



Biological Invasions Affect Resource Processing in Aquatic Ecosystems: The Invasive Amphipod Dikerogammarus villosus Impacts Detritus Processing through High Abundance Rather than Differential Response to Temperature


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
Data Analysis
3. Results
3.1. Shredding
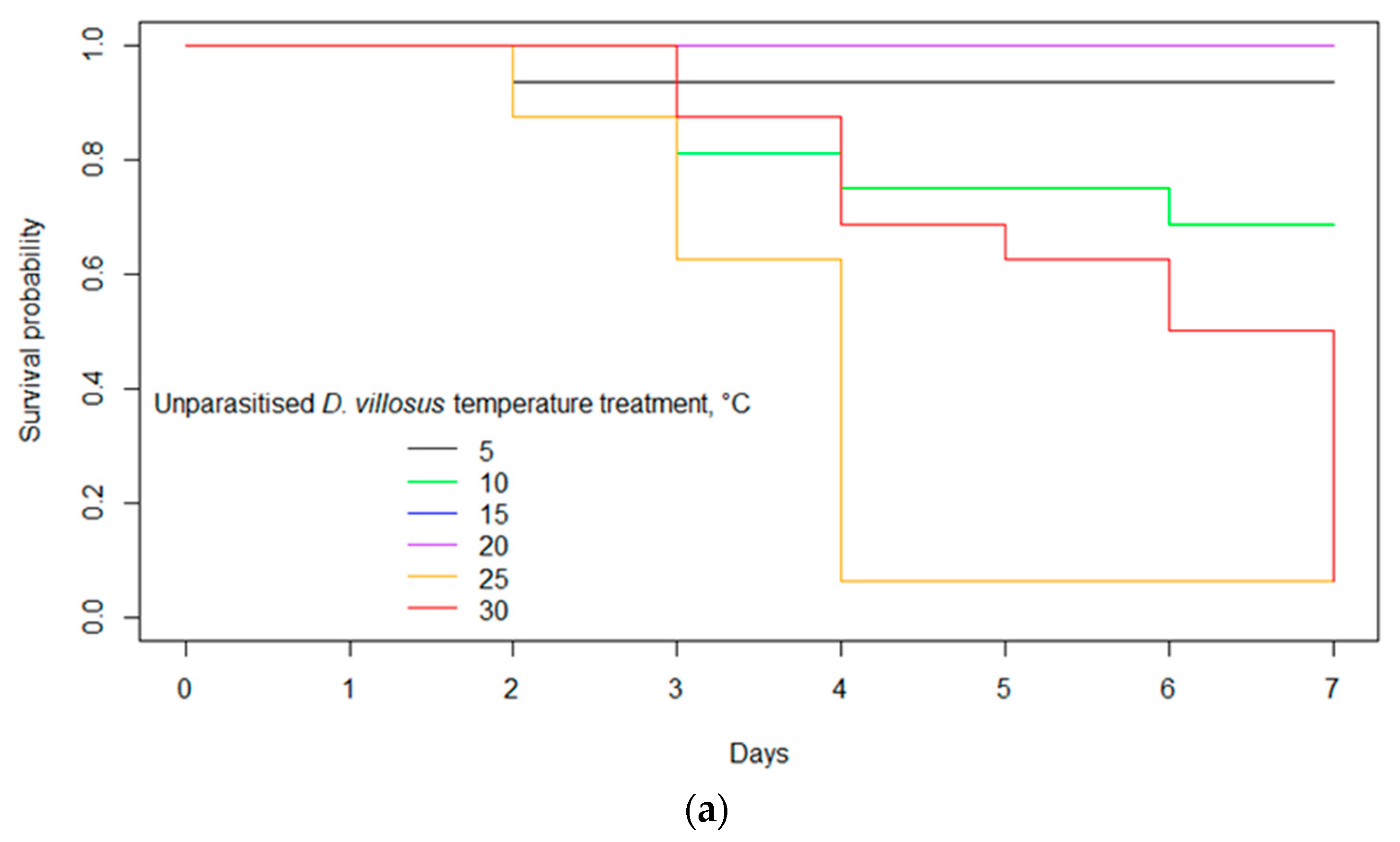
3.2. Survival
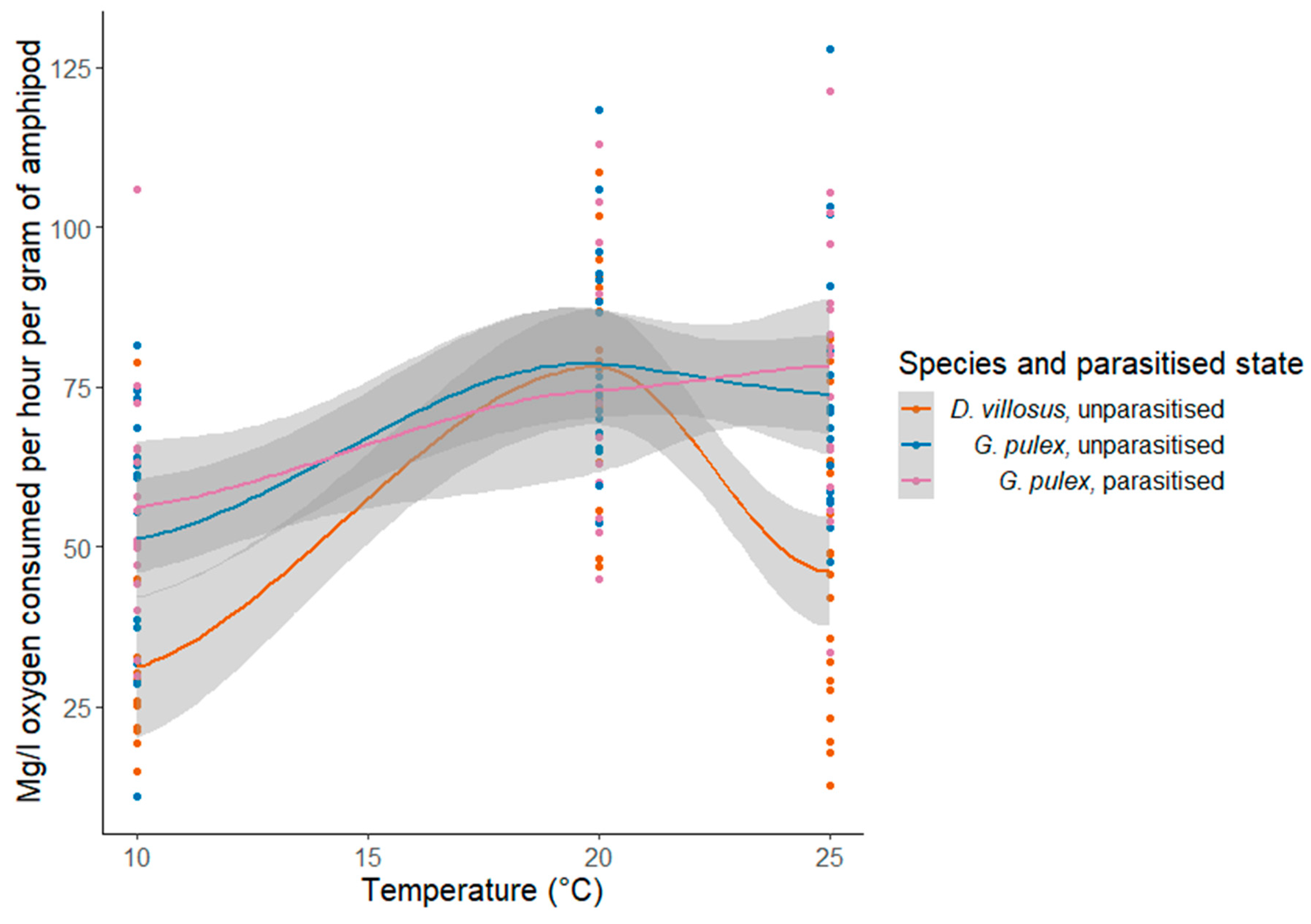
3.3. Metabolic Rate
3.4. Invader Relative Impact Potential
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orr, J.A.; Vinebrooke, R.D.; Jackson, M.C.; Kroeker, K.J.; Kordas, R.L.; Mantyka-Pringle, C.; Van den Brink, P.J.; De Laender, F.; Stoks, R.; Holmstrup, M.; et al. Towards a Unified Study of Multiple Stressors: Divisions and Common Goals across Research Disciplines. Proc. R. Soc. B 2020, 287, 20200421. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S.; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of Changing Biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, B.J.; Duffy, E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity Loss and Its Impact on Humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fincham, W.; Dunn, A.M.; Brown, L.E.; Hesketh, H.; Roy, H.E.; William, N. Invasion Success of a Widespread Invasive Predator May Be Explained by a High Predatory Efficacy but May Be Influenced by Pathogen Infection. Biol. Invasions 2019, 21, 3545–3560. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, A.L.; Dafforn, K.A.; Chariton, A.A.; Johnston, E.L.; Mayer-Pinto, M. After Decades of Stressor Research in Urban Estuarine Ecosystems the Focus Is Still on Single Stressors: A Systematic Literature Review and Meta-Analysis. Sci. Total Environ. 2019, 684, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Strayer, D.L. Alien Species in Fresh Waters: Ecological Effects, Interactions with Other Stressors, and Prospects for the Future. Freshw. Biol. 2010, 55 (Suppl. S1), 152–174. [Google Scholar] [CrossRef]
- Ormerod, S.J.; Dobson, M.; Hildrew, A.G.; Townsend, C.R. Multiple Stressors in Freshwater Ecosystems. Freshw. Biol. 2010, 55, 1–4. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater Biodiversity: Importance, Threats, Status and Conservation Challenges. Biol. Rev. 2006, 81, 163. [Google Scholar] [CrossRef]
- WWF. Freshwater Biodiversity. Available online: https://wwf.panda.org/discover/our_focus/freshwater_practice/freshwater_biodiversity_222/ (accessed on 22 May 2023).
