Sampling Method Affects HR-MAS NMR Spectra of Healthy Caprine Brain Biopsies

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
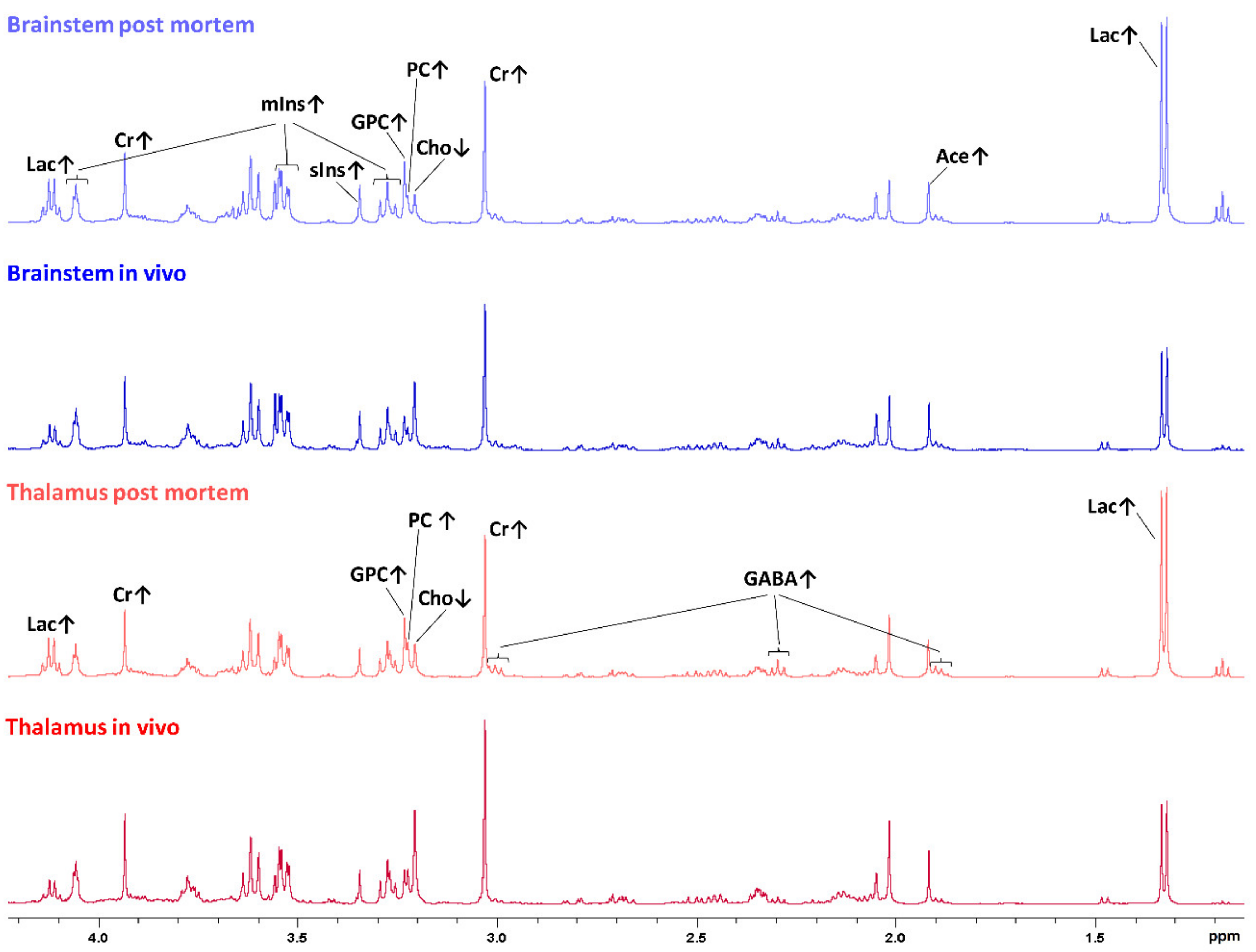
2.1. Metabolic Profiles of In Vivo and Postmortem Samples of the Brainstem and Thalamus with Regard to the Individual Animal
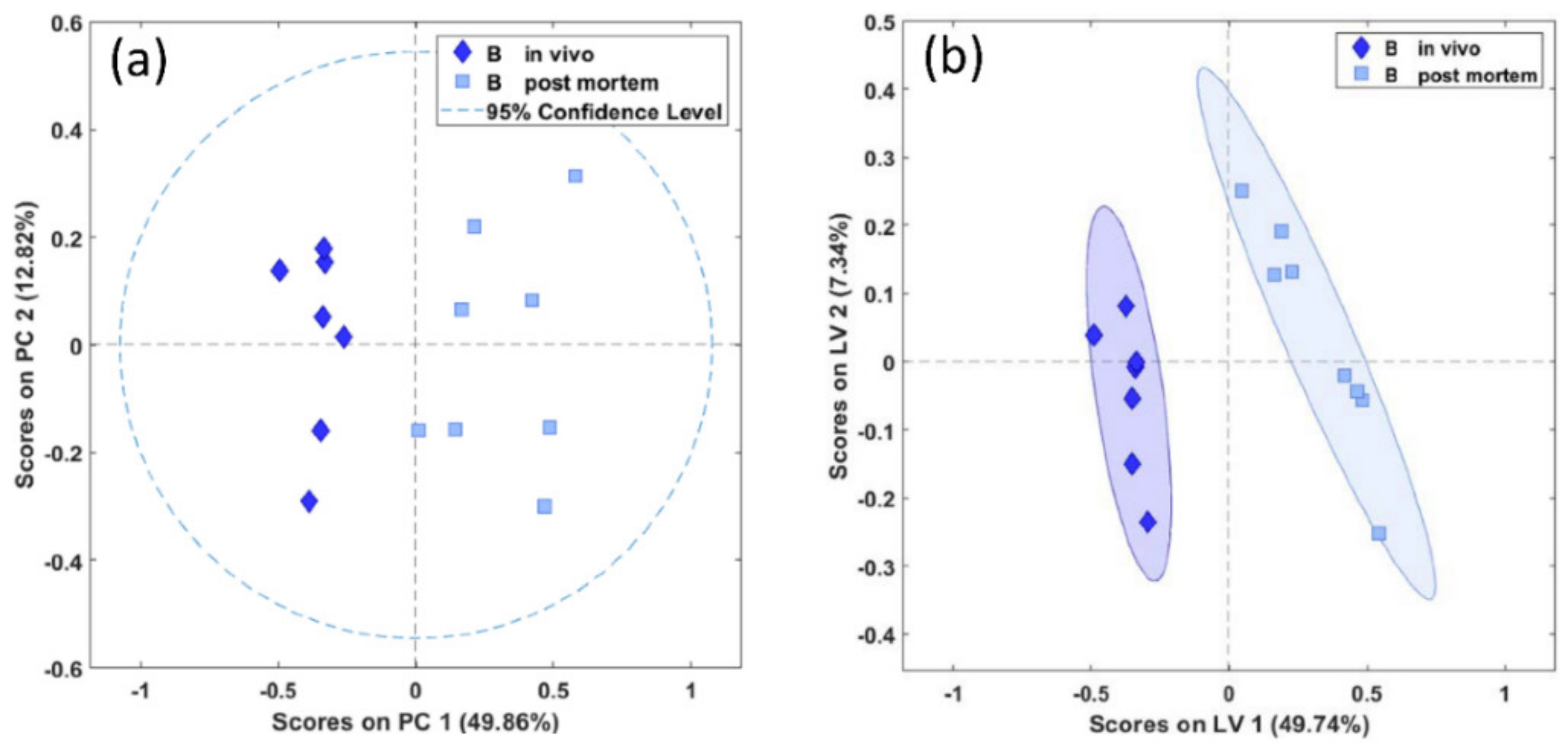
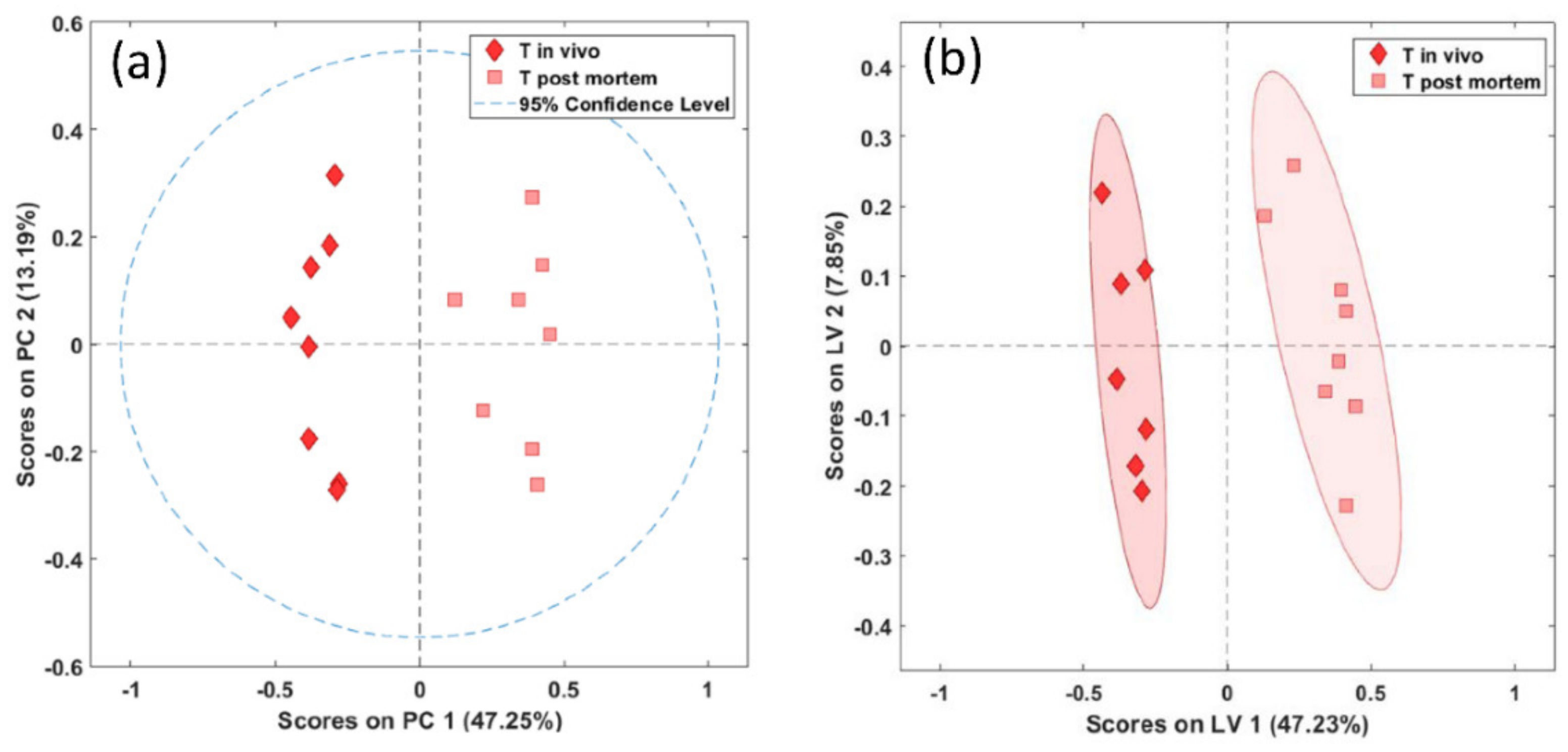
2.2. Metabolic Profiles of In Vivo and Postmortem Samples with Regard to the Brain Region (Brainstem and Thalamus)
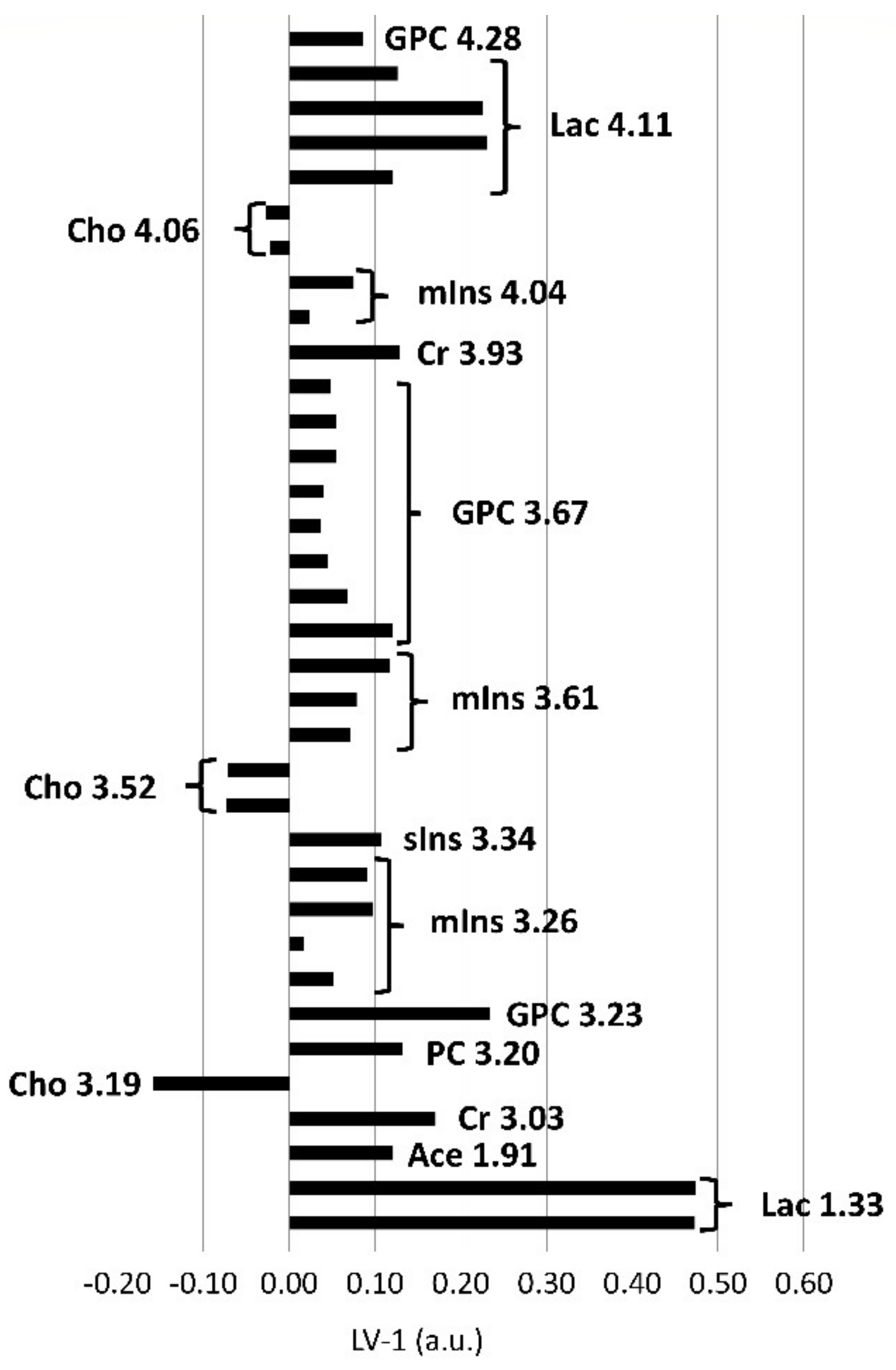
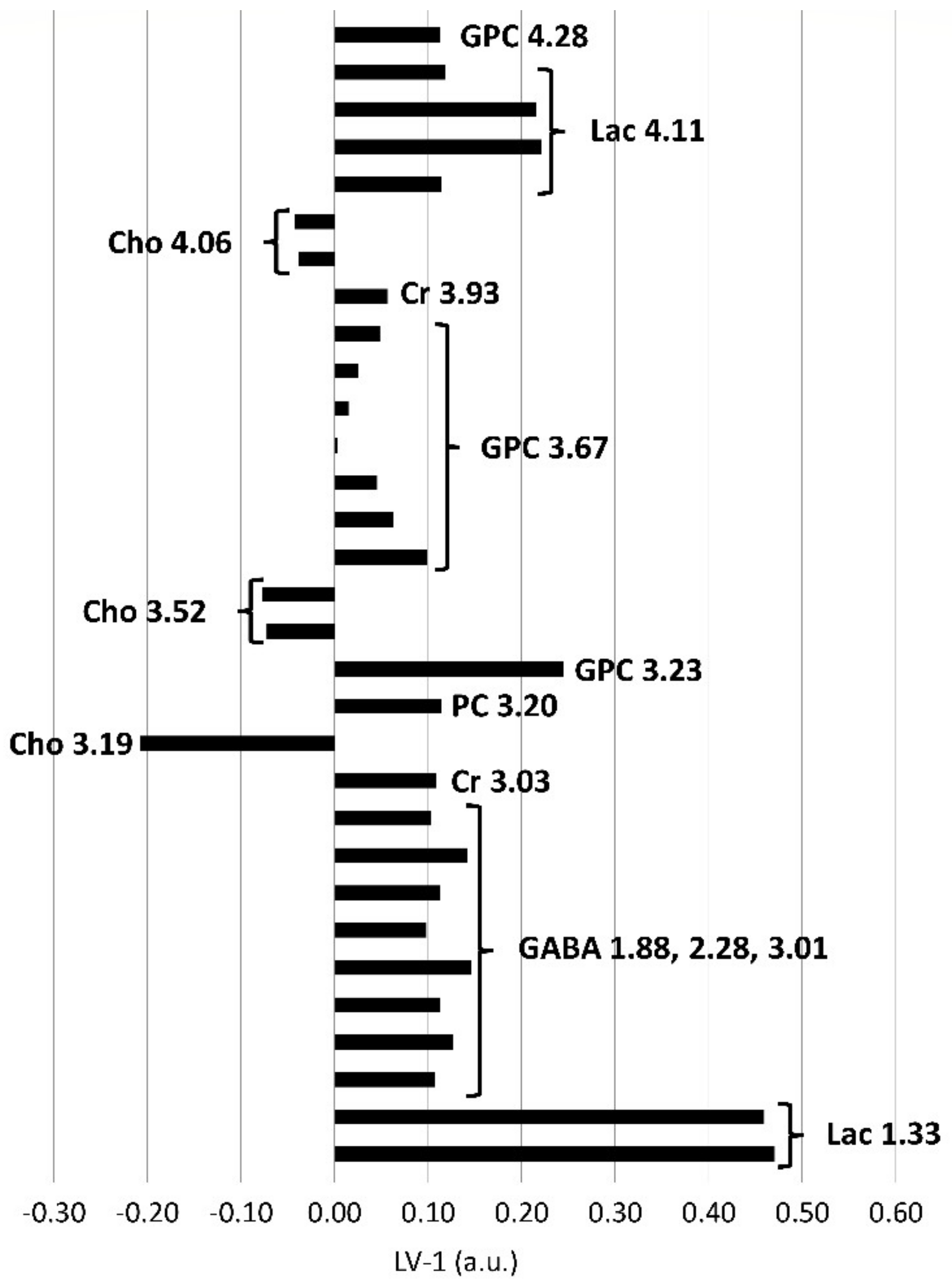
2.3. Metabolic Profiles of the Brainstem and Thalamus with Regard to the Sampling Method (In Vivo and Postmortem)
3. Discussion
3.1. Regional Differences in the Metabolic Profile
3.2. Increased Lactate as a Sign for the Switch of Energy Metabolism to Anaerobic Glycolysis
3.3. Creatine as a Marker of Impaired Brain Energy Homeostasis
3.4. Increase of Acetate and ƴ-Aminobutyric Acid (GABA) May Reflect the Breakdown of Lipids and Myelin
3.5. Alterations in the Choline Group and Increased Concentrations of Inositol Sugars Suggest Membrane Damage
3.6. Limitations
4. Materials and Methods
4.1. Brain Biopsies and Sampling Methods
