
Population Genetic Assessment Model Reveals Conservation Priorities for Gymnocypris Species Resources on the Qinghai-Tibetan Plateau

Abstract
Simple Summary
Abstract
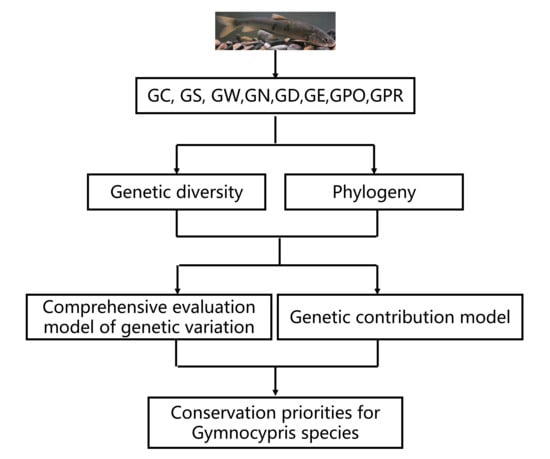
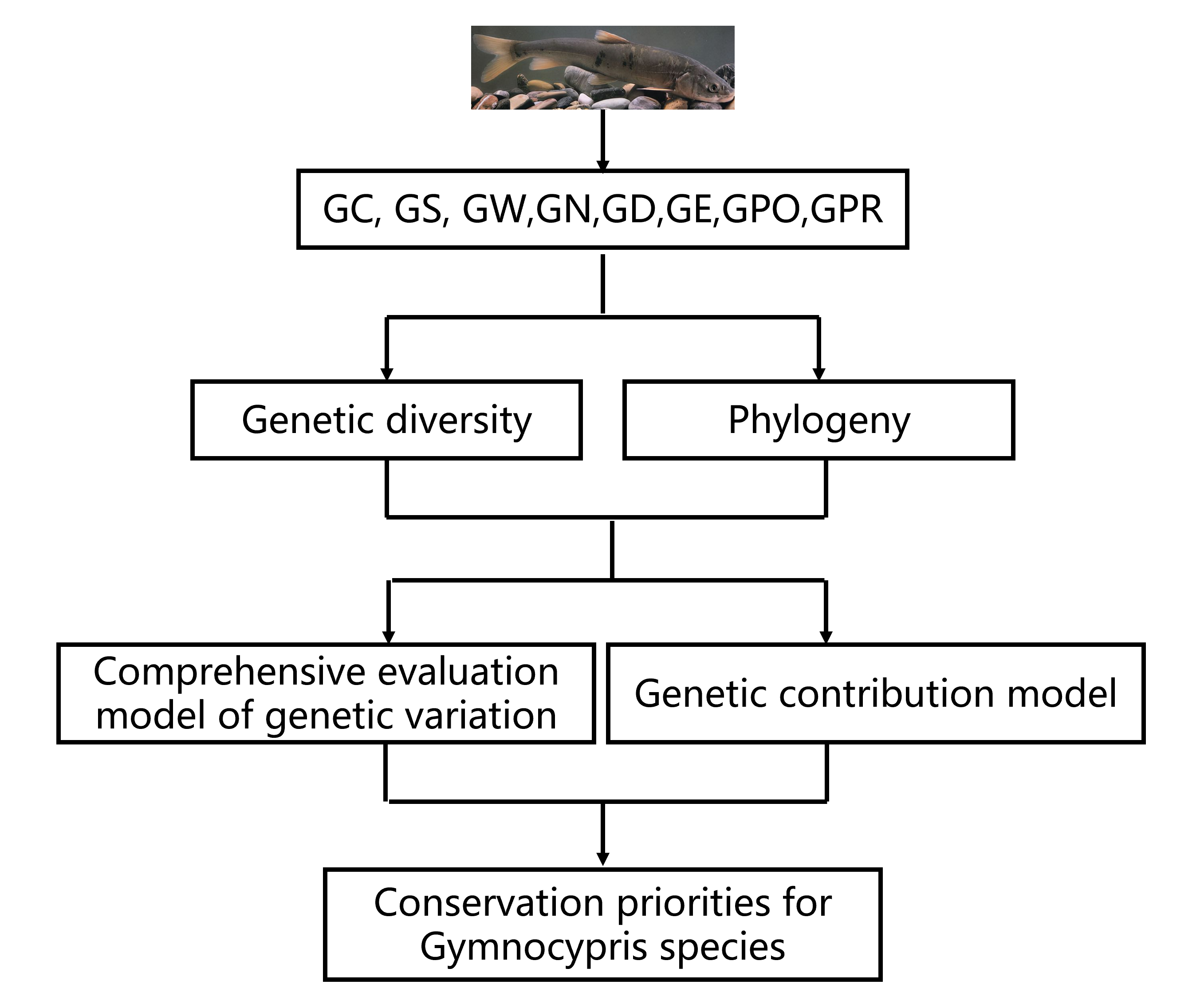
1. Introduction
2. Material and Methods
2.1. Animal and Sample Collection
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Data Analysis
3. Results
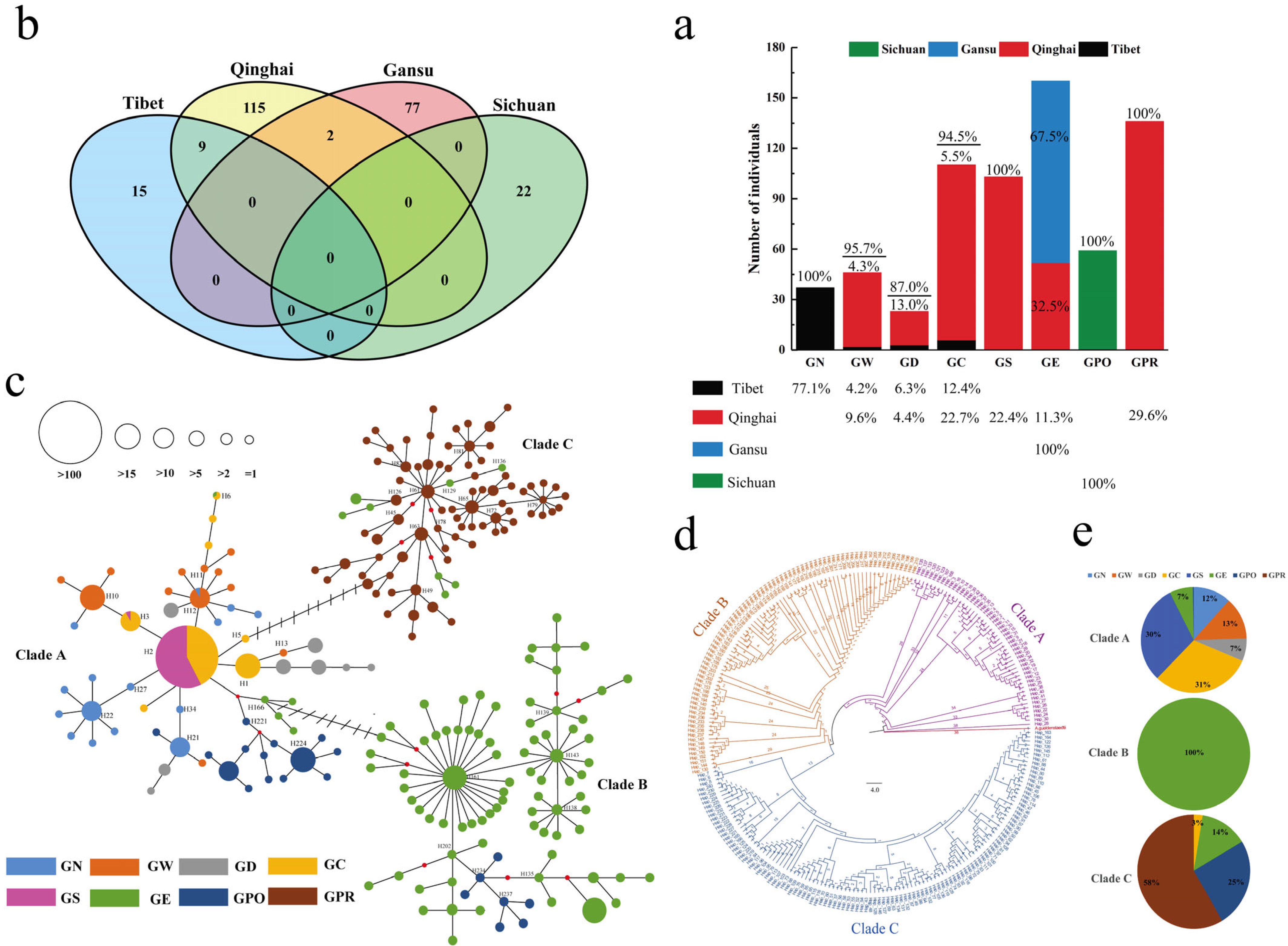
3.1. Genetic Diversity Analysis
3.2. Distribution, Expansion, and Phylogenetic Relationships of Populations
3.3. Establishment of a Comprehensive Genetic Diversity Evaluation Model
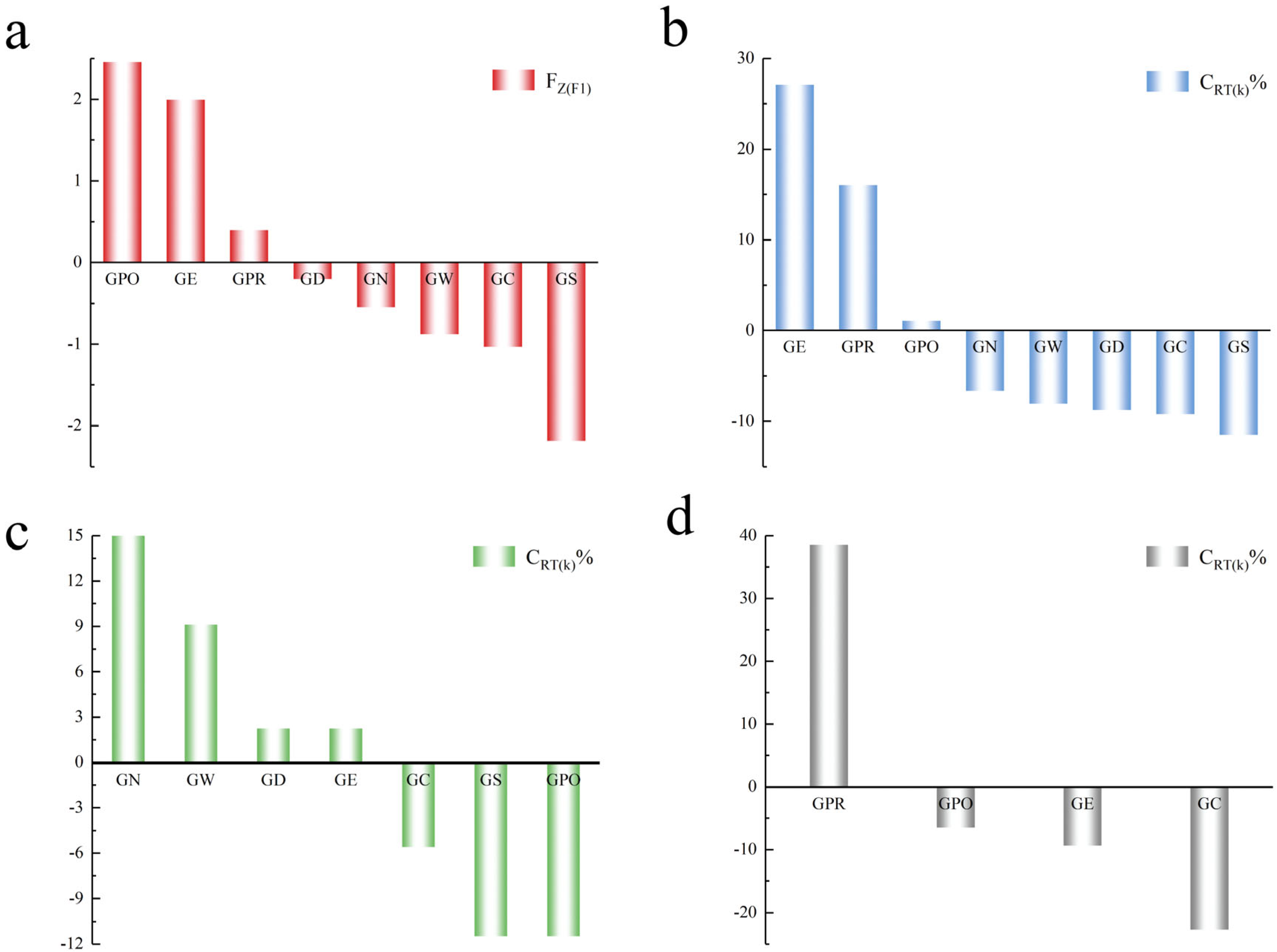
3.4. Establishment of a Genetic Contribution Rate Assessment Model
4. Discussion
4.1. Assessment of Gymnocypris Species for Priority Conservation on the Basis of Genetic Diversity
4.2. Model for Assessing Genetic Contribution on the Basis of Phylogeny and Genetic Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feng, X.; Jia, Y.; Zhu, R.; Chen, K.; Chen, Y. Characterization and analysis of the transcriptome in Gymnocypris selincuoensis on the Qinghai-Tibetan Plateau using single-molecule long-read sequencing and RNA-seq. DNA Res. 2019, 26, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Favre, A.; Packert, M.; Pauls, S.; Jahnig, S.; Uhl, D.; Michalak, I.; Muellner, A. The role of the uplift of the Qinghai-Tibetan Plateau for the evolution of Tibetan biotas. Biol. Rev. Camb. Philos. Soc. 2015, 90, 236–253. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Hoffmann, D.; Aldenderfer, M.; Haas, W.; Dahl, J.; Wang, Z.; Degering, D.; Schlütz, F. Permanent human occupation of the central Tibetan Plateau in the early Holocene. Science 2017, 355, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.J.; Yang, J.; Xie, X.L.; Lv, F.H.; Cao, Y.H.; Li, W.R.; Liu, M.J.; Wang, Y.T.; Li, J.Q.; Liu, Y.G.; et al. The Genome Landscape of Tibetan Sheep Reveals Adaptive Introgression from Argali and the History of Early Human Settlements on the Qinghai-Tibetan Plateau. Mol. Biol. Evol. 2019, 36, 283–303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yu, M.; Hu, P.; Peng, S.; Liu, Y.; Li, W.; Wang, C.; He, S.; Zhai, W.; Xu, Q.; et al. Genetic Adaptation of Schizothoracine Fish to the Phased Uplifting of the Qinghai-Tibetan Plateau. G3 Genes Genomes Genet. 2017, 7, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Zhao, G.; Li, L.; Zhang, J.; Luo, Z.; Kang, Y.; Liu, Z. Phylogeny and genetic diversity reveal the influence of Qinghai-Tibet Plateau uplift on the divergence and distribution of Gymnocypris species. Aquat. Sci. 2021, 83, 6. [Google Scholar] [CrossRef]
- Zhao, K.; Duan, Z.; Peng, Z.; Gan, X.; Zhang, R.; He, S.; Zhao, X. Phylogeography of the endemic Gymnocypris chilianensis (Cyprinidae): Sequential westward colonization followed by allopatric evolution in response to cyclical Pleistocene glaciations on the Tibetan Plateau. Mol. Phylogenet Evol. 2011, 59, 303–310. [Google Scholar] [CrossRef]
- Zhao, K.; Duan, Z.Y.; Peng, Z.G.; Guo, S.C.; Li, J.B.; He, S.P.; Zhao, X.Q. The youngest split in sympatric schizothoracine fish (Cyprinidae) is shaped by ecological adaptations in a Tibetan Plateau glacier lake. Mol. Ecol. 2009, 18, 3616–3628. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, L.; Liu, D.; Gao, Q.; Nie, M.; Zhu, S.; Chao, Y.; Yang, C.; Zhang, C.; Yi, R.; et al. Chromosome-level assembly of Gymnocypris eckloni genome. Sci. Data 2022, 9, 464. [Google Scholar] [CrossRef]
