
Cumulative Negative Impacts of Invasive Alien Species on Marine Ecosystems of the Aegean Sea

 , , and
, , and 
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
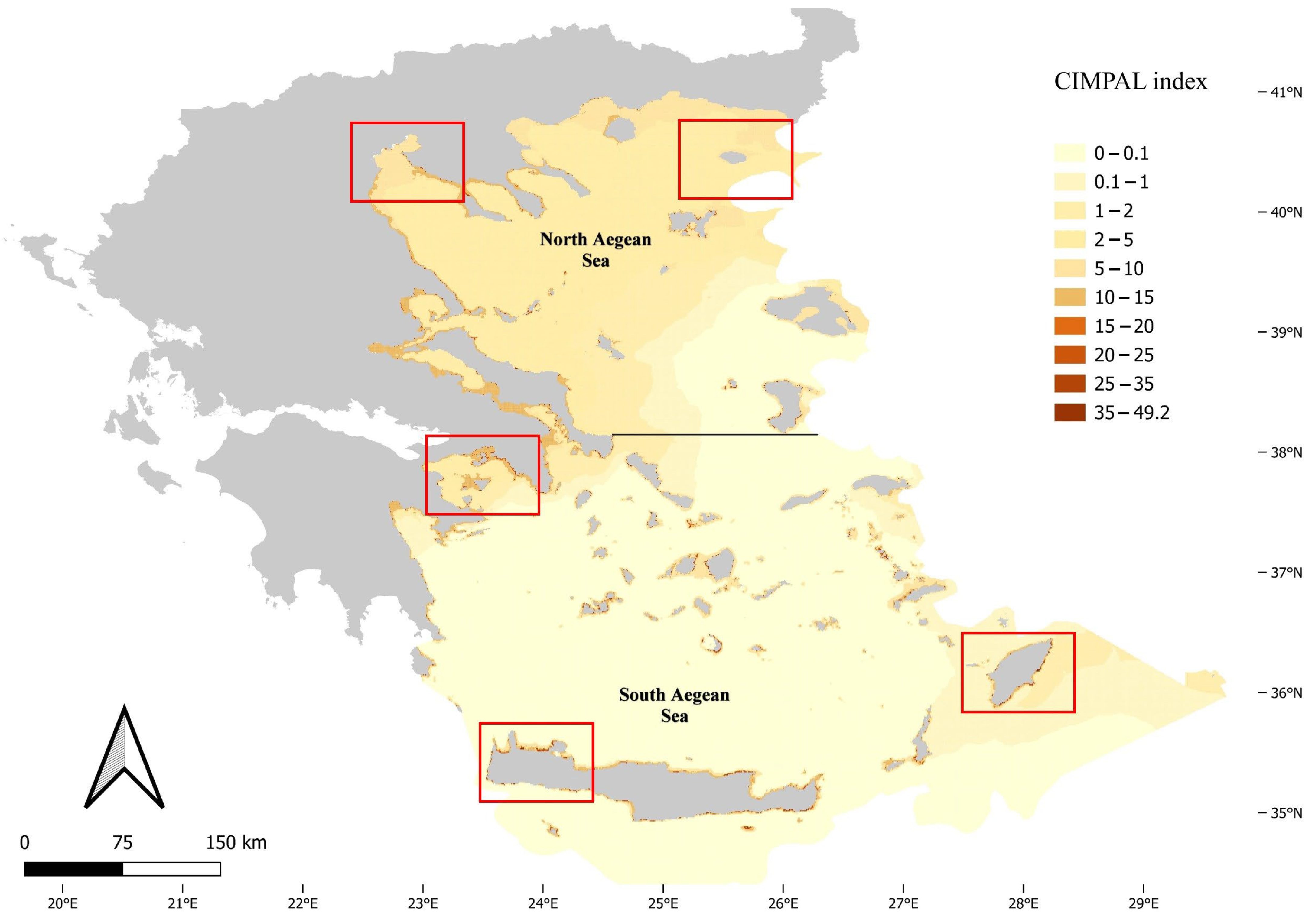
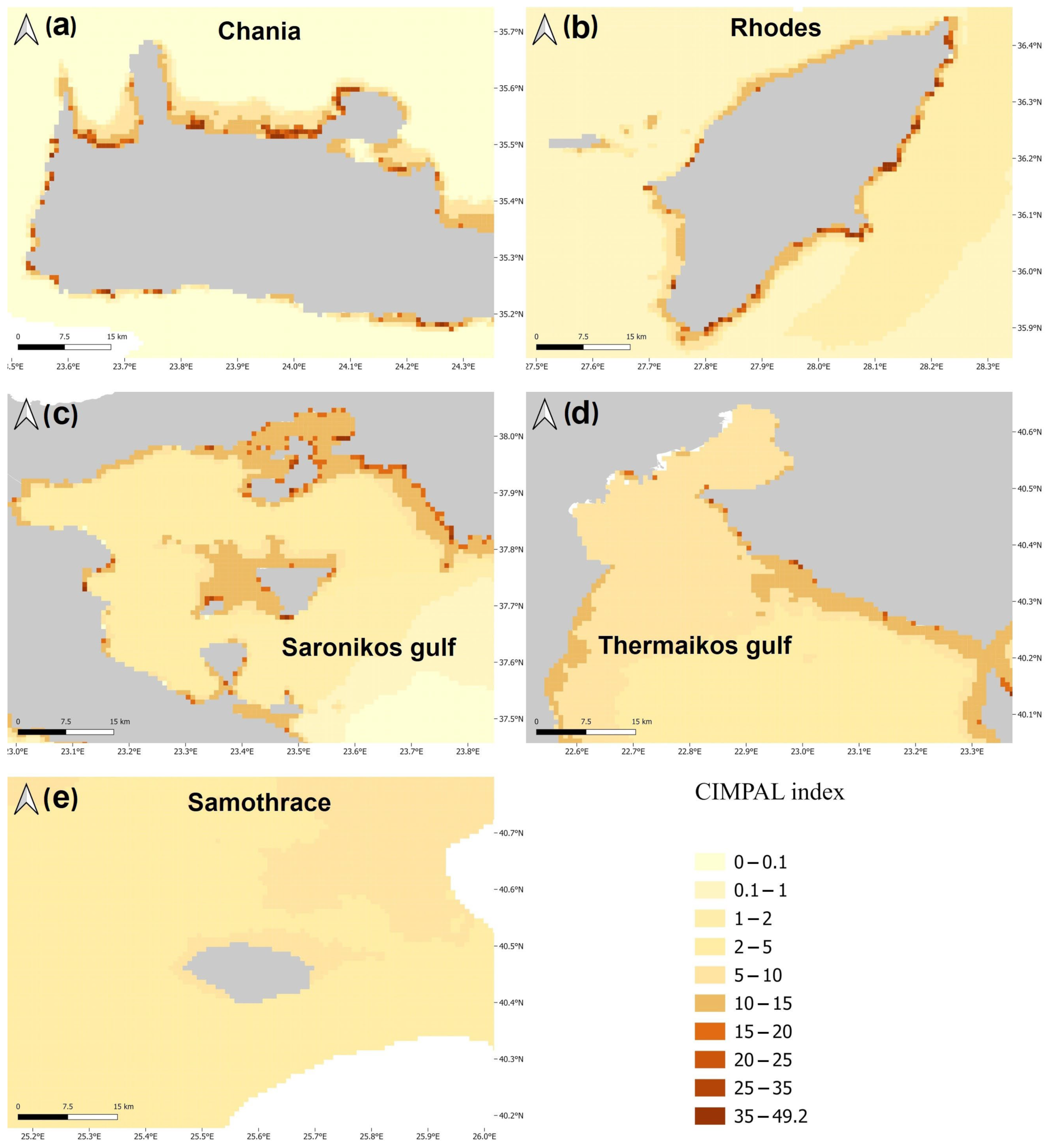
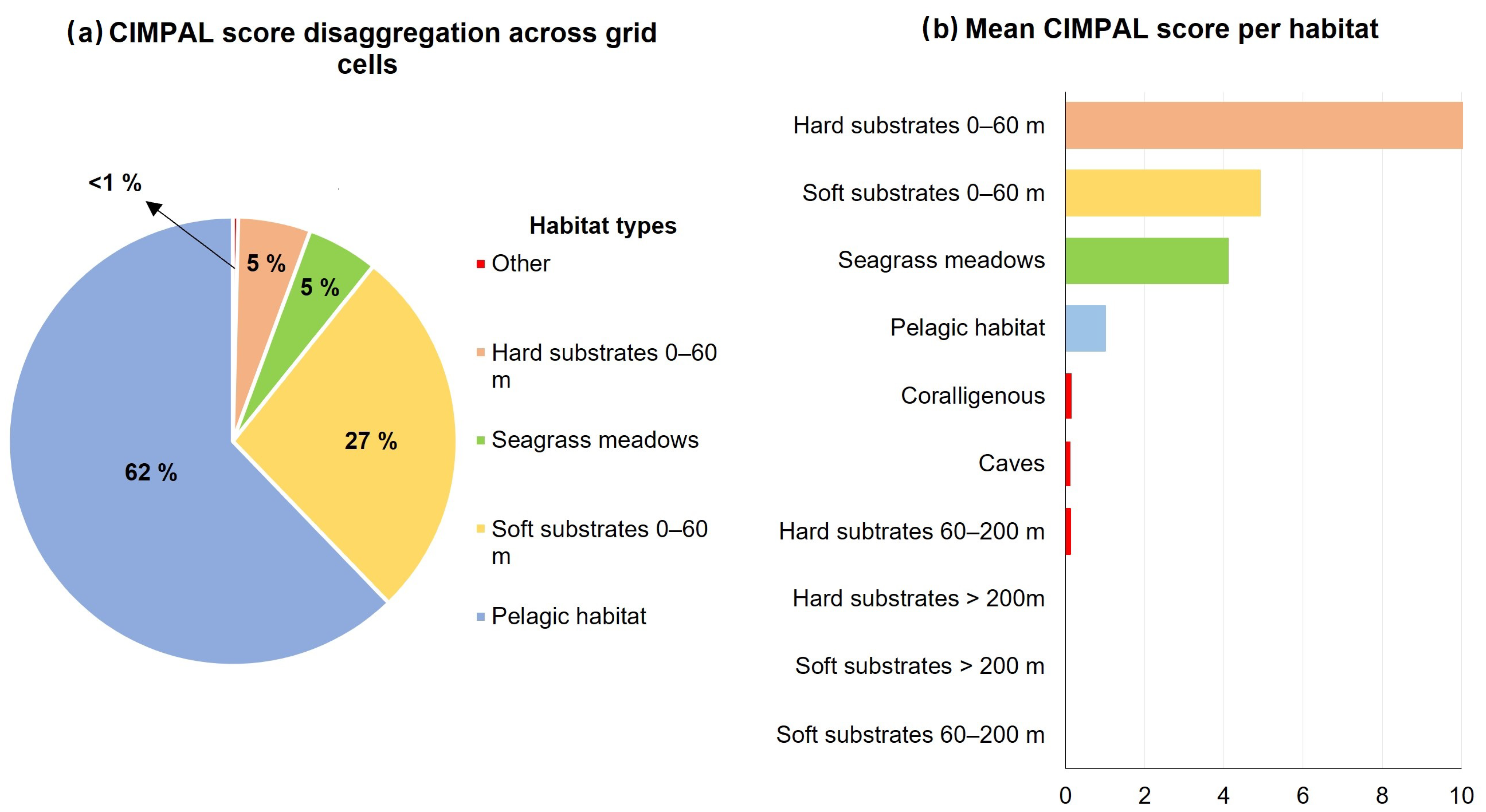
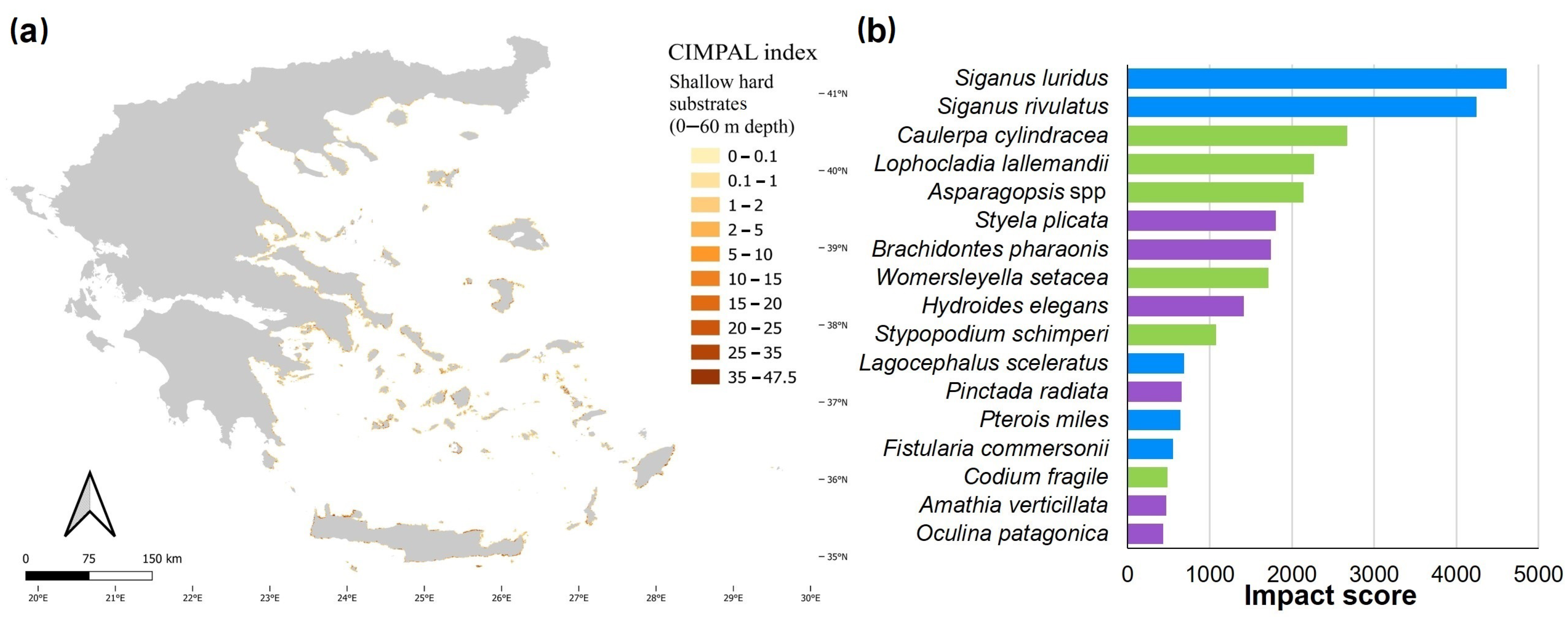
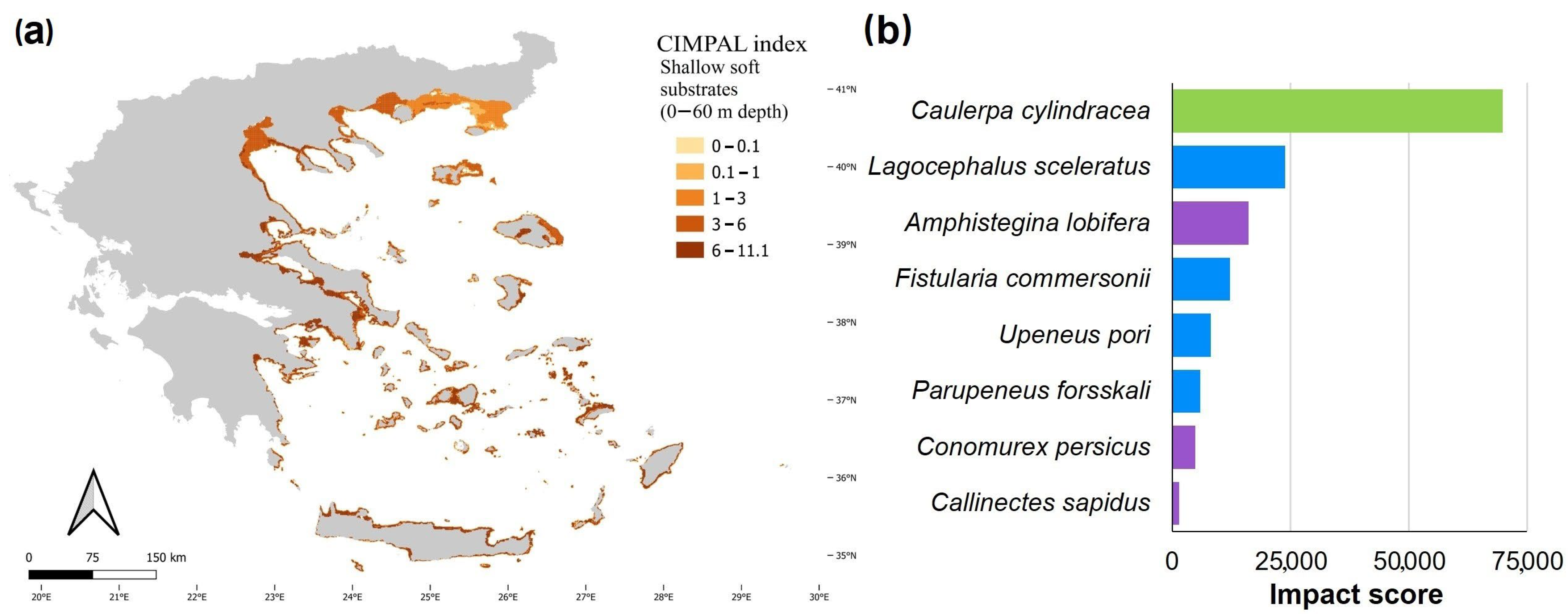
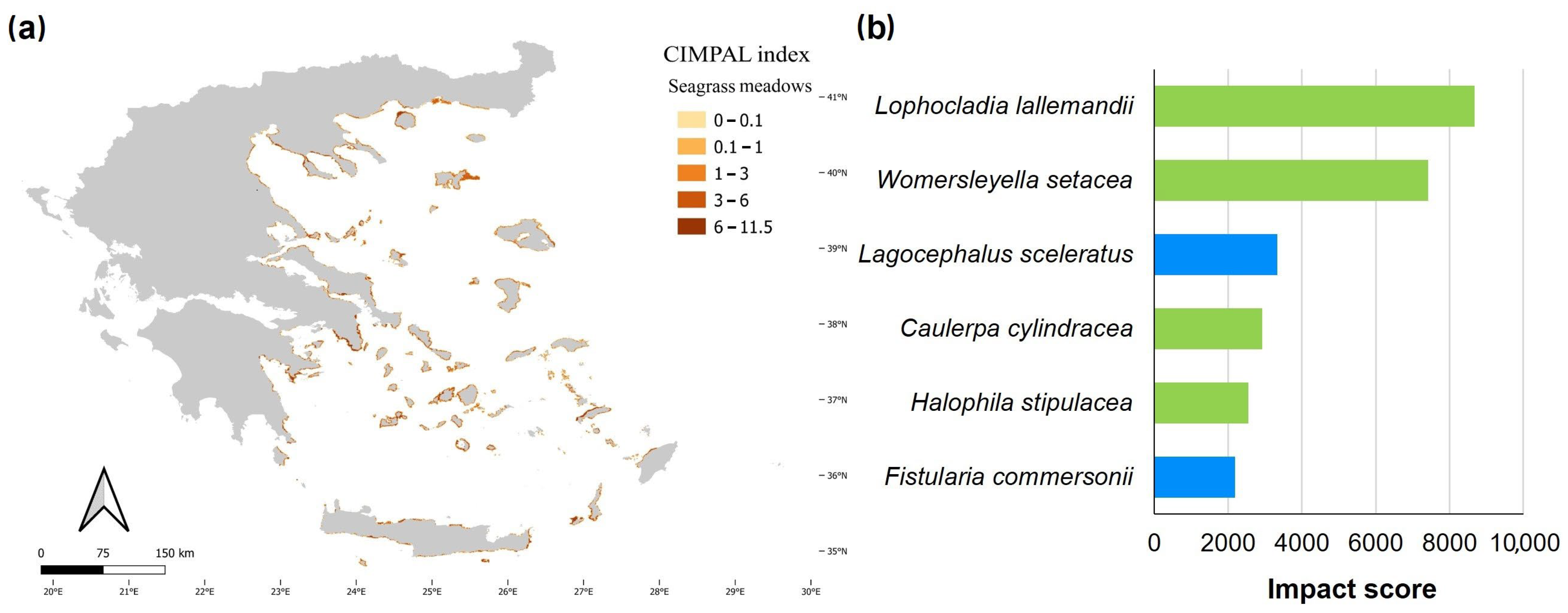
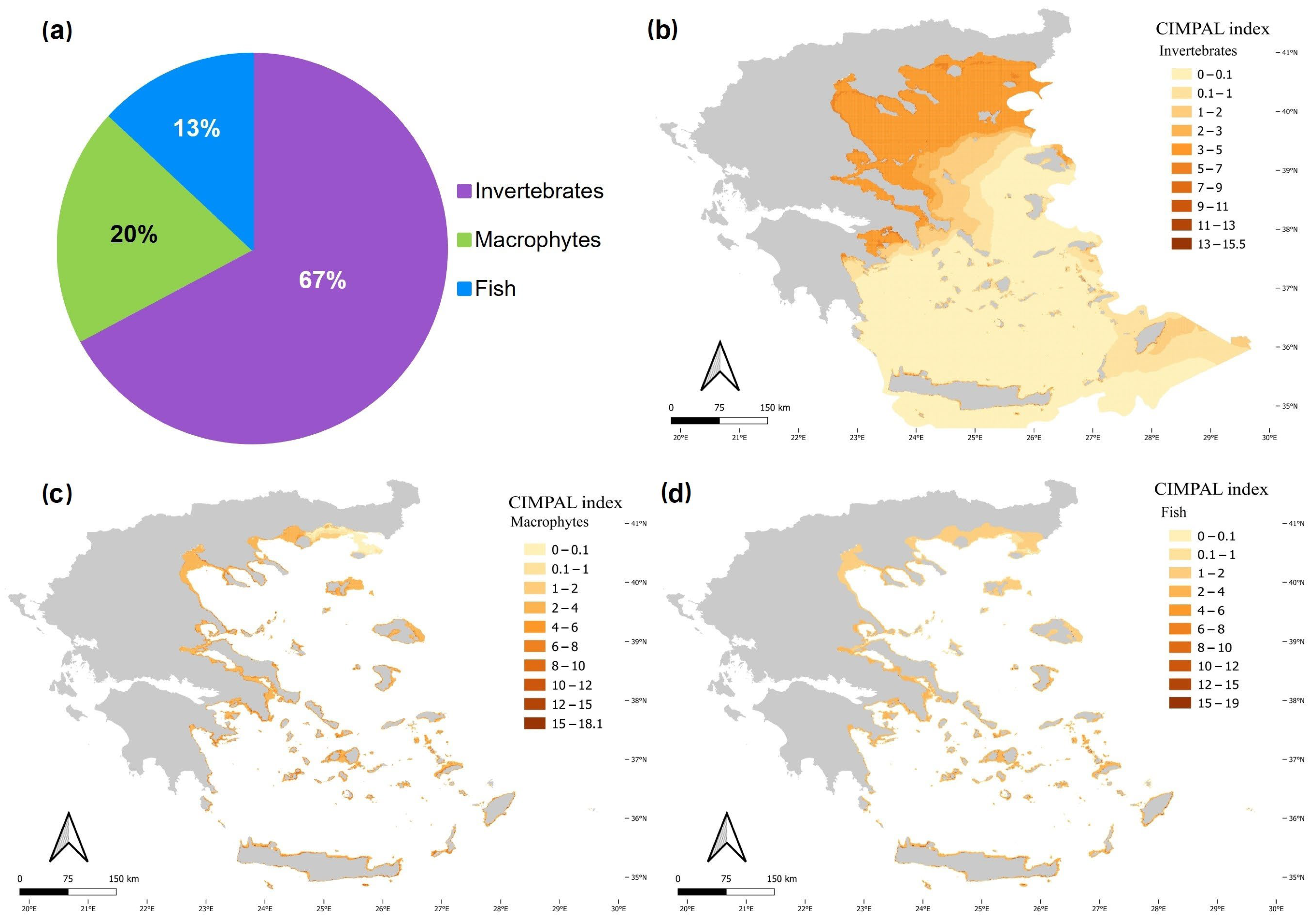
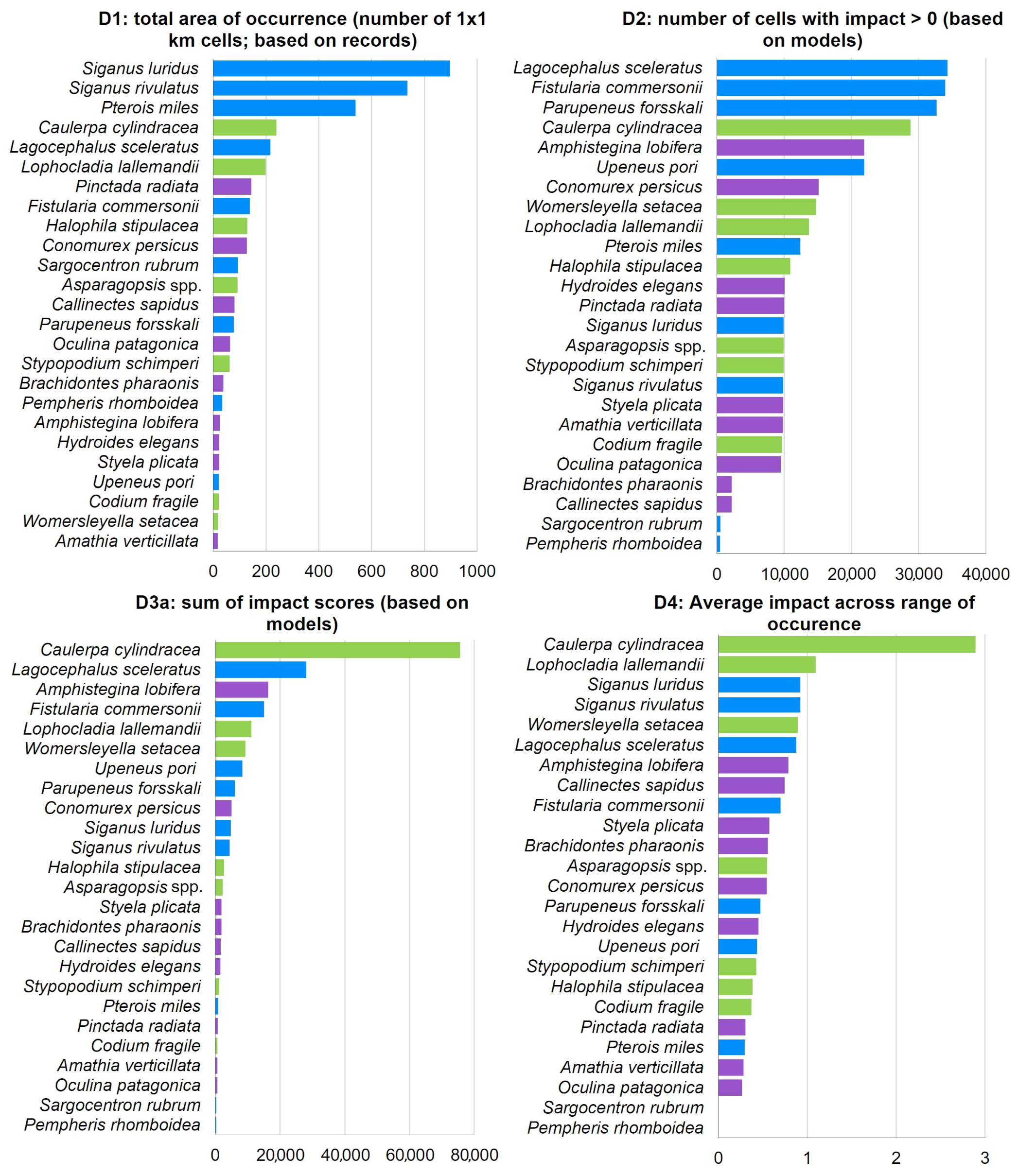
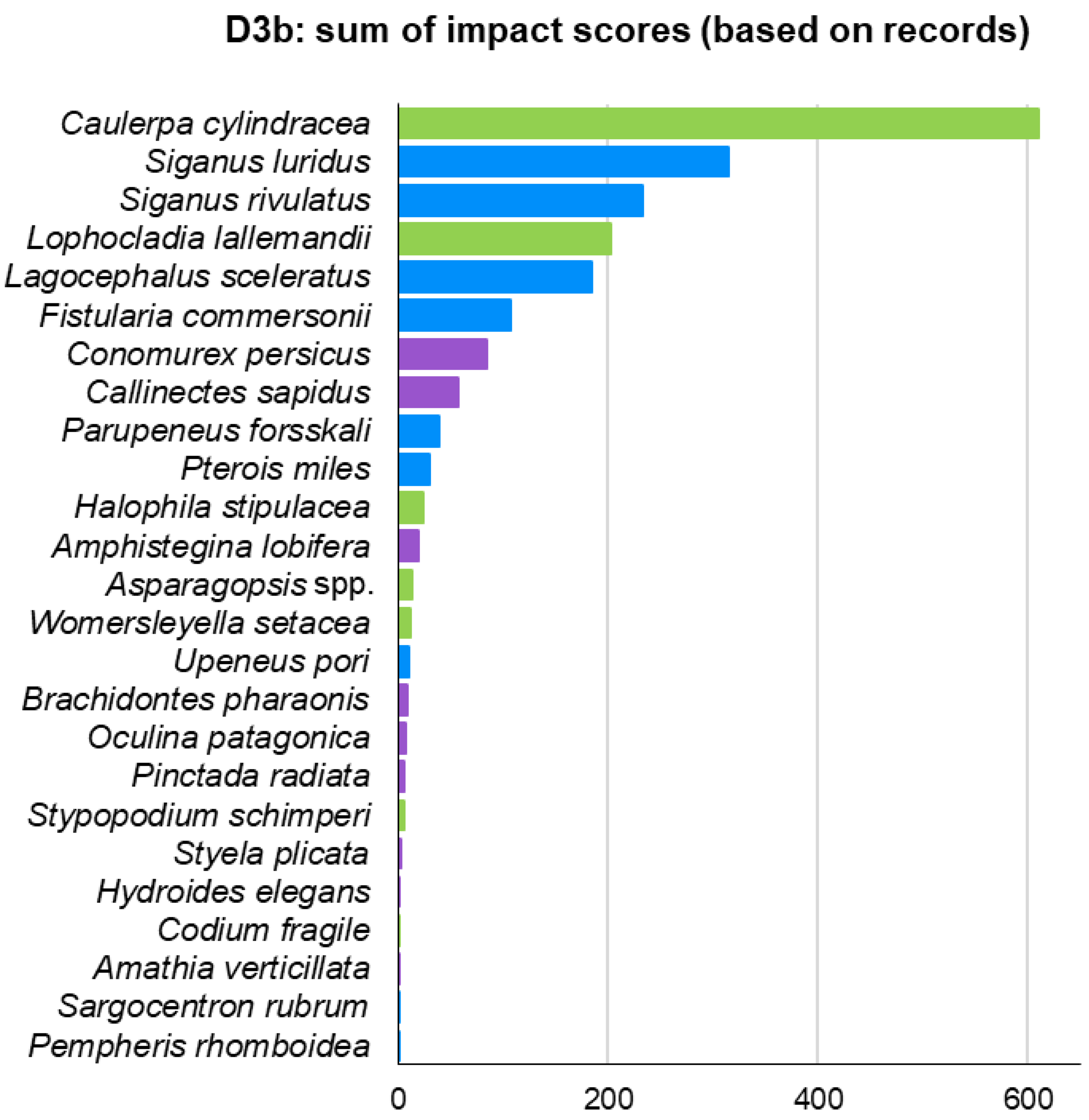
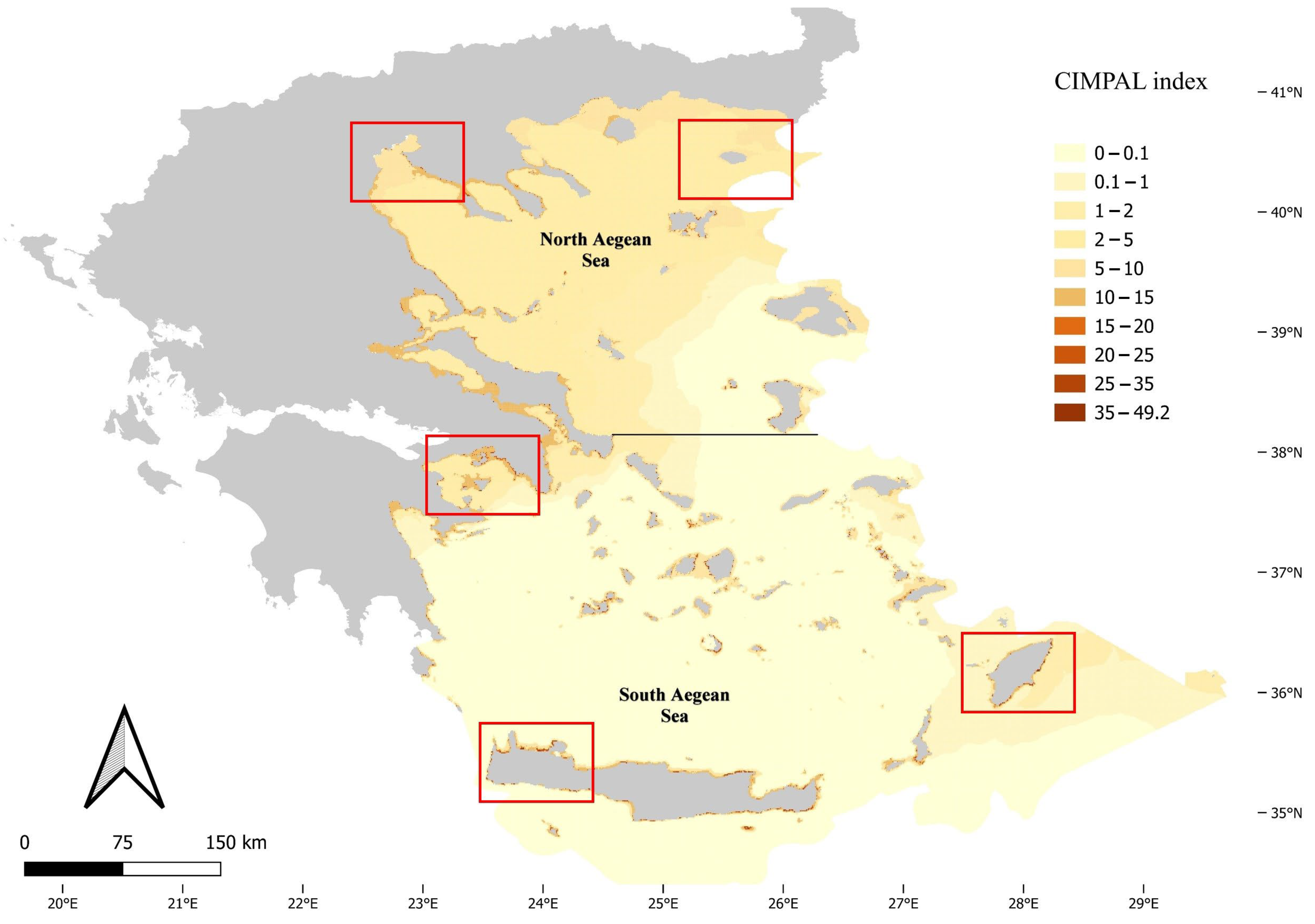
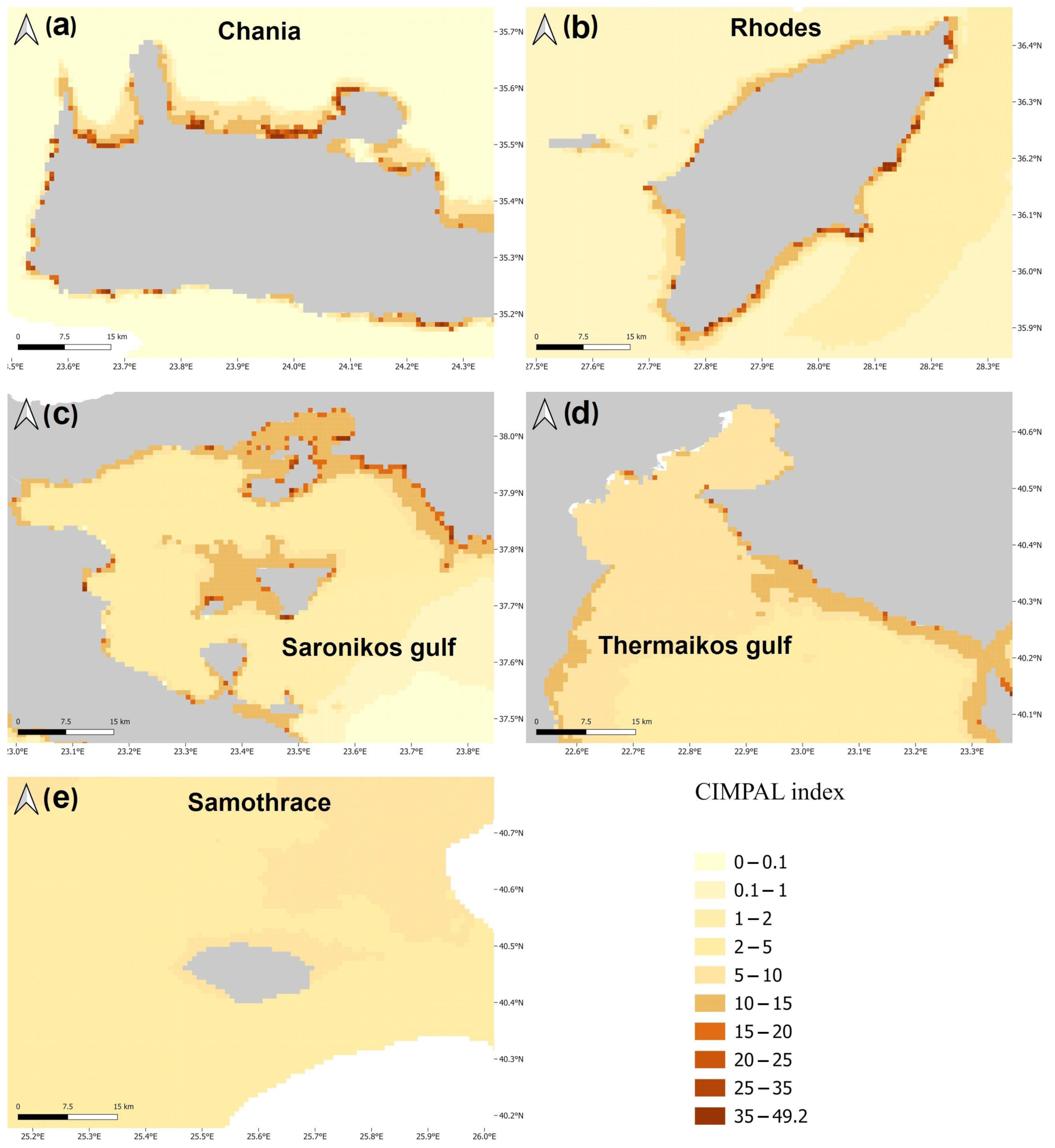
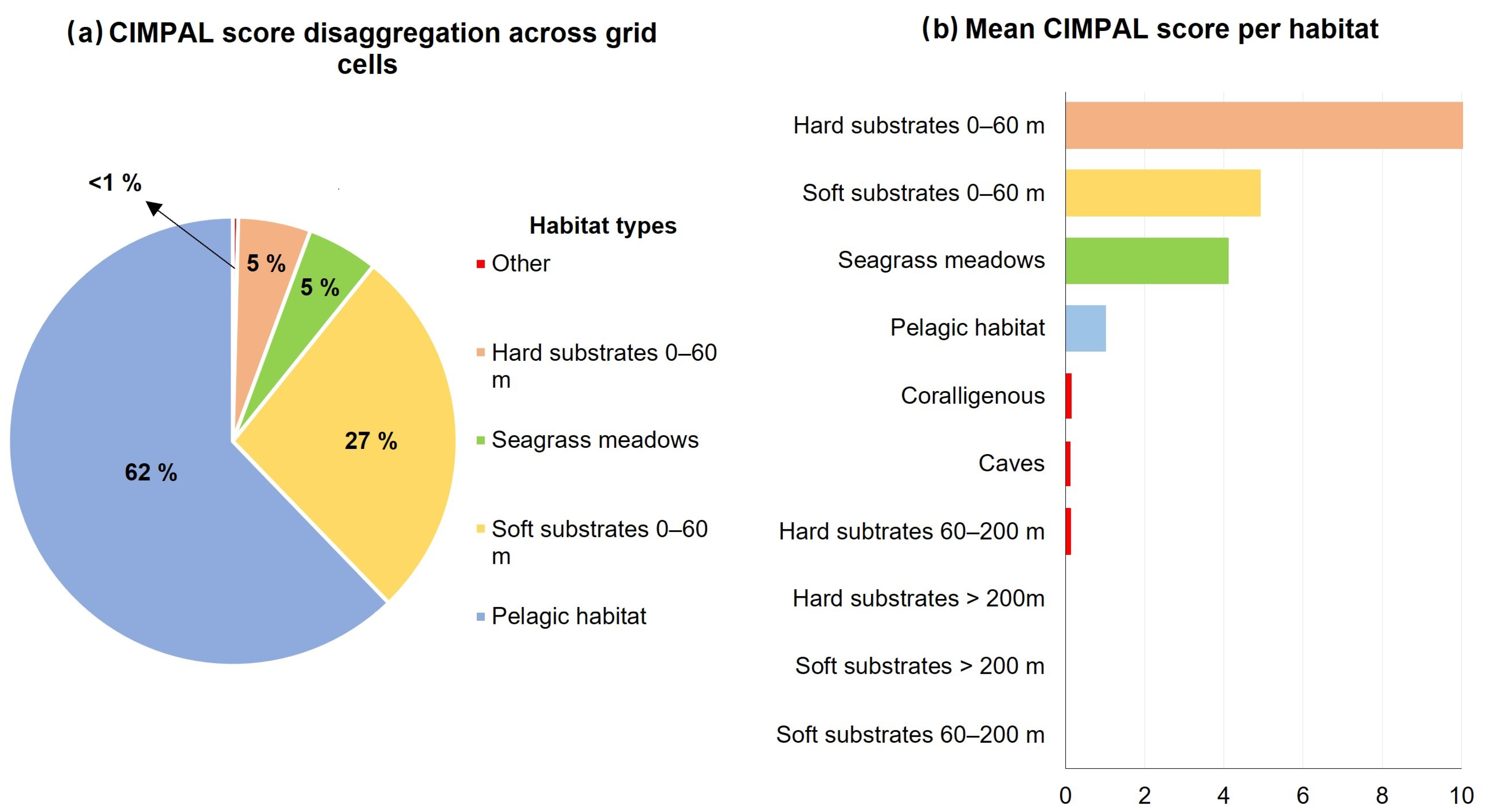
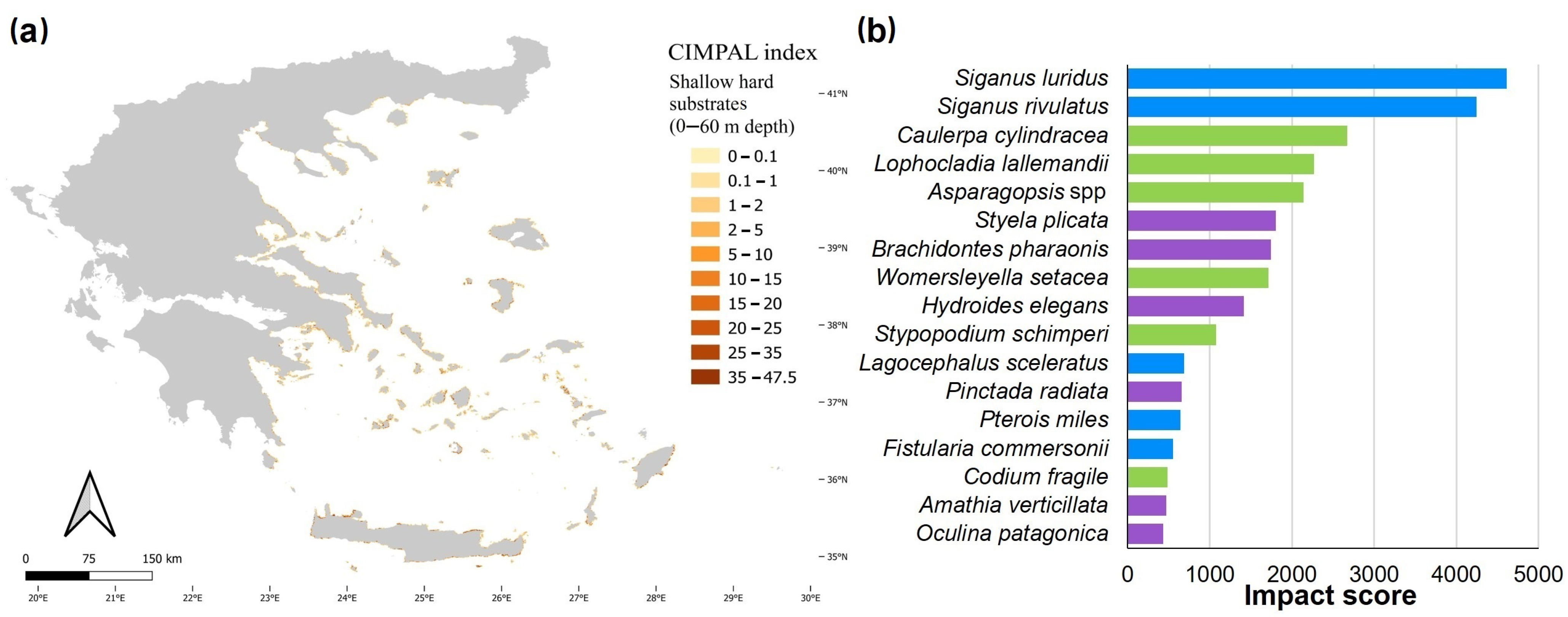
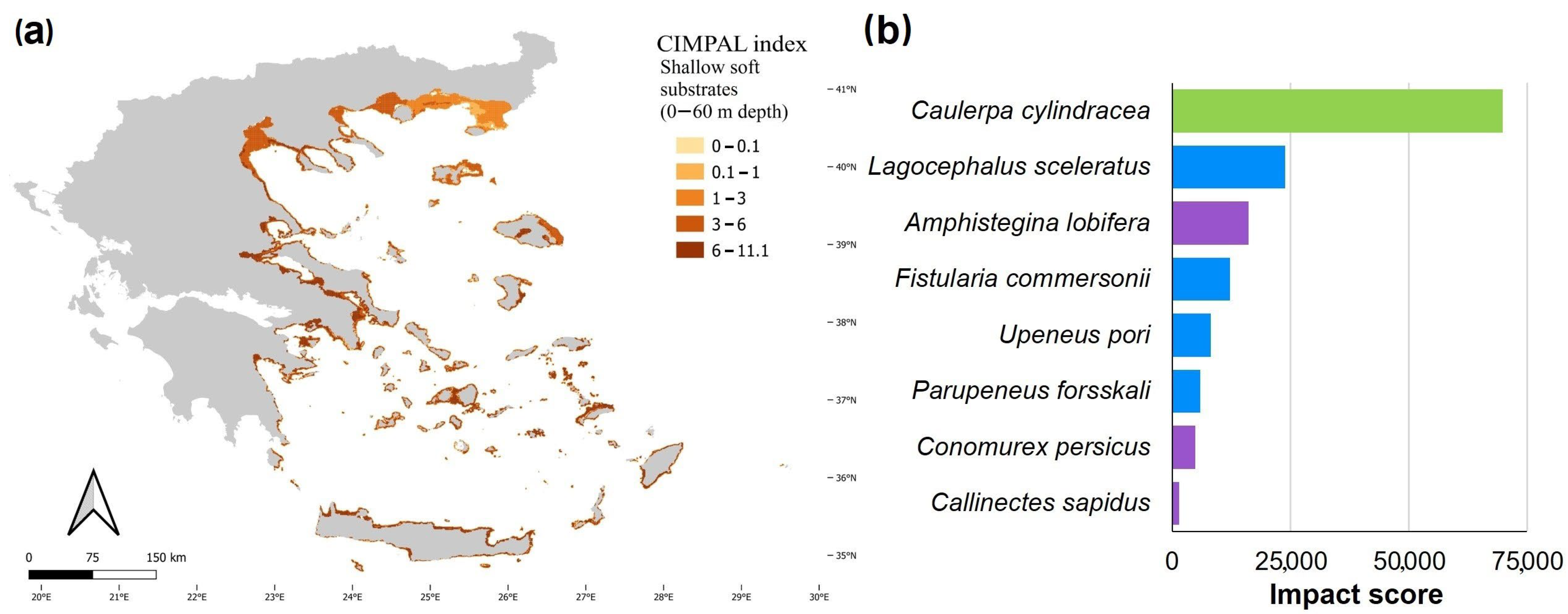
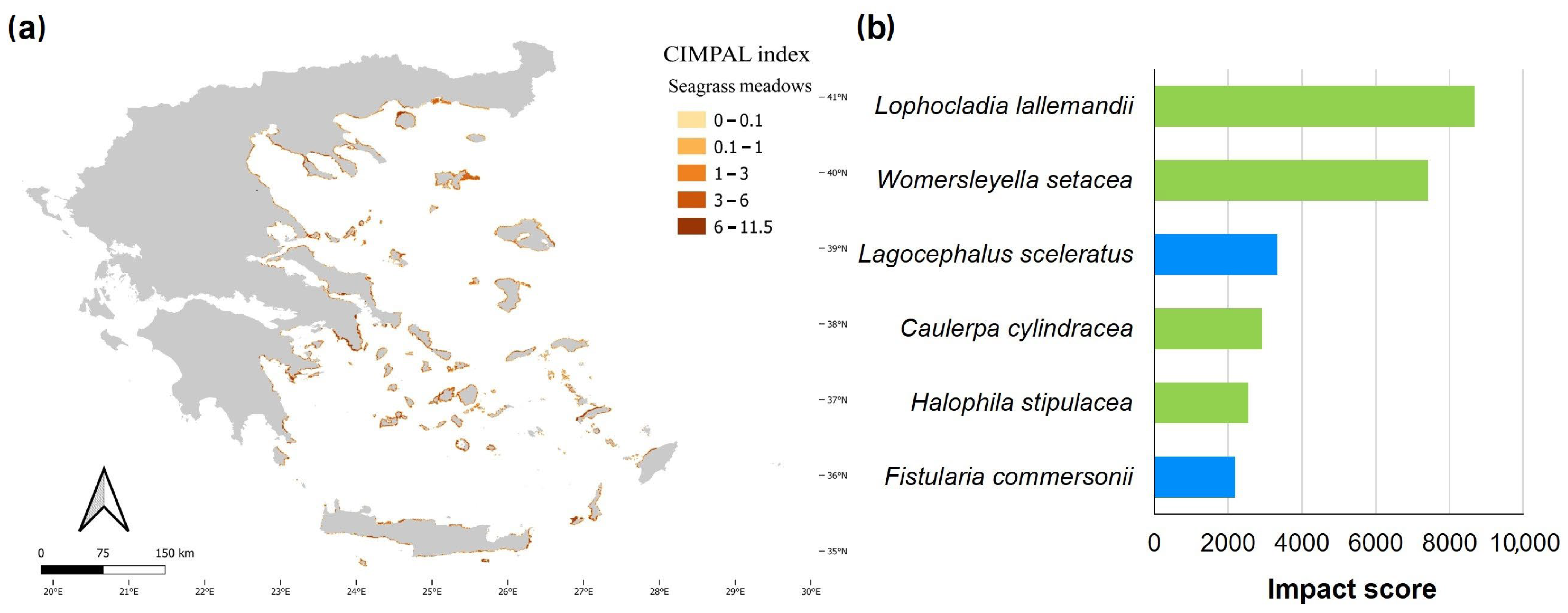
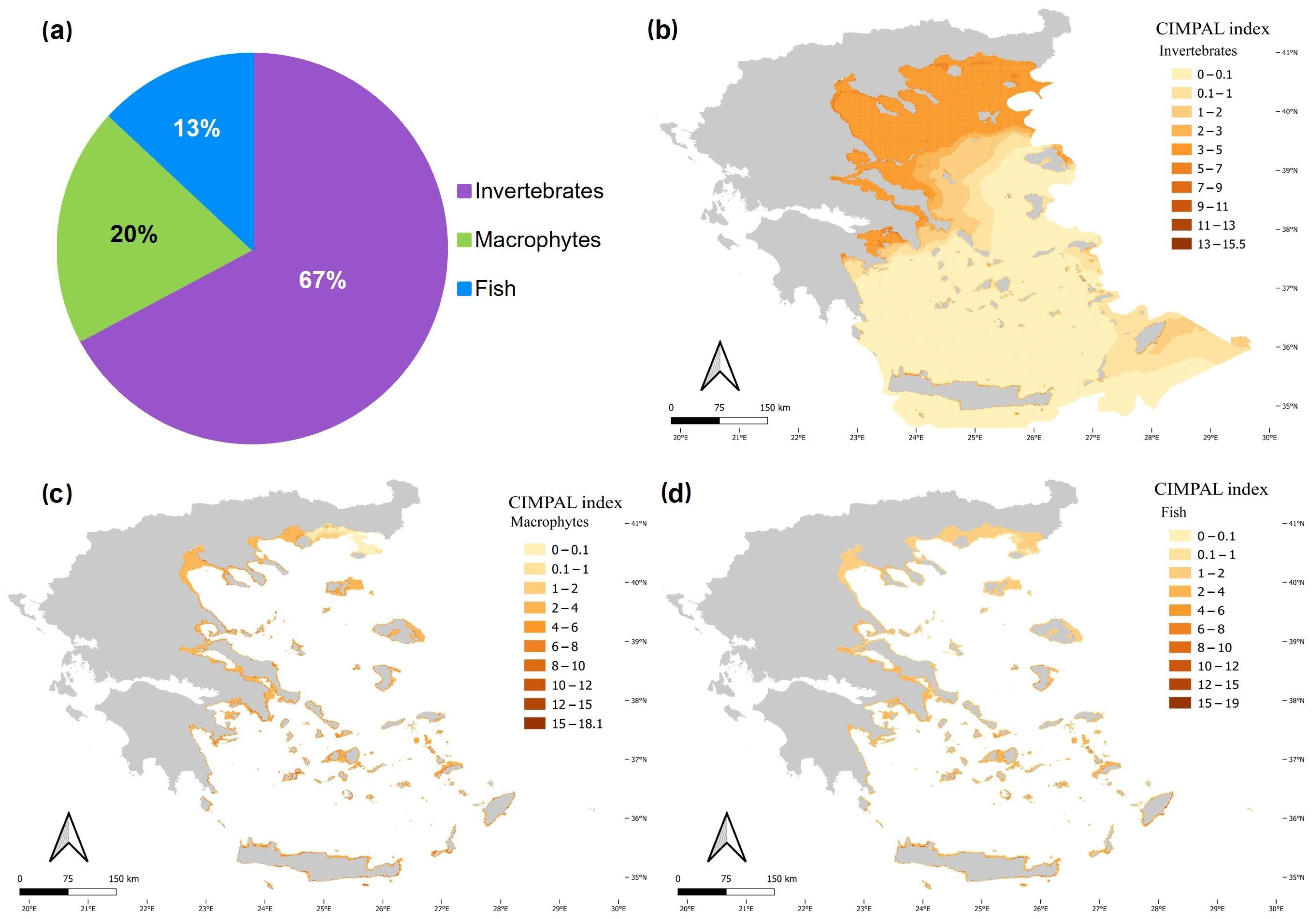
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doney, S.C.; Ruckelshaus, M.; Emmett Duffy, J.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N.; et al. Climate change impacts on marine ecosystems. Annu. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raven, J.; Caldeira, K.; Elderfield, H.; Hoegh-Guldberg, O.; Liss, P.; Riebesell, U.; Shepherd, J.; Turley, C.; Watson, A. Ocean Acidification Due to Increasing Atmospheric Carbon Dioxide; The Royal Society: London, UK, 2005; p. 68. [Google Scholar]
- IPCC. Summary for policymakers. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.O., Roberts, D.C., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2019; pp. 3–35. [Google Scholar] [CrossRef]
- Murphy, G.E.; Romanuk, T.N. A meta-analysis of declines in local species richness from human disturbances. Ecol. Evol. 2014, 4, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Seebens, H.; Bacher, S.; Blackburn, T.M.; Capinha, C.; Dawson, W.; Dullinger, S.; Genovesi, P.; Hulme, P.E.; Kleunen, M.; Kühn, I.; et al. Projecting the continental accumulation of alien species through to 2050. Glob. Change Biol. 2021, 27, 970–982. [Google Scholar] [CrossRef]
- Essl, F.; Lenzner, B.; Bacher, S.; Bailey, S.; Capinha, C.; Daehler, C.; Dullinger, S.; Genovesi, P.; Hui, C.; Hulme, P.E.; et al. Drivers of future alien species impacts: An expert-based assessment. Glob. Change Biol. 2020, 26, 4880–4893. [Google Scholar] [CrossRef]
- Lewis, S.L.; Maslin, M.A. Defining the Anthropocene. Nature 2015, 519, 171–180. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, T.M.; Essl, F.; Evans, T.; Hulme, P.E.; Jeschke, J.M.; Kühn, I.; Kumschick, S.; Marková, Z.; Mrugała, A.; Nentwig, W.; et al. A Unified Classification of Alien Species Based on the Magnitude of their Environmental Impacts. PLoS Biol. 2014, 12, e1001850. [Google Scholar] [CrossRef] [Green Version]
