
Molecular Detection and Genetic Variability of Hepatozoon canis in Golden Jackals (Canis aureus L. 1758) in Serbia

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
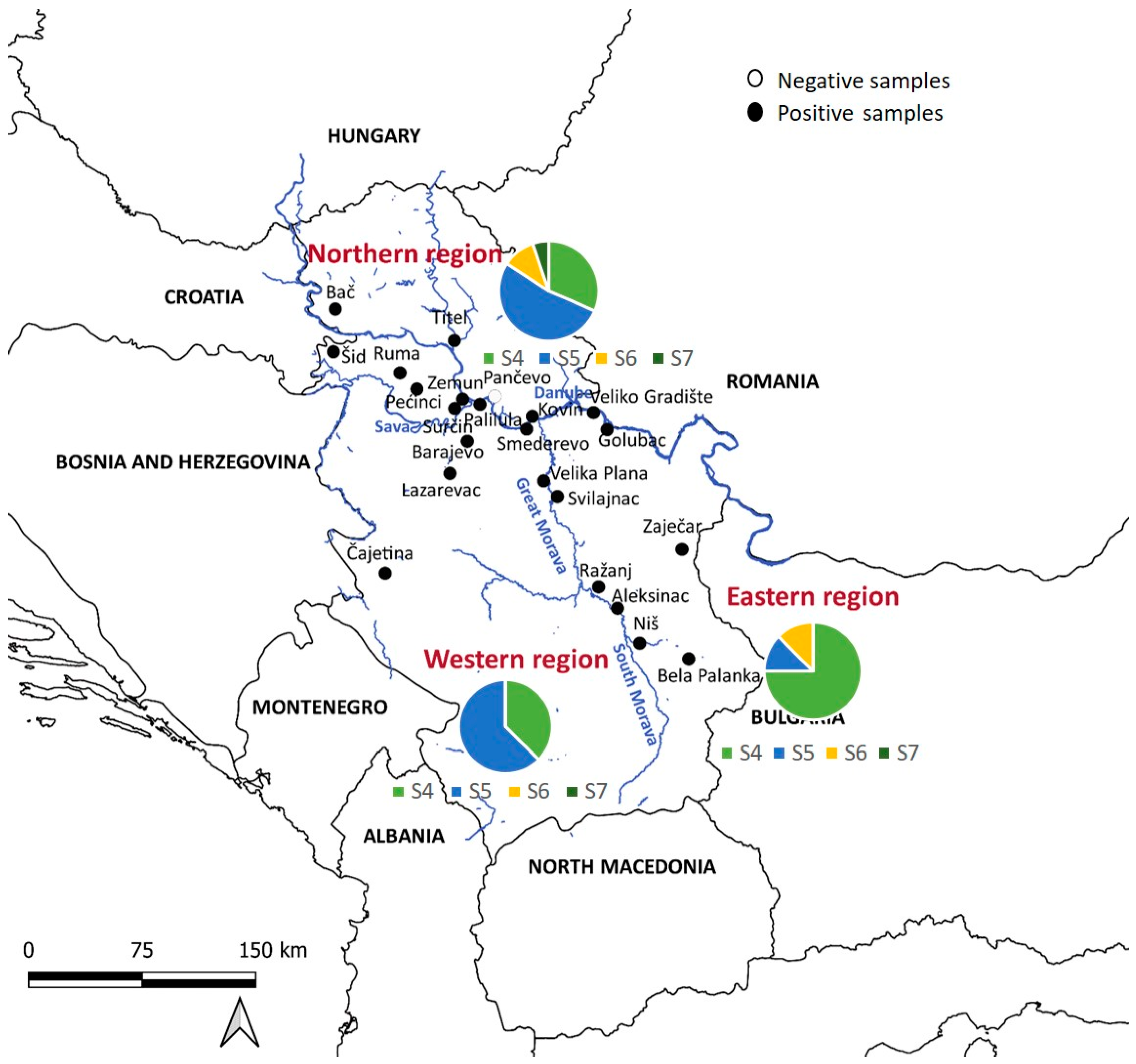
2.1. Study Area and Sampling
2.2. Molecular Detection and Characterization of H. canis
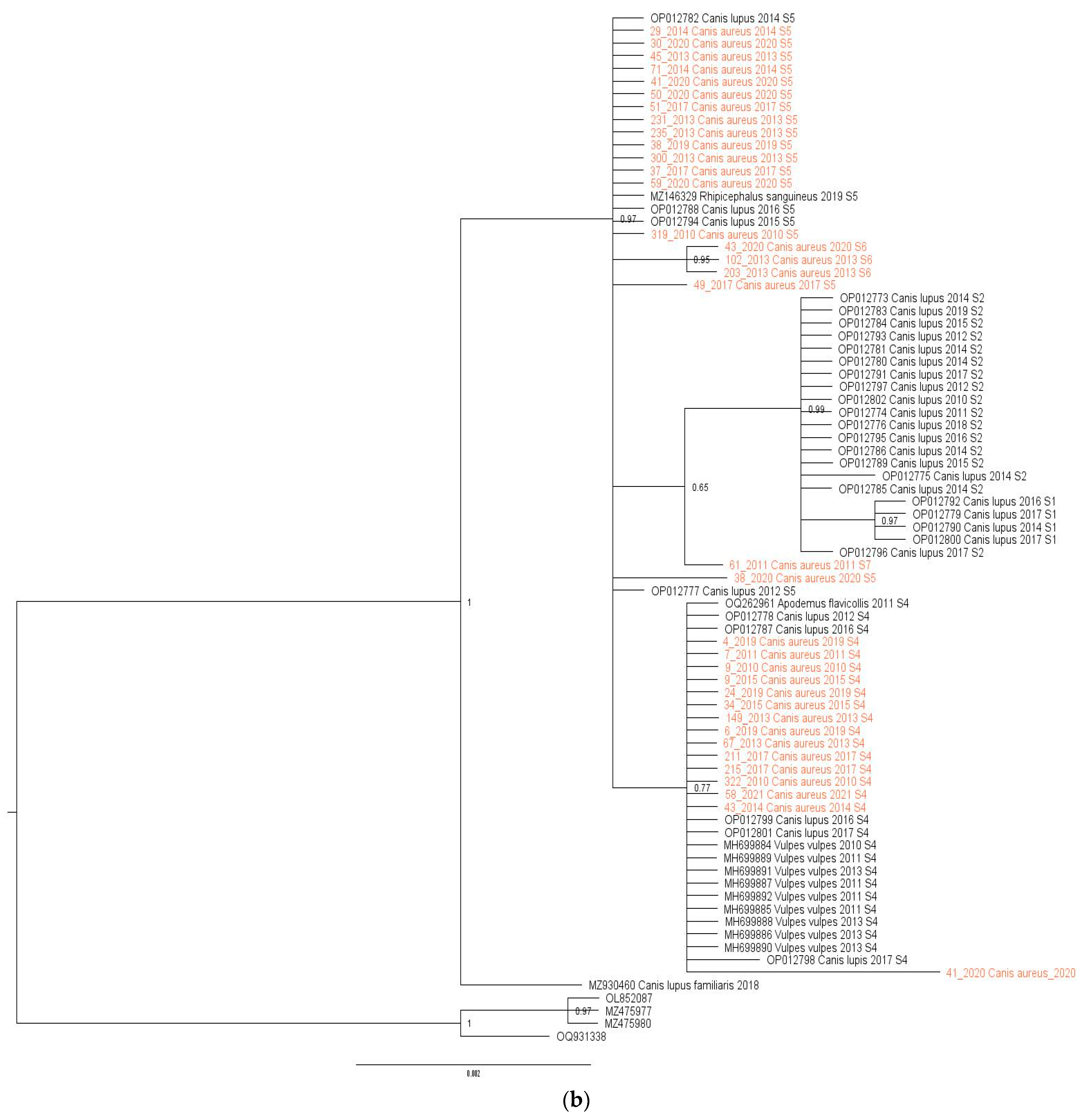
2.3. Phylogenetic Analysis
2.4. Statistics
3. Results
3.1. Demographic Characteristics and Regional Distribution of Sampled Golden Jackals
3.2. Genetic Variability of H. canis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arnold, J.; Humer, A.; Heltai, M.; Murariu, D.; Spassov, N.; Hacklander, K. Current status and distribution of golden jackal Canis aureus in Europe. Mammal. Rev. 2012, 42, 1–11. [Google Scholar] [CrossRef]
- Trouwborst, A.; Krofel, M.; Linnell, J.D. Legal implications of range expansions in a terrestrial carnivore: The case of the golden jackal (Canis aureus) in Europe. Biodivers. Conserv. 2015, 24, 2593–2610. [Google Scholar] [CrossRef]
- Hoffmann, M.; Arnold, J.; Duckworth, J.W.; Jhala, Y.W.; Kamler, J.F.; Krofel, M. Canis aureus. The IUCN Red List of Threatened Species. 2018, E. T118264161A146194820. Available online: https://doi.org/10.2305/IUCN.UK.2018-2.RLTS.T118264161A46194820.en (accessed on 1 January 2024).
