

Genetic Variability in Mediterranean Coastal Ecosystems: Insights into Ostrea spp. (Bivalvia: Ostreidae)
 , ,
, ,  , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling
2.2. Molecular Analysis
2.3. Sequence Data and Phylogenetic Analysis
2.4. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beck, M.W.; Brumbaugh, R.D.; Airoldi, L.; Carranza, A.; Coen, L.D.; Crawford, C.; Defeo, O.; Edgar, G.J.; Hancock, B.; Kay, M.C.; et al. Oyster reefs at risk and recommendations for conservation, restoration, and management. Bioscience 2011, 61, 107–116. [Google Scholar] [CrossRef]
- Cataudella, S.; Crosetti, D.; Massa, F. Mediterranean coastal lagoons: Sustainable management and interactions among aquaculture, capture fisheries and the environment. General Fisheries Commission for the Mediterranean. Stud. Rev. 2015, 95, I. [Google Scholar]
- Buestel, D.; Ropert, M.; Prou, J.; Goulletquer, P. History, status, and future of oyster culture in France. J. Shellfish Res. 2009, 28, 813–820. [Google Scholar] [CrossRef]
- FAO. Fisheries and Aquaculture Software. FishStatJ, a Tool for Fishery Statistics Analysis. Release: 4.02.07; Berger, T., Sibeni, F., Calderini, F., Eds.; FAO: Rome, Italy, 2022; Available online: https://www.fao.org/fishery/statistics/software/fishstatj/en (accessed on 5 June 2024).
- Günther, R.T. The oyster culture of the ancient Romans. J. Mar. Biol. Assoc. United Kingd. 1897, 4, 360–365. [Google Scholar] [CrossRef]
- MacKenzie, C.L.; Burrell, V.G.; Rosenfield, A.; Hobart, W.L. The History, Present Condition, and Future of the Molluscan Fisheries of North and Central America and Europe: Volume 3; Europe. NOAA/National Marine Fisheries Service, (NOAA Technical Report NMFS, 129); 1997. Available online: https://aquadocs.org/handle/1834/20478 (accessed on 5 June 2024).
- Voultsiadou, E.; Koutsoubas, D.; Achparaki, M. Bivalve mollusc exploitation in Mediterranean coastal communities: An historical approach. J. Biol. Res. 2010, 13, 35–45. [Google Scholar]
- Ruesink, J.L.; Lenihan, H.S.; Trimble, A.C.; Heiman, K.W.; Micheli, F.; Byers, J.E.; Kay, M.C. Introduction of non-native oysters: Ecosystem effects and restoration implications. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 643–689. [Google Scholar] [CrossRef]
- FAO. Cultured Aquatic Species Information Programme. Crassostrea Gigas. Cultured Aquatic Species Information Programme; Helm, M.M., Ed.; FAO Fisheries and Aquaculture Department: Rome, Italy, 2023. [Google Scholar]
- Airoldi, L.; Beck, M.W. Loss, status and trends for coastal marine habitats of Europe. Oceanogr. Mar. Biol. Annu. Rev. 2007, 45, 345–405. [Google Scholar]
- Helmer, L.; Farrell, P.; Hendy, I.; Harding, S.; Robertson, M.; Preston, J. Active management is required to turn the tide for depleted Ostrea edulis stocks from the effects of overfishing, disease and invasive species. PeerJ 2019, 7, e6431. [Google Scholar] [CrossRef]
- Salvi, D.; Macali, A.; Mariottini, P. Molecular phylogenetics and systematics of the bivalve family Ostreidae based on rRNA sequence-structure models and multilocus species tree. PLoS ONE 2014, 9, e108696. [Google Scholar] [CrossRef]
- Wehrmann, A.; Herlyn, M.; Bungenstock, F.; Hertweck, G.; Millat, G. The distribution gap is closed-first record of naturally settled Pacific oysters (Crassostrea gigas) in the East Frisian Wadden Sea, North Sea. Senckenberg. Maritima 2000, 30, 153–160. [Google Scholar] [CrossRef]
- Miossec, L.; Le Deuff, R.-M.; Goulletquer, P. Alien Species Alert: Crassostrea Gigas (Pacific Oyster); ICES Cooperative Research Report No. 299; 2009; 42p. Available online: https://ices-library.figshare.com/articles/report/Alien_species_alert_Crassostrea_gigas_Pacific_oyster_/18624182?file=33403241 (accessed on 5 June 2024).
