
Impacts of Invasive Plants on Native Vegetation Communities in Wetland and Stream Mitigation

Abstract
Simple Summary
Abstract
1. Introduction
1.1. Invasive Species Performance Standards in Compensatory Mitigation
1.2. Mitigation Plant Communities and Invasive Species
2. Materials and Methods
2.1. Target Invaders
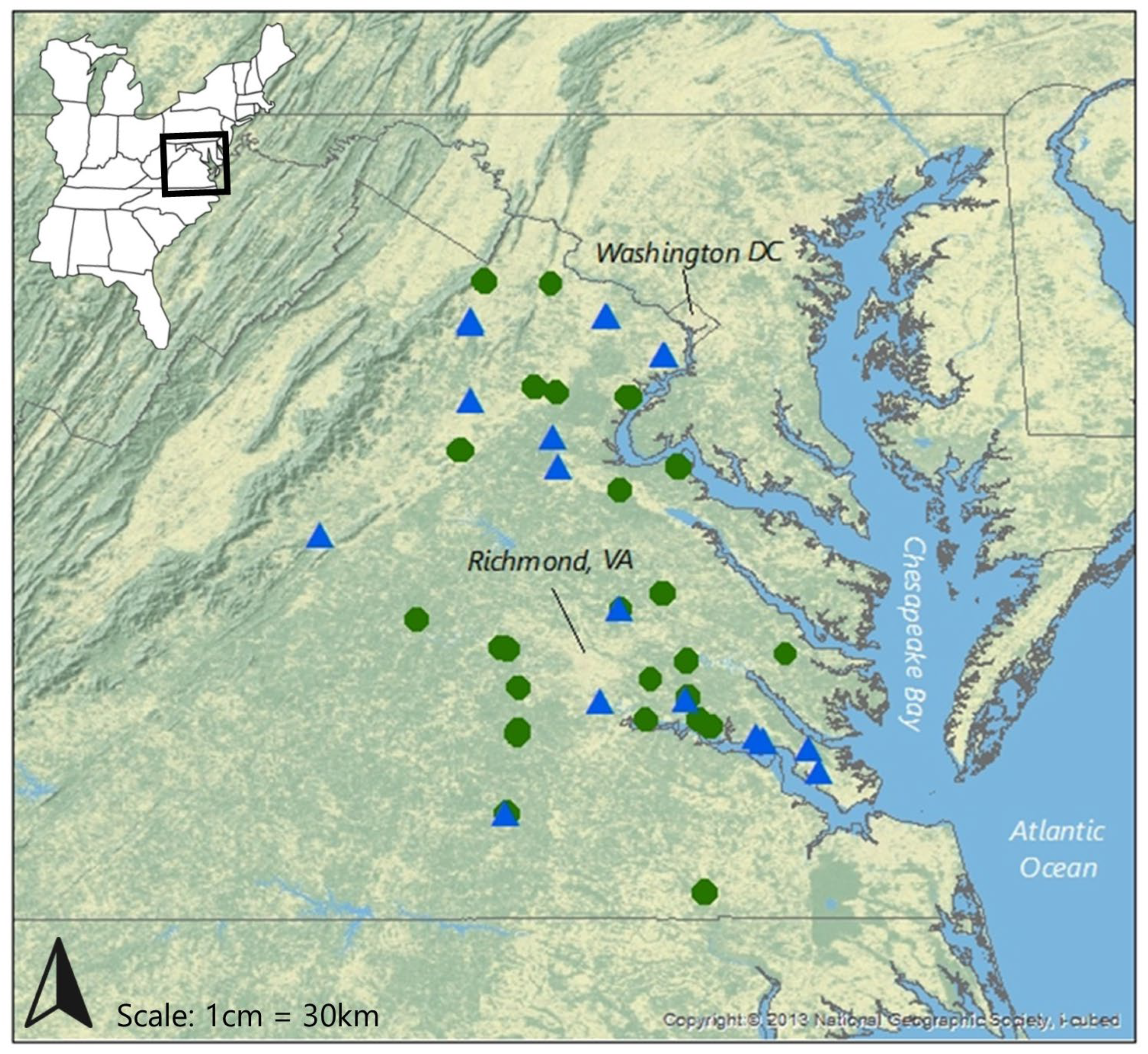
2.2. Study Sites
2.3. Vegetation Sampling
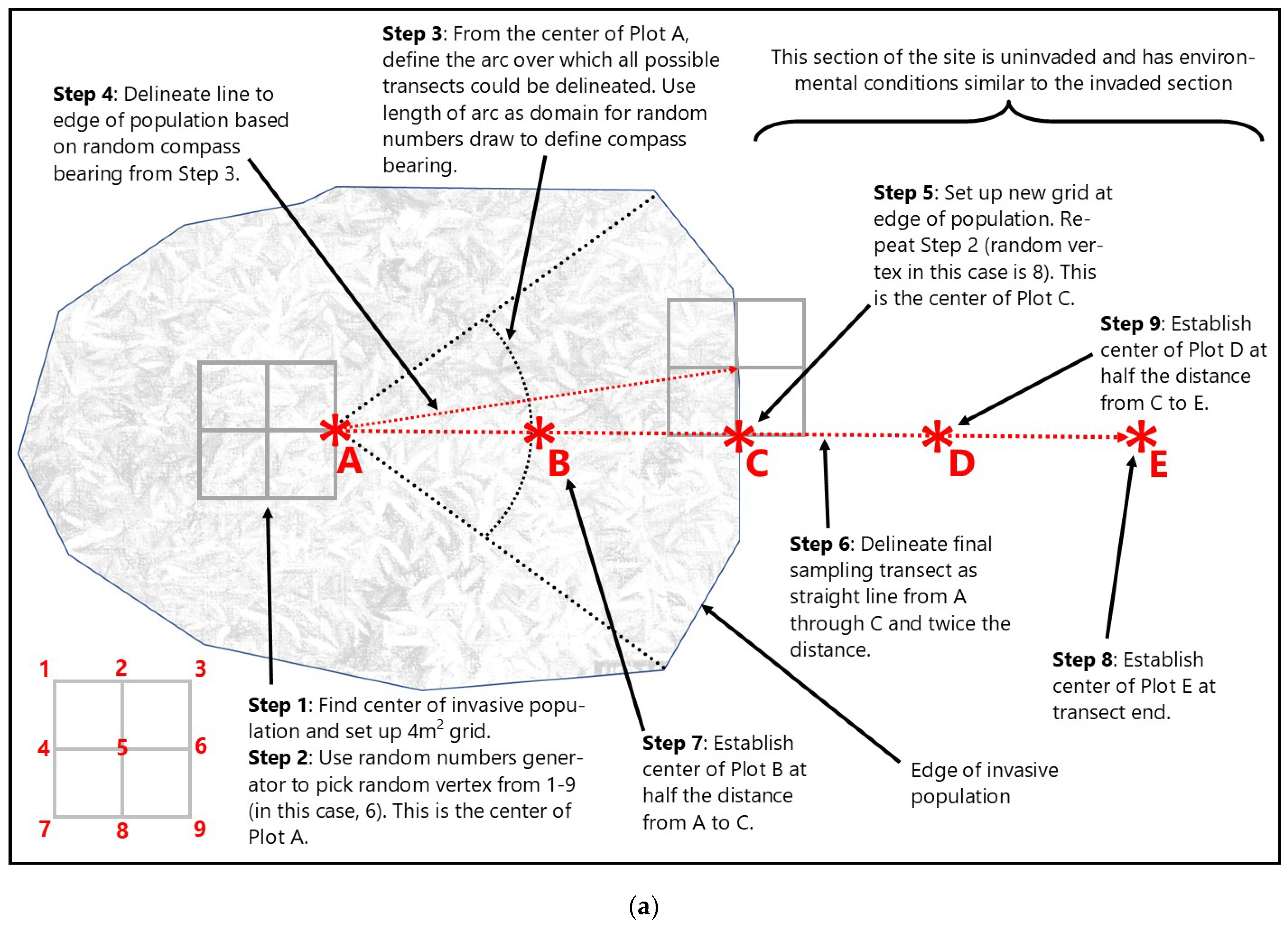
2.3.1. Wetland Mitigation Sampling Methods
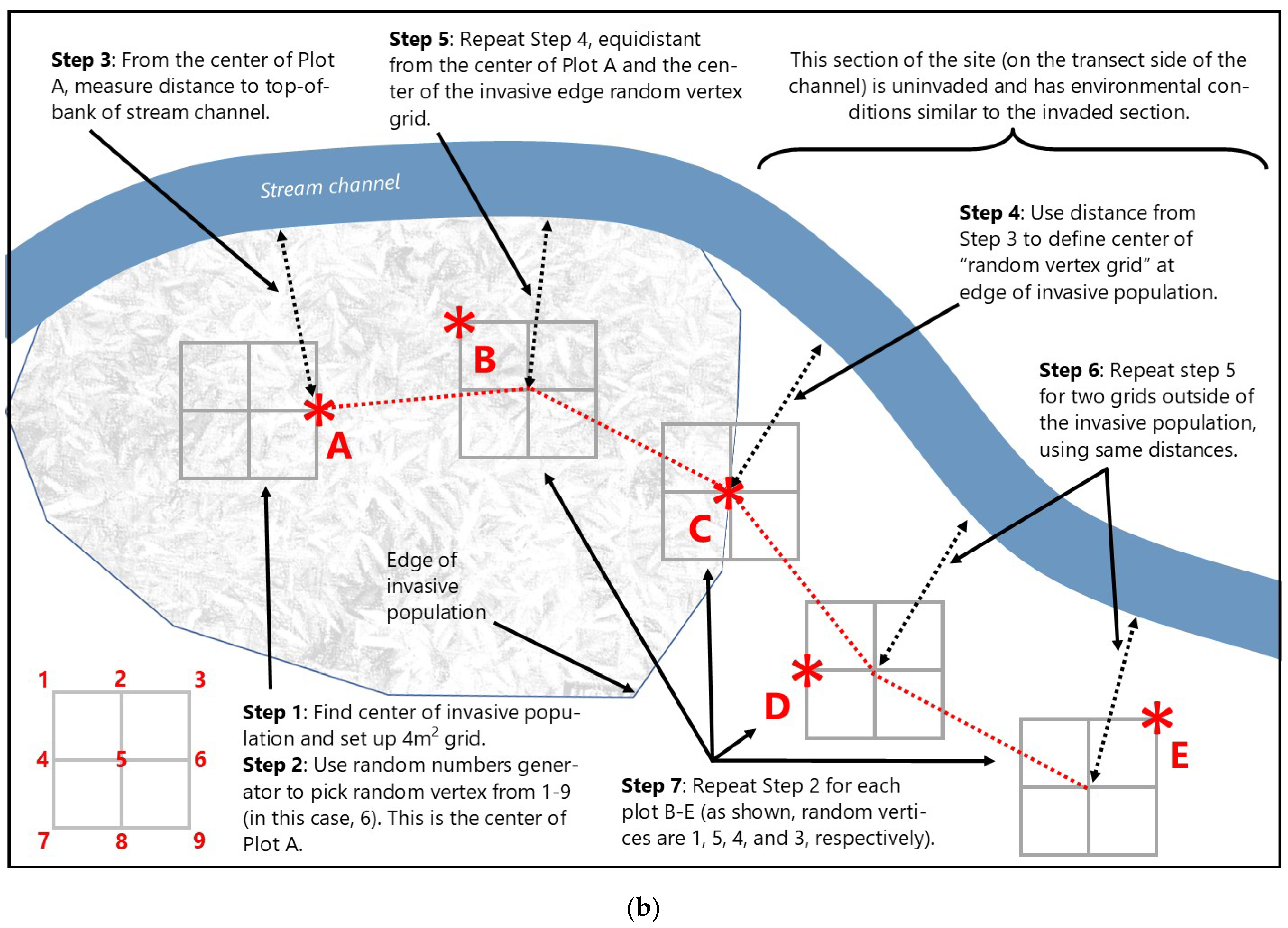
2.3.2. Stream Mitigation Sampling Methods
2.4. Data Synthesis and Analysis
2.5. Analysis for Ecological Performance Standard
3. Results
3.1. Wetland Study
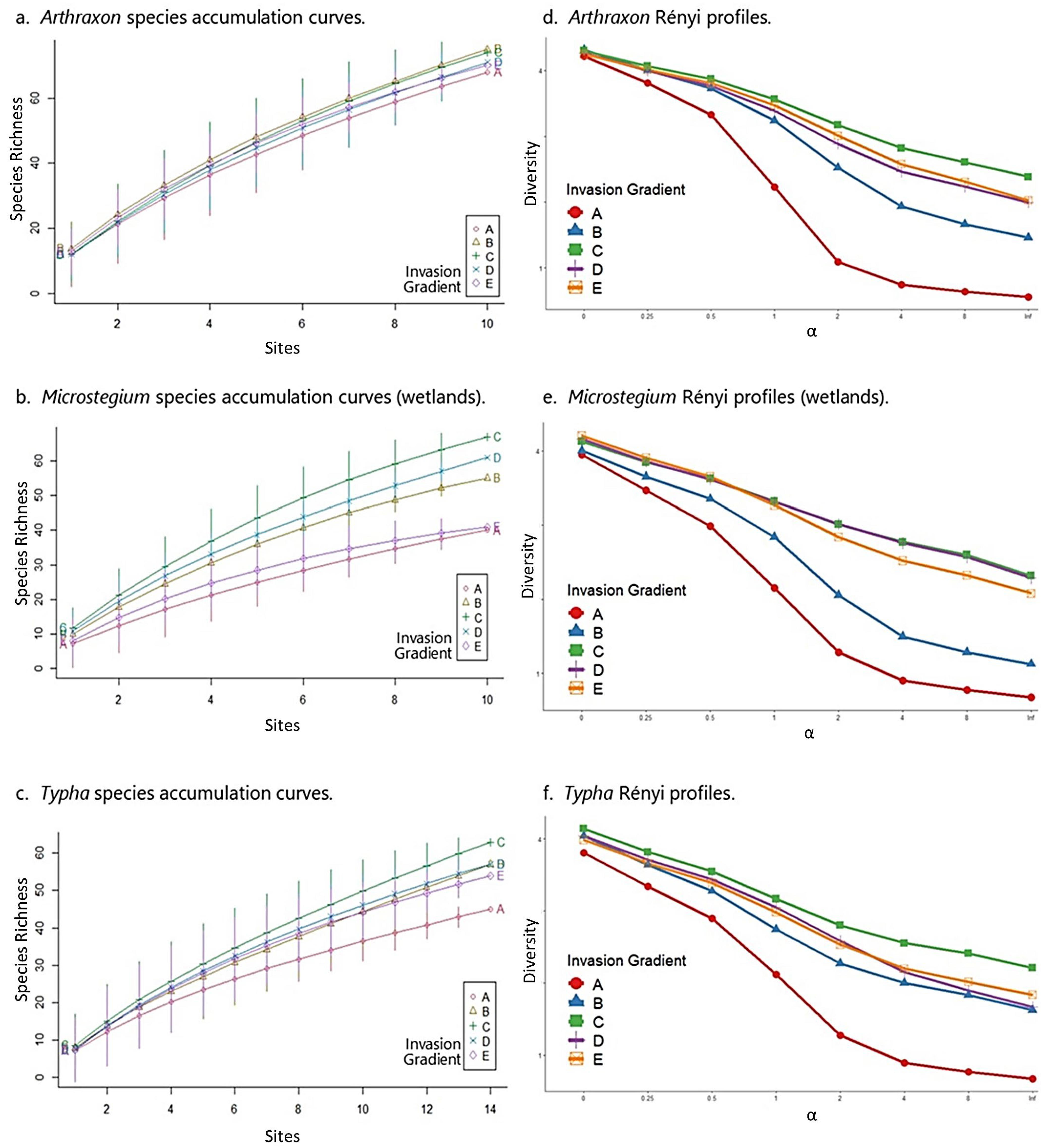
3.2. Species Composition—Wetlands
3.3. Community Properties—Wetlands
3.4. Stream Study
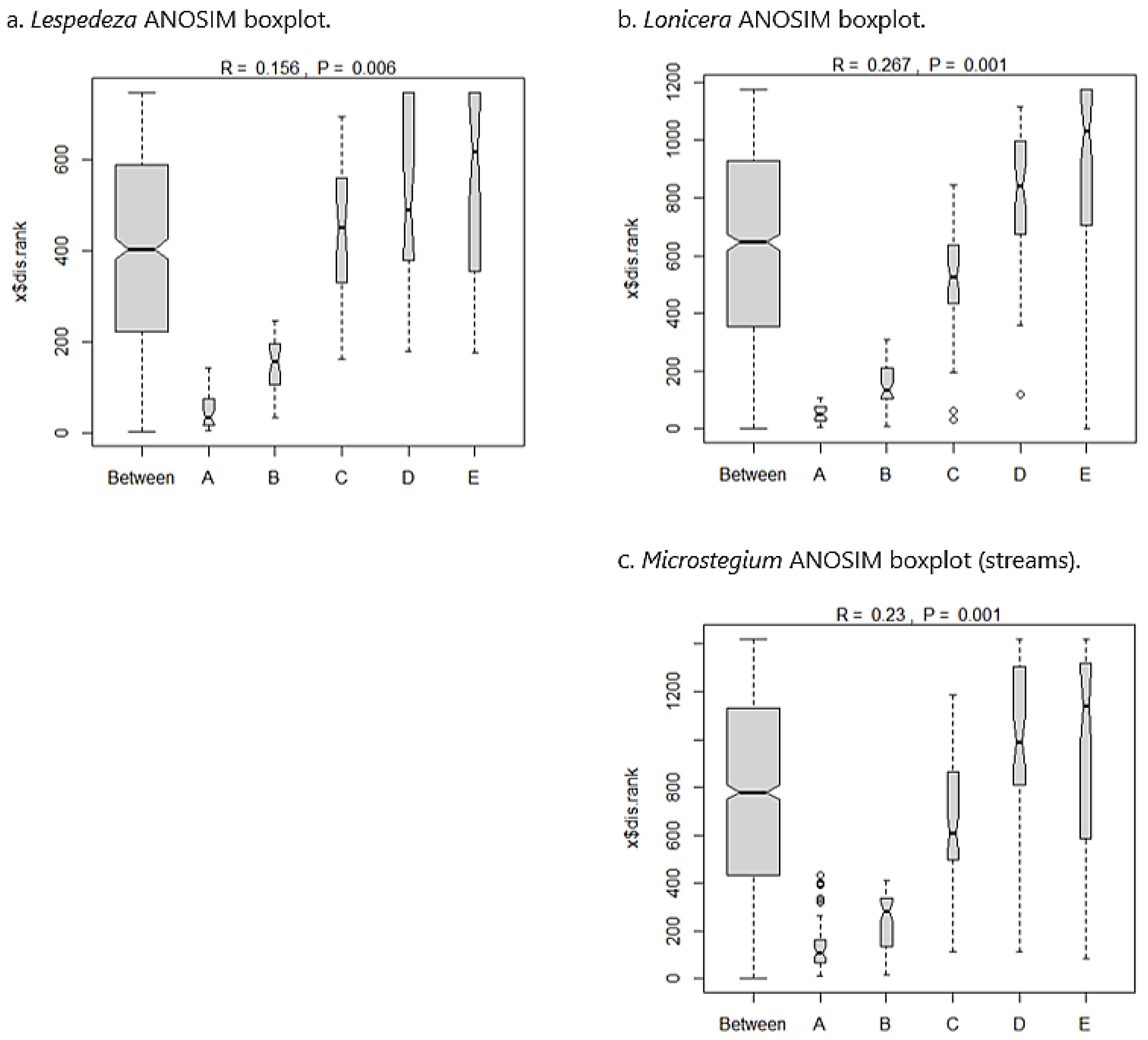
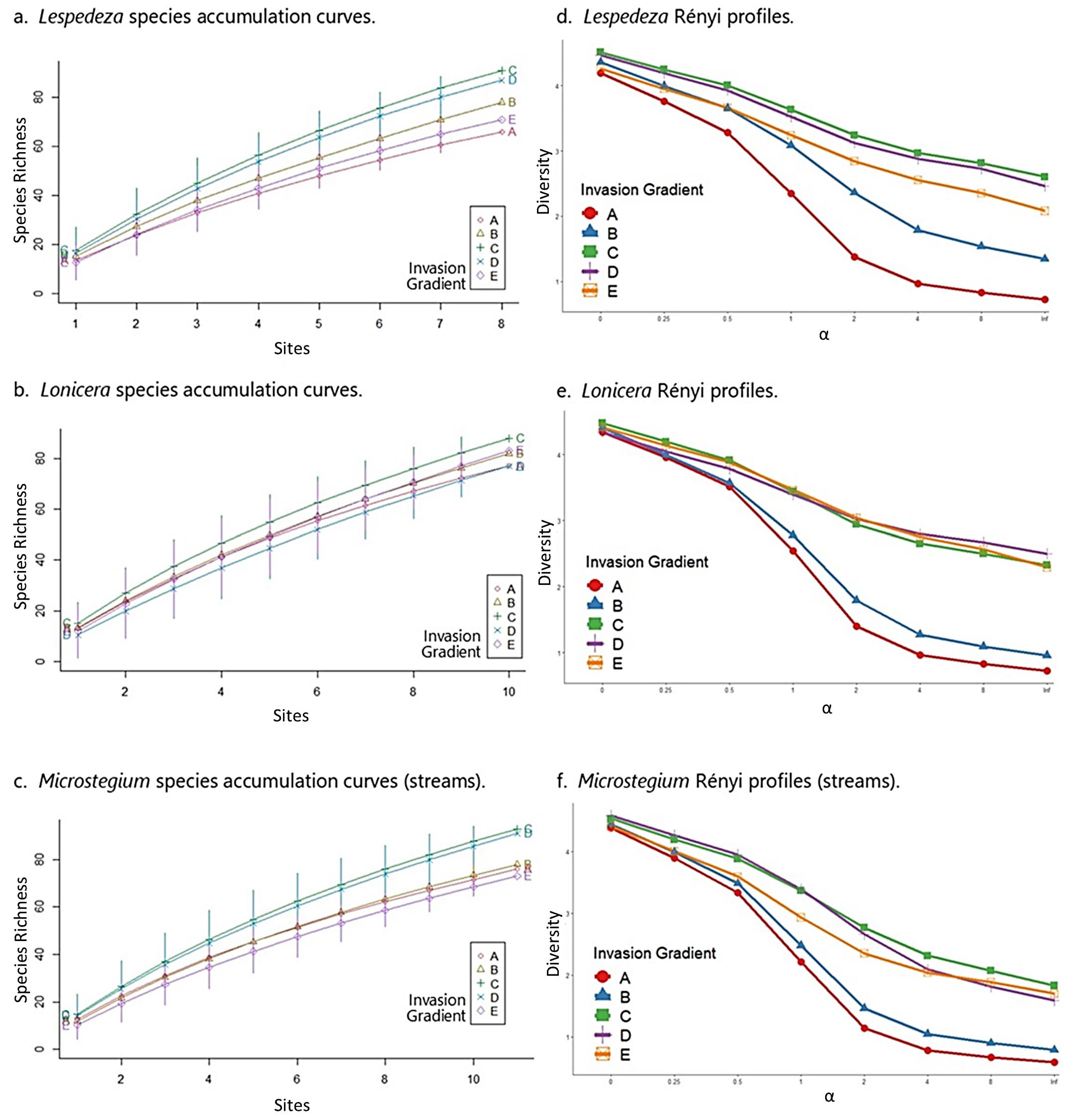
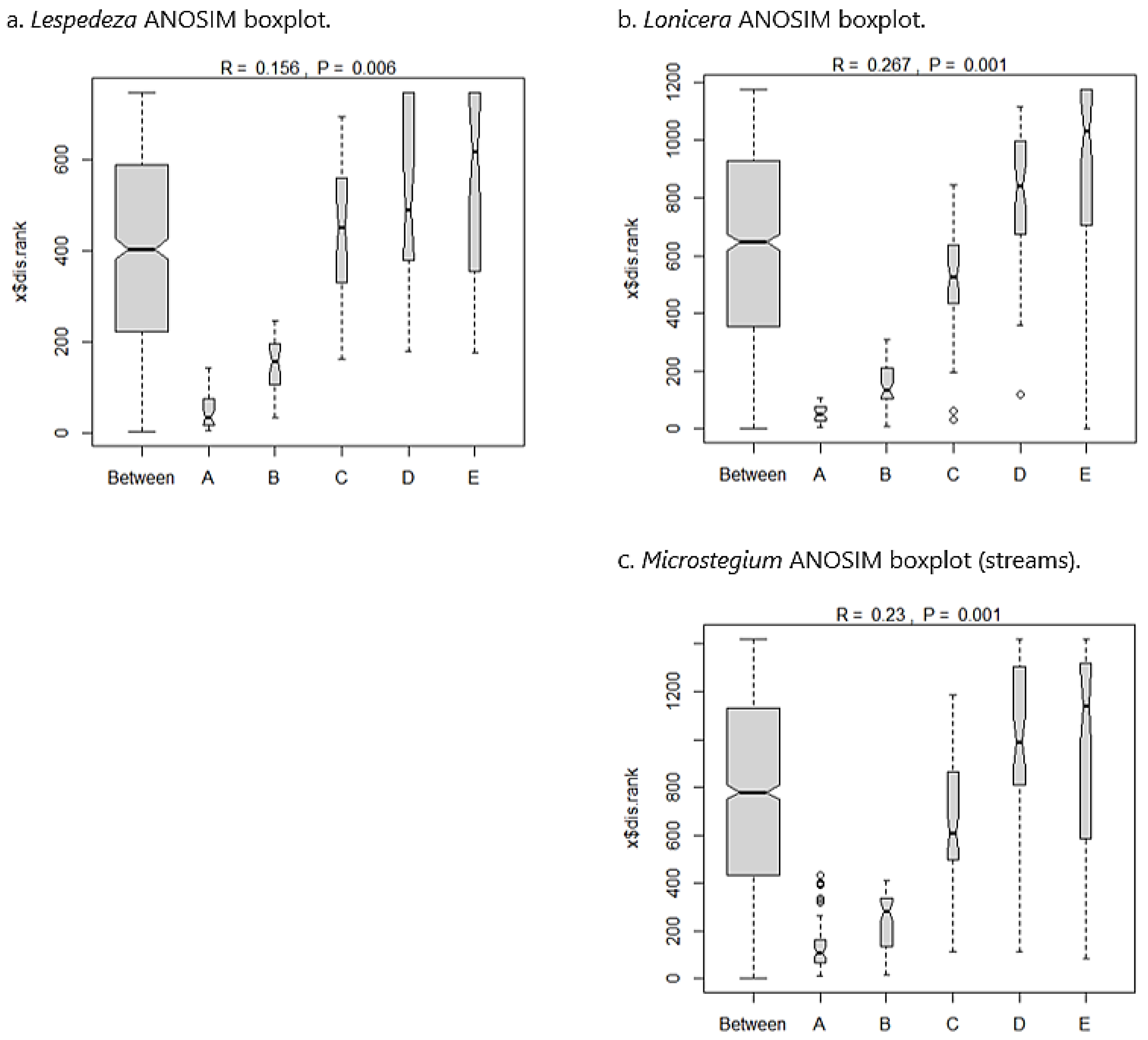
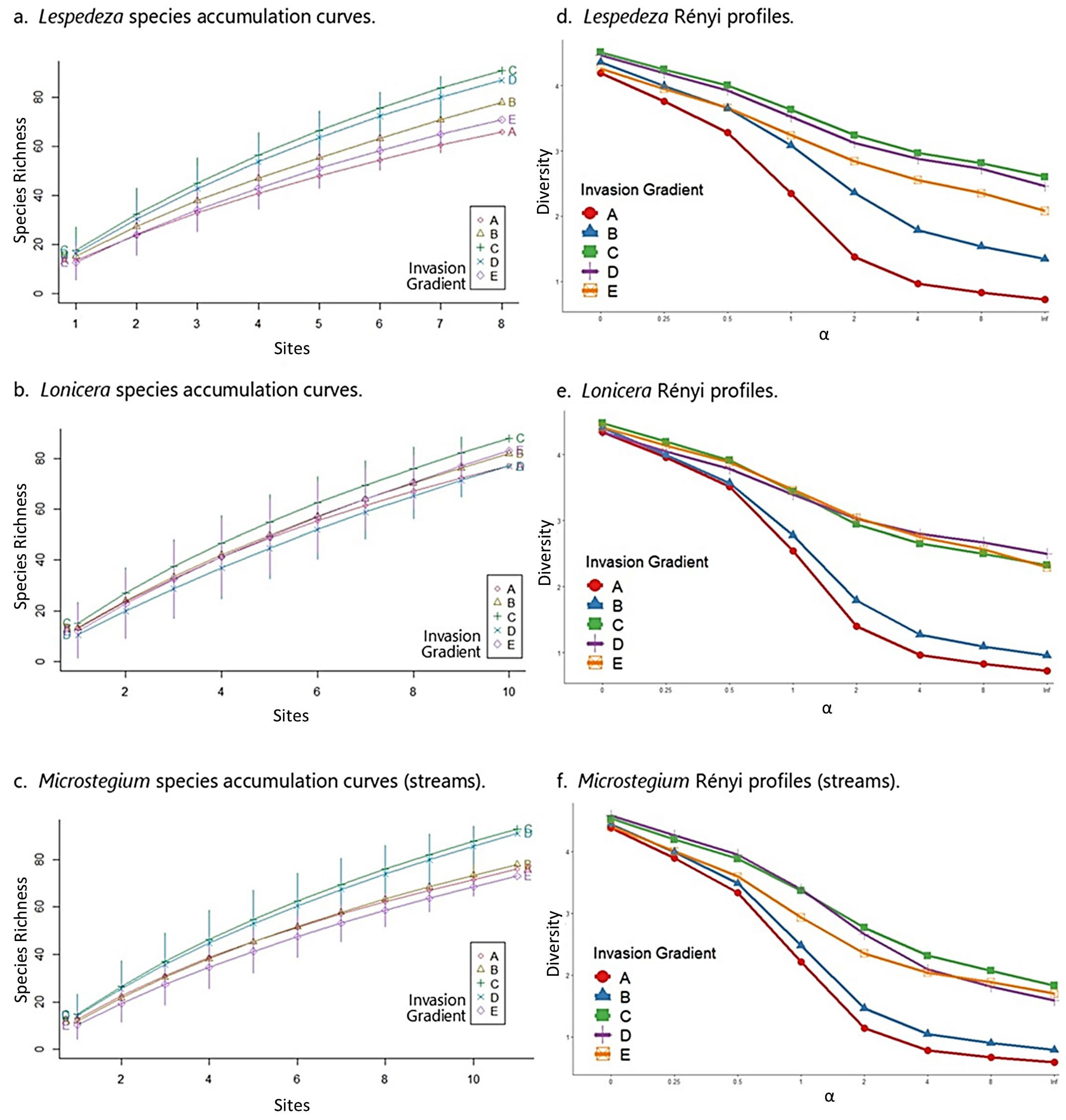
3.5. Species Composition—Streams
3.6. Community Properties—Streams
3.7. Invasive Species Impact Threshold
4. Discussion
4.1. Impact of Invaders on Species Composition
4.2. Impact of Invaders on Community Properties
4.3. Results-Based Invasive Species Performance Standard
