol. Mag. 2022, 159, 730–734. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, J. New Data on the Bivalved Arthropod Tuzoia From the Cambrian (Series 2, Stage 4) Guanshan Biota in Kunming, Yunnan, Southwest China. Front. Earth Sci. 2022, 10, 862679. [Google Scholar] [CrossRef]
- Zeng, H.; Zhao, F.; Yin, Z.; Zhu, M. A new radiodontan oral cone with a unique combination of anatomical features from the early Cambrian Guanshan Lagerstätte, eastern Yunnan, South China. J. Paleontol. 2017, 92, 40–48. [Google Scholar] [CrossRef]
- Liu, J.; Lerosey-Aubril, R.; Steiner, M.; Dunlop, J.A.; Shu, D.; Paterson, J.R. Origin of raptorial feeding in juvenile euarthropods revealed by a Cambrian radiodontan. Natl. Sci. Rev. 2018, 5, 863–869. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Zhang, Z.; Betts, M.J.; Zhang, Z.; Liu, F. First report on Guanshan Biota (Cambrian Stage 4) at the stratotype area of Wulongqing Formation in Malong County, Eastern Yunnan, China. Geosci. Front. 2019, 10, 1459–1476. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, J.; Chen, F.; Wang, X.D. Ichnology, palaeoenvironment, and ecosystem dynamics of the Early Cambrian (Stage 4, Series 2) Guanshan Biota, South China. Geol. J. 2018, 55, 77–94. [Google Scholar] [CrossRef]
- Guo, J.; Pates, S.; Cong, P.; Daley, A.C.; Edgecombe, G.D.; Chen, T.; Hou, X.; Álvaro, J. A new radiodont (stem Euarthropoda) frontal appendage with a mosaic of characters from the Cambrian (Series 2 Stage 3) Chengjiang biota. Pap. Palaeontol. 2019, 5, 99–110. [Google Scholar] [CrossRef]
- Haug, J.T.; Waloszek, D.; Maas, A.; Liu, Y.U.; Haug, C. Functional morphology, ontogeny and evolution of mantis shrimp–like predators in the Cambrian. Palaeontology 2012, 55, 369–399. [Google Scholar] [CrossRef]
- Wu, Y.; Fu, D.; Ma, J.; Lin, W.; Sun, A.; Zhang, X. Houcaris gen. nov. from the early Cambrian (Stage 3) Chengjiang Lagerstätte expanded the palaeogeographical distribution of tamisiocaridids (Panarthropoda: Radiodonta). PalZ 2021, 95, 209–221. [Google Scholar] [CrossRef]
- Nielsen, C. Animal Evolution: Interrelationships of the Living Phyla; Oxford University Press: Oxford, UK, 1995; p. 467. [Google Scholar]
- von Siebold, C.T.E. Lehrbuch der vergleichenden Anatomie der WirbellosenThiere. Erster Theil. In Lehrbuch Dervergleichenden Anatomie; von Siebold, C.T.E., Stannius, H., Eds.; Verlag von Veit & Company: Berlin, Germany, 1848; pp. 1–679. [Google Scholar]
- Briggs, D.E.G. Anomalocaris, the largest known Cambrian arthropod. Palaeontology 1979, 22, 631–664. [Google Scholar]
- Cong, P.; Ma, X.; Hou, X.; Edgecombe, G.D.; Strausfeld, N.J. Brain structure resolves the segmental affinity of anomalocaridid appendages. Nature 2014, 513, 538–542. [Google Scholar] [CrossRef]