- Larsen, S.; Muehlbauer, J.D.; Marti, E. Resource Subsidies between Stream and Terrestrial Ecosystems under Global Change. Glob. Chang. Biol. 2016, 22, 2489–2504. [Google Scholar] [CrossRef]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Synthesis; Millennium Ecosystem Assessment: Washington, DC, USA, 2005. [Google Scholar]
- Gessner, M.O.; Swan, C.M.; Dang, C.K.; McKie, B.G.; Bardgett, R.D.; Wall, D.H.; Hättenschwiler, S. Diversity Meets Decomposition. Trends Ecol. Evol. 2010, 25, 372–380. [Google Scholar] [CrossRef]
- Covich, A.P.; Austen, M.C.; Bärlocher, F.; Chauvet, E.; Cardinale, B.J.; Biles, C.L.; Inchausti, P.; Dangles, O.; Solan, M.; Gessner, M.O.; et al. The Role of Biodiversity in the Functioning of Freshwater and Marine Benthic Ecosystems. Bioscience 2004, 54, 767–775. [Google Scholar] [CrossRef]
- Cummins, K.W.; Merritt, R.W.; Andrade, P.C.N. The Use of Invertebrate Functional Groups to Characterize Ecosystem Attributes in Selected Streams and Rivers in South Brazil. Stud. Neotrop. Fauna Environ. 2005, 40, 69–89. [Google Scholar] [CrossRef]
- Alp, M.; Cucherousset, J.; Buoro, M.; Lecerf, A. Phenological Response of a Key Ecosystem Function to Biological Invasion. Ecol. Lett. 2016, 19, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R.; et al. Global Threats to Human Water Security and River Biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutowsky, L.F.G.; Giacomini, H.C.; de Kerckhove, D.T.; Mackereth, R.; McCormick, D.; Chu, C. Quantifying Multiple Pressure Interactions Affecting Populations of a Recreationally and Commercially Important Freshwater Fish. Glob. Chang. Biol. 2019, 25, 1049–1062. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F.W.; Vitule, J.R.S. The Largemouth Bass Micropterus Salmoides (Lacepède, 1802): Impacts of a Powerful Freshwater Fish Predator Outside of Its Native Range. Rev. Fish Biol. Fish. 2019, 29, 639–652. [Google Scholar] [CrossRef]
- Fletcher, R.A.; Brooks, R.K.; Lakoba, V.T.; Sharma, G.; Heminger, A.R.; Dickinson, C.C.; Barney, J.N. Invasive Plants Negatively Impact Native, but Not Exotic, Animals. Glob. Chang. Biol. 2019, 25, 3694–3705. [Google Scholar] [CrossRef]
- Gallardo, B.; Clavero, M.; Sánchez, M.I.; Vilà, M. Global Ecological Impacts of Invasive Species in Aquatic Ecosystems. Glob. Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, M.; Molofsky, J.; With, K.A.; Cabin, R.J.; Cohen, J.E.; Norman, C.; Mccauley, D.E.; et al. The Population Biology of Invasive Species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Vitousek, P.M.; D’Antonio, C.M.; Loope, L.L.; Westbrooks, R. Biological Invasions as Global Environmental Change. Am. Sci. 1996, 84, 468–478. [Google Scholar]
- Rewicz, T.; Grabowski, M.; MacNeil, C.; Bącela-Spychalska, K. The Profile of a ‘Perfect’ Invader—The Case of Killer Shrimp, Dikerogammarus villosus. Aquat. Invasions 2014, 9, 267–288. [Google Scholar] [CrossRef]
- Doherty-Bone, T.M.; Dunn, A.M.; Jackson, F.L.; Brown, L.E. Multi-Faceted Impacts of Native and Invasive Alien Decapod Species on Freshwater Biodiversity and Ecosystem Functioning. Freshw. Biol. 2019, 64, 461–473. [Google Scholar] [CrossRef]
- DeBoer, J.A.; Anderson, A.M.; Casper, A.F. Multi-Trophic Response to Invasive Silver Carp (Hypophthalmichthys Molitrix) in a Large Floodplain River. Freshw. Biol. 2018, 63, 597–611. [Google Scholar] [CrossRef]
- de Omena, P.M.; Srivastava, D.S.; Romero, G.Q. Does the Strength of Cross-Ecosystem Trophic Cascades Vary with Ecosystem Size? A Test Using a Natural Microcosm. Freshw. Biol. 2017, 62, 724–736. [Google Scholar] [CrossRef]
- MacNeil, C.; Dick, J.T.A.; Elwood, R.W. The Trophic Ecology of Freshwater Gammarus Spp. (Crustacea: Amphipoda): Problems and Perspectives Concerning the Functional Feeding Group Concept. Biol. Rev. Camb. Philos. Soc. 1997, 72, 349–364. [Google Scholar] [CrossRef]
- Dangles, O.; Malmqvist, B. Species Richness-Decomposition Relationships Depend on Species Dominance. Ecol. Lett. 2004, 7, 395–402. [Google Scholar] [CrossRef]
- Grabowski, M.; Bacela, K.; Konopacka, A. How to Be an Invasive Gammarid (Amphipoda: Gammaroidea)–Comparison of Life History Traits. Hydrobiologia 2007, 590, 75–84. [Google Scholar] [CrossRef]
- Species Alerts—GB Non-Native Species Secretariat. Available online: http://www.nonnativespecies.org/alerts/index.cfm?id=3 (accessed on 17 June 2018).