4.4. Monitoring Plant Invasion on Mitigation Sites—Recommendations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blossey, B. Before, during and after: The Need for Long-Term Monitoring in Invasive Plant Species Management. Biol. Invasions 1999, 1, 301–311. [Google Scholar] [CrossRef]
- Bryson, C.T.; Carter, R. Biology of Pathways for Invasive Weeds 1. Weed Technol. 2004, 18, 1216–1220. [Google Scholar] [CrossRef]
- Zedler, J.B.; Kercher, S. Causes and Consequences of Invasive Plants in Wetlands: Opportunities, Opportunists, and Outcomes. Crit. Rev. Plant Sci. 2004, 23, 431–452. [Google Scholar] [CrossRef]
- Shea, K.; Chesson, P. Community Ecology Theory as a Framework for Biological Invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- DeBerry, D.A.; McGoff, N.; Zinn, N.D. An Account of Schoenoplectus mucronatus (L.) Palla in Virginia with Comments on Species Introductions in Wetland Mitigation Sites. Castanea 2010, 75, 503–505. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. Wetlands; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- McElfish, J.M.; Brooks, R.P. Policy and Regulatory Programs Affecting Wetlands and Waters of the Mid-Atlantic Region. In Mid-Atlantic Freshwater Wetlands: Advances in Wetlands Science, Management, Policy, and Practice; Brooks, R.P., Wardrop, D.H., Eds.; Springer: New York, NY, USA, 2013; pp. 441–462. ISBN 978-1-4614-5595-0. [Google Scholar]