- Pari, G.; Briggs, D.E.G.; Gaines, R.R. The soft–bodied biota of the Cambrian Series 2 Parker Quarry Lagerstätte of northwestern Vermont, USA. J. Paleontol. 2022, 96, 770–790. [Google Scholar] [CrossRef]
- Hou, X.; Bergström, J.R.; Ahlberg, P.E. Anomalocaris and other large animals in the lower Cambrian Chengjiang fauna of southwest China. Gff 1995, 117, 163–183. [Google Scholar]
- Cong, P.; Daley, A.C.; Edgecombe, G.D.; Hou, X.; Chen, A. Morphology of the radiodontan Lyrarapax from the early Cambrian Chengjiang biota. J. Paleontol. 2016, 90, 663–671. [Google Scholar] [CrossRef]
- Nedin, C. Anomalocaris predation on nonmineralized and mineralized trilobites. Geology 1999, 27, 987–990. [Google Scholar] [CrossRef]
- Vannier, J.; Chen, J. Early Cambrian Food Chain: New Evidence from Fossil Aggregates in the Maotianshan Shale Biota, SW China. Palaios 2005, 20, 3–26. [Google Scholar] [CrossRef]
- Zacaï, A.; Vannier, J.; Lerosey-Aubril, R. Reconstructing the diet of a 505–million–year–old arthropod: Sidneyia inexpectans from the Burgess Shale fauna. Arthropod Struct. Dev. 2016, 45, 200–220. [Google Scholar] [CrossRef]
- Zhang, Z.; Holmer, L.E.; Robson, S.P.; Hu, S.; Wang, X.; Wang, H. First record of repaired durophagous shell damages in Early Cambrian lingulate brachiopods with preserved pedicles. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 302, 206–212. [Google Scholar] [CrossRef]
- Bicknell, R.D.C.; Paterson, J.R. Reappraising the early evidence of durophagy and drilling predation in the fossil record: Implications for escalation and the Cambrian Explosion. Biol. Rev. Camb. Philos. Soc. 2018, 93, 754–784. [Google Scholar] [CrossRef]
- Vannier, J.-F. L’Explosion cambrienne ou l’émergence des écosystèmes modernes. Comptes. Rendus. Palevol. 2009, 8, 133–154. [Google Scholar] [CrossRef]
- Hu, S. Taphonomy and Palaeoecology of the Early Cambrian Chengjiang Biota from Eastern Yunnan, China. Berl. Palaobiologische Abh. 2005, 7, 1–197. [Google Scholar]
- Hou, X.-g.; Siveter, D.; Siveter, D.; Aldridge, R.; Cong, P.; Gabbott, S.; Ma, X.; Purnell, M.; Williams, M. The Cambrian Fossils of Chengjiang, China: The Flowering of Early Animal Life, 2nd ed.; John Wiley & Sons Ltd.: Chichester, UK, 2017; pp. 154–160. [Google Scholar]
- Steiner, M.; Zhu, M.; Zhao, Y.-L.; Erdtmann, B.D. Lower Cambrian Burgess Shale–type fossil associations of South China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 220, 129–152. [Google Scholar] [CrossRef]
- Du, K.-S.; Ortega-Hernández, J.; Yang, J.; Yang, X.-Y.; Guo, Q.-H.; Li, W.; He, J.-F.; Li, K.-R.; Du, J.-L.; Hou, J.-B.; et al. A new early Cambrian Konservat–Lagerstätte expands the occurrence of Burgess Shale–type deposits on the Yangtze Platform. Earth–Sci. Rev. 2020, 211, 103409. [Google Scholar] [CrossRef]