4.2. 1H High-Resolution–Magic Angle Spinning (HR-MAS) NMR Spectroscopy
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gonzalez-Riano, C.; Garcia, A.; Barbas, C. Metabolomics studies in brain tissue: A review. J. Pharm. Biomed. Anal. 2016, 130, 141–168. [Google Scholar] [CrossRef]
- Sitter, B.; Bathen, T.F.; Tessem, M.B.; Gribbestad, I.S. High-resolution magic angle spinning (HR MAS) MR spectroscopy in metabolic characterization of human cancer. Prog. Nucl. Magn. Reson. Spectrosc. 2009, 54, 239–254. [Google Scholar] [CrossRef]
- Fuss, T.L.; Cheng, L.L. Evaluation of cancer metabolomics using ex vivo high resolution magic angle spinning (HRMAS) magnetic resonance spectroscopy (MRS). Metabolites 2016, 6, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.L.; Chang, I.W.; Louis, D.N.; Gonzalez, R.G. Correlation of high-resolution magic angle spinning proton magnetic resonance spectroscopy with histopathology of intact human brain tumor specimens. Cancer Res. 1998, 58, 1825–1832. [Google Scholar] [PubMed]
- Wright, A.J.; Fellows, G.A.; Griffiths, J.R.; Wilson, M.; Bell, B.A.; Howe, F.A. Ex-Vivo HRMAS of Adult Brain Tumors: Metabolite Quantification and Assignment of Tumor Biomarkers. Mol. Cancer 2010, 9, 66. [Google Scholar] [CrossRef] [Green Version]