- Brooks, R.P.; Gebo, N.A. Wetlands Restoration and Mitigation. In Mid-Atlantic Freshwater Wetlands: Advances in Wetlands Science, Management, Policy, and Practice; Brooks, R.P., Wardrop, D.H., Eds.; Springer: New York, NY, USA, 2013; pp. 421–440. ISBN 978-1-4614-5595-0. [Google Scholar]
- Bergdolt, F.S.; Prehmus, C.A.; Barham, J.B. An Evaluation of Wetland Mitigation Site Compliance at the Washington State Department of Transportation; Washington State Department of Transportation: Olympia, WA, USA, 2005.
- Pimentel, D. Environmental and Economic Costs Associated with Alien Invasive Species in the United States. In Biological Invasions: Economic and Environmental Costs of Alien Plant, Animal, and Microbe Species; Pimentel, D., Ed.; CRC Press: Boca Raton, FL, USA, 2011; pp. 411–430. [Google Scholar]
- Brooks, R.P.; Wardrop, D.H.; Cole, C.A.; Campbell, D.A. Are We Purveyors of Wetland Homogeneity? Ecol. Eng. 2005, 24, 331–340. [Google Scholar] [CrossRef]
- Matthews, J.W.; Endress, A.G. Performance Criteria, Compliance Success, and Vegetation Development in Compensatory Mitigation Wetlands. Environ. Manag. 2008, 41, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.W.; Peralta, A.L.; Soni, A.; Baldwin, P.; Kent, A.D.; Endress, A.G. Local and Landscape Correlates of Non-native Species Invasion in Restored Wetlands. Ecography 2009, 32, 1031–1039. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Zhang, L.; Stefanik, K.C.; Nahlik, A.M.; Anderson, C.J.; Bernal, B.; Hernandez, M.; Song, K. Creating Wetlands: Primary Succession, Water Quality Changes, and Self-Design over 15 Years. BioScience 2012, 62, 237–250. [Google Scholar] [CrossRef]
- U.S. Army Corps of Engineers. Virginia Department of Environmental Quality Mitigation Banking Instrument 2018 Template 2018; US Environmental Protection Agency: Washington, DC, USA, 2018.
- D’Antonio, C.; Meyerson, L.A. Exotic Plant Species as Problems and Solutions in Ecological Restoration: A Synthesis. Restor. Ecol. 2002, 10, 703–713. [Google Scholar] [CrossRef]
- Kettenring, K.M.; Adams, C.R. Lessons Learned from Invasive Plant Control Experiments: A Systematic Review and Meta-Analysis: Invasive Plant Control Experiments. J. Appl. Ecol. 2011, 48, 970–979. [Google Scholar] [CrossRef]
- Lawrence, B.A.; Lishawa, S.C.; Rodriguez, Y.; Tuchman, N.C. Herbicide Management of Invasive Cattail (Typha × Glauca) Increases Porewater Nutrient Concentrations. Wetl. Ecol. Manag. 2016, 24, 457–467. [Google Scholar] [CrossRef]