- Rewicz, T.; Wattier, R.; Grabowski, M.; Rigaud, T.; Bącela-Spychalska, K. Out of the Black Sea: Phylogeography of the Invasive Killer Shrimp Dikerogammarus villosus across Europe. PLoS ONE 2015, 10, e0118121. [Google Scholar] [CrossRef] [Green Version]
- Dodd, J.A.; Dick, J.T.A.; Alexander, M.E.; Macneil, C.; Dunn, A.M.; Aldridge, D.C. Predicting the Ecological Impacts of a New Freshwater Invader: Functional Responses and Prey Selectivity of the ‘Killer Shrimp’, Dikerogammarus villosus, Compared to the Native Gammarus pulex. Freshw. Biol. 2014, 59, 337–352. [Google Scholar] [CrossRef] [Green Version]
- MacNeil, C.; Platvoet, D. The Predatory Impact of the Freshwater Invader Dikerogammarus villosus on Native Gammarus pulex (Crustacea: Amphipoda); Influences of Differential Microdistribution and Food Resources. J. Zool. 2005, 267, 31. [Google Scholar] [CrossRef]
- van Riel, M.C.; Healy, E.P.; van der Velde, G.; bij de Vaate, A. Interference Competition among Native and Invader Amphipods. Acta Oecologica 2007, 31, 282–289. [Google Scholar] [CrossRef]
- Kinzler, W.; Kley, A.; Mayer, G.; Waloszek, D.; Maier, G. Mutual Predation between and Cannibalism within Several Freshwater Gammarids: Dikerogammarus villosus versus One Native and Three Invasives. Aquat. Ecol. 2009, 43, 457–464. [Google Scholar] [CrossRef]
- Bollache, L.; Dick, J.T.; Farnsworth, K.D.; Montgomery, W.I. Comparison of the Functional Responses of Invasive and Native Amphipods. Biol. Lett. 2008, 4, 166–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatcher, M.J.; Dunn, A.M. Parasites in Ecological Communities: From Interactions to Ecosystems; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Dunn, A.M.; Torchin, M.E.; Hatcher, M.J.; Kotanen, P.M.; Blumenthal, D.M.; Byers, J.E.; Coon, C.A.C.; Frankel, V.M.; Holt, R.D.; Hufbauer, R.A.; et al. Indirect Effects of Parasites in Invasions. Funct. Ecol. 2012, 26, 1262–1274. [Google Scholar] [CrossRef] [Green Version]
- Grabner, D.S. Hidden Diversity: Parasites of Stream Arthropods. Freshw. Biol. 2017, 62, 52–64. [Google Scholar] [CrossRef]
- Fielding, N.J.; MacNeil, C.; Dick, J.T.A.; Elwood, R.W.; Riddell, G.E.; Dunn, A.M. Effects of the Acanthocephalan Parasite Echinorhynchus truttae on the Feeding Ecology of Gammarus pulex (Crustacea: Amphipoda). J. Zool. 2003, 261, 321–325. [Google Scholar] [CrossRef]
- Macneil, C.; Fielding, N.; Hume, K.D.; Dick, J.T.A.; Elwood, R.W.; Hatcher, M.J.; Dunn, A.M. Parasite Altered Micro-Distribution of Gammarus pulex (Crustacea: Amphipoda). Int. J. Parasitol. 2003, 33, 57–64. [Google Scholar] [CrossRef]
- Lagrue, C.; Gueventam, A.; Bollache, L. Manipulative Parasites May Not Alter Intermediate Host Distribution but Still Enhance Their Transmission: Field Evidence for Increased Vulnerability to Definitive Hosts and Non-Host Predator Avoidance. Parasitology 2013, 140, 258–265. [Google Scholar] [CrossRef]
- Baldauf, S.A.; Thünken, T.; Frommen, J.G.; Bakker, T.C.M.; Heupel, O.; Kullmann, H. Infection with an Acanthocephalan Manipulates an Amphipod’s Reaction to a Fish Predator’s Odours. Int. J. Parasitol. 2007, 37, 61–65. [Google Scholar] [CrossRef]
- Perrot-Minnot, M.-J.; Kaldonski, N.; Cézilly, F. Increased Susceptibility to Predation and Altered Anti-Predator Behaviour in an Acanthocephalan-Infected Amphipod. Int. J. Parasitol. 2007, 37, 645–651. [Google Scholar] [CrossRef]
- Dick, J.T.A.; Armstrong, M.; Clarke, H.C.; Farnsworth, K.D.; Hatcher, M.J.; Ennis, M.; Kelly, A.; Dunn, A.M. Parasitism May Enhance Rather than Reduce the Predatory Impact of an Invader. Biol. Lett. 2010, 6, 636–638. [Google Scholar] [CrossRef] [PubMed]
- Perrot-Minnot, M.-J.; Maddaleno, M.; Cézilly, F. Parasite-Induced Inversion of Geotaxis in a Freshwater Amphipod: A Role for Anaerobic Metabolism? Funct. Ecol. 2016, 30, 780–788. [Google Scholar] [CrossRef] [Green Version]
- Labaude, S.; Cézilly, F.; Tercier, X.; Rigaud, T. Influence of Host Nutritional Condition on Post-Infection Traits in the Association between the Manipulative Acanthocephalan Pomphorhynchus Laevis and the Amphipod Gammarus pulex. Parasit. Vectors 2015, 8, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arundell, K.; Dunn, A.; Alexander, J.; Shearman, R.; Archer, N.; Ironside, J.E. Enemy Release and Genetic Founder Effects in Invasive Killer Shrimp Populations of Great Britain. Biol. Invasions 2015, 17, 1439–1451. [Google Scholar] [CrossRef] [Green Version]
- Gendron, A.D.; Marcogliese, D.J.; Thomas, M. Invasive Species Are Less Parasitized than Native Competitors, but for How Long? The Case of the Round Goby in the Great Lakes-St. Lawrence Basin. Biol. Invasions 2012, 14, 367–384. [Google Scholar] [CrossRef]
