
Rhizobacteria Isolated from Amazonian Soils Reduce the Effects of Water Stress on the Growth of Açaí (Euterpe oleracea Mart.) Palm Seedlings

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Isolation of Rhizobacteria
2.3. Pre-Inoculum and Selection of PGPRs
2.3.1. Test for Inorganic Phosphate Solubilization and Organic Phosphate Mineralization
2.3.2. Test for Siderophore Production
2.3.3. Indole-3-Acetic Acid (IAA) Production Test
2.3.4. Biofilm Production
2.3.5. Cellulolytic Activity
2.3.6. ACC Deaminase Enzyme Synthesis
2.3.7. Test for Production of Antimicrobial Substances (AMS)
2.4. Statistical Analyses
2.5. Selection of Drought-Tolerant Strains
2.6. DNA Extraction and Molecular Identification
2.7. Effect of PGPR Inoculation on Seed Germination and Initial Seedling Development of Açaí under Water Deficiency
3. Results
3.1. Isolation, Selection of PGPRs, and Evaluation of Drought Tolerance
3.2. Molecular Identification and Phylogenetic Analysis
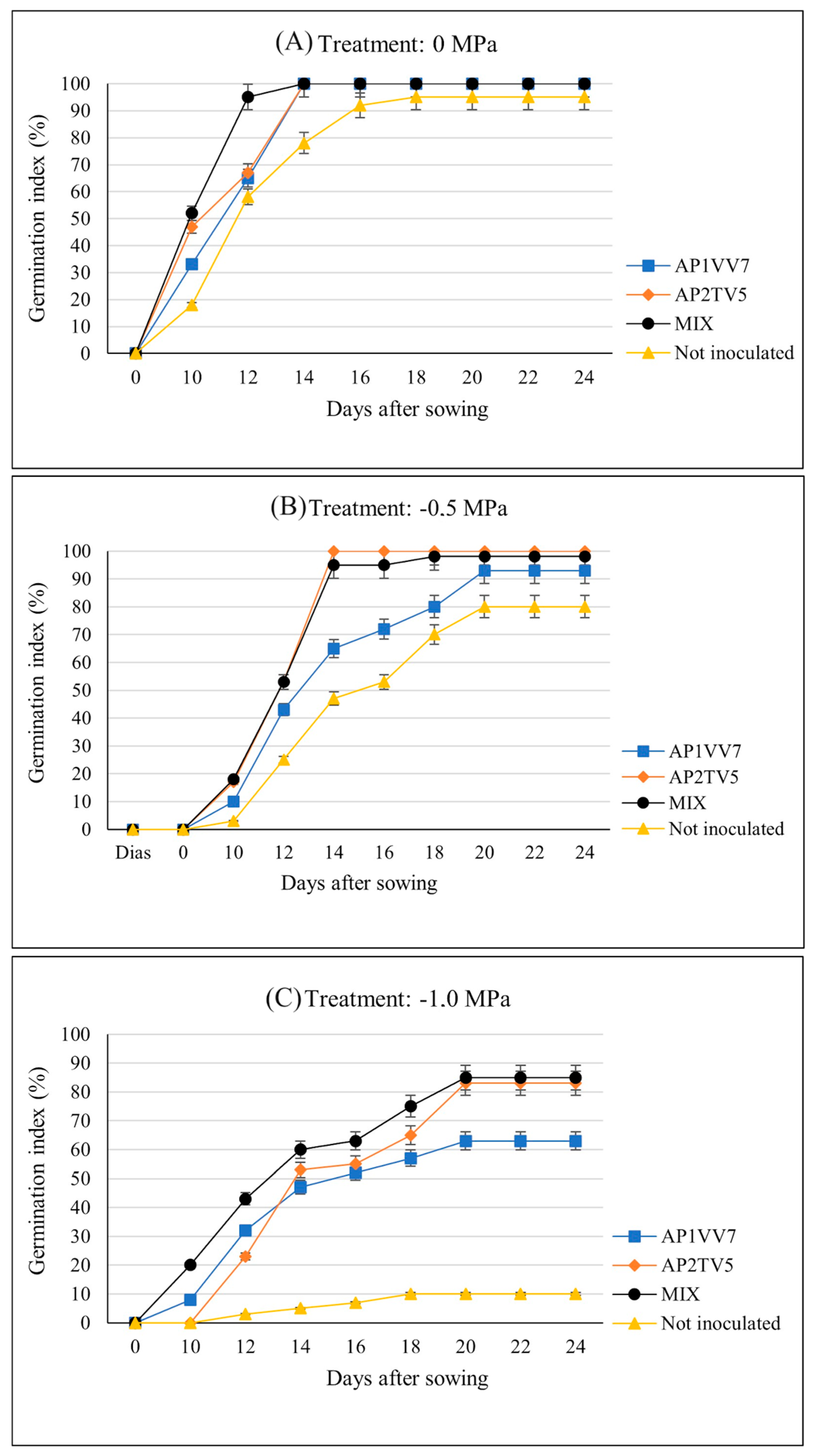
3.3. Assessment of Germination Percentage and Speed Index
3.4. Initial Growth of Açaí Seedlings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamaguchi, K.K.L.; Pereira, L.F.R.; Lamarão, C.V.; Lima, E.S.; Veiga-Junior, V.F. Amazon acai: Chemistry and biological activities: A review. Food Chem. 2015, 179, 137–151. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, L.A.S.; Jardim, M.A.G. Sobrevivência e Mortalidade de Plantas de Açaizeiro (Euterpe oleracea Mart.) Cultivadas em Capoeira no Nordeste Paraense. Rev. Bras. Biociências 2007, 5, 255–257. [Google Scholar]
- Instituto Brasileiro de Geografia e Estatística (IBGE). Açaí—Cultivo. Available online: https://www.ibge.gov.br/explica/producao-agropecuaria/acai-cultivo/pa (accessed on 21 April 2024).
- Santos, L.R.; de Campos Almeida, M.; Wittmann, F. Biometria e germinação de sementes de Macrolobium acaciifolium (Benth.) Benth. de várzea e igapó da Amazônia Central. Iheringia Série Botânica 2020, 75, e2020004. [Google Scholar] [CrossRef]
- Empresa Brasileira de Pesquisa Agropecuária—EMBRAPA. Açaí de Terra Firme. Available online: https://www.embrapa.br/amazonia-oriental/portal-do-acai/acai-de-terra-firme (accessed on 14 October 2020).
- Supremo Tribunal Federal. Agenda 2030. Available online: https://portal.stf.jus.br/hotsites/agenda-2030/ (accessed on 21 April 2024).
- Singh, R.P.; Jha, P.N. The PGPR Stenotrophomonas maltophilia SBP-9 augments resistance against biotic and abiotic stress in wheat plants. Front. Microbiol. 2017, 8, 275381. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Hameed, S.; Shahid, M.; Iqbal, M.; Lazarovits, G.; Imran, A. Functional characterization of potential PGPR exhibiting broad-spectrum antifungal activity. Microbiol. Res. 2020, 232, 126389. [Google Scholar] [CrossRef]