- Opstad, K.S.; Bell, B.A.; Griffiths, J.R.; Howe, F.A. An assessment of the effects of sample ischaemia and spinning time on the metabolic profile of brain tumour biopsy specimens as determined by high-resolution magic angle spinning 1 H NMR. NMR Biomed. 2008, 21, 1138–1147. [Google Scholar] [CrossRef]
- Tsang, T.M.; Griffin, J.L.; Haselden, J.; Fish, C.; Holmes, E. Metabolic Characterization of Distinct Neuroanatomical Regions in Rats by Magic Angle Spinning 1 H Nuclear Magnetic Resonance Spectroscopy. Magn. Reson. Med. 2005, 53, 1018–1024. [Google Scholar] [CrossRef]
- Geddes, J.W.; Chang, N.G.; Ackley, D.C.; Soultanian, N.S.; McGillis, J.P.; Yokel, R.A. Postmortem elevation in extracellular glutamate in the rat hippocampus when brain temperature is maintained at physiological levels: Implications for the use of human brain autopsy tissues. Brain Res. 1999, 831, 104–112. [Google Scholar] [CrossRef]
- Peeling, J.; Wong, D.; Sutherland, G.R. Nuclear magnetic resonance study of regional metabolism after forebrain ischemia in rats. Stroke 1989, 20, 633–640. [Google Scholar] [CrossRef] [Green Version]