- Foster, J.; Gaines, R. Taphonomy and paleoecology of the “middle” Cambrian (Series 3) formations in Utah’s West Desert: Recent finds and new data. Utah Geol. Assoc. Publ. 2016, 45, 291–336. [Google Scholar]
- Steiner, M.; Wallis, E.; Erdtmann, B.D.; Zhao, Y.-l.; Yang, R. Submarine–hydrothermal exhalative ore layers in black shales from South China and associated fossils—Insights into a Lower Cambrian facies and bio–evolution. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2001, 169, 165–191. [Google Scholar] [CrossRef]
- Lerosey-Aubril, R.; Kimmig, J.; Pates, S.; Skabelund, J.; Weug, A.; Ortega-Hernández, J. New exceptionally preserved panarthropods from the Drumian Wheeler Konservat–Lagerstätte of the House Range of Utah. Pap. Palaeontol. 2020, 6, 501–531. [Google Scholar] [CrossRef]
- Pari, G.; Briggs, D.E.G.; Gaines, R.R. The Parker Quarry Lagerstätte of Vermont—The first reported Burgess Shale–type fauna rediscovered. Geology 2021, 49, 693–697. [Google Scholar] [CrossRef]
- Trond, H.T.; Cocks, L.R.M. Chapter 2 New global palaeogeographical reconstructions for the Early Palaeozoic and their generation. Geol. Soc. Lond. Mem. 2013, 38, 5–24. [Google Scholar] [CrossRef]
- Mount, J.D. Characteristics of Early Cambrian faunas from eastern San Bernardino County, California. S. Calif. Paleont. Soc. Spec. Publ. 1980, 2, 19–29. [Google Scholar]
- Zhu, M.-Y.; Babcock, L.E.; Peng, S.-C. Advances in Cambrian stratigraphy and paleontology: Integrating correlation techniques, paleobiology, taphonomy and paleoenvironmental reconstruction. Palaeoworld 2006, 15, 217–222. [Google Scholar] [CrossRef]
- Hough, M.L.; Shields, G.A.; Evins, L.Z.; Strauss, H.; Henderson, R.A.; Mackenzie, S. A major sulphur isotope event atc. 510 Ma: A possible anoxia–extinction–volcanism connection during the Early-Middle Cambrian transition? Terra Nova 2006, 18, 257–263. [Google Scholar] [CrossRef]
- Jourdan, F.; Hodges, K.; Sell, B.; Schaltegger, U.; Wingate, M.T.D.; Evins, L.Z.; Söderlund, U.; Haines, P.W.; Phillips, D.; Blenkinsop, T. High–precision dating of the Kalkarindji large igneous province, Australia, and synchrony with the Early–Middle Cambrian (Stage 4–5) extinction. Geology 2014, 42, 543–546. [Google Scholar] [CrossRef]
- Li, C.; Jin, C.; Planavsky, N.J.; Algeo, T.J.; Cheng, M.; Yang, X.; Zhao, Y.; Xie, S. Coupled oceanic oxygenation and metazoan diversification during the early–middle Cambrian? Geology 2017, 45, 743–746. [Google Scholar] [CrossRef]
- Chang, C.; Hu, W.; Wang, X.; Yu, H.; Yang, A.; Cao, J.; Yao, S. Carbon isotope stratigraphy of the lower to middle Cambrian on the eastern Yangtze Platform, South China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 479, 90–101. [Google Scholar] [CrossRef]
- Chang, C.; Hu, W.; Wang, X.; Huang, K.-J.; Yang, A.; Zhang, X. Nitrogen isotope evidence for an oligotrophic shallow ocean during the Cambrian Stage 4. Geochim. Cosmochim. Acta 2019, 257, 49–67. [Google Scholar] [CrossRef]