- Panchami, P.S.; Geetha Thanuja, K.; Karthikeyan, S. Isolation and Characterization of Indigenous Plant Growth-Promoting Rhizobacteria (PGPR) from Cardamom Rhizosphere. Curr. Microbiol. 2020, 77, 2963–2981. [Google Scholar] [CrossRef] [PubMed]
- de Castro, G.L.S.; da Silva Júnior, D.D.; Viana, R.G.; Rêgo, M.C.F.; da Silva, G.B. Photosynthetic apparatus protection and drought effect mitigation in açaí palm seedlings by rhizobacteria. Acta Physiol. Planta. 2019, 41, 163. [Google Scholar] [CrossRef]
- Castro, G.L.; Júnior, D.D.S.; Bueno, A.C.S.; Silva, G.B. Anthracnose in açaí palm leaves reduces leaf gas exchange and chlorophyll a fluorescence. Trop. Plant Pathol. 2017, 42, 13–20. [Google Scholar] [CrossRef]
- Empresa Brasileira de Pesquisa Agropecuária—EMBRAPA. Manual de Métodos de Análise de Solo, 3rd ed.; Empresa Brasileira de Pesquisa Agropecuária—EMBRAPA: Brasília, Brazil, 2017; 573p, ISBN 978-85-7035-771-7. [Google Scholar]
- de Sousa, R.S.R.; Lima, G.V.S.; Garcias, J.T.; Gomes, G.O.; Mateus, J.R.; Madeira, L.D.P.d.S.; Seldin, L.; Rogez, H.L.G.; Marques, J.M. The Microbial Community Structure in the Rhizosphere of Theobroma cacao L. and Euterpe oleracea Mart. Is Influenced by Agriculture System in the Brazilian Amazon. Microorganisms 2024, 12, 398. [Google Scholar] [CrossRef]
- Paul, D.; Sinha, S.N. Phosphate solubilization potential and phosphatase activity of some bacterial strains isolated from thermal power plant effluent exposed water of river Ganga. CIBTech J. Microbiol. 2013, 2, 1–7. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Gusmiaty; Restu, M.; Bachtiar, B.; Larekeng, S.H. Gibberellin and IAA Production by Rhizobacteria from Various Private Forest. IOP Conf. Ser. Earth Environ. Sci. 2019, 270, 012018. [Google Scholar] [CrossRef]
- Mendonça, S.M. Rizobactérias Multifuncionais na Mitigação do Déficit Hídrico e da Brusone Foliar em Arroz de Terras Altas. 2021. Available online: https://bdtd.ibict.br/vufind/Record/UFG-2_f151909de7ced3bbee29f60aae628601 (accessed on 21 April 2024).
- Aguiar, K.P. Prospecção de Bactérias Promotoras do Crescimento Vegetal Associadas a Vermicompostos. Master’s Thesis, Plant Production—Campos dos Goytacazes: Universidade Estadual do Norte Fluminense Darcy Ribeiro, Rio de Janeiro, Brazil, 2012. [Google Scholar]
- Ali, S.Z.; Sandhya, V.; Venkateswar Rao, L. Isolation and characterization of drought-tolerant ACC deaminase and exopolysaccharide-producing fluorescent Pseudomonas sp. Ann. Microbiol. 2014, 64, 493–502. [Google Scholar] [CrossRef]