- Tsirintanis, K.; Azzurro, E.; Crocetta, F.; Dimiza, M.; Froglia, C.; Gerovasileiou, V.; Langeneck, J.; Mancinelli, G.; Rosso, A.; Stern, N.; et al. Bioinvasion impacts on biodiversity, ecosystem services, and human health in the Mediterranean Sea. Aquat. Invasions 2022, 17, 308–352. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Tempera, F.; Teixeira, H. Mapping the impact of alien species on marine ecosystems: The Mediterranean Sea case study. Divers. Distrib. 2016, 22, 694–707. [Google Scholar] [CrossRef] [Green Version]
- Strayer, D.L.; Eviner, V.T.; Jeschke, J.M.; Pace, M.L. Understanding the long-term effects of species invasions. Trends Ecol. Evol. 2006, 21, 645–651. [Google Scholar] [CrossRef]
- Vergés, A.; Tomas, F.; Cebrian, E.; Ballesteros, E.; Kizilkaya, Z.; Dendrinos, P.; Karamanlidis, A.A.; Spiegel, D.; Sala, E. Tropical rabbitfish and the deforestation of a warming temperate sea. J. Ecol. 2014, 102, 1518–1527. [Google Scholar] [CrossRef]
- Hulme, P.E. Climate change and biological invasions: Evidence, expectations, and response options. Biol. Rev. 2017, 92, 1297–1313. [Google Scholar] [CrossRef]
- Gissi, E.; Manea, E.; Mazaris, A.D.; Fraschetti, S.; Almpanidou, V.; Bevilacqua, S.; Coll, M.; Guarnieri, G.; Lloret-Lloret, E.; Pascual, M.; et al. A review of the combined effects of climate change and other human stressors on the marine environment. Sci. Total Environ. 2021, 755, 142564. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Moustakas, A. Uncertainty in Marine Invasion Science. Front. Mar. Sci. 2018, 5, 38. [Google Scholar] [CrossRef] [Green Version]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of invasive alien marine species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef] [Green Version]
- Halpern, B.; Frazier, M.; Potapenko, J.; Casey, K.S.; Koenig, K.; Longo, C.; Lowndes, J.S.; Rockwood, R.S.; Selig, E.R.; Selkoe, K.A.; et al. Spatial and temporal changes in cumulative human impacts on the world’s ocean. Nat. Commun. 2015, 6, 7615. [Google Scholar] [CrossRef] [Green Version]
- Ban, N.C.; Alidina, H.M.; Ardron, J.A. Cumulative impact mapping: Advances, relevance and limitations to marine management and conservation, using Canada’s Pacific waters as a case study. Mar. Policy 2010, 34, 876–886. [Google Scholar] [CrossRef]
- Halpern, B.S.; McLeod, K.L.; Rosenberg, A.A.; Crowder, L.B. Managing for cumulative impacts in ecosystem—Based management through ocean zoning. Ocean Coast. Manag. 2008, 51, 203–211. [Google Scholar] [CrossRef]