- Kryštufek, B.; Murariu, D.; Kurtonour, C. Present distribution of the Golden Jackal Canis aureus in Balkans and adjacent regions. Mammal. Rev. 1997, 27, 109–114. [Google Scholar] [CrossRef]
- Lanszki, J.; Schally, G.; Heltai, M.; Ranc, N. Golden jackal expansion in Europe: First telemetry evidence of a natal dispersal. Mamm. Biol. 2018, 88, 81–84. [Google Scholar] [CrossRef]
- Penezić, A.; Ćirović, D. Seasonal variation in diet of the golden jackal (Canis aureus) in Serbia. Mammal Res. 2015, 60, 309–317. [Google Scholar] [CrossRef]
- Spassov, N.; Acosta-Pankov, I. Dispersal history of the golden jackal (Canis aureus moreoticus Geoffroy, 1835) in Europe and possible causes of its recent population explosion. Biodivers. Data J. 2019, 7, e34825. [Google Scholar] [CrossRef] [PubMed]
- Ionică, A.M.; Matei, I.A.; D’Amico, G.; Daskalaki, A.A.; Juránková, J.; Ionescu, D.T.; Mihalca, A.D.; Modrý, D.; Gherman, C.M. Role of golden jackals (Canis aureus) as natural reservoirs of Dirofilaria spp. in Romania. Parasites Vectors 2016, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mitková, B.; Hrazdilová, K.; D’Amico, G.; Duscher, G.G.; Suchentrunk, F.; Forejtek, P.; Gherman, C.M.; Matei, I.A.; Ionică, A.M.; Daskalaki, A.A.; et al. Eurasian golden jackal as host of canine vector-borne protists. Parasites Vectors 2017, 10, 183. [Google Scholar] [CrossRef]
- Smith, T.G. The genus Hepatozoon (Apicomplexa: Adeleina). J. Parasitol. 1996, 82, 565–585. [Google Scholar] [CrossRef] [PubMed]
- Baneth, G. Perspectives on canine and feline hepatozoonosis. Vet. Parasitol. 2011, 181, 3–11. [Google Scholar] [CrossRef]
- Solano-Gallego, L.; Baneth, G. Babesiosis in dogs and cats—Expanding parasitological and clinical spectra. Vet. Parasitol. 2011, 181, 8–60. [Google Scholar] [CrossRef]
- Alvarado-Rybak, M.; Solano-Gallego, L.; Millán, J. A review of piroplasmid infections in wild carnivores worldwide: Importance for domestic animal health and wildlife conservation. Parasites Vectors 2016, 9, 538. [Google Scholar] [CrossRef] [PubMed]
- Baneth, G.; Samish, M.; Shkap, V. Life cycle of Hepatozoon canis (Apicomplexa: Adeleorina: Hepatozoidae) in the tick Rhipicephalus sanguineus and domestic dog (Canis familiaris). J. Parasitol. 2007, 93, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Uiterwijk, M.; Vojta, L.; Šprem, N.; Beck, A.; Jurković, D.; Kik, M.; Duscher, G.G.; Hodžić, A.; Reljić, S.; Sprong, H.; et al. Diversity of Hepatozoon species in wild mammals and ticks in Europe. Parasites Vectors 2023, 16, 27. [Google Scholar] [CrossRef]
- Murata, T.; Inoue, M.; Tateyama, S.; Taura, Y.; Nakama, S. Vertical transmission of Hepatozoon canis in dogs. J. Vet. Med. Sci. 1993, 55, 867–868. [Google Scholar] [CrossRef] [PubMed]
- Hodžić, A.; Mrowietz, N.; Cézanne, R.; Bruckschwaiger, P.; Punz, S.; Habler, V.E.; Tomsik, V.; Lazar, J.; Duscher, D.D.; Glawisching, W.; et al. Occurrence and diversity of arthropod-transmitted pathogens in red foxes (Vulpes vulpes) in western Austria, and possible vertical (transplacental) transmission of Hepatozoon canis. Parasitology 2018, 145, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, I.; Müller, E.; Nijhof, A.M.; Aupperle-Lellbach, H.; Loesenbeck, G.; Cramer, S.; Naucke, T.J. First evidence of vertical Hepatozoon canis transmission in dogs in Europe. Parasites Vectors 2022, 15, 296. [Google Scholar] [CrossRef] [PubMed]