- Wu, Y.; Pates, S.; Ma, J.; Lin, W.; Wu, Y.; Zhang, X.; Fu, D. Addressing the Chengjiang conundrum: A palaeoecological view on the rarity of hurdiid radiodonts in this most diverse early Cambrian Lagerstätte. Geosci. Front. 2022, 13, 101430. [Google Scholar] [CrossRef]
- Wu, Y.; Ma, J.; Lin, W.; Sun, A.; Zhang, X.; Fu, D. New anomalocaridids (Panarthropoda: Radiodonta) from the lower Cambrian Chengjiang Lagerstätte: Biostratigraphic and paleobiogeographic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 569, 110333. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
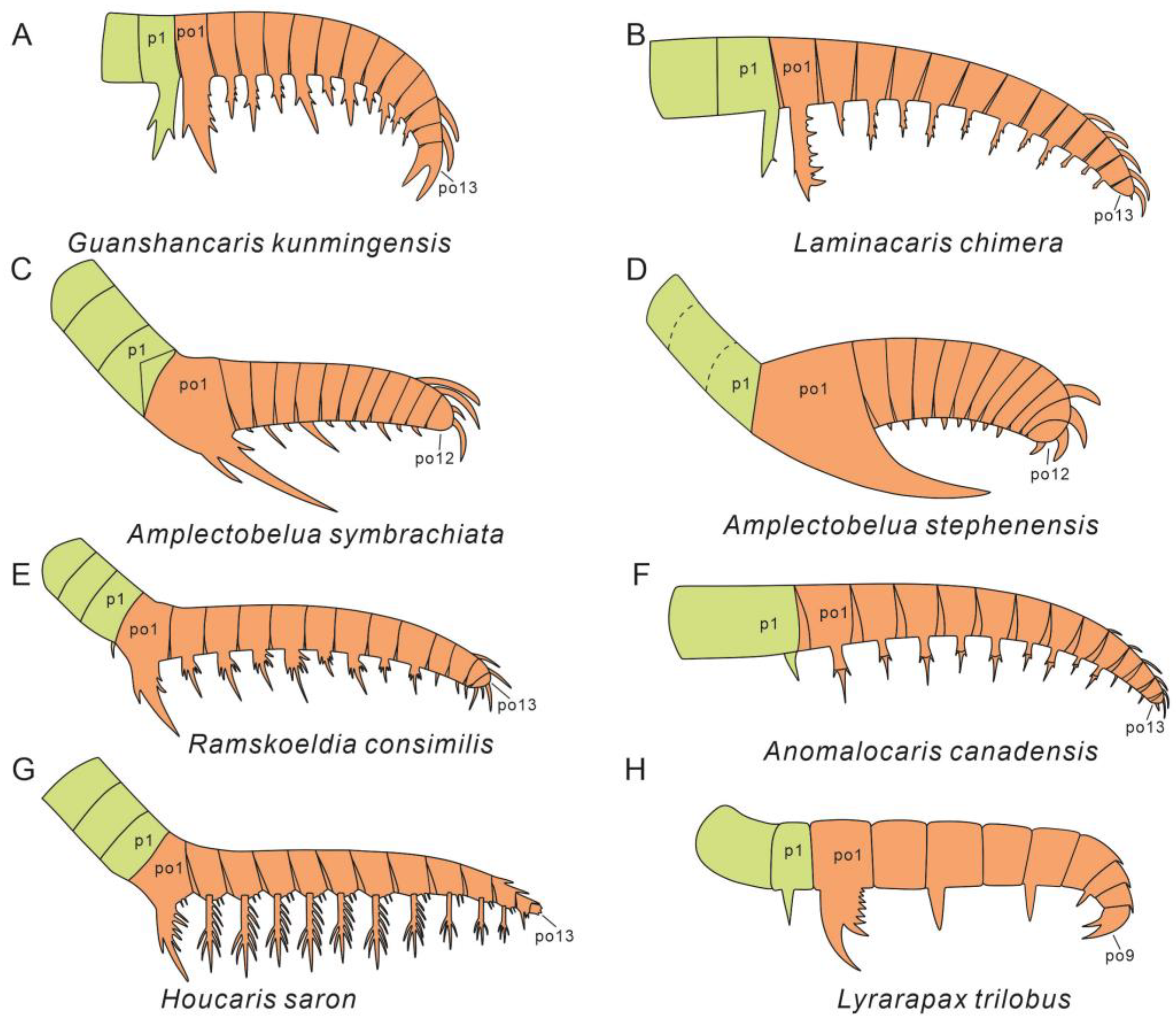
| Morphology of Frontal Appendage | Morphology of Largest en. | References | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| # pd in Peduncle | # pd in d.a.r. | t. m. between pd | # Rows En. | En. in d.a.r. Alternate Long/Short | en. on pd5 Longer than pd3 in d.a.r. | Aux. Paired | Aux. on Both Anterior and Posterior | Aux. Increase Distally Along En. | # Large Aux. | Small Aux. between Large Aux. | ||
| Guanshancaris kunmingensis | 2 | 13 | Yes | 2 | Yes | Yes | No | Yes | Yes | 2 | No | This study [7,10] |
| Laminacaris chimera | 2 | 13 | Yes | 1? | Yes | Yes? | No | Yes | Yes | 5 | Yes | [47] |
| Amplectobelua symbrachiata | 3 | 12 | Yes | 2 | Yes | Yes | No | Yes | No | 2 | No * | [24] |
| Amplectobelua stephenensis | 3 | 12 | Yes | 2 | Yes | Yes | No | No | No | 0 | No * | [12] |
| Ramskoeldia consimilis | 3 | 13 | Yes | 2 | Yes | Yes? | Yes | Yes | No | 2 | No | [25] |
| Anomalocaris canadensis | 1 | 13 | Yes | 2 | Yes | No | Yes | Yes | No | 2 | No * | [15] |
| Houcaris saron | 3 | 13 | Yes | 2 | Yes | No | Yes | Yes | No | 2 | No | [49] |
| Deposits | Age | Fossil Zones | Strata | Taxa | Body Parts | Abundance | Environment | References |
|---|---|---|---|---|---|---|---|---|
| Chengjiang biota, Yunnan, South China | Cam. Stage 3 | Yunnanocephalus Subzone, Eoredlichia–Wutingaspis Zone | Yu’anshan Mb., Chiungchussu Fm. | Amplectobelua symbrachiata | HS, OC, GLS, FA, FL | 1021 | a proximal offshore to lower shoreface setting | [24,25,44,47,53,55,56,64] |
| Lyrarapax unguispinus | HS, OC, EY, FA, FL, GT, TF, TK, NP, MS | 4 | ||||||
| L. trilobus | HS, OC, EY, FA, FL, GT, TK | 2 | ||||||
| Ramskoeldia platyacantha | HS, OC, GLS, FA, FL | 5 | ||||||
| R. consimilis | HS, OC, GLS, FA, FL | 5 | ||||||
| Niutitang Fm., Hunan South China | Cam. Stage 3 | Parabadiella–Mianxiandiscus Interval Zone | Niutitang Fm. | Amplectobelua symbrachiata | FA | 1 | an offshore shelf-basin facies of the Yangtze Platform; a marginal back-arc basin | [65,68] |
| Fandian biota, Sichuan, South China | Cam. Stage 3 | Yilangella–Zhangshania Zone | Yuxiansi and Jiulaodong Fm. | Amplectobelluidae indet. 1 | FA | 1 | an offshore to shoreface continental platform | [66] |