- Kennedy, R.J.; Roberts, D. A survey of the current status of the flat oyster Ostrea edulis in Strangford Lough, Northern Ireland, with a view to the restoration of its oyster beds. In Biology and Environment: Proceedings of the Royal Irish Academy; Royal Irish Academy: Dublin, Ireland, 1999; Volume 99B, pp. 79–88. [Google Scholar]
- Herbert RJ, H.; Humphreys, J.; Davies, C.J.; Roberts, C.; Fletcher, S.; Crowe, T.P. Ecological impacts of non-native Pacific oysters (Crassostrea gigas) and management measures for protected areas in Europe. Biodivers. Conserv. 2016, 25, 2835–2865. [Google Scholar] [CrossRef]
- Launey, S.; Ledu, C.; Boudry, P.; Bonhomme, F.; Naciri-Graven, Y. Geographic structure in the European flat oyster (Ostrea edulis L.) as revealed by microsatellite polymorphism. J. Hered. 2002, 93, 331–351. [Google Scholar] [CrossRef] [PubMed]
- Gaffney, P.M. The role of genetics in shellfish restoration. Aquat. Living Resour. 2006, 19, 277–282. [Google Scholar] [CrossRef]
- Yu, D.H.; Chu, K.H. Genetic variation in wild and cultured populations of the pearl oyster Pinctada fucata from southern China. Aquaculture 2006, 258, 220–227. [Google Scholar] [CrossRef]
- Hedgecock, D.; Banks, M.A.; McGoldrick, D.J. The status of the Kumamoto oyster Crassostrea sikamea (Amemiya 1928) in US commercial brood stocks. J. Shellfish Res. 1993, 12, 215–221. [Google Scholar]
- Tan, K.S.; Zhang, H.; Liu, H.; Cheng, D.; Ye, T.; Ma, H.; Zheng, H. Enhancing lipid nutritional quality of oysters by hybridization between Crassostrea gigas and C. angulata. Aquac. Res. 2019, 50, 3776–3782. [Google Scholar] [CrossRef]