- Shank, R.P.; Aprison, M.H. Post mortem changes in the content and specific radioactivity of several amino acids in four areas of the rat brain. J. Neurobiol. 1971, 2, 145–151. [Google Scholar] [CrossRef]
- Petroff, O.A.C.; Ogino, T.; Alger, J.R. High-Resolution Proton Magnetic Resonance Spectroscopy of Rabbit Brain: Regional Metabolite Levels and Postmortem Changes. J. Neurochem. 1988, 51, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Perry, T.L.; Hansen, S.; Gandham, S. Postmortem Changes of Amino Compounds in Human and Rat Brain. J. Neurochem. 1981, 36, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Precht, C.; Diserens, G.; Vermathen, M.; Oevermann, A.; Lauper, J.; Vermathen, P. Metabolic profiling of listeria rhombencephalitis in small ruminants by 1 H high-resolution magic angle spinning NMR spectroscopy. NMR Biomed. 2018, 31, e4023. [Google Scholar] [CrossRef] [PubMed]
- Govindaraju, V.; Young, K.; Maudsley, A.A. Proton NMR chemical shifts and coupling constants for brain metabolites. NMR Biomed. 2000, 13, 129–153. [Google Scholar] [CrossRef]
- Duarte, J.M.N.; Lei, H.; Mlynárik, V.; Gruetter, R. The neurochemical profile quantified by in vivo 1H NMR spectroscopy. Neuroimage 2012, 61, 342–362. [Google Scholar] [CrossRef] [Green Version]