| Guanshan biota, Yunnan, South China | Cam. Stage 4 | Palaeolenus Zone, Megapalaeolenus Zone | Wulongqing Fm. | Guanshancaris kunmingnesis | FA, HS | 149 | an offshore transition between fair-weather wave base and storm wave base | [7,10,28]; this study |
| Kinzers Fm., Pennsylvania, USA | Cam. Stage 4 | Bonnia–Olenellus Zone | Kinzers Fm. | Amplectobelua aff. symbrachiata | FA | 3 | seaward of a carbonate shelf | [8] |
| Parker Quarry Lagerstätte, northwest Vermont, USA | Cam. Stage 4 | Bolbolenellus euryparia Zone or overlying Nephrolenellus multinodus Zone | Parker Fm. | ?Guanshancaris | FA | 1 | seaward margin of a carbonate platform with a steep slope; part of the deep-water Franklin Basin | [54,69] |
| Latham Shale, California, USA | Cam. Stage 4 | Bristolia mohavensis–Peachella iddingsi Zones | Latham Shale, Marble and Providence Mountains | Ramskoeldia consimilis? | FA | 4 | a proximal shelf above the fair-weather wave | [22] |
| Burgess Fm. British Columbia, Canada | Cam. Wuliuan | Latest Glossopleura Zone to early–middle Bathyuriscus–Elrathina Zone | ’Thick Stephen’ (Burgess Shale) Fm. | Amplectobelua stephenensis | FA | 5 | seaward margin of a carbonate platform with a steep slope; below storm wave base | [12] |
| Wheeler Fm. House Range, Utah, USA | Cam. Drumian | Bolaspidella trilobite Zone, Ptychagnostus atavus agnostoid Zone | Wheeler Fm. | Amplectobelua cf. A. stephenensis | FA | 1 | offshore, outer shelf, adjacent to a carbonate ramp; deep-water settings; below storm wave base | [69] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Wu, Y.; Lin, W.; Ma, J.; Wu, Y.; Fu, D. Amplectobeluid Radiodont Guanshancaris gen. nov. from the Lower Cambrian (Stage 4) Guanshan Lagerstätte of South China: Biostratigraphic and Paleobiogeographic Implications. Biology 2023, 12, 583. https://doi.org/10.3390/biology12040583
Zhang M, Wu Y, Lin W, Ma J, Wu Y, Fu D. Amplectobeluid Radiodont Guanshancaris gen. nov. from the Lower Cambrian (Stage 4) Guanshan Lagerstätte of South China: Biostratigraphic and Paleobiogeographic Implications. Biology. 2023; 12(4):583. https://doi.org/10.3390/biology12040583
Chicago/Turabian StyleZhang, Mingjing, Yu Wu, Weiliang Lin, Jiaxin Ma, Yuheng Wu, and Dongjing Fu. 2023. "Amplectobeluid Radiodont Guanshancaris gen. nov. from the Lower Cambrian (Stage 4) Guanshan Lagerstätte of South China: Biostratigraphic and Paleobiogeographic Implications" Biology 12, no. 4: 583. https://doi.org/10.3390/biology12040583
APA StyleZhang, M., Wu, Y., Lin, W., Ma, J., Wu, Y., & Fu, D. (2023). Amplectobeluid Radiodont Guanshancaris gen. nov. from the Lower Cambrian (Stage 4) Guanshan Lagerstätte of South China: Biostratigraphic and Paleobiogeographic Implications. Biology, 12(4), 583. https://doi.org/10.3390/biology12040583