- Chi, W.; Ma, X.; Niu, J.; Zou, M. Genome-wide identification of genes probably relevant to the adaptation of schizothoracins (Teleostei: Cypriniformes) to the uplift of the Qinghai-Tibet Plateau. BMC Genom. 2017, 18, 310. [Google Scholar] [CrossRef]
- Mamiatis, T.; Fritsch, E.; Sambrook, J.; Engel, J. Molecular cloning—A laboratory manual. New York: Cold spring harbor laboratory 2010 1982, 545s. Acta Biotechnol. 2010, 5, 104. [Google Scholar]
- Petit, R.; Mousadik, A.; Pons, O. Identifying populations for conservation on the basis of genetic markers. Conserv. Biol. 1998, 12, 844–855. [Google Scholar] [CrossRef]
- Quan, J.; Gao, C.; Cai, Y.; Ge, Q.; Jiao, T.; Zhao, S. Population genetics assessment model reveals priority protection of genetic resources in native pig breeds in China. Glob. Ecol. Conserv. 2020, 21, e00829. [Google Scholar]
- Cao, Y.B.; Chen, X.Q.; Wang, S.; Wang, Y.X.; Du, J.Z. Evolution and regulation of the downstream gene of hypoxia-inducible factor-1alpha in naked carp (Gymnocypris przewalskii) from Lake Qinghai, China. J. Mol. Evol. 2008, 67, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jiang, P.; Gang, H.; Shi, J.Q.; Sun, X. Development and characterization of microsatellite markers to study population genetic diversity of Przewalski’s naked carp Gymnocypris przewalskii in China. J. Fish. Biol. 2009, 75, 2352–2356. [Google Scholar] [CrossRef]
- Quan, J.; Li, Y.; Yang, Y.; Yang, T.; Sha, Y.; Cai, Y.; Jiao, T.; Wu, J.; Zhao, S. Population genetic diversity and genetic evaluation models reveal the maternal genetic structure and conservation priority characteristics of indigenous cattle in China. Glob. Ecol. Conserv. 2021, 32, e01903. [Google Scholar] [CrossRef]