- Scheurer, E.; Ith, M.; Dietrich, D.; Kreis, R.; Hüsler, J.; Dirnhofer, R.; Boesch, C. Statistical evaluation of time-dependent metabolite concentrations: Estimation of post-mortem intervals based on in situ 1H-MRS of the brain. NMR Biomed. 2005, 18, 163–172. [Google Scholar] [CrossRef]
- Musshoff, F.; Klotzbach, H.; Block, W.; Traeber, F.; Schild, H.; Madea, B. Comparison of post-mortem metabolic changes in sheep brain tissue in isolated heads and whole animals using 1H-MR spectroscopy-preliminary results. Int. J. Leg. Med. 2011, 125, 741–744. [Google Scholar] [CrossRef]
- Rae, C.D. A Guide to the Metabolic Pathways and Function of Metabolites Observed in Human Brain 1 H Magnetic Resonance Spectra. Neurochem. Res. 2014, 39, 1–36. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Michaelis, T.; Helms, G.; Frahm, J. Metabolic alterations in brain autopsies: Proton NMR identification of free glycerol. NMR Biomed. 1996, 9, 121–124. [Google Scholar] [CrossRef]
- Fritschy, J.; Meskenaite, V.; Weinmann, O.; Honer, M.; Benke, D.; Mohler, H. GABA B-receptor splice variants GB1a and GB1b in rat brain: Developmental regulation, cellular distribution and extrasynaptic localization. Eur. J. Neurosci. 