- Goswami, M.; Deka, S. Isolation of a novel rhizobacteria having multiple plant growth promoting traits and antifungal activity against certain phytopathogens. Microbiol Res. 2020, 240, 126516. [Google Scholar] [CrossRef]
- Ashry, N.M.; Alaidaroos, B.A.; Mohamed, S.A.; Badr, O.A.; El-Saadony, M.T.; Esmael, A. Utilization of drought-tolerant bacterial strains isolated from harsh soils as a plant growth-promoting rhizobacteria (PGPR). Saudi J. Biol. Sci. 2022, 29, 1760–1769. [Google Scholar] [CrossRef]
- Seldin, L.; Dubnau, D. Deoxyribonucleic acid homology among Bacillus polymyxa, Bacillus macerans, Bacillus azotofixans, and other nitrogen-fixing Bacillus strains. Int. J. Syst. Evol. Microbiol. 1985, 35, 151–154. [Google Scholar] [CrossRef]
- Massol-Deya, A.A.; Odelson, D.A.; Hickey, R.F.; Tiedje, J.M. Bacterial community fingerprinting of amplified 16S and 16–23S ribosomal DNA gene sequences and restriction endonuclease analysis (ARDRA). In Molecular Microbial Ecology Manual; Springer: Dordrecht, The Netherlands, 1995; pp. 289–296. [Google Scholar] [CrossRef]
- Hall, T.A. BIOEDIT: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/ NT. Phytopathol Mediterr. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Zia, R.; Nawaz, M.S.; Yousaf, S.; Amin, I.; Hakim, S.; Mirza, M.S.; Imran, A. Seed inoculation of desert-plant growth-promoting rhizobacteria induce biochemical alterations and develop resistance against water stress in wheat. Physiol. Plant. 2021, 172, 990–1006. [Google Scholar] [CrossRef] [PubMed]
- Chukwuneme, C.F.; Babalola, O.O.; Kutu, F.R.; Ojuederie, O.B. Characterization of actinomycetes isolates for plant growth promoting traits and their effects on drought tolerance in maize. J. Plant Interact. 2020, 15, 93–105. [Google Scholar] [CrossRef]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ.-Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef]
- Dashti, N.; Al-Sarraf, N.Y.A.; Cherian, V.M.; Montasser, M.S. Isolation and characterization of novel plant growth-promoting rhizobacteria (PGPR) isolates from tomato (Solanum lycopersicum L.) rhizospherical soil: A novel IAA producing bacteria. Kuwait J. Sci. 2021, 48, 2. [Google Scholar] [CrossRef]
- Prasannakumar, S.P.; Gowtham, H.G.; Hariprasad, P.; Shivaprasad, K.; Niranjana, S.R. Delftia tsuruhatensis WGR–UOM–BT1, a novel rhizobacterium with PGPR properties from Rauwolfia serpentina (L.) Benth. ex Kurz also suppresses fungal phytopathogens by producing a new antibiotic—AMTM. Lett. Appl. Microbiol. 2015, 61, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Anderson, A.J.; Kim, Y.C. Root-Associated Bacteria Are Biocontrol Agents for Multiple Plant Pests. Microorganisms 2022, 10, 1053. [Google Scholar] [CrossRef] [PubMed]