- Margalit Levi, M.; Nachum-Biala, Y.; King, R.; Baneth, G. A survey of Babesia spp. and Hepatozoon spp. in wild canids in Israel. Parasites Vectors 2018, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Inokuma, H.; Okuda, M.; Ohno, K.; Shimoda, K.; Onishi, T. Analysis of the 18S rRNA gene sequence of a Hepatozoon detected in two Japanese dogs. Vet. Parasitol. 2002, 106, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Kuručki, M.; Tomanović, S.; Sukara, R.; Ćirović, D. High Prevalence and Genetic Variability of Hepatozoon canis in Grey Wolf (Canis lupus L. 1758) Population in Serbia. Animals 2022, 12, 3335. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K.; Mega, X. Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Posada, D. JModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Milutinović, M.; Petrović, Z.; Bobić, B.; Pavlović, I. Ecological notes on ticks (Acari: Ixodidae) collected in west Serbia, Yugoslavia. Parasitol. Hung. 1996, 29–30, 67–74. [Google Scholar]
- Milutinović, M.; Masuzawa, T.; Tomanović, S.; Radulović, Ž.; Fukui, T.; Okamoto, Y. Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum, Francisella tularensis and their co-infections in host-seeking Ixodes ricinus ticks collected in Serbia. Exp. Appl. Acarol. 2008, 45, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Potkonjak, A.; Gutiérrez, R.; Savić, S.; Vračar, V.; Nachum-Biala, Y.; Jurišić, A.; Kleinerman, G.; Rojas, A.; Petrović, A.; Baneth, G.; et al. Molecular detection of emerging tick-borne pathogens in Vojvodina, Serbia. Ticks Tick-Borne Dis. 2016, 7, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Tomanović, S.; Chochlakis, D.; Radulović, Ž.; Milutinović, M.; Ćakić, S.; Mihaljica, D.; Tselentis, Y.; Psaroulaki, A. Analysis of pathogen co-occurrence in host seeking adult hard ticks from Serbia. Exp. Appl. Acarol. 2013, 59, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Tomanović, S.; Radulović, Ž.; Ćakić, S.; Mihaljica, D.; Sukara, R.; Penezić, A.; Burazerović, J.; Ćirović, D. Tick species (acari: Ixodidae) of red foxes (Vulpes vulpes) in Serbia. In Proceedings of the 2nd International Symposium on Hunting, Novi Sad, Serbia, 17–20 October 2013. [Google Scholar]
- Juwaid, S.; Sukara, R.; Penezić, A.; Mihaljica, D.; Veinović, G.; Kavallieratos, N.G.; Ćirović, D.; Tomanović, S. First evidence of tick-borne protozoan pathogens, Babesia sp. and Hepatozoon canis, in red foxes (Vulpes vulpes) in Serbia. Acta Vet. Hung. 2019, 67, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Duscher, G.; Ćirović, D.; Heltai, M.; Szabó, L.; Lanszki, J.; Bošković, I.; Floriančić, T.; Knauer, F.; Suchentrunk, F. Hepatozoonozis in golden jackal (Canis aureus) from southeastern and central Europe: Prevalence data from a first molecular screening. In Book of Abstracts of First International Jackal Symposium; University of Belgrade: Veliko Gradiste, Serbia, 2014; p. 70. [Google Scholar]