1999, 11, 761–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valonen, P.K.; Griffin, J.L.; Lehtimäki, K.K.; Liimatainen, T.; Nicholson, J.K.; Gröhn, O.H.J.; Kauppinen, R.A. High-resolution magic-angle-spinning 1H NMR spectroscopy reveals different responses in choline-containing metabolites upon gene therapy-induced programmed cell death in rat brain glioma. NMR Biomed. 2005, 18, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W.; Kim, S.Y.; Lee, T.; Nam, Y.K.; Ju, A.; Woo, D.C.; You, S.J.; Han, J.S.; Lee, S.H.; Choi, C.B.; et al. Ex vivo detection for chronic ethanol consumption-induced neurochemical changes in rats. Brain Res. 2012, 1429, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Jernerén, F.; Söderquist, M.; Karlsson, O. Post-sampling release of free fatty acids—Effects of heat stabilization and methods of euthanasia. J. Pharmacol. Toxicol. Methods 2015, 71, 13–20. [Google Scholar] [CrossRef]
- Staudacher, A.; Oevermann, A.; Stoffel, M.H.; Gorgas, D. Validation of a magnetic resonance imaging guided stereotactic access to the ovine brainstem. BMC Vet. Res. 2014, 10, 216. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, J.; Nilsson, M.; Bodenhausen, G.; Morris, G. Spin echo NMR spectra without J modulation. Chem. Commun. 2012, 48, 811–813. [Google Scholar] [CrossRef] [Green Version]