- Sena, I.S.; Ferreira, A.M.; Marinho, V.H.; Holanda, F.H.; Borges, S.F.; de Souza, A.A.; de Carvalho, R.; Koga, R.; Lima, A.L.; Florentino, A.C.; et al. Euterpe oleracea Mart (Açaizeiro) from the Brazilian Amazon: A Novel Font of Fungi for Lipase Production. Microorganisms 2022, 10, 2394. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Rai, S.; Bano, A.; Sharma, S.; Kumar, M.; Binsuwaidan, R.; Suhail Khan, M.; Upadhyay, T.K.; Alshammari, N.; Saeed, M.; et al. ACC Deaminase Produced by PGPR Mitigates the Adverse Effect of Osmotic and Salinity Stresses in Pisum sativum through Modulating the Antioxidants Activities. Plants 2022, 11, 3419. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Saraf, M. ACC deaminase producing PGPR modulates nutrients uptake, soil properties and growth of cluster bean (Cyamopsis tetragonoloba L.) under deficit irrigation. Biologia 2023, 78, 2303–2316. [Google Scholar] [CrossRef]
- Xue, P.P.; Carrillo, Y.; Pino, V.; Minasny, B.; McBratney, A.B. Soil properties drive microbial community structure in a large scale transect in South Eastern Australia. Sci. Rep. 2018, 8, 11725. [Google Scholar] [CrossRef] [PubMed]
- Lori, M.; Armengot, L.; Schneider, M.; Schneidewind, U.; Bodenhausen, N.; Mäder, P.; Krause, H.M. Organic management enhances soil quality and drives microbial community diversity in cocoa production systems. Sci. Total Environ. 2022, 834, 155223. [Google Scholar] [CrossRef]
- Meza, C.; Valenzuela, F.; Echeverría-Vega, A.; Gomez, A.; Sarkar, S.; Cabeza, R.A.; Arencibia, A.D.; Quiroz, K.; Carrasco, B.; Banerjee, A. Plant-growth-promoting bacteria from rhizosphere of Chilean common bean ecotype (Phaseolus vulgaris L.) supporting seed germination and growth against salinity stress. Front. Plant Sci. 2022, 13, 1052263. [Google Scholar] [CrossRef]
- Nie, M.; Wu, C.; Tang, Y.; Shi, G.; Wang, X.; Hu, C.; Cao, J.; Zhao, X. Selenium and Bacillus proteolyticus SES synergistically enhanced ryegrass to remediate Cu–Cd–Cr contaminated soil. Environ. Pollut. 2023, 323, 121272. [Google Scholar] [CrossRef]
- Zeng, Z.; He, X.; Li, F.; Zhang, Y.; Huang, Z.; Wang, Y.; Li, K.; Bao, Y.; Iqbal, M.; Fakhar-e-Alam Kulyar, M.; et al. Probiotic properties of Bacillus proteolyticus isolated from Tibetan yaks, China. Front. Microbiol. 2021, 12, 649207. [Google Scholar] [CrossRef]
- Ahmad, M.; Adil, Z.; Hussain, A.; Mumtaz, M.Z.; Nafees, M.; Ahmad, I.; Jamil, M. Potential of phosphate solubilizing Bacillus strains for improving growth and nutrient uptake in mungbean and maize crops. Pak. J. Agric. Sci. 2019, 56, 283–289. [Google Scholar] [CrossRef]