- Jenkins, G.J.; Murphy, J.M.; Sexton, D.M.H.; Lowe, J.; Jones, P.D.; Chris, K. UK Climate Projections: Briefing Report; Met Office Hadley Centre: Exeter, UK, 2009. [Google Scholar]
- Pachauri, R.K.; Meyer, L.; Van Ypersele, J.-P.; Brinkman, S.; Van Kesteren, L.; Leprince-Ringuet, N.; Van Boxmeer, F. IPCC, 2014: Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Leung, J.Y.S.; Connell, S.D.; Russell, B.D. Heatwaves Diminish the Survival of a Subtidal Gastropod through Reduction in Energy Budget and Depletion of Energy Reserves. Sci. Rep. 2017, 7, 17688. [Google Scholar] [CrossRef] [Green Version]
- Bertani, I.; Primicerio, R.; Rossetti, G. Extreme Climatic Event Triggers a Lake Regime Shift That Propagates Across Multiple Trophic Levels. Ecosystems 2016, 19, 16–31. [Google Scholar] [CrossRef]
- Jankowski, T.; Livingstone, D.M.; Bührer, H.; Forster, R.; Niederhauser, P. Consequences of the 2003 European Heat Wave for Lake Temperature Profiles, Thermal Stability, and Hypolimnetic Oxygen Depletion: Implications for a Warmer World. Limnol. Oceanogr. 2006, 51, 815–819. [Google Scholar] [CrossRef] [Green Version]
- Daufresne, M.; Bady, P.; Fruget, J.-F. Impacts of Global Changes and Extreme Hydroclimatic Events on Macroinvertebrate Community Structures in the French Rhône River. Oecologia 2007, 151, 544–559. [Google Scholar] [CrossRef]
- Hupało, K.; Riss, H.W.; Grabowski, M.; Thiel, J.; Bącela-Spychalska, K.; Meyer, E.I. Climate Change as a Possible Driver of Invasion and Differential in HSP70 Expression in Two Genetically Distinct Populations of the Invasive Killer Shrimp, Dikerogammarus villosus. Biol. Invasions 2018, 20, 2047–2059. [Google Scholar] [CrossRef] [Green Version]
- Iacarella, J.C.; Dick, J.T.A.; Alexander, M.E.; Ricciardi, A. Ecological Impacts of Invasive Alien Species along Temperature Gradients: Testing the Role of Environmental Matching. Ecol. Appl. 2015, 25, 706–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diez, J.M.; D’Antonio, C.M.; Dukes, J.S.; Grosholz, E.D.; Olden, J.D.; Sorte, C.J.B.; Blumenthal, D.M.; Bradley, B.A.; Early, R.; Ibanez, I.; et al. Will Extreme Climatic Events Facilitate Biological Invasions? Front. Ecol. Environ. 2012, 10, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Tepolt, C.K.; Somero, G.N. Master of All Trades: Thermal Acclimation and Adaptation of Cardiac Function in a Broadly Distributed Marine Invasive Species, the European Green Crab, Carcinus Maenas. J. Exp. Biol. 2014, 217, 1129–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truhlar, A.M.; Dodd, J.A.; Aldridge, D.C. Differential Leaf-Litter Processing by Native (Gammarus pulex) and Invasive (Dikerogammarus villosus) Freshwater Crustaceans under Environmental Extremes. Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 24, 56–65. [Google Scholar] [CrossRef]
- Kenna, D.; Fincham, W.N.W.; Dunn, A.M.; Brown, L.E.; Hassall, C. Antagonistic Effects of Biological Invasion and Environmental Warming on Detritus Processing in Freshwater Ecosystems. Oecologia 2017, 183, 875–886. [Google Scholar] [CrossRef] [Green Version]
- Gehman, A.L.M.; Harley, C.D.G. Symbiotic Endolithic Microbes Alter Host Morphology and Reduce Host Vulnerability to High Environmental Temperatures. Ecosphere 2019, 10, 02683. [Google Scholar] [CrossRef] [Green Version]
- Greenspan, S.E.; Bower, D.S.; Roznik, E.A.; Pike, D.A.; Marantelli, G.; Alford, R.A.; Schwarzkopf, L.; Scheffers, B.R. Infection Increases Vulnerability to Climate Change via Effects on Host Thermal Tolerance. Sci. Rep. 2017, 7, 9349. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Loras, A.; Boyero, L.; Correa-Araneda, F.; Tejedo, M.; Hettyey, A.; Bosch, J. Infection with Batrachochytrium dendrobatidis Lowers Heat Tolerance of Tadpole Hosts and Cannot Be Cleared by Brief Exposure to CTmax. PLoS ONE 2019, 14, e0216090. [Google Scholar] [CrossRef] [Green Version]
- Musgrave, K.; Bartlow, A.W.; Fair, J.M. Long-Term Variation in Environmental Conditions Influences Host-Parasite Fitness. Ecol. Evol. 2019, 9, 7688–7703. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H.; Burger, J.R.; Hou, C.; Hall, C.A.S. The Pace of Life: Metabolic Energy, Biological Time, and Life History. Integr. Comp. Biol. 2022, 62, 1479–1491. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a Metabolic Theory of Ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Glazier, D.S.; Gring, J.P.; Holsopple, J.R.; Gjoni, V. Temperature Effects on Metabolic Scaling of a Keystone Freshwater Crustacean Depend on Fish-Predation Regime. J. Exp. Biol. 2020, 223, jeb232322. [Google Scholar] [CrossRef] [PubMed]