- Halpern, B.S.; Fujita, R. Assumptions, challenges, and future directions in cumulative impact analysis. Ecosphere 2013, 4, 131. [Google Scholar] [CrossRef]
- Micheli, F.; Halpern, B.S.; Walbridge, S.; Ciriaco, S.; Ferretti, F.; Fraschetti, S.; Lewison, R.; Nykjaer, L.; Rosenberg, A.A. Cumulative human impacts on Mediterranean and Black Sea marine ecosystems: Assessing current pressures and opportunities. PLoS ONE 2013, 8, e79889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, M.J.; Dekeyzer, D.; Galil, B.S.; Hutchings, P.; Katsanevakis, S.; Pagad, S.; Robinson, T.B.; Turon, X.; Vandepitte, L.; Vanhoorne, B.; et al. Introducing the World Register of Introduced Marine Species (WRiMS). Manag. Biol. Invasion 2021, 12, 792–811. [Google Scholar] [CrossRef]
- Zenetos, A.; Albano, P.G.; López Garcia, E.; Stern, N.; Tsiamis, K.; Galanidi, M. Established non-indigenous species increased by 40% in 11 years in the Mediterranean Sea. Mediterr. Mar. Sci. 2022, 23, 196–212. [Google Scholar] [CrossRef]
- Zenetos, A.; Albano, P.G.; López Garcia, E.; Stern, N.; Tsiamis, K.; Galanidi, M. Corrigendum to the Review Article (Medit. Mar. Sci. 23/1 2022, 196–212). Mediterr. Mar. Sci. 2022, 23, 876–878. [Google Scholar] [CrossRef]
- Galanidi, M.; Zenetos, A. Data-Driven Recommendations for Establishing Threshold Values for the NIS Trend Indicator in the Mediterranean Sea. Diversity 2022, 14, 57. [Google Scholar] [CrossRef]
- Carlton, J.T. Biological invasions and cryptogenic species. Ecology 1996, 77, 1653–1655. [Google Scholar] [CrossRef]
- Essl, F.; Bacher, S.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Katsanevakis, S.; Kowarik, I.; Kühn, I.; Pyšek, P.; Rabitsch, W.; et al. Which taxa are alien? Criteria, applications, and uncertainties. BioScience 2018, 68, 496–509. [Google Scholar] [CrossRef] [Green Version]
- Gerovasileiou, V.; Bancila, R.I.; Katsanevakis, S.; Zenetos, A. Introduced species in Mediterranean marine caves: An increasing but neglected threat. Mediterr. Mar. Sci. 2022, 23, 995–1005. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Coll, M.; Piroddi, C.; Steenbeek, J.; Ben Rais Lasram, F.; Zenetos, A.; Cardoso, A.C. Invading the Mediterranean Sea: Biodiversity patterns shaped by human activities. Front. Mar. Sci. 2014, 1, 32. [Google Scholar] [CrossRef] [Green Version]
- Rilov, G.; Peleg, O.; Yeruham, E.; Garval, T.; Vichik, A.; Raveh, O. Alien turf: Overfishing, overgrazing and invader domination in south-eastern Levant reef ecosystems. Aquat. Conserv. 2018, 28, 351–369. [Google Scholar] [CrossRef]