- Shen, F.T.; Yen, J.H.; Liao, C.S.; Chen, W.C.; Chao, Y.T. Screening of rice endophytic biofertilizers with fungicide tolerance and plant growth-promoting characteristics. Sustainability 2019, 11, 1133. [Google Scholar] [CrossRef]
- Park, J.M.; Park, S.J.; Kim, W.J.; Ghim, S.Y. Application of antifungal CFB to increase the durability of cement mortar. J. Microbiol. Biotechnol. 2012, 22, 1015–1020. [Google Scholar] [CrossRef]
- Delshadi, S.; Ebrahimi, M.; Shirmohammadi, E. Reducing Water Stress, Germination and Shoot Nutrients Accumulation of Avena sativa L. Following Application of Plant Growth-Promoting Bacteria. Commun. Soil Sci. Plant Anal. 2023, 55, 1175–1189. [Google Scholar] [CrossRef]
- Kálmán, C.D.; Nagy, Z.; Berényi, A.; Kiss, E.; Posta, K. Investigating PGPR bacteria for their competence to protect hybrid maize from the factor drought stress. Cereal Res. Commun. 2023, 52, 129–150. [Google Scholar] [CrossRef]
- Ahmed, B.; Shahid, M.; Syed, A.; Rajput, V.D.; Elgorban, A.M.; Minkina, T.; Bahkali, A.H.; Lee, J. Drought Tolerant Enterobacter sp./Leclercia adecarboxylata Secretes Indole-3-acetic Acid and Other Biomolecules and Enhances the Biological Attributes of Vigna radiata (L.) R. Wilczek in Water Deficit Conditions. Biology 2021, 10, 1149. [Google Scholar] [CrossRef]
- Wang, C.J.; Yang, W.; Wang, C.; Gu, C.; Niu, D.D.; Liu, H.X.; Wang, Y.P.; Guo, J.H. Induction of Drought Tolerance in Cucumber Plants by a Consortium of Three Plant Growth-Promoting Rhizobacterium Strains. PLoS ONE 2012, 7, e52565. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | Depth | C | MO | N | N | Ratio | P | K | Na | Al | Ca | Ca+Mg | pH |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (cm) | * g/kg | % | * g/kg | C/N | ** mg/dm3 | *** cmolc/dm3 | H2O | ||||||
| Summer | |||||||||||||
| Floodplain soil | 20 | 6.84 | 11.8 | 0.18 | 1.8 | 3.81 | 5 | 47 | 40 | 0.13 | 7.1 | 10.13 | 6.01 |
| Solid ground | 20 | 12.5 | 21.59 | 0.04 | 0.36 | 34.71 | 2 | 14 | 7 | 1.7 | 0.2 | 0.37 | 5.24 |
| Winter | |||||||||||||
| Floodplain soil | 20 | 14.8 | 25.46 | 0.03 | 0.31 | 47.79 | 9.59 | 65.3 | 63.7 | 0.11 | 5.7 | 9.54 | 5.18 |
| Solid ground | 20 | 15.3 | 26.46 | 0.01 | 0.1 | 153.4 | 10.47 | 22.7 | 19.7 | 1.6 | 0.2 | 0.8 | 4.78 |
| Strain | Sampling Area | Season | Cell Morphology (Gram) | IAA (µg/mL) | Organic Phosphate (MI) | Inorganic Phosphate (SI) | Siderophores (%US) | Cellulolytic Activity (EI) | Biofilm | ACC Deaminase | AMS Production | Water Stress Tolerance | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Curvularia | Colletotrichum | ||||||||||||