- Gjoni, V.; Basset, A.; Glazier, D.S. Temperature and Predator Cues Interactively Affect Ontogenetic Metabolic Scaling of Aquatic Amphipods. Biol. Lett. 2020, 16, 20200267. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, M.J.; Dick, J.T.A.; Bojko, J.; Stentiford, G.D.; Stebbing, P.; Dunn, A.M. Infection and Invasion: Study Cases from Aquatic Communities. In Wildlife Disease Ecology Linking Theory to Data and Application; Cambridge University Press: Cambridge, UK, 2019; pp. 262–295. [Google Scholar] [CrossRef]
- Dick, J.T.A.; Laverty, C.; Lennon, J.J.; Barrios-O’Neill, D.; Mensink, P.J.; Robert Britton, J.; Médoc, V.; Boets, P.; Alexander, M.E.; Taylor, N.G.; et al. Invader Relative Impact Potential: A New Metric to Understand and Predict the Ecological Impacts of Existing, Emerging and Future Invasive Alien Species. J. Appl. Ecol. 2017, 54, 1259–1267. [Google Scholar] [CrossRef] [Green Version]
- Warren, D.A.; Bradbeer, S.J.; Dunn, A.M. Superior Predatory Ability and Abundance Predicts Potential Ecological Impact towards Early-Stage Anurans by Invasive ‘Killer Shrimp’ (Dikerogammarus villosus). Sci. Rep. 2021, 11, 4570. [Google Scholar] [CrossRef]
- Worrall, F.; Howden, N.J.K.; Burt, T.P.; Hannah, D.M. River Water Temperature Demonstrates Resistance to Long-Term Air Temperature Change. Hydrol. Process. 2022, 36, e14732. [Google Scholar] [CrossRef]
- Laize, C.L.R.; Bruna Meredith, C. Water Temperatures for the Period 1984 to 2007 at 35 Sites on 21 UK Rivers—EIDC; NERC Environmental Information Data Centre: Lancaster, UK, 2015. [Google Scholar] [CrossRef]
- Awachie, J. The Development and Life History of Echinorhynchus truttae Schrank, 1788 (Acanthocephala). J. Helminthol. 1966, 40, 11–32. [Google Scholar] [CrossRef]
- Perrot-Minnot, M.-J.; Gaillard, M.; Dodet, R.; Cézilly, F. Interspecific Differences in Carotenoid Content and Sensitivity to UVB Radiation in Three Acanthocephalan Parasites Exploiting a Common Intermediate Host. Int. J. Parasitol. 2011, 41, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Bloor, M.C. Dietary Preference of Gammarus pulex and Asellus aquaticus during a Laboratory Breeding Programme for Ecotoxicological Studies. Int. J. Zool. 2011, 2011, 294394. [Google Scholar] [CrossRef] [Green Version]
- Waring, R.H.; Running, S.W. Forest Ecosystems: Analysis at Multiple Scales, 3rd ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: San Diego, CA, USA, 2007. [Google Scholar]
- Bjelke, U.; Boberg, J.; Oliva, J.; Tattersdill, K.; Mckie, B.G. Dieback of Riparian Alder Caused by the Phytophthora alni Complex: Projected Consequences for Stream Ecosystems. Freshw. Biol. 2016, 61, 565–579. [Google Scholar] [CrossRef] [Green Version]
- Shokri, M.; Cozzoli, F.; Vignes, F.; Bertoli, M.; Pizzul, E.; Basset, A. Metabolic Rate and Climate Change across Latitudes: Evidence of Mass-Dependent Responses in Aquatic Amphipods. J. Exp. Biol. 2022, 225, jeb244842. [Google Scholar] [CrossRef] [PubMed]
- Kohler, S.A.; Parker, M.O.; Ford, A.T. Shape and Size of the Arenas Affect Amphipod Behaviours: Implications for Ecotoxicology. PeerJ 2018, 2018, e5271. [Google Scholar] [CrossRef] [PubMed]
- Perrot-Minnot, M.J.; Banchetry, L.; Cézilly, F. Anxiety-like Behaviour Increases Safety from Fish Predation in an Amphipod Crustacea. R. Soc. Open Sci. 2017, 4, 171558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowe, C.L.; Hopkins, W.A.; Zehnder, C.; Congdon, J.D. Metabolic Costs Incurred by Crayfish (Procambarus acutus) in a Trace Element-Polluted Habitat: Further Evidence of Similar Responses among Diverse Taxonomic Groups. Comp. Biochem. Physiol. Part C 2001, 129, 275–283. [Google Scholar] [CrossRef]
- Becker, J.; Ortmann, C.; Wetzel, M.A.; Koop, J.H.E. Metabolic Activity and Behavior of the Invasive Amphipod Dikerogammarus villosus and Two Common Central European Gammarid Species (Gammarus fossarum, Gammarus roeselii): Low Metabolic Rates May Favor the Invader. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 191, 119–126. [Google Scholar] [CrossRef]
- Kemp, J.S.; Tang, F.; Aldridge, D.C. Quantifying Invader Impact: Applying Functional Response Metrics to a Rapidly Spreading Non-Native Species. Freshw. Biol. 2018, 63, 1514–1522. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wood, S.N. Fast Stable Restricted Maximum Likelihood and Marginal Likelihood Estimation of Semiparametric Generalized Linear Models. J. R. Stat. Soc. Ser. B Stat. Methodol. 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Therneau, T.M.; A Package for Survival Analysis in R. R Package Version 3.2-11. 2021. Available online: https://CRAN.R-Project.Org/Package=survival (accessed on 16 April 2022).