- Yeruham, E.; Shpigel, M.; Abelson, A.; Rilov, G. Ocean warming and tropical invaders erode the performance of a key herbivore. Ecology 2020, 101, e02925. [Google Scholar] [CrossRef]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Sini, M.; Katsanevakis, S.; Koukourouvli, N.; Gerovasileiou, V.; Dailianis, T.; Buhl-Mortensen, L.; Damalas, D.; Dendrinos, P.; Dimas, X.; Frantzis, A.; et al. Assembling Ecological Pieces to Reconstruct the Conservation Puzzle of the Aegean Sea. Front. Mar. Sci. 2017, 4, 347. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Zenetos, A.; Corsini-Foka, M.; Tsiamis, K. Biological invasions in the Aegean sea: Temporal trends, pathways, and impacts. In The Handbook of Environmental Chemistry; Anagnostou, C.L., Kostianoy, A.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–34. [Google Scholar] [CrossRef]
- Ragkousis, M.; Sini, M.; Koukourouvli, N.; Zenetos, A.; Katsanevakis, S. Invading the Greek Seas: Spatiotemporal Patterns of Marine Impactful Alien and Cryptogenic Species. Diversity 2023, 15, 353. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Tsirintanis, K.; Sini, M.; Gerovasileiou, V.; Koukourouvli, N. Aliens in the Aegean—A sea under siege (ALAS). Res. Ideas Outcomes 2020, 6, e53057. [Google Scholar] [CrossRef]
- Pancucci-Papadopoulou, M.; Zenetos, A.; Corsini-Foka, M.; Politou, C. Update of marine alien species in Hellenic waters. Mediterr. Mar. Sci. 2005, 6, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Zenetos, A.; Corsini-Foka, M.; Crocetta, F.; Gerovasileiou, V.; Karachle, V.; Simboura, M.; Tsiamis, K.; Pancucci-Papadopoulou, M. Deep cleaning of alien and cryptogenic species records in the Greek Seas (2018 update). Manag. Biol. Invasion 2018, 9, 209–226. [Google Scholar] [CrossRef] [Green Version]
- Zenetos, A.; Arianoutsou, M.; Bazos, I.; Balopoulou, S.; Corsini-Foka, M.; Dimiza, M.; Drakopoulou, P.; Katsanevakis, S.; Kondylatos, G.; Koutsikos, N.; et al. ELNAIS: A collaborative network on aquatic alien species in Hellas (Greece). Manag. Biol. Invasion 2015, 6, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Karachle, P.; Corsini Foka, M.; Crocetta, F.; Dulcic, J.; Dzhembekova, N.; Galanidi, M.; Ivanova, P.; Shenkar, N.; Skolka, M.; Stefanova, E.; et al. Setting-up a billboard of marine invasive species in the ESENIAS area: Current situation and future expectancies. Acta Adriat. 2017, 58, 429–458. [Google Scholar] [CrossRef] [Green Version]