- Banović, P.; Díaz-Sánchez, A.A.; Galon, C.; Foucault-Simonin, A.; Simin, V.; Mijatović, D.; Papić, L.; Wu-Chuang, A.; Obregón, D.; Moutailler, S.; et al. A One Health approach to study the circulation of tick-borne pathogens: A preliminary study. One Health 2021, 13, 100270. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, S.; Otašević, S.; Ignjatović, A.; Savić, S.; Fraulo, M.; Arsić-Arsenijević, V.; Momčilović, S.; Cancrini, G. Canine babesioses in noninvestigated areas of Serbia. Vector-Borne Zoonotic Dis. 2015, 15, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Sukara, R.; Andrić, N.; Andrić, J.F.; Mihaljica, D.; Veinović, G.; Ranković, V.; Tomanović, S. Autochthonous infection with Ehrlichia canis and Hepatozoon canis in dogs from Serbia. Vet. Med. Sci. 2023, 9, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Veinović, G.; Sukara, R.; Mihaljica, D.; Penezić, A.; Ćirović, D.; Tomanović, S. The Occurrence and Diversity of Tick-Borne Pathogens in Small Mammals from Serbia. Vector-Borne Zoonotic Dis. 2024, 24, 5. [Google Scholar] [CrossRef] [PubMed]
- Farkas, R.; Solymosi, N.; Takács, N.; Hornyák, Á.; Hornok, S.; Nachum-Biala, Y.; Baneth, G. First molecular evidence of Hepatozoon canis infection in red foxes and golden jackals from Hungary. Parasites Vectors 2014, 7, 303. [Google Scholar] [CrossRef] [PubMed]
- Hodžić, A.; Georges, I.; Postl, M.; Duscher, G.G.; Jeschke, D.; Szentiks, A.C.; Ansorge, H.; Heddergott, M. Molecular survey of tick-borne pathogens reveals a high prevalence and low genetic variability of Hepatozoon canis in free-ranging grey wolves (Canis lupus) in Germany. Ticks Tick Borne Dis. 2020, 11, 101389. [Google Scholar] [CrossRef] [PubMed]
- Battisti, E.; Zanet, S.; Khalili, S.; Trisciuoglio, A.; Hertel, B.; Ferroglio, E. Molecular survey on vector-borne pathogens in alpine wild carnivorans. Front. Vet. Sci. 2020, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Criado-Fornelio, A.; Martín-Pérez, T.; Verdú-Expósito, C.; Reinoso-Ortiz, S.A.; Pérez-Serrano, J. Molecular epidemiology of parasitic protozoa and Ehrlichia canis in wildlife in Madrid (central Spain). Parasitol. Res. 2018, 117, 2291–2298. [Google Scholar] [CrossRef] [PubMed]
- Ortuño, M.; Nachum-Biala, Y.; García-Bocanegra, I.; Resa, M.; Berriatua, E.; Baneth, G. An epidemiological study in wild carnivores from Spanish Mediterranean ecosystems reveals association between Leishmania infantum, Babesia spp. and Hepatozoon spp. infection and new hosts for Hepatozoon martis, Hepatozoon canis and Sarcocystis spp. Transbound. Emerg. Dis. 2022, 69, 2110–2125. [Google Scholar] [CrossRef] [PubMed]
- Helm, S.C.; Samson-Himmelstjerna, V.G.; Liesner, M.J.; Kohn, B.; Müller, E.; Schaper, R.; Pachnicke, S.; Schulze, C.; Krücken, J. Identical 18S rRNA haplotypes of Hepatozoon canis in dogs and foxes in Brandenburg, Germany. Ticks Tick Borne Dis. 2020, 11, 101520. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, L.; Cortes, H.C.; Eyal, O.; Reis, A.; Lopes, A.P.; Vila-Viçosa, M.J.; Rodrigues, P.A.; Baneth, G. Molecular and histopathological detection of Hepatozoon canis in red foxes (Vulpes vulpes) from Portugal. Parasites Vectors 2014, 7, 113. [Google Scholar] [CrossRef] [PubMed]
- Dordio, A.M.; Beck, R.; Nunes, T.; Pereira da Fonseca, I.; Gomes, J. Molecular survey of vector-borne diseases in two groups of domestic dogs from Lisbon, Portugal. Parasites Vectors 2021, 14, 163. [Google Scholar] [CrossRef] [PubMed]
- Criado-Fornelio, A.; Buling, A.; Pingret, J.L.; Etievant, M.; Boucraut-Baralon, C.; Alongi, A.; Agnone, A.; Torina, A. Hemoprotozoa of domestic animals in France: Prevalence and molecular characterization. Vet. Parasitol. 2009, 159, 73–76. [Google Scholar] [CrossRef]