- Hu, J.; Cai, Y.; Xiong, S.; He, Z.; Li, S.; Wang, X.; He, Z.; Yan, T. The complete mitochondrial DNA of Gymnocypris potanini firmispinatus and comparative mitogenomic analyses of the Gymnocypris species. Mitochondrial DNA B Resour. 2016, 1, 436–437. [Google Scholar] [CrossRef] [PubMed]
- Kai, Z.; Ziyuan, D.; Gongshe, Y.; Zuogang, P.; Shunping, H.; Yiyu, C. Origin and population evolution of the nudibranchs of Qinghai Lake. Adv. Nat. Sci. 2007, 3, 320–328, (In Chinese with English abstract). [Google Scholar]
- Chen, J.; Yang, L.; Zhang, R.; Uebbing, S.; Zhang, C.; Jiang, H.; Lei, Y.; Lv, W.; Tian, F.; Zhao, K.; et al. Transcriptome-Wide Patterns of the Genetic and Expression Variations in Two Sympatric Schizothoracine Fishes in a Tibetan Plateau Glacier Lake. Genome Biol. Evol. 2020, 12, 3725–3737. [Google Scholar] [CrossRef]
- Wang, C.; Shi, J.; Wei, F.; Qi, D.; Shi, Y.; Zhang, Y.; Qi, H.; Yang, M.; Qin, Q.; Li, C. AFLP genetic diversity analysis of Qinghai Lake nudibranchs in different tributaries of Qinghai Lake. Genom. Appl. Biol. 2015, 34, 82–89, In Chinese with English abstract. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, J.; Wang, W.; Zhou, J.; Pan, Y.; Mou, Z. The complete mitochondrial genome of the Gymnocypris scleracanthus (Cypriniformes: Cyprinidae). Mitochondrial DNA B Resour. 2017, 2, 634–635. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; He, S.; Peng, Z.; Li, J. Population structure and mitochondrial DNA variation in the naked carp (Cyprinus carpio) from Qinghai Lake. J. Qinghai Univ. Nat. Sci. Ed. 2006, 24, 1–4+9, (In Chinese with English abstract). [Google Scholar]