- Yvon-Durocher, G.; Caffrey, J.M.; Cescatti, A.; Dossena, M.; Del Giorgio, P.; Gasol, J.M.; Montoya, J.M.; Pumpanen, J.; Staehr, P.A.; Trimmer, M.; et al. Reconciling the Temperature Dependence of Respiration across Timescales and Ecosystem Types. Nature 2012, 487, 472–476. [Google Scholar] [CrossRef]
- Soto, I.; Cuthbert, R.N.; Ahmed, D.A.; Kouba, A.; Domisch, S.; Marquez, J.R.G.; Beidas, A.; Amatulli, G.; Kiesel, J.; Shen, L.Q.; et al. Tracking a Killer Shrimp: Dikerogammarus villosus Invasion Dynamics across Europe. Divers. Distrib. 2023, 29, 157–172. [Google Scholar] [CrossRef]
- GB Non-Native Species Secretariat. Killer Shrimp. NNSS. Available online: https://www.nonnativespecies.org/non-native-species/information-portal/view/1219 (accessed on 22 May 2023).
- Laverty, C.; Brenner, D.; McIlwaine, C.; Lennon, J.J.; Dick, J.T.A.; Lucy, F.E.; Christian, K.A. Temperature Rise and Parasitic Infection Interact to Increase the Impact of an Invasive Species. Int. J. Parasitol. 2017, 47, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Wijnhoven, S.; Van Riel, M.C.; Van Der Velde, G.; van Riel, M.C.; van der Velde, G. Exotic and Indigenous Freshwater Gammarid Species: Physiological Tolerance to Water Temperature in Relation to Ionic Content of the Water. Aquat. Ecol. 2003, 37, 151–158. [Google Scholar] [CrossRef]
- Verberk, W.C.E.P.; Leuven, R.S.E.W.; van der Velde, G.; Gabel, F. Thermal Limits in Native and Alien Freshwater Peracarid Crustacea: The Role of Habitat Use and Oxygen Limitation. Funct. Ecol. 2018, 32, 926–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottin, D.; Roussel, D.; Foucreau, N.; Hervant, F.; Piscart, C. Disentangling the Effects of Local and Regional Factors on the Thermal Tolerance of Freshwater Crustaceans. Naturwissenschaften 2012, 99, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, T.; Prevodnik, A.; Sundelin, B. Combined Effects of Temperature and a Pesticide on the Baltic Amphipod Monoporeia affinis. Aquat. Biol. 2008, 1, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Maranhão, P.; Marques, J.C. The Influence of Temperature and Salinity on the Duration of Embryonic Development, Fecundity and Growth of the Amphipod Echinogammarus marinus Leach (Gammaridae). Acta Oecologica 2003, 24, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, P.G.; Loganimoce, E.M.; Neuparth, T.; Rocha, M.J.; Rocha, E.; Arenas, F. Interactive Effects of Increased Temperature, pCO2 and the Synthetic Progestin Levonorgestrel on the Fitness and Breeding of the Amphipod Gammarus locusta. Environ. Pollut. 2018, 236, 937–947. [Google Scholar] [CrossRef]
- Dianne, L.; Bollache, L.; Lagrue, C.; Franceschi, N.; Rigaud, T. Larval Size in Acanthocephalan Parasites: Influence of Intraspecific Competition and Effects on Intermediate Host Behavioural Changes. Parasit. Vectors 2012, 5, 166. [Google Scholar] [CrossRef]
- Kirk, D.; Jones, N.; Peacock, S.; Phillips, J.; Molnár, P.K.; Krkošek, M.; Luijckx, P. Empirical Evidence That Metabolic Theory Describes the Temperature Dependency of Within-Host Parasite Dynamics. PLoS Biol. 2018, 16, e2004608. [Google Scholar] [CrossRef] [Green Version]
- Dickey, J.W.E.; Cuthbert, R.N.; Rea, M.; Laverty, C.; Crane, K.; South, J.; Briski, E.; Chang, X.; Coughlan, N.E.; MacIsaac, H.J.; et al. Assessing the Relative Potential Ecological Impacts and Invasion Risks of Emerging and Future Invasive Alien Species. NeoBiota 2018, 40, 1–24. [Google Scholar] [CrossRef]
- Dickey, J.W.E.; Cuthbert, R.N.; South, J.; Britton, J.R.; Caffrey, J.; Chang, X.; Crane, K.; Coughlan, N.E.; Fadaei, E.; Farnsworth, K.D.; et al. On the RIP: Using Relative Impact Potential to Assess the Ecological Impacts of Invasive Alien Species. NeoBiota 2020, 55, 27–60. [Google Scholar] [CrossRef] [Green Version]
- Platvoet, D.; van der Velde, G.; Dick, J.; Li, S. Flexible Omnivory in Dikerogammarus villosus (Sowinsky, 1894) (Amphipoda). Crustaceana 2009, 82, 703–720. [Google Scholar] [CrossRef] [Green Version]
- Mancini, F.; De Giorgi, R.; Ludovisi, A.; Vizzini, S.; Mancinelli, G. Ontogenetic Shift in the Trophic Role of the Invasive Killer Shrimp Dikerogammarus villosus: A Stable Isotope Study. Biol. Invasions 2021, 23, 1803–1817. [Google Scholar] [CrossRef]