- Meng, L.; Li, Q.; Xu, C.; Liu, S.; Kong, L.; Yu, H. Hybridization improved stress resistance in the Pacific oyster: Evidence from physiological and immune responses. Aquaculture 2021, 545, 737227. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Zhang, Y.; Ma, H.; Xiao, S.; Xiang, Z.; Yu, Z. Performance evaluation of reciprocal hybrids derived from the two brackish oysters, Crassostrea hongkongensis and Crassostrea sikamea in southern China. Aquaculture 2017, 473, 310–316. [Google Scholar] [CrossRef]
- Hajovsky, P.; Beseres Pollack, J.; Anderson, J. Morphological Assessment of the Eastern Oyster Crassostrea virginica throughout the Gulf of Mexico. Mar. Coast. Fish. 2021, 13, 309–319. [Google Scholar] [CrossRef]
- Bayne, B.L. Biology of Oysters; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Guo, X.; Li, C.; Wang, H.; Xu, Z. Diversity and evolution of living oysters. J. Shellfish Res. 2018, 37, 755–771. [Google Scholar] [CrossRef]
- Harry, H.W. Synopsis of the supraspecific classification of living oysters (Bivalvia: Gryphaeidae and Ostreidae). Veliger 1985, 28, 121–158. [Google Scholar]
- Coan, E.V.; Valentich-Scott, P.; Bernard, F.R. Bivalve Seashells of Western North America: Marine Bivalve Mollusks from Artic Alaska to Baja California; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 2000; 746p. [Google Scholar]
- Kirkendale, L.; Lee, T.; Baker, P.; Foighil, D.O. Oysters of the Conch Republic (Florida keys): A molecular phylogenetic study of Parahyotissa mcgintyi, Teskeyostrea weberi and Ostreola equestris. Malacologia 2004, 46, 309–326. [Google Scholar]
- Lapègue, S.; Salah, I.B.; Batista, F.M.; Heurtebise, S.; Neifar, L.; Boudry, P. Phylogeographic study of the dwarf oyster, Ostreola stentina, from Morocco, Portugal and Tunisia: Evidence of a geographic disjunction with the closely related taxa, Ostrea aupouria and Ostreola equestris. Mar. Biol. 2006, 150, 103–110. [Google Scholar] [CrossRef]