- Giovos, I.; Kleitou, P.; Poursanidis, D.; Batjakas, I.; Bernardi, G.; Crocetta, F.; Doumpas, N.; Kalogirou, S.; Kampouris, T.E.; Keramidas, I.; et al. Citizen-science for monitoring marine invasions and stimulating public engagement: A case project from the eastern Mediterranean. Biol. Invasions 2019, 21, 3707–3721. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Poursanidis, D.; Hoffman, R.; Rizgalla, J.; Rothman, S.B.-S.; Levitt-Barmats, Y.; Hadjioannou, L.; Trkov, D.; Garmendia, J.M.; Rizzo, M.; et al. Unpublished Mediterranean records of marine alien and cryptogenic species. BioInvasions Rec. 2020, 9, 165–182. [Google Scholar] [CrossRef]
- Ragkousis, M.; Zenetos, A.; Ben Souissi, J.; Hoffman, R.; Ghanem, R.; Taşkın, E.; Muresan, M.; Karpova, E.; Slynko, E.; Dağlı, E.; et al. Unpublished Mediterranean and Black Sea records of marine alien, cryptogenic, and neonative species. BioInvasions Rec. 2023, 12, 339–369. [Google Scholar] [CrossRef]
- Zenetos, A.; Karachle, P.K.; Corsini-Foka, M.; Gerovasileiou, V.; Simboura, N.; Xentidis, N.J.; Tsiamis, K. Is the trend in new introductions of marine non-indigenous species a reliable criterion for assessing good environmental status? Τhe case study of Greece. Mediterr. Mar. Sci. 2020, 21, 775–793. [Google Scholar] [CrossRef]
- Topouzelis, K.; Makri, D.; Stoupas, N.; Papakonstantinou, A.; Katsanevakis, S. Seagrass mapping in Greek territorial waters using Landsat-8 satellite images. Int. J. Appl. Earth Obs. Geoinf. 2018, 67, 98–113. [Google Scholar] [CrossRef]
- Ragkousis, Μ.; Papazekou, M.; Sini, M.; Zenetos, A.; Mazaris, A.D.; Katsanevakis, S. Modelling the distribution of impactful alien and cryptogenic species in the Aegean Sea. Mediterr. Mar. Sci. 2023; submitted. [Google Scholar]
- Yemshanov, D.; Koch, F.H.; Ducey, M.; Koehler, K. Mapping ecological risks with a portfolio-based technique: Incorporating uncertainty and decision-making preferences. Divers. Distrib. 2013, 19, 567–579. [Google Scholar] [CrossRef] [Green Version]
- Volery, L.; Blackburn, T.M.; Bertolino, S.; Evans, T.; Genovesi, P.; Kumschick, S.; Roy, H.E.; Smith, K.G.; Bacher, S. Improving the environmental impact classification for alien taxa (EICAT): A summary of revisions to the framework and guidelines. NeoBiota 2020, 62, 547–567. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Gueguen, M.; Engler, R.; Breiner, F.; Lafourcade, B.; Patin, R. Package ‘biomod2′ Ensemble Platform for Species Distribution Modeling. 2023. Available online: https://cran.r-project.org/web/packages/biomod2/biomod2.pdf (accessed on 28 April 2023).