- Giannelli, A.; Latrofa, M.S.; Nachum-Biala, Y.; Hodžić, A.; Greco, G.; Attanasi, A.; Annosica, A.; Otranto, D.; Baneth, G. Three different Hepatozoon species in domestic cats from southern Italy. Ticks Tick Borne Dis. 2017, 8, 721–724. [Google Scholar] [CrossRef]
- Baneth, G.; Mathew, J.S.; Shkap, V.; Macintire, D.K.; Barta, J.R.; Ewing, S.A. Canine hepatozoonosis—Two disease syndromes caused by separate Hepatozoon species. Trends Parasitol. 2003, 19, 27–31. [Google Scholar] [CrossRef]
- Ćirović, D.; Penezić, A. Importance of slaughter waste in winter diet of wolves (Canis lupus) in Serbia. North-West. J. Zool. 2019, 15, 175–178. [Google Scholar]
- Kobryn, H.T.; Swinhoe, E.J.; Bateman, P.W.; Adams, P.J.; Shephard, J.M.; Fleming, P.A. Foxes at your front door? Habitat selection and home range estimation of suburban red foxes (Vulpes vulpes). Urban Ecosyst. 2023, 26, 1–17. [Google Scholar] [CrossRef]
- Lanszki, J.; Heltai, M.; Szabo, L. Feeding habits and trophic niche overlap betw sympatric golden jackal (Canis aureus) and re (Vulpes vulpes) in the Pannonian ecoregion (Hungary). Can. J. Zool. 2006, 84, 1647–1656. [Google Scholar] [CrossRef]
- Lainson, R.; Paperna, I.; Naiff, R.D. Development of Hepatozoon caimani (Carini, 1909) Pess a, De Biasi & De Souza, 1972 in the Caiman Caiman c. crocodilus, the frog Rana catesbeiana and the mosquito Culex fatigans. Mem. Inst. Oswaldo Cruz. 2003, 98, 103–113. [Google Scholar] [CrossRef]
- Viana, L.A.; Soares, P.; Silva, J.E.; Paiva, F.; Coutinho, M.E. Anurans as paratenic hosts in the transmission of Hepatozoon caimani to caimans Caiman yacare and Caiman latirostris. Parasitol. Res. 2012, 110, 883–886. [Google Scholar] [CrossRef]
- Sloboda, M.; Kamler, M.; Bulantová, J.; Votýpka, J.; Modrý, D. Rodents as intermediate hosts of Hepatozoon ayorgbor (Apicomplexa: Adeleina: Hepatozoidae) from the African ball python, Python regius? Folia Parasitol. 2008, 55, 13–16. [Google Scholar] [CrossRef]
- Smith, T.G.; Desser, S.S.; Martin, D.S. The development of Hepatozoon sipedon sp. nov. (Apicomplexa: Adeleina: Hepatozoidae) in its natural host, the Northern water snake (Nerodia sipedon sipedon), in the culicine vectors Culex pipiens and C. territans, and in an intermediate host, the Northern leopard frog (Rana pipiens). Parasitol. Res. 1994, 80, 559–568. [Google Scholar] [CrossRef]
- Johnson, E.M.; Allen, K.E.; Panciera, R.J.; Ewing, S.A.; Little, S.E. Experimental transmission of Hepatozoon americanum to New Zealand White rabbits (Oryctolagus cuniculus) and infectivity of cystozoites for a dog. Vet. Par. 2009, 164, 162–166. [Google Scholar] [CrossRef]
- Johnson, E.M.; Panciera, R.J.; Allen, K.E.; Sheets, M.E.; Beal, J.D.; Ewing, S.A.; Little, S.E. Alternate pathway of infection with Hepatozoon americanum and the epidemiologic importance of predation. J Vet. Intern. Med./ACVIM 2009, 23, 1315–1318. [Google Scholar] [CrossRef]
- Ćirović, D.; Penezić, A.; Milenković, M.; Paunović, M. Present distribution and factors of range spread of golden jackal (Canis aureus L. 1758) in Serbia. In Proceedings of the International Conference on Large Carnivores, Žagubica, Serbia; Hunting Association of Serbia, Ed.; Hunting Association of Serbia: Beograd, Serbia, 2008; pp. 93–102. [Google Scholar]