- Shilts, M.H.; Pascual, M.S.; Foighil, D.O. Systematic, taxonomic and biogeographic relationships of Argentine flat oysters. Mol. Phylogenetics Evol. 2007, 44, 467–473. [Google Scholar] [CrossRef]
- Polson, M.P.; Hewson, W.E.; Eernisse, D.J.; Baker, P.K.; Zacherl, D.C. You say conchaphila, I say lurida: Molecular evidence for restricting the Olympia oyster (Ostrea lurida Carpenter 1864) to temperate western North America. J. Shellfish Res. 2009, 28, 11–21. [Google Scholar] [CrossRef]
- Salvi, D.; Mariottini, P. Revision shock in Pacific oysters taxonomy: The genus Magallana (formerly Crassostrea in part) is well-founded and necessary. Zool. J. Linn. Soc. 2021, 192, 43–58. [Google Scholar] [CrossRef]
- Hamaguchi, M.; Manabe, M.; Kajihara, N.; Shimabukuro, H.; Yamada, Y.; Nishi, E. DNA barcoding of flat oyster species reveals the presence of Ostrea stentina Payraudeau, 1826 (Bivalvia: Ostreidae) in Japan. Mar. Biodivers. Rec. 2017, 10, 4. [Google Scholar] [CrossRef]
- Hu, L.; Wang, H.; Zhang, Z.; Li, C.; Guo, X. Classification of small flat oysters of Ostrea stentina species complex and a new species Ostrea neostentina sp. nov. (Bivalvia: Ostreidae). J. Shellfish Res. 2019, 38, 295–308. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Bertolini, C.; Pastres, R. Understanding the influence of swarming timing on the dispersal of Ostrea edulis larvae in the Northern Adriatic Sea. Open Res. Eur. 2023, 3, 212. [Google Scholar] [CrossRef]
- Raith, M.; Zacherl, D.C.; Pilgrim, E.M.; Eernisse, D.J. Phylogeny and species diversity of Gulf of California oysters (Ostreidae) inferred from mitochondrial DNA. Am. Malacol. Bull. 2015, 33, 263–283. [Google Scholar] [CrossRef]