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Komac, B.; Esteban, P.; Trapero, L.; Caritg, R. Modelization of the Current and Future Habitat Suitability of Rhododendron ferrugineum Using Potential Snow Accumulation. PLoS ONE 2016, 11, e0147324. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rias Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [Green Version]
- Siapatis, A.; Giannoulaki, M.; Valavanis, V.D.; Palialexis, A.; Schismenou, E.; Machias, A.; Somarakis, S. Modelling potential habitat of the invasive ctenophore Mnemiopsis leidyi in Aegean sea. In Essential Fish Habitat Mapping in the Mediterranean. Developments in Hydrobiology; Valavanis, V.D., Ed.; Springer: Dordrecht, Germany, 2008; Volume 203, pp. 281–295. [Google Scholar] [CrossRef]
- Ikhtiyar, S.; Durgham, H. The biochemical composition of two marine gelatin species Marivagia stellata and Mnemiopsis leidyi in coastal waters of Latakia city. Al-Baath Univ. J. Med. Eng. Basic Appl. Sci. 2020, 42, 1–20. [Google Scholar]
- Galil, B.S.; Kress, N.; Shiganova, T.A. First record of Mnemiopsis leidyi A. Agassiz, 1865 (Ctenophora; Lobata; Mnemiidae) off the Mediterranean coast of Israel. Aquat. Invasions 2009, 4, 357–360. [Google Scholar] [CrossRef]
- Gómez, F. Phytoplankton invasions: Comments on the validity of categorizing the nonindigenous dinoflagellates and diatoms in European Seas. Mar. Pollut. Bull. 2008, 56, 620–628. [Google Scholar] [CrossRef]
- Gómez, F. Comments on the non-indigenous microalgae in the European seas. Mar. Pollut. Bull. 2019, 148, 1–2. [Google Scholar] [CrossRef]
- Tsiamis, K.; Azzurro, E.; Bariche, M.; Çinar, M.E.; Crocetta, F.; De Clerck, O.; Galil, B.; Gomez, F.; Hoffman, R.; Jensen, K.; et al. Prioritizing marine invasive alien species in the EU through Horizon. Scanning. Aquat. Conserv. 2020, 30, 794–845. [Google Scholar] [CrossRef]
- Karachle, P.; Oikonomou, A.; Pantazi, M.; Stergiou, K.I.; Zenetos, A. Can biological traits serve as predictors for fishes’ introductions, establishment, and interactions? The Mediterranean Sea as a case study. Biology 2022, 11, 1625. [Google Scholar] [CrossRef]
- Androulidakis, Y.S.; Krestenitis, Y.N. Sea Surface Temperature Variability and Marine Heat Waves over the Aegean, Ionian, and Cretan Seas from 2008–2021. J. Mar. Sci. Eng. 2022, 10, 42. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Yadav, N.S.; Barak, S.; Lima, F.P.; Sapir, Y.; Winters, G. Responses of Invasive and Native Populations of the Seagrass Halophila stipulacea to Simulated Climate Change. Front. Mar. Sci. 2020, 6, 812. [Google Scholar] [CrossRef]
- Bernal-Ibáñez, A.; Gestoso, I.; Ramalhosa, P.; Campanati, C.; Cacabelos, E. Interaction of marine heatwaves and grazing on two canopy-forming algae. J. Exp. Mar. Biol. Ecol. 2022, 556, 151795. [Google Scholar] [CrossRef]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- Costello, K.E.; Lynch, S.A.; McAllen, R.; O’Riordan, R.M.; Culloty, S.C. Assessing the potential for invasive species introductions and secondary spread using vessel movements in maritime ports. Mar. Pollut. Bull. 2022, 177, 113496. [Google Scholar] [CrossRef]
- Tempesti, J.; Mangano, M.C.; Langeneck, J.; Lardicci, C.; Maltagliati, F.; Castelli, A. Nonindigenous species in Mediterranean ports: A knowledge baseline. Mar. Environ. Res. 2020, 161, 105056. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Morri, C.; Rosso, A.; Violanti, D.; García Raso, J.E.; Çinar, M.E.; Almogi Labin, A.; Ates, A.S.; Azzuro, E.; et al. Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Mediterr. Mar. Sci. 2012, 13, 328–352. [Google Scholar] [CrossRef] [Green Version]
- Zenetos, A.; Ovalis, P.; Giakoumi, S.; Kontadakis, C.; Lefkaditou, E.; Mpazios, G.; Simboura, N.; Tsiamis, K. Saronikos Gulf: A hotspot area for alien species in the Mediterranean Sea. BioInvasions Rec. 2020, 9, 873–889. [Google Scholar] [CrossRef]
- Zenetos, A.; Tsiamis, K.; Galanidi, M.; Carvalho, N.; Bartilotti, C.; Canning Clode, J.; Castriota, L.; Chainho, P.; Comas-González, R.; Costa, A.C.; et al. Status and Trends in the Rate of Introduction of Marine Non-Indigenous Species in European Seas. Diversity 2022, 14, 1077. [Google Scholar] [CrossRef]
- Sala, E.; Kizilkaya, Z.; Yildirim, D.; Ballesteros, E. Alien marine fishes deplete algal biomass in the eastern Mediterranean. PLoS ONE 2011, 6, e17356. [Google Scholar] [CrossRef] [Green Version]
- Giakoumi, S. Distribution patterns of the invasive herbivore Siganus luridus (Rüppell, 1829) and its relation to native benthic communities in the central Aegean Sea, Northeastern Mediterranean. Mar. Ecol. 2013, 35, 96–105. [Google Scholar] [CrossRef]
- Salomidi, M.; Giakoumi, S.; Gerakaris, V.; Issaris, Y.; Sini, M.; Tsiamis, K. Setting an ecological baseline prior to the bottom-up establishment of a marine protected area in Santorini island, Aegean Sea. Mediterr. Mar. Sci. 2016, 17, 720–737. [Google Scholar] [CrossRef] [Green Version]
- Nikolaou, A.; Tsirintanis, K.; Rilov, G.; Katsanevakis, S. Invasive Fish and Sea Urchins Drive the Status of Canopy Forming Macroalgae in the Eastern Mediterranean. Biology 2023, 12, 763. [Google Scholar] [CrossRef]
- Kytinou, E.; Issaris, Y.; Sini, M.; Salomidi, M.; Katsanevakis, S. ECOfast—An integrative ecological evaluation index for an ecosystem-based assessment of shallow rocky reefs. J. Environ. Manag. 2023, 344, 118323. [Google Scholar] [CrossRef]
- Savin, A.; Sini, M.; Xynogala, I.; Vougioukalou, V.; Stamatis, K.; Noe, S.; Ragkousis, M.; Gerovasileiou, V.; Dailianis, T.; Katsanevakis, S. Assessment of macroalgal communities on shallow rocky reefs in the Aegean Sea indicates an impoverished ecological status. Mediterr. Mar. Sci. 2023, 24, 241–258. [Google Scholar] [CrossRef]
- Lodola, A. Distribution and abundance of the tropical macroalgae Caulerpa racemosa var. cylindracea (Chlorophyta: Caulerpaceae) and Asparagopsis taxiformis (Rhodophyta: Bonnemaisoniaceae) in the upper infralittoral fringe of Linosa island (Pelagian Islands, Italy). Sci. Acta 2013, 7, 3–11. [Google Scholar]
- Cebrian, E.; Tomas, F.; López-Sendino, P.; Vilà, M.; Ballesteros, E. Biodiversity influences invasion success of a facultative epiphytic seaweed in a marine forest. Biol. Invasions 2018, 20, 2839–2848. [Google Scholar] [CrossRef]
- Rueda, J.L.; Gofas, S.; Aguilar, R.; de la Torriente, A.; García Raso, J.E.; Lo Iacono, C.; Luque, Á.A.; Marina, P.; Mateo-Ramírez, Á.; Moya-Urbano, E.; et al. Benthic fauna of littoral and deep-sea habitats of the Alboran Sea: A hotspot of biodiversity. In Alboran Sea—Ecosystems and Marine Resources; Báez, J.C., Vázquez, J.T., Eds.; Springer: Cham, Switzerland; Berlin/Heidelberg, Germany, 2021; pp. 285–358. [Google Scholar] [CrossRef]
- Bedini, R.; Bedini, M.; Bonechi, L.; Piazzi, L. Effects of non-native turf-forming Rhodophyta on mobile macro-invertebrate assemblages in the north-western Mediterranean Sea. Mar. Biol. Res. 2015, 11, 430–437. [Google Scholar] [CrossRef]
- Mancuso, F.P.; D’Agostaro, R.; Milazzo, M.; Badalamenti, F.; Musco, L.; Mikac, B.; Brutto, S.L.; Chemello, R. The invasive seaweed Asparagopsis taxiformis erodes the habitat structure and biodiversity of native algal forests in the Mediterranean Sea. Mar. Environ. Res. 2022, 173, 105515. [Google Scholar] [CrossRef]
- Pica, D.; Bloecher, N.; Dell’Anno, A.; Bellucci, A.; Pinto, T.; Pola, L.; Puce, S. Dynamics of a biofouling community in finfish aquaculture: A case study from the South Adriatic Sea. Biofouling 2019, 35, 696–709. [Google Scholar] [CrossRef]
- Argyrou, M.; Demetropoulos, A.; Hadjichristophorou, M. Expansion of the macroalga Caulerpa racemosa and changes in softbottom macrofaunal assemblages in Moni Bay, Cyprus. Oceanol. Acta 1999, 22, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, L.; Pusceddu, A.; Stabili, L.; Alifano, P.; Fraschetti, S. Potential effects of an invasive seaweed (Caulerpa cylindracea, Sonder) on sedimentary organic matter and microbial metabolic activities. Sci. Rep. 2017, 7, 12113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, L.; Pusceddu, A.; Bianchelli, S.; Fraschetti, S. Potentially combined effect of the invasive seaweed Caulerpa cylindracea (Sonder) and sediment deposition rates on organic matter and meiofaunal assemblages. Mar. Environ. Res. 2020, 159, 104966. [Google Scholar] [CrossRef] [PubMed]
- Kalogirou, S. Ecological characteristics of the invasive pufferfish Lagocephalus sceleratus (Gmelin, 1789) in the eastern Mediterranean Sea—A case study from Rhodes. Mediterr. Mar. Sci. 2013, 14, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Kalogirou, S.; Corsini, M.; Kondilatos, G.; Wennhage, H. Diet of the invasive piscivorous fish Fistularia commersonii in a recently colonized area of the eastern Mediterranean. Biol. Invasions 2007, 9, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Bariche, M.; Alwan, N.; El-Assi, H.; Zurayk, R. Diet composition of the Lessepsian bluespotted cornetfish Fistularia commersonii in the eastern Mediterranean. J. Appl. Ichthyol. 2009, 25, 460–465. [Google Scholar] [CrossRef]
- Pancucci-Papadopoulou, M.A.; Raitsos, D.E.; Corsini-Foka, M. Biological invasions and climatic warming: Implications for south-eastern Aegean ecosystem functioning. J. Mar. Biol. Assoc. UK 2012, 92, 777–789. [Google Scholar] [CrossRef]
- Saygu, İ.; Heymans, J.J.; Fox, C.J.; Özbilgin, H.; Eryaşar, A.R.; Gökçe, G. The importance of alien species to the food web and bottom trawl fisheries of the Northeastern Mediterranean, a modelling approach. J. Mar. Syst. 2020, 202, 103253. [Google Scholar] [CrossRef]
- Caruso, A.; Cosentino, C. The first colonization of the Genus Amphistegina and other exotic benthic foraminifera of the Pelagian Islands and south-eastern Sicily (central Mediterranean Sea). Mar. Micropaleontol. 2014, 111, 38–52. [Google Scholar] [CrossRef]
- Mouanga, G.H. Impact and Range Extension of Invasive Foraminifera in the NW Mediterranean Sea: Implications for Diversity and Ecosystem Functioning. Ph.D. Thesis, University of Bonn, Bonn, Germany, 2018. [Google Scholar]
- Streftaris, N.; Zenetos, A. Alien Marine Species in the Mediterranean—The 100 ‘Worst Invasives’ and their Impact. Mediterr. Mar. Sci. 2006, 7, 87–118. [Google Scholar] [CrossRef] [Green Version]
- Piazzi, L.; Cinelli, F. Distribution and dominance of two introduced turf-forming macroalgae on the coast of Tuscany, Italy, Northwestern Mediterranean sea in relation to different habitats and sedimentation. Bot. Mar. 2001, 44, 509–520. [Google Scholar] [CrossRef]
- Ballesteros, E.; Cebrian, E.; Alcoverro, T. Mortality of shoots of Posidonia oceanica following meadow invasion by the red alga Lophocladia lallemandii. Bot. Mar. 2007, 50, 8–13. [Google Scholar] [CrossRef]
- Sureda, A.; Box, A.; Terrados, J.; Deudero, S.; Pons, A. Antioxidant response of the seagrass Posidonia oceanica when epiphytized by the invasive macroalgae Lophocladia lallemandii. Mar. Environ. Res. 2008, 66, 359–363. [Google Scholar] [CrossRef]
- Piazzi, L.; Cinelli, F. Effets de l’expansion des Rhodophyceae introduites Acrothamnion preissii et Womersleyella setacea sur les communautés algales des rhizomes de Posidonia oceanica de Méditerranée occidentale. Cryptogam. Algol. 2000, 21, 291–300. [Google Scholar] [CrossRef]
- Deudero, S.; Blanco, A.; Box, A.; Mateu-Vicens, G.; Cabanellas-Reboredo, M.; Sureda, A. Interaction between the invasive macroalga Lophocladia lallemandii and the bryozoan Reteporella grimaldii at seagrass meadows: Density and physiological responses. Biol. Invasions 2010, 12, 41–52. [Google Scholar] [CrossRef]
- Marchessaux, G.; Faure, V.; Chevalier, C.; Thibault, D. Refugia area for the ctenophore Mnemiopsis leidyi A. Agassiz 1865 in the Berre Lagoon (southeast France): The key to its persistence. Reg. Stud. Mar. Sci. 2020, 39, 101409. [Google Scholar] [CrossRef]