- Chen, D.Q.; Zhang, C.H.L.; Lu, C.; Zhang, X. Sequence polymorphism in the D-loop region of the mitochondrial genome of the Qinghai Lake naked carp breeding population. Chin. Aquat. Sci. 2006, 13, 800–806, (In Chinese with English abstract). [Google Scholar]
- Tang, L.; Duan, X.; Kong, F.; Zhang, F.; Zheng, Y.; Li, Z.; Mei, Y.; Zhao, Y.; Hu, S. Influences of climate change on area variation of Qinghai Lake on Qinghai-Tibetan Plateau since 1980s. Sci. Rep. 2018, 8, 7331. [Google Scholar] [CrossRef]
- MORITZC Defining“evolutionary significant units”for conservation. Trends Ecol. Evol. 1994, 9, 373–375. [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Abbreviation | S | H | Nps | Pis | Svs | Hd | Pi | K |
|---|---|---|---|---|---|---|---|---|
| GC | 110 | 8 | 20 | 19 | 1 | 0.500 ± 0.053 | 0.00307 | 2.013 |
| GS | 103 | 2 | 1 | 0 | 1 | 0.019 ± 0.019 | 0.00003 | 0.019 |
| GW | 44 | 12 | 12 | 6 | 6 | 0.718 ± 0.058 | 0.00187 | 1.227 |
| GN | 39 | 15 | 15 | 6 | 9 | 0.833 ± 0.043 | 0.00295 | 1.938 |
| GD | 23 | 8 | 9 | 8 | 1 | 0.858 ± 0.044 | 0.00511 | 3.352 |
| GPR | 136 | 77 | 86 | 35 | 51 | 0.985 ± 0.003 | 0.00795 | 5.207 |
| GE | 159 | 100 | 140 | 65 | 75 | 0.986 ± 0.004 | 0.01907 | 12.452 |
| GPO | 60 | 24 | 74 | 62 | 12 | 0.880 ± 0.030 | 0.02333 | 15.305 |
| Population | GC | GS | GW | GN | GD | GPR | GE | GPO |
|---|---|---|---|---|---|---|---|---|
| GC | 0.003 | |||||||
| GS | 0.002 | 0 | ||||||
| GW | 0.004 | 0.003 | 0.002 | |||||
| GN | 0.005 | 0.003 | 0.003 | 0.003 | ||||
| GD | 0.007 | 0.006 | 0.005 | 0.006 | 0.005 | |||
| GPR | 0.033 | 0.034 | 0.031 | 0.033 | 0.031 | 0.008 | ||
| GE | 0.024 | 0.024 | 0.024 | 0.025 | 0.025 | 0.028 | 0.02 | |
| GPO | 0.031 | 0.031 | 0.031 | 0.033 | 0.034 | 0.04 | 0.036 | 0.023 |