- Kenchington, E.; Bird, C.J.; Osborne, J.; Reith, M. Novel repeat elements in the nuclear ribosomal RNA operon of the flat oysters Ostrea edulis Linnaeus, 1758 and O. angasi Sowerby, 1871. J. Shellfish Res. 2002, 21, 697–705. [Google Scholar]
- Hurwood, D.A.; Heasman, M.P.; Mather, P.B. Gene flow, colonisation and demographic history of the flat oyster Ostrea angasi. Mar. Freshw. Res. 2005, 56, 1099–1106. [Google Scholar] [CrossRef]
- Salvi, D.; Mariottini, P. Molecular taxonomy in 2D: A novel ITS2 rRNA sequence-structure approach guides the description of the oysters’ subfamily Saccostreinae and the genus Magallana (Bivalvia: Ostreidae). Zool. J. Linn. Soc. 2017, 179, 263–276. [Google Scholar] [CrossRef]
- Sutton, J.T.; Nishimoto, J.; Schrader, J.; Agonias, K.; Antonio, N.; Bautista, B.; Haws, M. Genetic analysis identifies the Ostrea stentina/aupouria/equestris oyster species complex in Hawai’i, and resolves its lineage as the western Pacific O. equestris. bioRxiv 2020. [Google Scholar] [CrossRef]
- Crocetta, F.; Bitar, G.; Zibrowius, H.; Oliverio, M. Biogeographical homogeneity in the eastern Mediterranean Sea. II. Temporal variation in Lebanese bivalve biota. Aquat. Biol. 2013, 19, 75–84. [Google Scholar] [CrossRef]
- Pejovic, I.; Ardura, A.; Miralles, L.; Arias, A.; Borrell, Y.J.; Garcia-Vazquez, E. DNA barcoding for assessment of exotic molluscs associated with maritime ports in northern Iberia. Mar. Biol. Res. 2016, 12, 168–176. [Google Scholar] [CrossRef]
- Wang, H.; Qian, L.; Liu, X.; Zhang, G.; Guo, X. Classification of a common cupped oyster from southern China. J. Shellfish Res. 2010, 29, 857–866. [Google Scholar] [CrossRef]
- Li, C.; Haws, M.; Wang, H.; Guo, X. Taxonomic classification of three oyster (Ostreidae) species from Myanmar. J. Shellfish Res. 2017, 36, 365–372. [Google Scholar] [CrossRef]