- Marchessaux, G.; Belloni, B.; Gadreaud, J.; Thibault, D. Predation assessment of the invasive ctenophore Mnemiopsis leidyi in a French Mediterranean lagoon. J. Plankton Res. 2021, 43, 161–179. [Google Scholar] [CrossRef]
- Fiori, E.; Benzi, M.; Ferrari, C.R.; Mazziotti, C. Zooplankton community structure before and after Mnemiopsis leidyi arrival. J. Plankton Res. 2019, 41, 803–820. [Google Scholar] [CrossRef]
- Korpinen, S.; Klančnik, K.; Peterlin, M.; Nurmi, M.; Laamanen, L.; Zupančič, G.; Popit, A.; Murray, C.; Harvey, T.; Andersen, J.H.; et al. Multiple Pressures and Their Combined Effects in Europe’s Seas; ETC/ICM Technical Report 4/2019; European Topic Centre on Inland, Coastal and Marine Waters: Magdeburg, Germany, 2019; p. 164. [Google Scholar]
- Magliozzi, C.; Tsiamis, K.; Vigiak, O.; Deriu, I.; Gervasini, E.; Cardoso, A.C. Assessing invasive alien species in European catchments: Distribution and impacts. Sci. Total Environ. 2020, 732, 138677. [Google Scholar] [CrossRef]
- Arianoutsou, M.; Adamopoulou, C.; Andriopoulos, P.; Bazos, I.; Christopoulou, A.; Galanidis, A.; Kalogianni, E.; Karachle, P.K.; Kokkoris, Y.; Martinou, A.F.; et al. HELLAS-ALIENS. The Invasive Alien Species of Greece: Time trends, origin and pathways. NeoBiota, 2023; accepted. [Google Scholar]
- Airoldi, L. Effects of disturbance, life histories, and overgrowth on coexistence of algal crusts and turfs. Ecology 2000, 81, 798–814. [Google Scholar] [CrossRef]
- Arndt, E.; Givan, O.; Edelist, D.; Sonin, O.; Belmaker, J. Shifts in Eastern Mediterranean Fish Communities: Abundance Changes, Trait Overlap, and Possible Competition between Native and Non-Native Species. Fishes 2018, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Baldacconi, R.; Corriero, G. Effects of the spread of the alga Caulerpa racemosa var. cylindracea on the sponge assemblage from coralligenous concretions of the Apulian coast (Ionian Sea, Italy). Mar. Ecol. 2009, 30, 337–345. [Google Scholar] [CrossRef]
- Balestri, E.; Vallerini, F.; Menicagli, V.; Barnaba, S.; Lardicci, C. Biotic resistance and vegetativepropagule pressure co-regulate the invasion success of a marine clonal macrophyte. Sci. Rep. 2018, 8, 16621. [Google Scholar] [CrossRef] [Green Version]
- Bedini, R.; Bonechi, L.; Piazzi, L. Spread of the introduced red alga Lophocladia lallemandii in the Tuscan Archipelago (NW Mediterranean Sea). Cryptogamie Algol. 2011, 32, 383–391. [Google Scholar] [CrossRef]
- Bulleri, F.; Benedetti-Cecchi, L.; Ceccherelli, G.; Tamburello, L. A few is enough: A low cover of a non-native seaweed reduces the resilience of Mediterranean macroalgal stands to disturbances of varying extent. Biol. Invasions 2017, 19, 2291–2305. [Google Scholar] [CrossRef]
- Cebrian, E.; Linares, C.; Marschal, C.; Garrabou, J. Exploring the effects of invasive algae on the persistence of gorgonian populations. Biol. Invasions 2012, 14, 2647–2656. [Google Scholar] [CrossRef]
- Cebrian, E.; Rodríguez-Prieto, C. Marine Invasion in the Mediterranean Sea: The Role of Abiotic Factors When There Is No Biological Resistance. PLoS ONE 2012, 7, e31135. [Google Scholar] [CrossRef] [Green Version]
- Çinar, M.E.; Katağan, T.; Koçak, F.; Öztürk, B.; Ergen, Z.; Kocatas, A.; Önen, M.; Kirkim, F.; Bakir, K.; Kurt, G.; et al. Faunal assemblages of the mussel Mytilus galloprovincialis in and around Alsancak Harbour (Izmir Bay, eastern Mediterranean) with special emphasis on alien species. J. Marine Syst. 2008, 71, 1–17. [Google Scholar] [CrossRef]
- Coma, R.; Serrano, E.; Linares, C.; Ribes, M.; Díaz, D.; Ballesteros, E. Sea Urchins Predation Facilitates Coral Invasion in a Marine Reserve. PLoS ONE 2011, 6, e22017. [Google Scholar] [CrossRef] [Green Version]
- Crocetta, F.; Gofas, S.; Salas, C.; Tringali, L.; Zenetos, A. Local ecological knowledge versus published literature: A review of non-indigenous Mollusca in Greek marine waters. Aquat. Invasions 2017, 12, 415–434. [Google Scholar] [CrossRef]
- de Caralt, S.; Cebrian, E. Impact of an invasive alga (Womersleyella setacea) on sponge assemblages: Compromising the viability of future populations. Biol. Invasions 2013, 15, 1591–1600. [Google Scholar] [CrossRef]
- Deudero, S.; Box, A.; Vázquez-Luis, M.; Arroyo, N.L. Benthic community responses to macroalgae invasions in seagrass beds: Diversity, isotopic niche and food web structure at community level. Estuar. Coast. Shelf Sci. 2014, 142, 12–22. [Google Scholar] [CrossRef]
- Dumay, O.; Fernandez, C.; Pergent, G. Primary production and vegetative cycle in Posidonia oceanica when in competition with the green algae Caulerpa taxifolia and Caulerpa racemosa. J. Mar. Biol. Assoc. U.K. 2002, 82, 379–387. [Google Scholar] [CrossRef]
- Evagelopoulos, A.; Nikolaou, A.; Michailidis, N.; Kampouris, T.E.; Batjakas, I.E. Progress of the dispersal of the alien goatfish Parupeneus forsskali (Fourmanoir & Guézé, 1976) in the Mediterranean, with preliminary information on its diet composition in Cyprus. BioInvasions Rec. 2020, 9, 209–222. [Google Scholar] [CrossRef]
- Fortič, A.; Mavrič, B. First record of the bryozoan Tricellaria inopinata (d’Hondt & Occhipinti-Ambrogi, 1985) from the Slovenian sea. Ann. Ser. Hist. Nat. 2018, 28, 155–160. [Google Scholar] [CrossRef]
- Galanidi, M.; Zenetos, A.; Bacher, S. Assessing the socio-economic impacts of priority marine invasive fishes in the Mediterranean with the newly proposed SEICAT methodology. Mediterr. Mar. Sci. 2018, 19, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Gerovasileiou, V.; Voultsiadou, E.; Issaris, Y.; Zenetos, A. Alien biodiversity in Mediterranean marine caves. Mar. Ecol. 2016, 37, 239–256. [Google Scholar] [CrossRef]
- Green, S.J.; Akins, J.L.; Maljkovic, A.; Cote, I.M. Invasive Lionfish Drive Atlantic Coral Reef Fish Declines. PLoS ONE 2012, 7, e32596. [Google Scholar] [CrossRef]
- Hixon, M.A.; Green, S.J.; Albins, M.A.; Akins, J.L.; Morris, J.A. Lionfish: A major marine invasion. Mar. Ecol. Prog. Ser. 2016, 558, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Kampouris, T.E.; Porter, J.S.; Sanderson, W.G. Callinectes sapidus Rathbun, 1896 (Brachyura: Portunidae): An assessment on its diet and foraging behaviour, Thermaikos Gulf, NW Aegean Sea, Greece: Evidence for ecological and economic impacts. Crustacean Res. 2019, 48, 23–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kersting, D.K.; Cebrian, E.; Casado, C.; Teixidó, N.; Garrabou, J.; Linares, C. Experimental evidence of the synergistic effects of warming and invasive algae on a temperate reef-builder coral. Sci. Rep. 2015, 5, 18635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, J.; Verlaque, M. The Caulerpa racemosa invasion: A critical review. Mar. Pollut. Bull. 2008, 56, 205–225. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.C.; Verlaque, M. Experimental removal of the invasive Caulerpa racemosa triggers partial assemblage recovery. J. Mar. Biol. Assoc. UK 2011, 91, 117–125. [Google Scholar] [CrossRef]
- Kružić, P.; Žuljević, A.; Nikolić, V. The highly invasive alga Caulerpa racemosa var. cylindracea poses a new threat to the banks of the coral Cladocora caespitosa in the Adriatic Sea. Coral Reefs 2008, 27, 441. [Google Scholar] [CrossRef]
- Marchini, A.; Ferrario, J.; Minchin, D. Marinas may act as hubs for the spread of the pseudoindigenous bryozoan Amathia verticillata (Delle Chiaje, 1822) and its associates. Sci. Mar. 2015, 79, 355–365. [Google Scholar] [CrossRef] [Green Version]
- McCann, L.; Keith, I.; Carlton, J.; Ruiz, G.; Dawson, T.; Collins, K. First record of the non-native bryozoan Amathia (=Zoobotryon) verticillata (delle Chiaje, 1822) (Ctenostomata) in the Galápagos Islands. BioInvasions Rec. 2015, 4, 255–260. [Google Scholar] [CrossRef]
- Mouanga, G.H.; Langer, M.R. At the front of expanding ranges: Shifting community structures at amphisteginid species range margins in the Mediterranean Sea. Neues Jahrb. Geol. Paläontologie Abh. 2014, 271, 141–150. [Google Scholar] [CrossRef]
- Mutlu, E.; Ergev, B. Temporal variability of density and diverse shell occupancy of Diogenes pugilator on a sandy bottom of the Levantine Sea and their biometrical relationship. Cah. Biol. Mar. 2010, 51, 55–67. [Google Scholar] [CrossRef]
- Navarro-Barranco, C.; Florido, M.; Ros, M.; González-Romero, P.; Guerra-García, J.M. Impoverished mobile epifaunal assemblages associated with the invasive macroalga Asparagopsis taxiformis in the Mediterranean Sea. Mar. Environ. Res. 2018, 141, 44–52. [Google Scholar] [CrossRef]
- Otero, M.; Cebrian, E.; Francour, P.; Galil, B.; Savini, D. Monitoring Marine Invasive Species in Mediterranean Marine Protected Areas (MPAs): A Strategy and Practical Guide for Managers; Medpan North project; IUCN: Malaga, Spain, 2013; p. 136. [Google Scholar]
- Ounifi-Ben Amor, K.; Rifi, Μ.; Ghanem, R.; Draeif, I.; Zaouali, J.; Ben Souissi, J. Update of alien fauna and new records from Tunisian marine waters. Mediterr. Mar. Sci. 2016, 17, 124–143. [Google Scholar] [CrossRef]
- Piazzi, L.; Cinelli, F. Evaluation of benthic macroalgal invasion in a harbour area of the western Mediterranean Sea. Eur. J. Phycol. 2003, 38, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Piazzi, L.; Balata, D. Invasion of alien macroalgae in different Mediterranean habitats. Biol. Invasions 2009, 11, 193–204. [Google Scholar] [CrossRef]
- Piazzi, L.; Ceccherelli, G. Alpha and beta diversity in Mediterranean macroalgal assemblages: Relevancy and type of effect of anthropogenic stressors vs natural variability. Mar. Biol. 2020, 167, 32. [Google Scholar] [CrossRef]
- Piazzi, L.; Balata, D.; Ceccherelli, G.; Cinelli, F. Interactive effect of sedimentation and Caulerpa racemosa var. cylindracea invasion on macroalgal assemblages in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2005, 64, 467–474. [Google Scholar] [CrossRef]
- Piazzi, L.; Ceccherelli, G. Persistence of biological invasion effects: Recovery of macroalgal assemblages after removal of Caulerpa racemosa var. cylindracea. Estuar. Coast. Shelf Sci. 2006, 68, 455–461. [Google Scholar] [CrossRef]
- Piazzi, L.; Ceccherelli, G.; Cinelli, F. Threat to macroalgal diversity: Effects of the introduced green alga Caulerpa racemosa in the Mediterranean. Mar. Ecol. Prog. Ser. 2001, 210, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Piazzi, L.; Balata, D.; Foresi, L.; Cristaudo, C.; Cinelli, F. Sediment as a constituent of Mediterranean benthic communities dominated by Caulerpa racemosa var. cylindracea. Sci. Mar. 2007, 71, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Pinnegar, J.K.; Tomczak, M.T.; Link, J.S. How to determine the likely indirect food-web consequences of a newly introduced non-native species: A worked example. Ecol. Modell. 2014, 272, 379–387. [Google Scholar] [CrossRef]
- Pusceddu, A.; Fraschetti, S.; Scopa, M.; Rizzo, L.; Danovaro, R. Meiofauna communities, nematode diversity and C degradation rates in seagrass (Posidonia oceanica L.) and unvegetated sediments invaded by the algae Caulerpa cylindracea (Sonder). Mar. Environ. Res. 2016, 119, 88–99. [Google Scholar] [CrossRef]
- Safriel, U.N.; Sasson-Frostig, Z. Can colonizing mussel outcompete indigenous mussel? J. Exp. Mar. Biol. Ecol. 1988, 117, 211–226. [Google Scholar] [CrossRef]
- Savva, I.; Chartosia, N.; Antoniou, C.; Kleitou, P.; Georgiou, A.; Stern, N.; Hadjioannou, L.; Jimenez, C.; Andreou, V.; Hall-Spencer, J.M.; et al. They are here to stay: The biology and ecology of lionfish (Pterois miles) in the Mediterranean Sea. J. Fish Biol. 2020, 97, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Serrano, E.; Coma, R.; Ribes, M. A phase shift from macroalgal to coral dominance in the Mediterranean. Coral Reefs 2012, 31, 1199. [Google Scholar] [CrossRef] [Green Version]
- Sghaier, Y.R.; Zakhama-Sraieb, R.; Charfi-Cheikhrouha, F. Effects of the invasive seagrass Halophila stipulacea on the native seagrass Cymodocea nodosa. In Proceedings of the 5th Mediterranean Symposium on Marine Vegetation 2014, Portorož, Slovenia, 27–28 October; pp. 167–171.