- Reiss, K.C.; Hernandez, E.; Brown, M.T. Evaluation of Permit Success in Wetland Mitigation Banking: A Florida Case Study. Wetlands 2009, 29, 907–918. [Google Scholar] [CrossRef]
- Kozich, A.T.; Halvorsen, K.E. Compliance with Wetland Mitigation Standards in the Upper Peninsula of Michigan, USA. Environ. Manag. 2012, 50, 97–105. [Google Scholar] [CrossRef]
- Washington State Department of Ecology. Washington State Wetland Mitigation Evaluation Study Phase 2: Evaluating Success; Washington State Department of Ecology: Olympia, WA, USA, 2002.
- Mack, J.J.; Fennessy, M.S.; Micacchion, M.; Porej, D. Standardized Monitoring Protocols, Data Analysis and Reporting Requirements for Mitigation Wetlands in Ohio, v. 1.0; Ohio Environmental Protection Agency, Division of Surface Water, Wetland Ecology Group: Columbus, OH, USA, 2004.
- Spieles, D.J.; Coneybeer, M.; Horn, J. Community Structure and Quality After 10 Years in Two Central Ohio Mitigation Bank Wetlands. Environ. Manag. 2006, 38, 837–852. [Google Scholar] [CrossRef]
- Walbeck, D.; Clearwater, D.; Neff, K. Maryland Nontidal Wetland Mitigation Guidance; Maryland Department of Environment Nontidal Wetlands and Waterways Division: Baltimore, MD, USA, 2011.
- U.S. Army Corps of Engineers. Wilmington District Stream and Wetland Compensatory Mitigation Update 2016; North Carolina Department of Transportation: Raleigh, NC, USA, 2016.