- Li, C.; Wang, H.; Guo, X. Classification and taxonomic revision of two oyster species from Peru: Ostrea megodon (Hanley, 1846) and Crassostrea talonata (Li & Qi, 1994). J. Shellfish Res. 2017, 36, 359–365. [Google Scholar] [CrossRef]
- Willer, D.F.; Aldridge, D.C. Sustainable bivalve farming can deliver food security in the tropics. Nat. Food 2020, 1, 384–388. [Google Scholar] [CrossRef]
- Grazioli, E.; Guerranti, C.; Pastorino, P.; Esposito, G.; Bianco, E.; Simonetti, E.; Ranis, S.; Renzi, M.; Terlizzi, A. Review of the scientific literature on biology, ecology, and aspects related to the fishing sector of the striped Venus (Chamelea gallina) in Northern Adriatic Sea. J. Mar. Sci. Eng. 2022, 10, 1328. [Google Scholar] [CrossRef]
- Prado, P.; Fernández, M.; Cordero, D.; Saavedra, C.; Carella, F.; Alcaraz, C.; Gairin, I. Molecular identification, life cycle characterization, and hatchery seed production of dwarf oysters from the Ebro Delta (Spain). Aquat. Living Resour. 2022, 35, 5. [Google Scholar] [CrossRef]

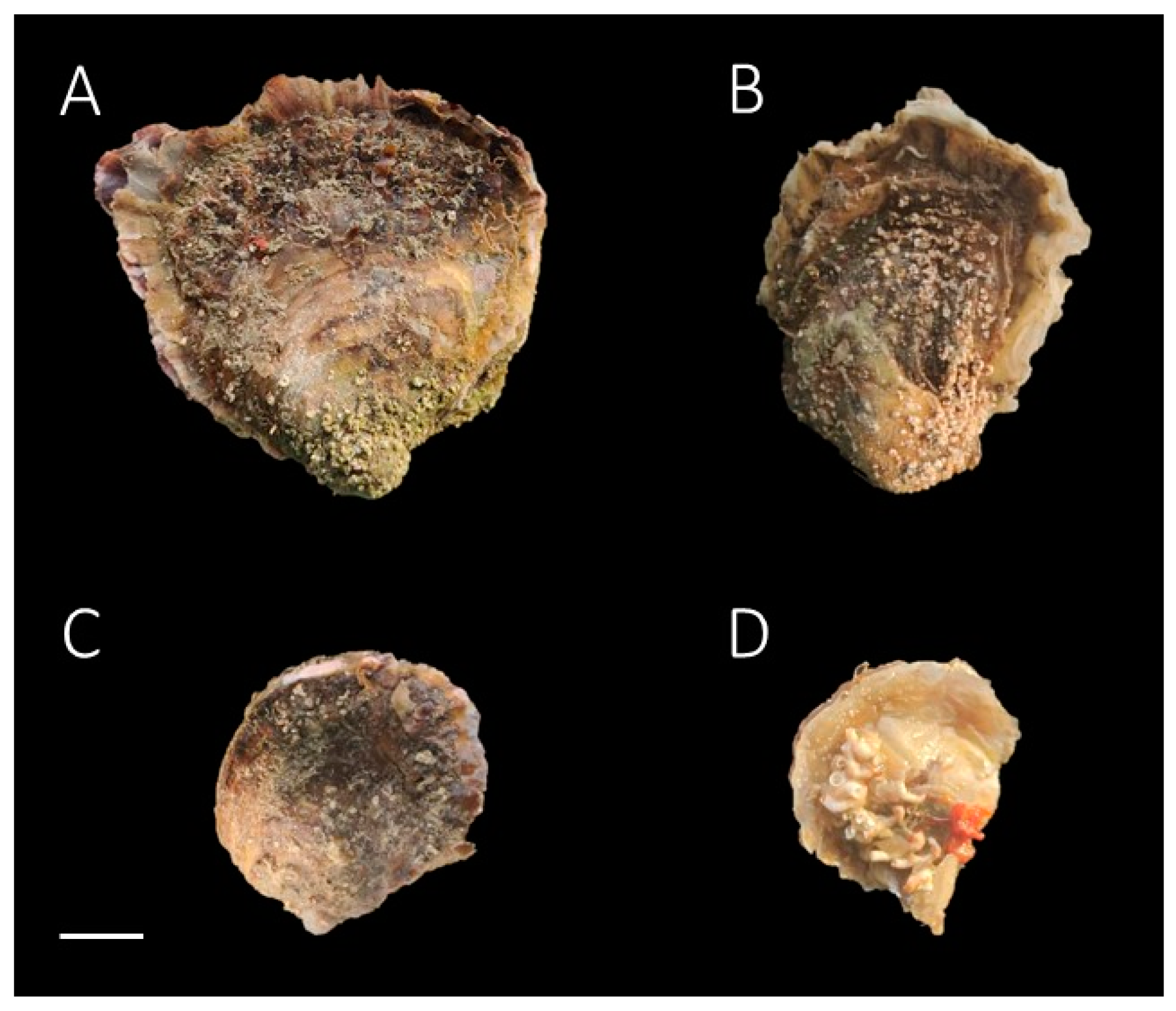
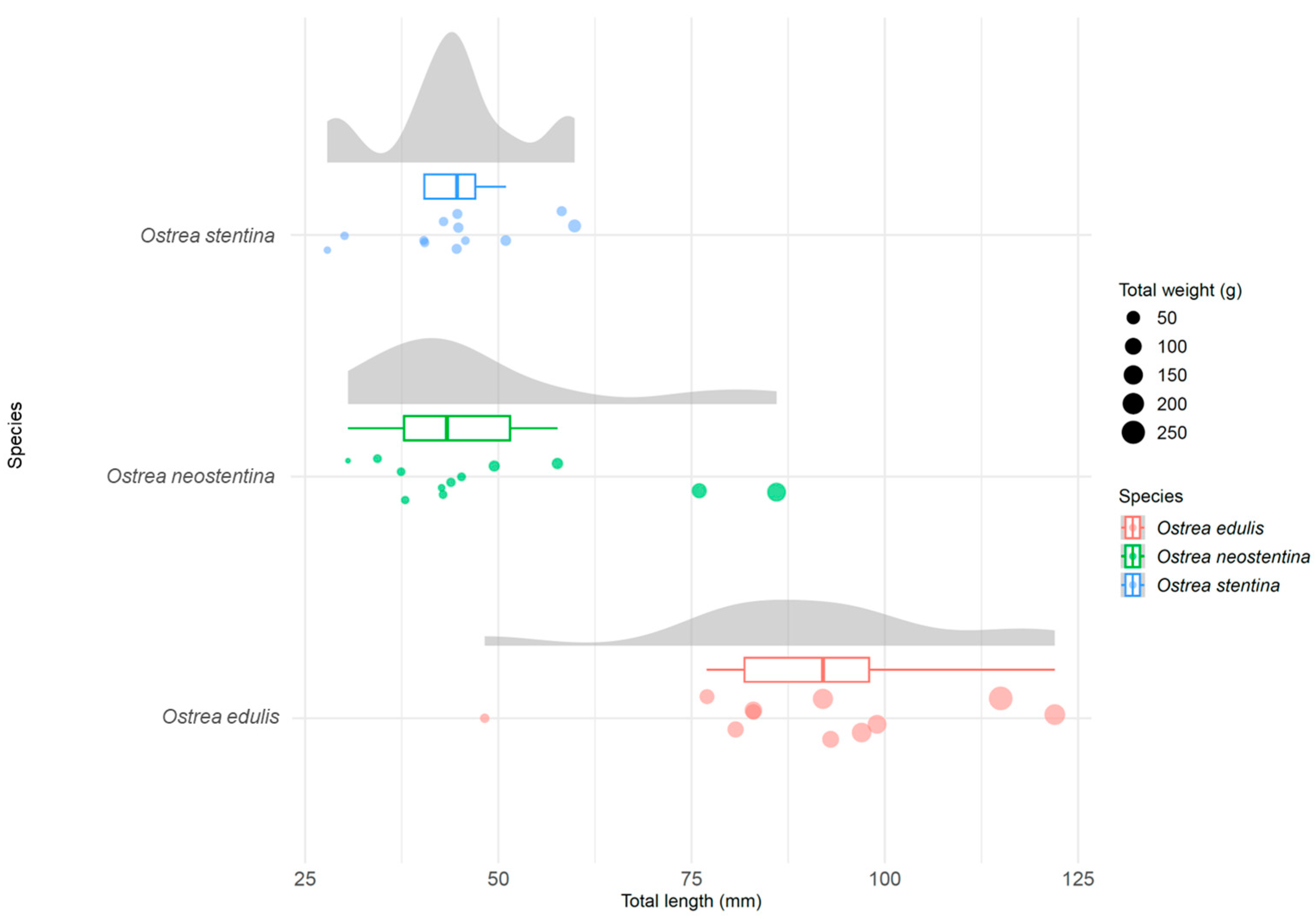
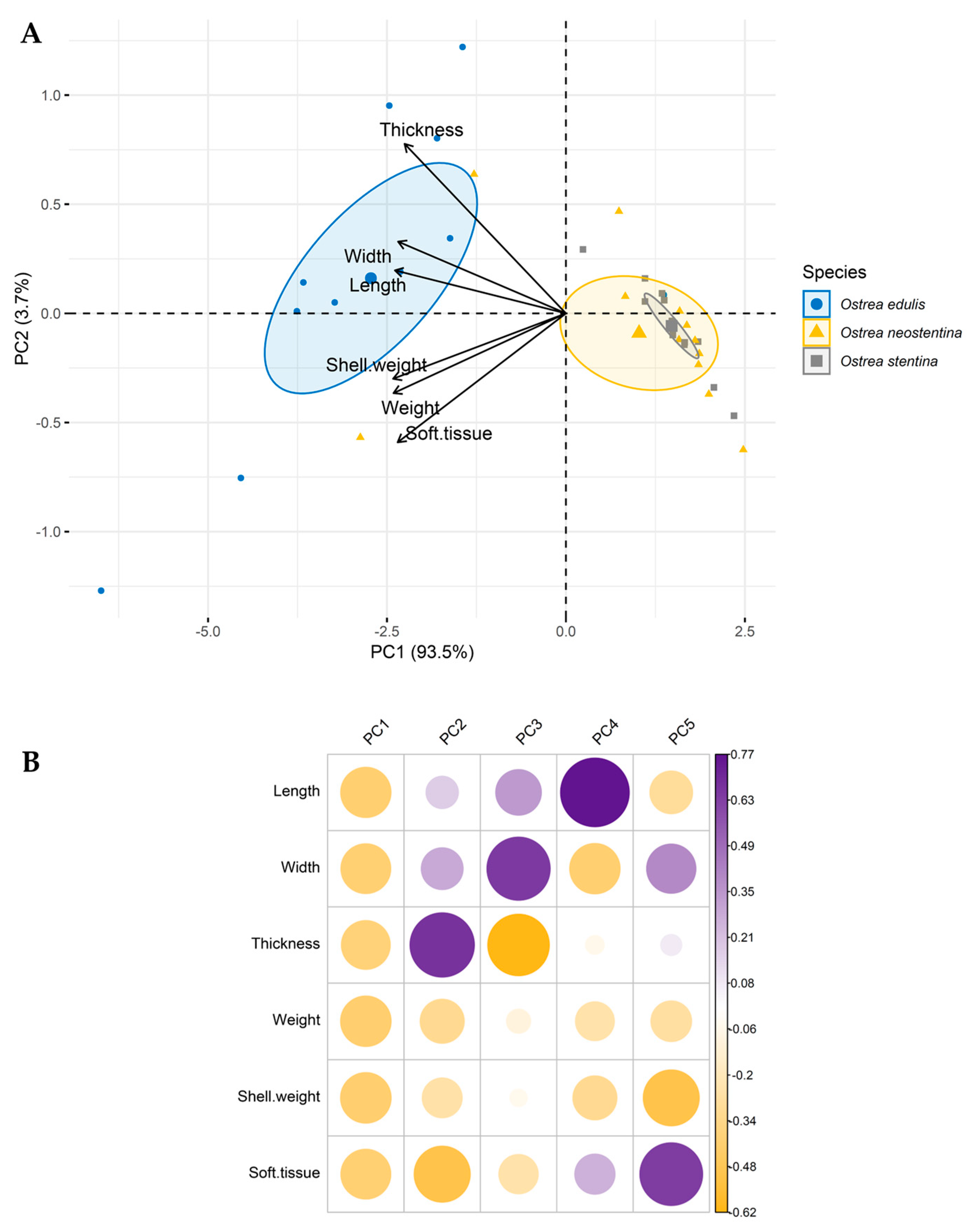
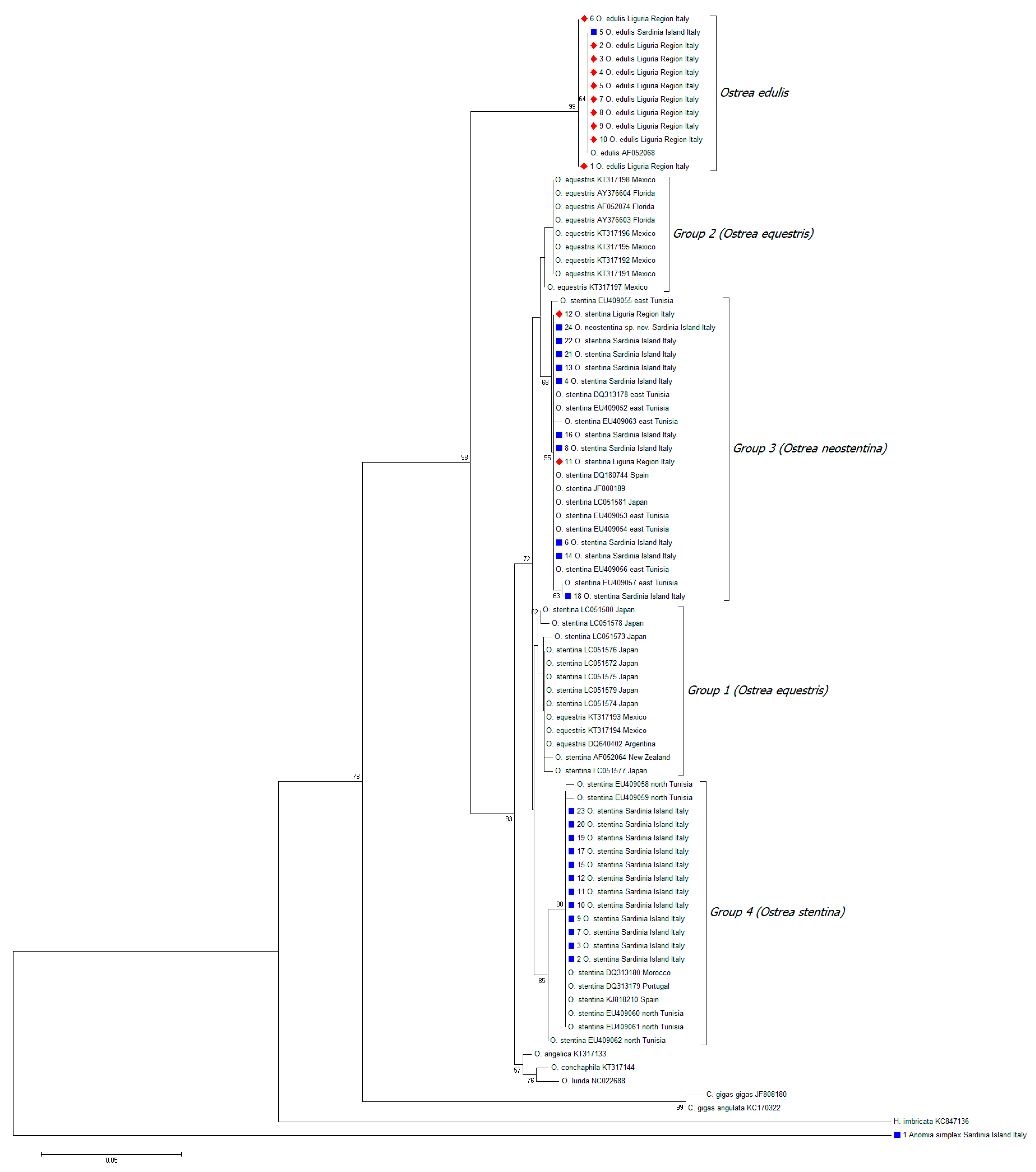

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | No. | Species (Morphology) | Species (After Phylogeny) | Total Length (mm) | Width (mm) | Thickness (mm) | Total Weight (g) | Shell Weight (g) | Soft Tissue (g) |
|---|---|---|---|---|---|---|---|---|---|
| Liguria S1 | 1 | Ostrea edulis | O. edulis | 80.7 | 81 | 25 | 90.8 | 76.04 | 6.39 |
| 2 | O. edulis | O. edulis | 99 | 77 | 28 | 140.51 | 109.73 | 16.57 | |
| 3 | O. edulis | O. edulis | 122 | 91 | 23 | 184.69 | 147.27 | 22.65 | |
| 4 | O. edulis | O. edulis | 115 | 86 | 37 | 263.69 | 199.85 | 35.62 | |
| 5 | O. edulis | O. edulis | 97 | 79 | 32 | 157.89 | 118.59 | 18.12 | |