| AP1TV1 | Solid ground | Summer | Streptobacilli (+) | - | 14.04 bc | - | - | 12.58 a | - | + | + | + | Highly sensitive |
| AP1TV5 | Solid ground | Summer | Streptobacilli (+) | - | - | 7.67 c | - | 7.10 d | - | + | + | + | Highly tolerant |
| AP1TV7 | Solid ground | Summer | Streptobacilli (+) | - | 12.07 cde | - | - | 7.08 d | - | + | + | + | Sensitive |
| AP2TV1 | Solid ground | Summer | Streptobacilli (+) | - | 19.24 a | - | - | 8.44 cd | - | + | + | - | Sensitive |
| AP2TV4 | Solid ground | Summer | Diplobacilli (+) | - | 12.42 cde | 7.38 c | - | - | - | + | + | + | Tolerant |
| AP2TV5 | Solid ground | Summer | Diplobacilli (+) | 2.70 ab | - | 7.52 c | - | 8.89 bc | - | + | + | + | Tolerant |
| AP2TV8 | Solid ground | Summer | Bacilli (+) | 0.98 c | - | - | 96.56 a | 7.73 cd | - | + | + | + | Sensitive |
| AP3TV2 | Solid ground | Summer | Bacilli (+) | - | - | - | - | 7.78 cd | + | + | + | + | Highly tolerant |
| AP3TV5 | Solid ground | Summer | Bacilli (+) | - | 16.62 ab | - | - | 7.10 d | - | - | + | + | Sensitive |
| AP4TV1 | Solid ground | Summer | Diplobacilli (+) | 2.09 bc | - | - | 96.56 a | - | - | - | + | + | Sensitive |
| AP4TV3 | Solid ground | Summer | Bacilli (+) | 1.21 c | - | - | - | 9.92 b | - | + | + | + | Sensitive |
| AP4TV4 | Solid ground | Summer | Bacilli (-) | 0.92 c | 7.81 f | - | - | - | - | - | + | - | Highly sensitive |
| AP1VV1 | Floodplain | Summer | Streptobacilli (+) | 1.92 bc | - | 7.38 c | - | - | - | + | + | + | Highly tolerant |
| AP1VV7 | Floodplain | Summer | Diplobacilli (+) | 4.00 a | - | 11.01 a | - | 9.97 b | + | + | + | - | Tolerant |
| AP3VV1 | Floodplain | Summer | Streptobacilli (+) | 1.80 bc | - | - | - | - | + | + | + | - | Highly sensitive |
| AP3VV9 | Floodplain | Summer | Streptobacilli (+) | - | 11.67 cde | 7.83 c | - | - | - | + | + | + | Tolerant |
| AP4VV4 | Floodplain | Summer | Coccos (-) | 1.24 c | 12.83 cd | - | - | - | - | - | + | - | Highly sensitive |
| AP4VV5 | Floodplain | Summer | Streptobacilli (+) | - | 10.24 def | - | - | 7.78 cd | - | - | + | - | Highly sensitive |
| AP4VVA3 | Floodplain | Summer | Coccos (-) | 1.55 bc | - | 10.57 a | 96.13 a | 7.67 cd | - | + | - | - | Highly tolerant |
| AP2TI5 | Solid ground | Winter | Coccobacillus (-) | - | 10.07 def | 10.11 ab | 96.20 a | - | + | + | - | - | Tolerant |
| AP1VI3 | Floodplain | Winter | Coccobacillus (-) | 1.86 bc | 9.28 ef | 9.17 b | 96.60 a | - | - | + | + | - | Highly tolerant |
| AP2VI7 | Floodplain | Winter | Coccobacillus (-) | - | 10.08 def | 10.96 a | 94.86 b | - | + | + | + | - | Sensitive |
| Treatment | Germination % | ||
|---|---|---|---|
| 0 MPa | −0.5 MPa | −1.0 MPa | |