- DeMeester, J.E.; de Richter, D.B. Restoring Restoration: Removal of the Invasive Plant Microstegium Vimineum from a North Carolina Wetland. Biol. Invasions 2010, 12, 781–793. [Google Scholar] [CrossRef]
- Dee, S.M.; Ahn, C. Soil Properties Predict Plant Community Development of Mitigation Wetlands Created in the Virginia Piedmont, USA. Environ. Manag. 2012, 49, 1022–1036. [Google Scholar] [CrossRef]
- Perry, J.E.; Morgan, E.; Bevington, A.E.; DeBerry, D. Understanding Cattail (Typha spp.) Invasion and Persistence in Forested Wetlands Created by the Virginia Department of Transportation; Virginia Transportation Research Council: Charlottesville, VA, USA, 2009.
- DeBerry, D.A.; Perry, J.E. Vegetation Dynamics across a Chronosequence of Created Wetland Sites in Virginia, USA. Wetl. Ecol. Manag. 2012, 20, 521–537. [Google Scholar] [CrossRef]
- Swearingen, J.; Slattery, B.; Reshetiloff, K.; Zwicker, S. Plant Invaders of Mid-Atlantic Natural Areas; National Park Service: Washington, DC, USA, 2010.
- Heffernan, K.; Engle, E.; Richardson, C. Virginia Invasive Plant Species List; Natural Heritage Technical Document 14–11; Virginia Department of Conservation and Recreation, Division of Natural Heritage: Richmond, VA, USA, 2014.
- White, L.; Catterall, C.; Taffs, K. The Habitat and Management of Hairy Jointgrass (Arthraxon hispidus, Poaceae) on the North Coast of New South Wales, Australia. Pac. Conserv. Biol. 2020, 26, 45. [Google Scholar] [CrossRef]
- Barden, L.S. Invasion of Microstegium vimineum (Poaceae), An Exotic, Annual, Shade-Tolerant, C 4 Grass, into a North Carolina Floodplain. Am. Midl. Nat. 1987, 118, 40. [Google Scholar] [CrossRef]
- Oswalt, C.M.; Oswalt, S.N.; Clatterbuck, W.K. Effects of Microstegium vimineum (Trin.) A. Camus on Native Woody Species Density and Diversity in a Productive Mixed-Hardwood Forest in Tennessee. For. Ecol. Manag. 2007, 242, 727–732. [Google Scholar] [CrossRef]
- Adams, S.N.; Engelhardt, K.A.M. Diversity Declines in Microstegium vimineum (Japanese Stiltgrass) Patches. Biol. Conserv. 2009, 142, 1003–1010. [Google Scholar] [CrossRef]
- Miller, N.P.; Matlack, G.R. Biodiversity Research: Population Expansion in an Invasive Grass, Microstegium Vimineum: A Test of the Channelled Diffusion Model. Divers. Distrib. 2010, 16, 816–826. [Google Scholar] [CrossRef]
- Warren, R.J.; Wright, J.P.; Bradford, M.A. The Putative Niche Requirements and Landscape Dynamics of Microstegium Vimineum: An Invasive Asian Grass. Biol. Invasions 2011, 13, 471–483. [Google Scholar] [CrossRef]
- Ziska, L.H.; Tomecek, M.B.; Valerio, M.; Thompson, J.P. Evidence for Recent Evolution in an Invasive Species, M Icrostegium Vimineum, Japanese Stiltgrass. Weed Res. 2015, 55, 260–267. [Google Scholar] [CrossRef]
- Green, E.K.; Galatowitsch, S.M. Differences in Wetland Plant Community Establishment with Additions of Nitrate-N and Invasive Species (Phalaris arundinacea and Typha × Glauca). Can. J. Bot. 2001, 79, 170–178. [Google Scholar] [CrossRef]
- Woo, I.; Zedler, J.B. Can Nutrients Alone Shift a Sedge Meadow towards Dominance by the Invasive Typha × Glauca. Wetlands 2002, 22, 509–521. [Google Scholar] [CrossRef]
- Angeloni, N.L.; Jankowski, K.J.; Tuchman, N.C.; Kelly, J.J. Effects of an Invasive Cattail Species (Typha × Glauca) on Sediment Nitrogen and Microbial Community Composition in a Freshwater Wetland. FEMS Microbiol. Lett. 2006, 263, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, L.; Wildova, R.; Goldberg, D.; Vogel, C. Growth of Three Cattail (Typha) Taxa in Response to Elevated CO2. Plant Ecol. 2010, 207, 121–129. [Google Scholar] [CrossRef]
- Wiltermuth, M.T.; Anteau, M.J. Is Consolidation Drainage an Indirect Mechanism for Increased Abundance of Cattail in Northern Prairie Wetlands? Wetl. Ecol. Manag. 2016, 24, 533–544. [Google Scholar] [CrossRef]
- Bansal, S.; Lishawa, S.C.; Newman, S.; Tangen, B.A.; Wilcox, D.; Albert, D.; Anteau, M.J.; Chimney, M.J.; Cressey, R.L.; DeKeyser, E.; et al. Typha (Cattail) Invasion in North American Wetlands: Biology, Regional Problems, Impacts, Ecosystem Services, and Management. Wetlands 2019, 39, 645–684. [Google Scholar] [CrossRef]
- Kalburtji, K.L.; Mosjidis, J.A.; Mamolos, A.P. Litter Dynamics of Low and High Tannin Sericea Lespedeza Plants under Field Conditions. Plant Soil 1999, 208, 271–281. [Google Scholar] [CrossRef]
- Eddy, T.A.; Davidson, J.; Obermeyer, B. Invasion dynamics and biological control prospects for sericea lespedeza in kansas. Great Plains Res. 2003, 13, 217–230. [Google Scholar]
- Brandon, A.L.; Gibson, D.J.; Middleton, B.A. Mechanisms for Dominance in An Early Successional Old Field by the Invasive Non-Native Lespedeza cuneata (Dum. Cours.) G. Don. Biol. Invasions 2004, 6, 483–493. [Google Scholar] [CrossRef]
- Cummings, D.C.; Bidwell, T.G.; Medlin, C.R.; Fuhlendorf, S.D.; Elmore, R.D.; Weir, J.R. Ecology and Management of Sericea Lespedeza; Oklahoma Cooperative Extension Service: Durant, OK, USA, 2007. [Google Scholar]
- Steele, K.L.; Kabrick, J.M.; Dey, D.C.; Jensen, R.G. Restoring Riparian Forests in the Missouri Ozarks. N. J. Appl. For. 2013, 30, 109–117. [Google Scholar] [CrossRef]
- Coykendall, K.E.; Houseman, G.R. Lespedeza Cuneata Invasion Alters Soils Facilitating Its Own Growth. Biol. Invasions 2014, 16, 1735–1742. [Google Scholar] [CrossRef]
- Reichenborn, M.M.; Houseman, G.R.; Foster, B.L. Plant Community Recovery Following Sericea Lespedeza (Lespedeza cuneata) Removal: Testing for a Soil Legacy Effect. Restor. Ecol. 2020, 28, 1192–1200. [Google Scholar] [CrossRef]
- Robertson, D.J.; Robertson, M.C.; Tague, T. Colonization Dynamics of Four Exotic Plants in a Northern Piedmont Natural Area. Bull. Torrey Bot. Club 1994, 121, 107. [Google Scholar] [CrossRef]
- Hidayati, S.N.; Baskin, J.M.; Baskin, C.C. Dormancy-Breaking and Germination Requirements of Seeds of Four Lonicera Species (Caprifoliaceae) with Underdeveloped Spatulate Embryos. Seed Sci. Res. 2000, 10, 459–469. [Google Scholar] [CrossRef]
- Shelton, M.G.; Cain, M.D. Potential Carry-over of Seeds from 11 Common Shrub and Vine Competitors of Loblolly and Shortleaf Pines. Can. J. For. Res. 2002, 32, 412–419. [Google Scholar] [CrossRef]
- Miller, J.H. Nonnative Invasive Plants of Southern Forests: A Field Guide for Identification and Control; US Department of Agriculture, Forest Service, Southern Research Station: Ashville, NC, USA, 2006; Volume 62.