| 6 | O. edulis | O. edulis | 92 | 71 | 34 | 171.92 | 143.84 | 17.22 | |
| 7 | O. edulis | O. edulis | 77 | 70 | 30 | 69.23 | 60.09 | 5.76 | |
| 8 | O. edulis | O. edulis | 83 | 76 | 25 | 115.91 | 94.45 | 11.88 | |
| 9 | O. edulis | O. edulis | 83 | 61 | 26 | 79.54 | 65.02 | 12.37 | |
| 10 | O. edulis | O. edulis | 93 | 66 | 34 | 102.24 | 84.93 | 11.16 | |
| 11 | O. stentina | O. neostentina | 76 | 57 | 28 | 74.72 | 59.39 | 9.26 | |
| 12 | O. stentina | O. neostentina | 86 | 80 | 21 | 146.19 | 120.56 | 16.03 | |
| Sardinia S2 | 1 | Anomia simplex | A. simplex | 49.17 | 45.82 | 7.02 | 8.38 | 6.75 | 1.28 |
| 2 | O. stentina | O. stentina | 59.87 | 46.22 | 19.04 | 42.68 | 31.95 | 4.47 | |
| 3 | O. stentina | O. stentina | 50.96 | 42.86 | 12.81 | 20.80 | 16.04 | 2.02 | |
| 4 | O. stentina | O. neostentina | 57.65 | 45.66 | 13.05 | 27.95 | 21.26 | 2.32 | |
| 5 | O. edulis | O. edulis | 48.24 | 39.47 | 12.52 | 12.04 | 10.26 | 1.26 | |
| 6 | O. stentina | O. neostentina | 49.47 | 44.63 | 18.91 | 26.45 | 22.31 | 2.07 | |
| 7 | O. stentina | O. stentina | 58.20 | 41.91 | 12.55 | 16.64 | 15.26 | 0.96 | |
| 8 | O. stentina | O. neostentina | 43.87 | 34.52 | 11.23 | 12.85 | 11.37 | 1.26 | |
| 9 | O. stentina | O. stentina | 42.91 | 32.59 | 11.17 | 11.78 | 10.39 | 1.18 | |
| 10 | O. stentina | O. stentina | 44.70 | 37.14 | 14.07 | 15.88 | 12.65 | 1.63 | |
| 11 | O. stentina | O. stentina | 44.61 | 37.80 | 11.58 | 16.22 | 12.22 | 0.81 | |
| 12 | O. stentina | O. stentina | 44.83 | 38.09 | 13.13 | 17.64 | 13.71 | 1.01 | |
| 13 | O. stentina | O. neostentina | 42.85 | 32.30 | 11.79 | 8.99 | 7.90 | 0.84 | |
| 14 | O. stentina | O. neostentina | 42.66 | 32.01 | 6.79 | 4.73 | 3.23 | 0.48 | |
| 15 | O. stentina | O. stentina | 40.36 | 30.90 | 10.82 | 9.76 | 2.17 | 0.69 | |