| Not inoculated | 95.00 ± 0.96 Aa | 80.00 ± 2.16 Ba | 10.00 ± 0.58 Bb |
| AP2TV5 | 100.00 ± 0.00 Aa | 100.00 ± 0.00 Aa | 83.00 ± 1.29 Ab |
| AP1VV7 | 100.00 ± 0.00 Aa | 93.00 ± 0.81 ABa | 63.00 ± 2.38 Ab |
| Association | 100.00 ± 0.00 Aa | 98.00 ± 0.50 Aab | 85.00 ± 1.70 Ab |
| Treatment | Root Length (cm) | Seedling Length (cm) | Vigor Index | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 MPa | −0.5 MPa | −1.0 MPa | 0 MPa | −0.5 MPa | −1.0 MPa | 0 MPa | −0.5 MPa | −1.0 MPa | |
| Not inoculated | 2.92 ± 1.09 Aa | 2.42 ± 0.98 Ba | 0.50 ± 0.08 Cb | 2.90 ± 0.17 Ba | 2.03 ± 0.05 Ba | 0.90 ± 0.10 Bb | 275.5 | 162.4 | 9.0 |
| AP2TV5 | 4.22 ± 1.78 Aa | 4.15 ± 0.44 Aa | 1.27 ± 0.48 Bb | 5.33 ± 0.11 Aa | 4.37 ± 0.15 Aa | 2.17 ± 0.15 Ab | 533.0 | 437.0 | 157.7 |
| AP1VV7 | 4.22 ± 0.22 Aa | 5.27 ± 1.05 Aa | 2.12 ± 0.30 Ab | 5.30 ± 0.26 Aa | 4.17 ± 0.15 Aa | 1.90 ± 0.10 Ab | 530.0 | 387.8 | 136.7 |
| Association | 5.05 ± 0.90 Aa | 4.70 ± 0.48 Aa | 1.65 ± 0.24 Ab | 5.10 ± 0.10 Aa | 4.33 ± 0.15 Aa | 2.20 ± 0.09 Ab | 510.0 | 424.3 | 187.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, S.M.d.N.; Garcias, J.T.; Farias, M.R.D.O.; Lima, A.L.A.; Sousa, R.d.S.d.R.d.; Philippsen, H.K.; Madeira, L.D.P.d.S.; Rogez, H.; Marques, J.M. Rhizobacteria Isolated from Amazonian Soils Reduce the Effects of Water Stress on the Growth of Açaí (Euterpe oleracea Mart.) Palm Seedlings. Biology 2024, 13, 757. https://doi.org/10.3390/biology13100757
Sousa SMdN, Garcias JT, Farias MRDO, Lima ALA, Sousa RdSdRd, Philippsen HK, Madeira LDPdS, Rogez H, Marques JM. Rhizobacteria Isolated from Amazonian Soils Reduce the Effects of Water Stress on the Growth of Açaí (Euterpe oleracea Mart.) Palm Seedlings. Biology. 2024; 13(10):757. https://doi.org/10.3390/biology13100757
Chicago/Turabian StyleSousa, Suania Maria do Nascimento, Josinete Torres Garcias, Marceli Ruani De Oliveira Farias, Allana Laís Alves Lima, Rosiane do Socorro dos Reis de Sousa, Hellen Kempfer Philippsen, Lucimar Di Paula dos Santos Madeira, Herve Rogez, and Joana Montezano Marques. 2024. "Rhizobacteria Isolated from Amazonian Soils Reduce the Effects of Water Stress on the Growth of Açaí (Euterpe oleracea Mart.) Palm Seedlings" Biology 13, no. 10: 757. https://doi.org/10.3390/biology13100757
APA StyleSousa, S. M. d. N., Garcias, J. T., Farias, M. R. D. O., Lima, A. L. A., Sousa, R. d. S. d. R. d., Philippsen, H. K., Madeira, L. D. P. d. S., Rogez, H., & Marques, J. M. (2024). Rhizobacteria Isolated from Amazonian Soils Reduce the Effects of Water Stress on the Growth of Açaí (Euterpe oleracea Mart.) Palm Seedlings. Biology, 13(10), 757. https://doi.org/10.3390/biology13100757