- Schaffelke, B.; Hewitt, C.L. Impacts of introduced seaweeds. Bot. Mar. 2007, 50, 397–417. [Google Scholar] [CrossRef]
- Tsiamis, K. Alien Macroalgae in the Sublittoral Zone of the Greek Coasts. PhD Thesis, National and Kapodistrian University of Athens, Athens, Greece, 2012; p. 335. [Google Scholar]
- Zanolla, M.; Carmona, R.; De La Rosa, J.; Altamirano, M. Structure and temporal dynamics of a seaweed assemblage dominated by the invasive lineage 2 of Asparagopsis taxiformis (Bonnemaisoniaceae, Rhodophyta) in the Alboran Sea. Mediterr. Mar. Sci. 2018, 19, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Žuljević, A.; Thibaut, T.; Despalatović, M.; Cottalorda, J.M.; Nikolić, V.; Cvitković, I.; Antolić, B. Invasive alga Caulerpa racemosa var. cylindracea makes a strong impact on the Mediterranean sponge Sarcotragus spinosulus. Biol. Invasions 2011, 13, 2303. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
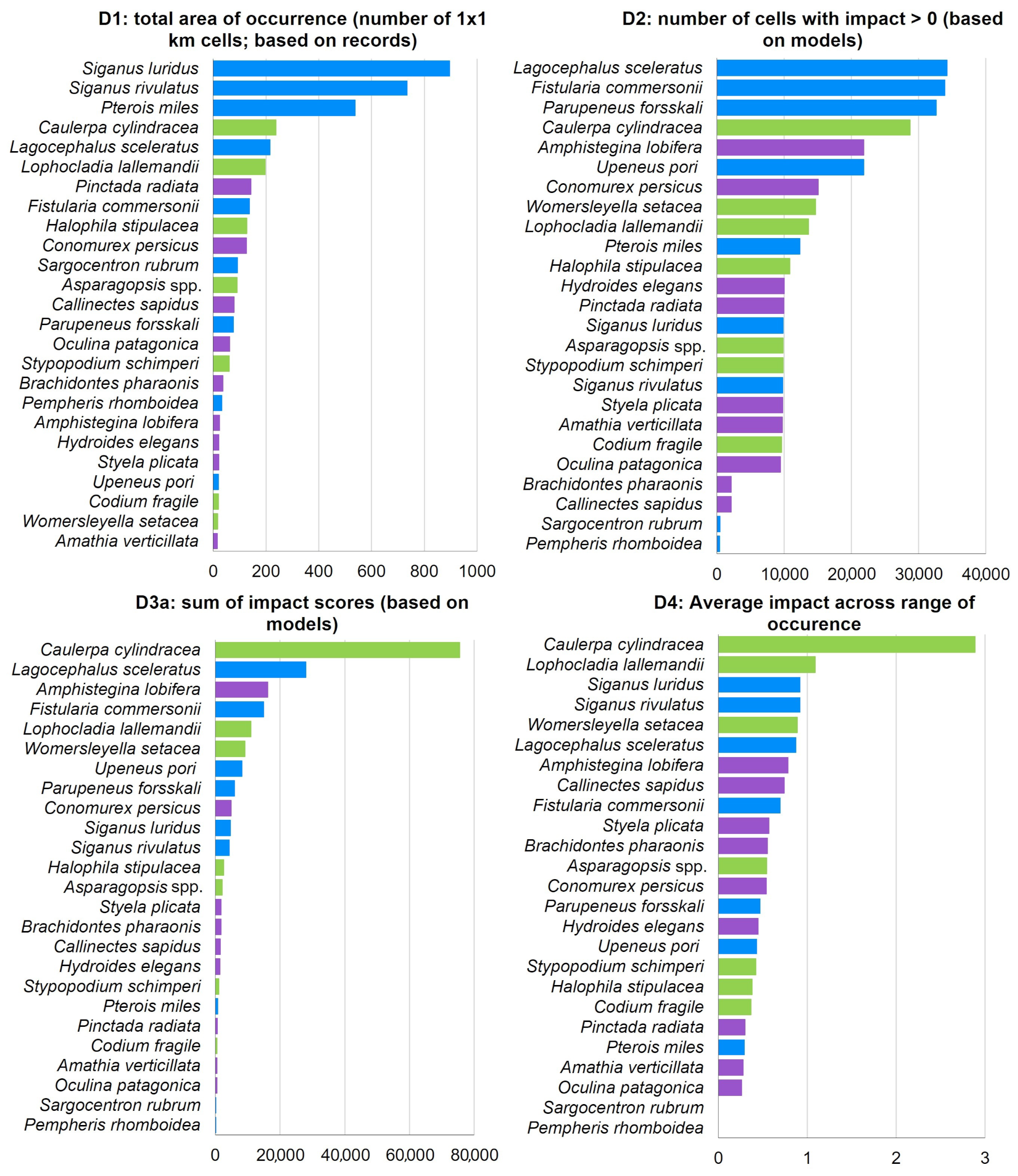
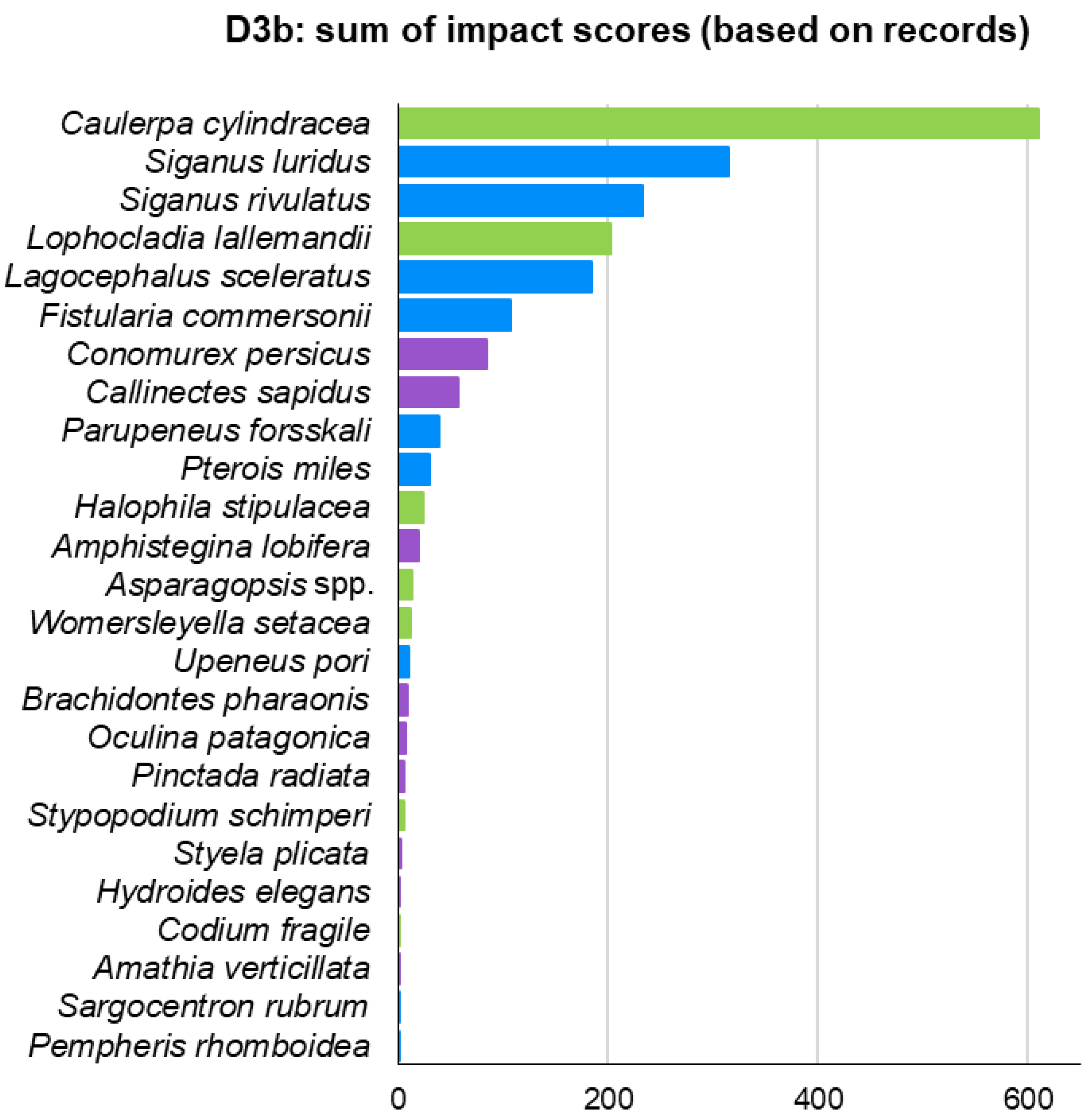
| Biotic Group | Phylum: Class | Species | Status | Number of Records |
|---|---|---|---|---|
| Macrophytes | Ochrophyta: Phaeophyceae | Stypopodium schimperi | Alien | 67 |
| Macrophytes | Chlorophyta: Ulvophyceae | Caulerpa cylindracea | Alien | 283 |
| Macrophytes | Chlorophyta: Ulvophyceae | Codium fragile | Alien | 22 |
| Macrophytes | Rhodophyta: Florideophyceae | Asparagopsis spp. | Alien | 117 |
| Macrophytes | Rhodophyta: Florideophyceae | Lophocladia lallemandii | Alien | 224 |
| Macrophytes | Rhodophyta: Florideophyceae | Womersleyella setacea | Alien | 24 |
| Macrophytes | Tracheophyta: Magnoliopsida | Halophila stipulacea | Alien | 132 |
| Invertebrates | Foraminifera: Globothalamea | Amphistegina lobifera | Alien | 26 |
| Invertebrates | Ctenophora: Tentaculata | Mnemiopsis leidyi | Alien | 77 |
| Invertebrates | Cnidaria: Anthozoa | Oculina patagonica | Cryptogenic | 51 |
| Invertebrates | Bryozoa: Gymnolaemata | Amathia verticillata | Cryptogenic | 19 |
| Invertebrates | Mollusca: Bivalvia | Brachidontes pharaonis | Alien | 35 |
| Invertebrates | Mollusca: Bivalvia | Pinctada radiata | Alien | 159 |
| Invertebrates | Mollusca: Gastropoda | Conomurex persicus | Alien | 129 |
| Invertebrates | Annelida: Polychaeta | Hydroides elegans | Alien | 24 |
| Invertebrates | Arthropoda: Malacostraca | Callinectes sapidus | Alien | 115 |
| Invertebrates | Chordata: Ascidiacea | Styela plicata | Alien | 21 |
| Fish | Chordata: Teleostei | Fistularia commersonii | Alien | 171 |
| Fish | Chordata: Teleostei | Lagocephalus sceleratus | Alien | 254 |
| Fish | Chordata: Teleostei | Parupeneus forsskali | Alien | 70 |
| Fish | Chordata: Teleostei | Pempheris rhomboidea | Alien | 31 |
| Fish | Chordata: Teleostei | Pterois miles | Alien | 574 |
| Fish | Chordata: Teleostei | Sargocentron rubrum | Alien | 88 |
| Fish | Chordata: Teleostei | Siganus luridus | Alien | 919 |
| Fish | Chordata: Teleostei | Siganus rivulatus | Alien | 748 |
| Fish | Chordata: Teleostei | Upeneus pori | Alien | 23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsirintanis, K.; Sini, M.; Ragkousis, M.; Zenetos, A.; Katsanevakis, S. Cumulative Negative Impacts of Invasive Alien Species on Marine Ecosystems of the Aegean Sea. Biology 2023, 12, 933. https://doi.org/10.3390/biology12070933
Tsirintanis K, Sini M, Ragkousis M, Zenetos A, Katsanevakis S. Cumulative Negative Impacts of Invasive Alien Species on Marine Ecosystems of the Aegean Sea. Biology. 2023; 12(7):933. https://doi.org/10.3390/biology12070933
Chicago/Turabian StyleTsirintanis, Konstantinos, Maria Sini, Michail Ragkousis, Argyro Zenetos, and Stelios Katsanevakis. 2023. "Cumulative Negative Impacts of Invasive Alien Species on Marine Ecosystems of the Aegean Sea" Biology 12, no. 7: 933. https://doi.org/10.3390/biology12070933
APA StyleTsirintanis, K., Sini, M., Ragkousis, M., Zenetos, A., & Katsanevakis, S. (2023). Cumulative Negative Impacts of Invasive Alien Species on Marine Ecosystems of the Aegean Sea. Biology, 12(7), 933. https://doi.org/10.3390/biology12070933