- Schierenbeck, K.A. Japanese Honeysuckle (Lonicera japonica) as an Invasive Species; History, Ecology, and Context. Crit. Rev. Plant Sci. 2004, 23, 391–400. [Google Scholar] [CrossRef]
- Sweeney, B.W.; Czapka, S.J. Riparian Forest Restoration: Why Each Site Needs an Ecological Prescription. For. Ecol. Manag. 2004, 192, 361–373. [Google Scholar] [CrossRef]
- Surrette, S.B.; Brewer, J.S. Inferring Relationships between Native Plant Diversity and Lonicera Japonica in Upland Forests in North Mississippi, USA. Appl. Veg. Sci. 2008, 11, 205–214. [Google Scholar] [CrossRef]
- Ward, E.B.; Pregitzer, C.C.; Kuebbing, S.E.; Bradford, M.A. Invasive Lianas Are Drivers of and Passengers to Altered Soil Nutrient Availability in Urban Forests. Biol. Invasions 2020, 22, 935–955. [Google Scholar] [CrossRef]
- Flory, S.L. Management of Microstegium vimineum Invasions and Recovery of Resident Plant Communities. Restor. Ecol. 2010, 18, 103–112. [Google Scholar] [CrossRef]
- Noon, K.F. A Model of Created Wetland Primary Succession. Landsc. Urban Plan. 1996, 34, 97–123. [Google Scholar] [CrossRef]
- Hunter, D.M.; DeBerry, D.A. Environmental Drivers of Plant Invasion in Wetland Mitigation. Wetlands 2023, 43, 81. [Google Scholar] [CrossRef]
- Mueller-Dombois, D.; Ellenberg, D. Aims and Methods of Vegetation Ecology; John Wiley and Sons, Inc.: New York, NY, USA, 1974. [Google Scholar]
- Weakley, A.; Ludwig, J.; Townsend, J.; Fleming, G. Flora of Virginia—With Significant Additions and Updates; Mobile App.; Foundation of the Flora of Virginia Project Inc. High Country Apps: Bozeman, MT, USA; Richmond, VA, USA, 2020. [Google Scholar]
- Virginia Botanical Associates Digital Atlas of the Virginia Flora. Available online: https://vaplantatlas.org/index.php?do=start (accessed on 3 March 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Kindt, R.; Coe, R. Tree Diversity Analysis: A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies; World Agroforestry Centre: Nairobi, Kenya, 2005. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011; ISBN 978-1-4419-7975-9. [Google Scholar]
- Harrell, F.E.; Dupont, C. Hmisc—Harrell Miscellaneous, R Package Version 4.4-2; Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 1 March 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2020. Available online: https://github.com/vegandevs/vegan (accessed on 1 March 2021).
- DeBerry, D.A.; Perry, J.E. Using the Floristic Quality Concept to Assess Created and Natural Wetlands: Ecological and Management Implications. Ecol. Indic. 2015, 53, 247–257. [Google Scholar] [CrossRef]
- DeBerry, D.A.; Thompson, J.A.; Bradford, Z.R.; Davis, D.L.; Ludwig, J.C.; Simmons, R.H.; Wieboldt, T.F. Coefficients of Conservatism (C-Values) for the Flora of Virginia. Banisteria 2021, 55, 112–149. [Google Scholar]
- Van Den Bosch, K.; Matthews, J.W. An Assessment of Long-Term Compliance with Performance Standards in Compensatory Mitigation Wetlands. Environ. Manag. 2017, 59, 546–556. [Google Scholar] [CrossRef]
- Tiner, R.W. Wetland Indicators: A Guide to Wetland Identification, Delineation, Classification, and Mapping, 2nd ed.; Taylor & Francis: Boca Raton, FL, USA, 2017; ISBN 978-1-4398-5369-6. [Google Scholar]
- Ehrenfeld, J.G. Ecosystem Consequences of Biological Invasions. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 59–80. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Hoopes, M.F.; Marchetti, M.P. Invasion Ecology; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Hooper, D.U.; Vitousek, P.M. The Effects of Plant Composition and Diversity on Ecosystem Processes. Science 1997, 277, 1302–1305. [Google Scholar] [CrossRef]
- DeBerry, D.A.; Perry, J.E. Primary Succession in a Created Freshwater Wetland. Castanea 2004, 69, 185–193. [Google Scholar] [CrossRef]
- Atkinson, R.B.; Perry, J.E.; Cairns, J. Vegetation Communities of 20-Year-Old Created Depressional Wetlands. Wetl. Ecol. Manag. 2005, 13, 469–478. [Google Scholar] [CrossRef]
- Huston, M.A. Biological Diversity: The Coexistence of Species on Changing Landscapes; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Gunderson, L.H. Ecological Resilience—In Theory and Application. Annu. Rev. Ecol. Syst. 2000, 31, 425–439. [Google Scholar] [CrossRef]
- Galatowitsch, S.M.; Anderson, N.O.; Ascher, P.D. Invasiveness in Wetland Plants in Temperate North America. Wetlands 1999, 19, 733–755. [Google Scholar] [CrossRef]
- DeBerry, D.; Beisch, D.; Crayosky, T. Integrated Wetland Design: Sustainability within the Landscape. Geo-Strat. Geo Inst. ASCE 2004, 5, 23–25. [Google Scholar]
- Bradley, B.A.; Blumenthal, D.M.; Wilcove, D.S.; Ziska, L.H. Predicting Plant Invasions in an Era of Global Change. Trends Ecol. Evol. 2010, 25, 310–318. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Huenneke, L.F. Disturbance, Diversity, and Invasion: Implications for Conservation. Conserv. Biol. 1992, 6, 324–337. [Google Scholar] [CrossRef]
- Alpert, P.; Bone, E.; Holzapfel, C. Invasiveness, Invasibility and the Role of Environmental Stress in the Spread of Non-Native Plants. Perspect. Plant Ecol. Evol. Syst. 2000, 3, 52–66. [Google Scholar] [CrossRef]
- Naiman, R.J.; Décamps, H. The Ecology of Interfaces: Riparian Zones. Annu. Rev. Ecol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef]
- Hupp, C.R.; Osterkamp, W.R. Riparian Vegetation and Fluvial Geomorphic Processes. Geomorphology 1996, 14, 277–295. [Google Scholar] [CrossRef]
- Bendix, J.; Hupp, C.R. Hydrological and Geomorphological Impacts on Riparian Plant Communities. Hydrol. Process. 2000, 14, 2977–2990. [Google Scholar] [CrossRef]
- Hughes, F.M.R.; Colston, A.; Mountford, J.O. Restoring Riparian Ecosystems: The Challenge of Accommodating Variability and Designing Restoration Trajectories. Ecol. Soc. 2005, 10, 12. [Google Scholar] [CrossRef]
- Biswas, S.R.; Mallik, A.U. Disturbance Effects on Species Diversity and Functional Diversity in Riparian and Upland Plant Communities. Ecology 2010, 91, 28–35. [Google Scholar] [CrossRef]