| 16 | O. stentina | O. neostentina | 37.95 | 35.26 | 9.02 | 7.60 | 6.68 | 0.74 | |
| 17 | O. stentina | O. stentina | 45.74 | 41.07 | 10.09 | 7.80 | 8.81 | 1.81 | |
| 18 | O. stentina | O. neostentina | 34.36 | 33.06 | 10.67 | 9.82 | 6.26 | 0.90 | |
| 19 | O. stentina | O. stentina | 40.47 | 35.54 | 10.94 | 10.67 | 8.48 | 1.37 | |
| 20 | O. stentina | O. stentina | 30.10 | 32.15 | 8.80 | 6.90 | 4.77 | 0.81 | |
| 21 | O. stentina | O. neostentina | 37.42 | 30.72 | 11.78 | 9.19 | 7.02 | 1.05 | |
| 22 | O. stentina | O. neostentina | 45.24 | 31.13 | 13.07 | 9.90 | 8.51 | 1.21 | |
| 23 | O. stentina | O. stentina | 27.88 | 23.26 | 8.37 | 3.72 | 3.07 | 0.50 | |
| 24 | O. neostentina | O. neostentina | 30.55 | 22.33 | 5.75 | 1.83 | 1.29 | 0.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esposito, G.; Peletto, S.; Guo, X.; Pastorino, P.; Arillo, A.; Martini, I.; Acutis, P.L.; Mugetti, D.; Meloni, D.; Prearo, M.; et al. Genetic Variability in Mediterranean Coastal Ecosystems: Insights into Ostrea spp. (Bivalvia: Ostreidae). Biology 2024, 13, 702. https://doi.org/10.3390/biology13090702
Esposito G, Peletto S, Guo X, Pastorino P, Arillo A, Martini I, Acutis PL, Mugetti D, Meloni D, Prearo M, et al. Genetic Variability in Mediterranean Coastal Ecosystems: Insights into Ostrea spp. (Bivalvia: Ostreidae). Biology. 2024; 13(9):702. https://doi.org/10.3390/biology13090702
Chicago/Turabian StyleEsposito, Giuseppe, Simone Peletto, Ximing Guo, Paolo Pastorino, Alessandra Arillo, Isabella Martini, Pier Luigi Acutis, Davide Mugetti, Domenico Meloni, Marino Prearo, and et al. 2024. "Genetic Variability in Mediterranean Coastal Ecosystems: Insights into Ostrea spp. (Bivalvia: Ostreidae)" Biology 13, no. 9: 702. https://doi.org/10.3390/biology13090702