| Population | GC | GS | GW | GN | GD | GPR | GE | GPO |
|---|---|---|---|---|---|---|---|---|
| GC | 0.0172 | 0.0598 | 0.0585 | 0.0596 | 0.0346 | 0.0624 | 0.0612 | |
| GS | 0.0743 | 0.0553 | 0.0618 | 0.0619 | 0.0261 | 0.0610 | 0.0589 | |
| GW | 0.3961 | 0.6695 | 0.0494 | 0.0574 | 0.0328 | 0.0620 | 0.0595 | |
| GN | 0.3738 | 0.5529 | 0.2713 | 0.0547 | 0.0342 | 0.0620 | 0.0595 | |
| GD | 0.3917 | 0.5504 | 0.3569 | 0.3239 | 0.0415 | 0.0625 | 0.0607 | |
| GPR | 0.8338 | 0.8817 | 0.8446 | 0.8366 | 0.7901 | 0.0625 | 0.0596 | |
| GE | 0.5153 | 0.5775 | 0.5439 | 0.5469 | 0.5024 | 0.4982 | 0.0602 | |
| GPO | 0.5718 | 0.6198 | 0.5990 | 0.6088 | 0.5845 | 0.6071 | 0.4044 |
| Item | Hd | Pi | K |
|---|---|---|---|
| Hd | 1.000 | ||
| Pi | 0.568 ** | 1.000 | |
| K | 0.568 ** | 1.000 ** | 1.000 |
| Population | RS(k) | RD(k) | RT(k) | CRS(k)% | CRD(k) % | CRT(k) % |
|---|---|---|---|---|---|---|
| GN | 1.000 | 12.625 | 13.625 | −1.1560 | −5.5132 | −6.6692 |
| GW | 0.250 | 10.000 | 10.250 | −1.4609 | −6.5803 | −8.0412 |
| GD | 1.500 | 7.000 | 8.500 | −0.9527 | −7.7998 | −8.7525 |
| GC | 1.875 | 5.500 | 7.375 | −0.8003 | −8.4096 | −9.2099 |
| GS | 1.000 | 0.750 | 1.750 | −1.1560 | −10.3404 | −11.4964 |
| GE | 9.625 | 87.000 | 96.625 | 2.3501 | 24.7205 | 27.0706 |
| GPO | 12.500 | 20.125 | 32.625 | 3.5188 | −2.4644 | 1.0544 |
| GPR | 3.000 | 66.500 | 69.500 | −0.3430 | 16.3872 | 16.0442 |
| Population | Clade A | Clade C | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RS(k) | RD(k) | RT(k) | CRS(k)% | CRD(k) % | CRT(k) % | RS(k) | RD(k) | RT(k) | CRS(k)% | CRD(k) % | CRT(k) % | |
| GN | 2.143 | 12.357 | 14.500 | 2.1609 | 12.8252 | 14.9861 | ||||||
| GW | 1.714 | 9.786 | 11.500 | 1.3206 | 7.7832 | 9.1037 | ||||||