- Milenković, M.; Paunović, M. Phenomenon of golden jackal (Canis aureus Linnaeus, 1758), expantion in Serbia. In Carpathian Workshop on Large Carnivores Conservation. Convention on the Conservation of European Wildlife and Natural Habitats; The Council of Europe: Salzburg, Austria; Brasov, Romania, 2003. [Google Scholar]
- Ćirović, D.; Penezić, A.; Plećaš, M. Šakal (Canis aureus) u predelima sa dominantnim antropogenim uticajem. In Slovenski lovski dan, Šakal v Sloveniji in Na Balkanu: Stanje in Upravljavski Izzivi; Pokorny, B., Kmetec, U., Eds.; Lovska Zveza Slovenije: Koper, Slovenia, 2018; pp. 13–14. [Google Scholar]
- Hodžić, A.; Alić, A.; Prašović, S.; Otranto, D.; Baneth, G.; Duscher, G.G. Hepatozoon silvestris sp. nov.: Morphological and molecular characterization of a new species of Hepatozoon (Adeleorina: Hepatozoidae) from the European wild cat (Felis silvestris silvestris). Parasitology 2017, 144, 650–661. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Region | Locality (No. of Collected Samples) | H. canis-Positive | |
|---|---|---|---|
| Male | Female | ||
| NORTHERN REGION | Kovin (3) | 2/3 (66.6) | 0/0 (0) |
| Titel (1) | 1/1 (100) | 0/0 (0) | |
| Surčin (29) | 10/13 (76.9) | 11/16 (68.8) | |
| Bač (5) | 1/3 (33.3) | 1/2 (50) | |
| Palilula (2) | 0/1 (0) | 1/1 (100) | |
| Pećinci (13) | 7/9 (77.7) | 3/4 (75) | |
| Pančevo (1) | 0/1 (0) | 0/0 (0) | |
| Šid (2) | 1/2 (50) | 0/0 (0) | |
| Ruma (1) | 1/1 (100) | 0/0 (0) | |
| Zemun (1) | 1/1 (100) | 0/0 (0) | |
| Total * (%) | 40/58 (68.9) | 24/35 (68.57) | 16/23 (69.57) |
| WESTERN REGION | Barajevo (1) | 0/0 (0) | 1/1 (100) |
| Lazarevac (2) | 1/1 (100) | 1/1 (100) | |
| Smederevo (10) | 3/3 (100) | 7/7 (100) | |
| Velika Plana (5) | 2/2 (100) | 3/3 (100) | |
| Čajetina (5) | 3/4 (75) | 1/1 (100) | |
| Total * (%) | 22/23 (95.6) | 9/10 (90) | 12/13 (92.30) |
| EASTERN REGION | Veliko Gradište (23) | 11/14 (78.5) | 8/9 (88.8) |
| Golubac (1) | 1/1 (100) | 0/0 (0) | |
| Svilajnac (4) | 2/2 (100) | 1/2 (50) | |
| Ražanj (1) | 0/0 (0) | 1/1 (100) | |
| Zaječar (1) | 1/1 (100) | 0/0 (0) | |
| Aleksinac (1) | 0/0 (0) | 1/1 (100) | |
| Niš (1) | 1/1 (100) | 0/0 (0) | |
| Bela Palanka (1) | 1/1 (100) | 0/0 (0) | |
| Total * (%) | 28/33 (84.8) | 17/20 (85) | 11/13 (84.62) |
| Ʃ | 90/114 (78.95) | 51/64 (79.69) | 29/50 (58) |
| Sequence Type | Position of the Variable Site | ||
|---|---|---|---|
| 85 bp | 549 bp | 550 bp | |
| Consensus | A | T | A |
| S4 | G | ||
| S5 | A | ||
| S6 | C | ||
| S7 | G | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuručki, M.; Sukara, R.; Ćirković, V.; Ćirović, D.; Tomanović, S. Molecular Detection and Genetic Variability of Hepatozoon canis in Golden Jackals (Canis aureus L. 1758) in Serbia. Biology 2024, 13, 411. https://doi.org/10.3390/biology13060411
Kuručki M, Sukara R, Ćirković V, Ćirović D, Tomanović S. Molecular Detection and Genetic Variability of Hepatozoon canis in Golden Jackals (Canis aureus L. 1758) in Serbia. Biology. 2024; 13(6):411. https://doi.org/10.3390/biology13060411
Chicago/Turabian StyleKuručki, Milica, Ratko Sukara, Valentina Ćirković, Duško Ćirović, and Snežana Tomanović. 2024. "Molecular Detection and Genetic Variability of Hepatozoon canis in Golden Jackals (Canis aureus L. 1758) in Serbia" Biology 13, no. 6: 411. https://doi.org/10.3390/biology13060411