- Doherty-Bone, T.M.; Dunn, A.M.; Liddell, C.; Brown, L.E. Transformation of Detritus by a European Native and Two Invasive Alien Freshwater Decapods. Biol. Invasions 2018, 20, 1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamczuk, M.; Ferencz, B.; Mieczan, T.; Dawidek, J. Allochthonous Subsidies as Driving Forces for Development of Plankton in an Autotrophic, Temperate, and Small Lake. Hydrobiology 2019, 846, 59–73. [Google Scholar] [CrossRef] [Green Version]
- Cummins, K.W.; Klug, M.J. Feeding Ecology of Stream Invertebrates. Annu. Rev. Ecol. Syst. 1979, 10, 147–172. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amphipod Treatment | Temperature (°C) | Mean Shredding Rate (g Leaf Eaten Amphipod g−1 day−1) | St. Dev. Shredding Rate |
|---|---|---|---|
| G. pulex unparasitised | 5 | 0.0263 | 0.0083 |
| G. pulex infected with E. truttae | 5 | 0.0301 | 0.0153 |
| D. villosus | 5 | 0.0093 | 0.0033 |
| G. pulex unparasitised | 10 | 0.0337 | 0.0127 |
| G. pulex infected with E. truttae | 10 | 0.0498 | 0.0151 |
| D. villosus | 10 | 0.016 | 0.0059 |
| G. pulex unparasitised | 15 | 0.0479 | 0.0233 |
| G. pulex infected with E. truttae | 15 | 0.0644 | 0.0793 |
| D. villosus | 15 | 0.0343 | 0.0111 |
| G. pulex unparasitised | 20 | 0.0548 | 0.0149 |
| G. pulex infected with E. truttae | 20 | 0.063 | 0.0636 |
| D. villosus | 20 | 0.0327 | 0.015 |
| G. pulex unparasitised | 25 | 0.0247 | 0.0128 |
| G. pulex infected with E. truttae | 25 | 0.0626 | 0.023 |
| D. villosus | 25 | 0.0325 | 0.0182 |
| G. pulex unparasitised | 30 | 0.036 | 0.0072 |
| G. pulex infected with E. truttae | 30 | 0.135 | 0.116 |
| D. villosus | 30 | 0.021 | 0.008 |
| Amphipod Treatment | Intercept ± 1 SE | Multiplier 95% CI | p-Value | R2 |
|---|---|---|---|---|
| G. pulex unparasitised | −3.16 ± 0.10 | 0.39 (0.17, 0.60) | <0.001 | 0.24 |
| G. pulex parasitised | −2.99 ± 0.12 | 0.36 (0.09, 0.63) | <0.001 | 0.14 |
| D. villosus | −3.47 ± 0.09 | 0.90 (0.72, 1.09) | <0.001 | 0.69 |
| Amphipod Treatment | Temperature (°C) | Mean Metabolic Rate (mg/L Oxygen Consumed Hour−1 Amphipod Gram−1) | St. Dev. Metabolic Rate |
|---|---|---|---|
| G. pulex unparasitised | 10 | 51.2 | 20.5 |
| G. pulex infected with E. truttae | 10 | 56.2 | 18.1 |
| D. villosus | 10 | 31.2 | 16.9 |
| G. pulex unparasitised | 20 | 78.6 | 15.9 |
| G. pulex infected with E. truttae | 20 | 74.4 | 23.0 |
| D. villosus | 20 | 78.1 | 17.2 |
| G. pulex unparasitised | 25 | 73.8 | 21.3 |
| G. pulex infected with E. truttae | 25 | 78.3 | 22.5 |
| D. villosus | 25 | 46.1 | 21.7 |
| Temperature (°C) | Mean RIP | 95% Confidence Interval | % Probability RIP > 1 | % Probability RIP > 10 | |
|---|---|---|---|---|---|
| Lower Limit | Upper Limit | ||||
| 5 | 4.82 | 0.08 | 29.83 | 61.0 | 10.8 |
| 10 | 6.80 | 0.10 | 42.31 | 68.6 | 15.4 |
| 15 | 11.07 | 0.16 | 69.42 | 78.2 | 23.9 |
| 20 | 8.01 | 0.12 | 49.92 | 72.3 | 18.1 |
| 25 | 20.85 | 0.24 | 133.94 | 86.0 | 36.3 |
| 30 | 7.56 | 0.13 | 46.49 | 72.3 | 17.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pile, B.; Warren, D.; Hassall, C.; Brown, L.E.; Dunn, A.M. Biological Invasions Affect Resource Processing in Aquatic Ecosystems: The Invasive Amphipod Dikerogammarus villosus Impacts Detritus Processing through High Abundance Rather than Differential Response to Temperature. Biology 2023, 12, 830. https://doi.org/10.3390/biology12060830
Pile B, Warren D, Hassall C, Brown LE, Dunn AM. Biological Invasions Affect Resource Processing in Aquatic Ecosystems: The Invasive Amphipod Dikerogammarus villosus Impacts Detritus Processing through High Abundance Rather than Differential Response to Temperature. Biology. 2023; 12(6):830. https://doi.org/10.3390/biology12060830
Chicago/Turabian StylePile, Benjamin, Daniel Warren, Christopher Hassall, Lee E. Brown, and Alison M. Dunn. 2023. "Biological Invasions Affect Resource Processing in Aquatic Ecosystems: The Invasive Amphipod Dikerogammarus villosus Impacts Detritus Processing through High Abundance Rather than Differential Response to Temperature" Biology 12, no. 6: 830. https://doi.org/10.3390/biology12060830
APA StylePile, B., Warren, D., Hassall, C., Brown, L. E., & Dunn, A. M. (2023). Biological Invasions Affect Resource Processing in Aquatic Ecosystems: The Invasive Amphipod Dikerogammarus villosus Impacts Detritus Processing through High Abundance Rather than Differential Response to Temperature. Biology, 12(6), 830. https://doi.org/10.3390/biology12060830