| GD | 1.143 | 6.857 | 8.000 | 0.2001 | 2.0409 | 2.2410 | ||||||
| GC | 0.714 | 3.286 | 4.000 | −0.6402 | −4.9619 | −5.6022 | 0.750 | 1.750 | 2.500 | −5.6250 | −17.0833 | −22.7083 |
| GS | 0.286 | 0.714 | 1.000 | −1.4806 | −10.0039 | −11.4845 | ||||||
| GE | 1.143 | 6.857 | 8.000 | 0.2001 | 2.0409 | 2.2410 | 4.750 | 13.750 | 18.500 | −2.2917 | −7.0833 | −9.3750 |
| GPO | 0.143 | 0.857 | 1.000 | −1.7607 | −9.7238 | −11.4845 | 5.500 | 16.500 | 22.000 | −1.6667 | −4.7917 | −6.4583 |
| GPR | 19.000 | 57.000 | 76.000 | 9.5833 | 28.9583 | 38.5417 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quan, J.; Qu, Y.; Li, Y.; Ren, Y.; Zhao, G.; Li, L.; Lu, J. Population Genetic Assessment Model Reveals Conservation Priorities for Gymnocypris Species Resources on the Qinghai-Tibetan Plateau. Biology 2024, 13, 259. https://doi.org/10.3390/biology13040259
Quan J, Qu Y, Li Y, Ren Y, Zhao G, Li L, Lu J. Population Genetic Assessment Model Reveals Conservation Priorities for Gymnocypris Species Resources on the Qinghai-Tibetan Plateau. Biology. 2024; 13(4):259. https://doi.org/10.3390/biology13040259
Chicago/Turabian StyleQuan, Jinqiang, Yuling Qu, Yongqing Li, Yue Ren, Guiyan Zhao, Lanlan Li, and Junhao Lu. 2024. "Population Genetic Assessment Model Reveals Conservation Priorities for Gymnocypris Species Resources on the Qinghai-Tibetan Plateau" Biology 13, no. 4: 259. https://doi.org/10.3390/biology13040259
APA StyleQuan, J., Qu, Y., Li, Y., Ren, Y., Zhao, G., Li, L., & Lu, J. (2024). Population Genetic Assessment Model Reveals Conservation Priorities for Gymnocypris Species Resources on the Qinghai-Tibetan Plateau. Biology, 13(4), 259. https://doi.org/10.3390/biology13040259