- Catford, J.A.; Daehler, C.C.; Murphy, H.T.; Sheppard, A.W.; Hardesty, B.D.; Westcott, D.A.; Rejmánek, M.; Bellingham, P.J.; Pergl, J.; Horvitz, C.C.; et al. The Intermediate Disturbance Hypothesis and Plant Invasions: Implications for Species Richness and Management. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 231–241. [Google Scholar] [CrossRef]
- Apfelbaum, S.I.; Sams, C.E. Ecology and Control of Reed Canary Grass (Phalaris arundinacea L.). Nat. Areas J. 1987, 7, 69–74. [Google Scholar]
- DeBerry, D.A. Vegetation Sampling Concepts for Compensatory Mitigation Sites. Wetl. Sci. Pract. 2020, 37, 174–182. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Scientific Name | Family | Dur. | Habit | Origin | Comments | Refs. |
|---|---|---|---|---|---|---|---|
| Wetland mitiga-tion | Arthraxon hispidus (Thunb.) Makino | Poaceae | A | Graminoid | East Asia | Little attention in the literature. Moderately invasive throughout Mid-Atlantic region; problematic on mitigation sites. | [30,31,32] |
| Microstegium vimineum (Trin.) A. Camus | Poaceae | A | Graminoid | Asia | Tolerant of shading and temporary flooding. Prolific seeder. Forms persistent seed banks. Inhabits variety of wetland and upland habitats. Alters community structure and reduces native plant diversity. | [31,33,34,35,36,37,38] | |
| Typha spp. L. | Typhaceae | P | Forb | U.S. | Two cattail species (Typha latifolia L. and Typha angustifolia L.) and their hybrid (Typha x glauca Godron). Native to U.S. (T. angustifolia putatively introduced from Europe) but regulated as invasive on wetland mitigation sites. Tolerant of prolonged inundation. Impacts on wetland communities have been questioned. | [3,28,39,40,41,42,43,44] | |
| Stream mitigation | Lespedeza cuneata (Dum.-Cours.) G. Don | Fabaceae | P | Forb | East Asia | Occupies well-drained soils. A nitrogen fixer, its extensive taproot allows survival in drought conditions and a wide range of soil pH. Modifies habitat to facilitate invasion. Herbivory-resistant with allelopathic properties. Modifies nutrient pools by rapid uptake/slow release via slower decomposition (through concentrated tannins and phenolic compounds). | [30,31,45,46,47,48,49,50,51] |
| Lonicera japonica Thunb. | Caprifoliaceae | P | Vine | East Asia | Dispersed by birds but expansion generally occurs vegetatively. Due to high transpiration rates, does not tolerate prolonged drought and therefore tends to prefer mesic habitats, making riparian zones, streambanks, and floodplains susceptible to invasion. Somewhat shade tolerant, but prefers canopy openings to promote localized dominance. | [30,31,52,53,54,55,56,57,58,59] | |
| Microstegium vimineum (Trin.) A. Camus | Poaceae | A | Graminoid | Asia | See comments above. Factors contributing to invasion potential are perhaps more important in streams, floodplains, and riparian zones due to use of flowing water as a dispersal mechanism. | [26,33,37,60] |
| a. Arthraxon | B | C | D | E | |
| A | 0.57 | 0.59 | 0.67 | 0.44 | |
| B | 0.61 | 0.60 | 0.57 | ||
| C | 0.56 | 0.46 | |||
| D | 0.50 | ||||
| b. Microstegium | B | C | D | E | |
| A | 0.48 | 0.56 | 0.52 | 0.58 | |
| B | 0.48 | 0.64 | 0.50 | ||
| C | 0.52 | 0.51 | |||
| D | 0.56 | ||||
| c. Typha | B | C | D | E | |
| A | 0.65 | 0.49 | 0.64 | 0.57 | |
| B | 0.51 | 0.57 | 0.54 | ||
| C | 0.56 | 0.57 | |||
| D | 0.70 |
| Mean Native Species Richness | |||||
| Invasion Gradient: | A | B | C | D | E |
| Arthraxon | 8.6 | 11.9 | 12.1 | 9.5 | 12.4 |
| Microstegium | 5.7 | 8.3 | 9.8 | 8.6 | 8.4 |
| Typha | 6.1 | 5.4 | 9.0 | 5.9 | 7.2 |
| Floristic Quality Index (FQI) | |||||
| Invasion Gradient: | A | B | C | D | E |
| Arthraxon | 10.4 | 12.7 | 12.8 | 11.3 | 12.2 |
| Microstegium | 9.4 | 11.0 | 12.3 | 11.8 | 11.6 |
| Typha | 8.5 | 7.6 | 10.1 | 9.3 | 9.1 |
| Mean Native Species Richness | |||||
| Invasion Gradient: | A | B | C | D | E |
| Arthraxon | 66.4 | 23.2 | 6.6 | 0.7 | 0.0 |
| Microstegium | 58.9 | 28.8 | 8.1 | 0.9 | 0.0 |
| Typha | 58.2 | 26.6 | 5.1 | 0.0 | 0.0 |
| a. Lespedeza | B | C | D | E | ||
| A | 0.61 | 0.57 | 0.58 | 0.69 | ||
| B | 0.70 | 0.67 | 0.66 | |||
| C | 0.67 | 0.62 | ||||
| D | 0.66 | |||||
| b. Lonicera | B | C | D | E | ||
| A | 0.58 | 0.56 | 0.48 | 0.46 | ||
| B | 0.64 | 0.52 | 0.53 | |||
| C | 0.63 | 0.55 | ||||
| D | 0.65 | |||||
| c. Microstegium | B | C | D | E | ||
| A | 0.56 | 0.51 | 0.49 | 0.53 | ||
| B | 0.54 | 0.55 | 0.50 | |||
| C | 0.62 | 0.55 | ||||
| D | 0.56 | |||||
| Mean Native Species Richness | |||||
| Invasion Gradient: | A | B | C | D | E |
| Lespedeza | 11.4 | 13.3 | 15.1 | 14.4 | 10.6 |
| Lonicera | 10.3 | 10.7 | 12.4 | 8.6 | 10.1 |
| Microstegium | 9.8 | 9.5 | 12.1 | 11.9 | 9.0 |
| Floristic Quality Index (FQI) | |||||
| Invasion Gradient: | A | B | C | D | E |
| Lespedeza | 11.1 | 12.8 | 14.0 | 11.5 | 12.6 |
| Lonicera | 10.7 | 12.4 | 13.5 | 10.2 | 11.3 |
| Microstegium | 11.1 | 11.1 | 12.9 | 12.2 | 10.7 |
| Mean Native Species Richness | |||||
| Invasion Gradient: | A | B | C | D | E |
| Lespedeza | 48.4 | 26.1 | 3.2 | 0.6 | 0.2 |
| Lonicera | 48.6 | 38.5 | 6.5 | 1.1 | 0.4 |
| Microstegium | 56.9 | 44.1 | 14.0 | 2.6 | 0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
DeBerry, D.A.; Hunter, D.M. Impacts of Invasive Plants on Native Vegetation Communities in Wetland and Stream Mitigation. Biology 2024, 13, 275. https://doi.org/10.3390/biology13040275
DeBerry DA, Hunter DM. Impacts of Invasive Plants on Native Vegetation Communities in Wetland and Stream Mitigation. Biology. 2024; 13(4):275. https://doi.org/10.3390/biology13040275
Chicago/Turabian StyleDeBerry, Douglas A., and Dakota M. Hunter. 2024. "Impacts of Invasive Plants on Native Vegetation Communities in Wetland and Stream Mitigation" Biology 13, no. 4: 275. https://doi.org/10.3390/biology13040275
APA StyleDeBerry, D. A., & Hunter, D. M. (2024). Impacts of Invasive Plants on Native Vegetation Communities in Wetland and Stream Mitigation. Biology, 13(4), 275. https://doi.org/10.3390/biology13040275