- Diserens, G.; Vermathen, M.; Zurich, M.; Vermathen, P. Longitudinal investigation of the metabolome of 3D aggregating brain cell cultures at different maturation stages by 1 H HR-MAS NMR. Anal. Bioanal. Chem. 2018, 410, 6733–6749. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Häni, A.; Diserens, G.; Oevermann, A.; Vermathen, P.; Precht, C. Sampling Method Affects HR-MAS NMR Spectra of Healthy Caprine Brain Biopsies. Metabolites 2021, 11, 38. https://doi.org/10.3390/metabo11010038
Häni A, Diserens G, Oevermann A, Vermathen P, Precht C. Sampling Method Affects HR-MAS NMR Spectra of Healthy Caprine Brain Biopsies. Metabolites. 2021; 11(1):38. https://doi.org/10.3390/metabo11010038
Chicago/Turabian StyleHäni, Annakatrin, Gaëlle Diserens, Anna Oevermann, Peter Vermathen, and Christina Precht. 2021. "Sampling Method Affects HR-MAS NMR Spectra of Healthy Caprine Brain Biopsies" Metabolites 11, no. 1: 38. https://doi.org/10.3390/metabo11010038
APA StyleHäni, A., Diserens, G., Oevermann, A., Vermathen, P., & Precht, C. (2021). Sampling Method Affects HR-MAS NMR Spectra of Healthy Caprine Brain Biopsies. Metabolites, 11(1), 38. https://doi.org/10.3390/metabo11010038