
Human Stool Metabolome Differs upon 24 h Blood Pressure Levels and Blood Pressure Dipping Status: A Prospective Longitudinal Study

 , , ,
, , ,
Abstract
:1. Introduction
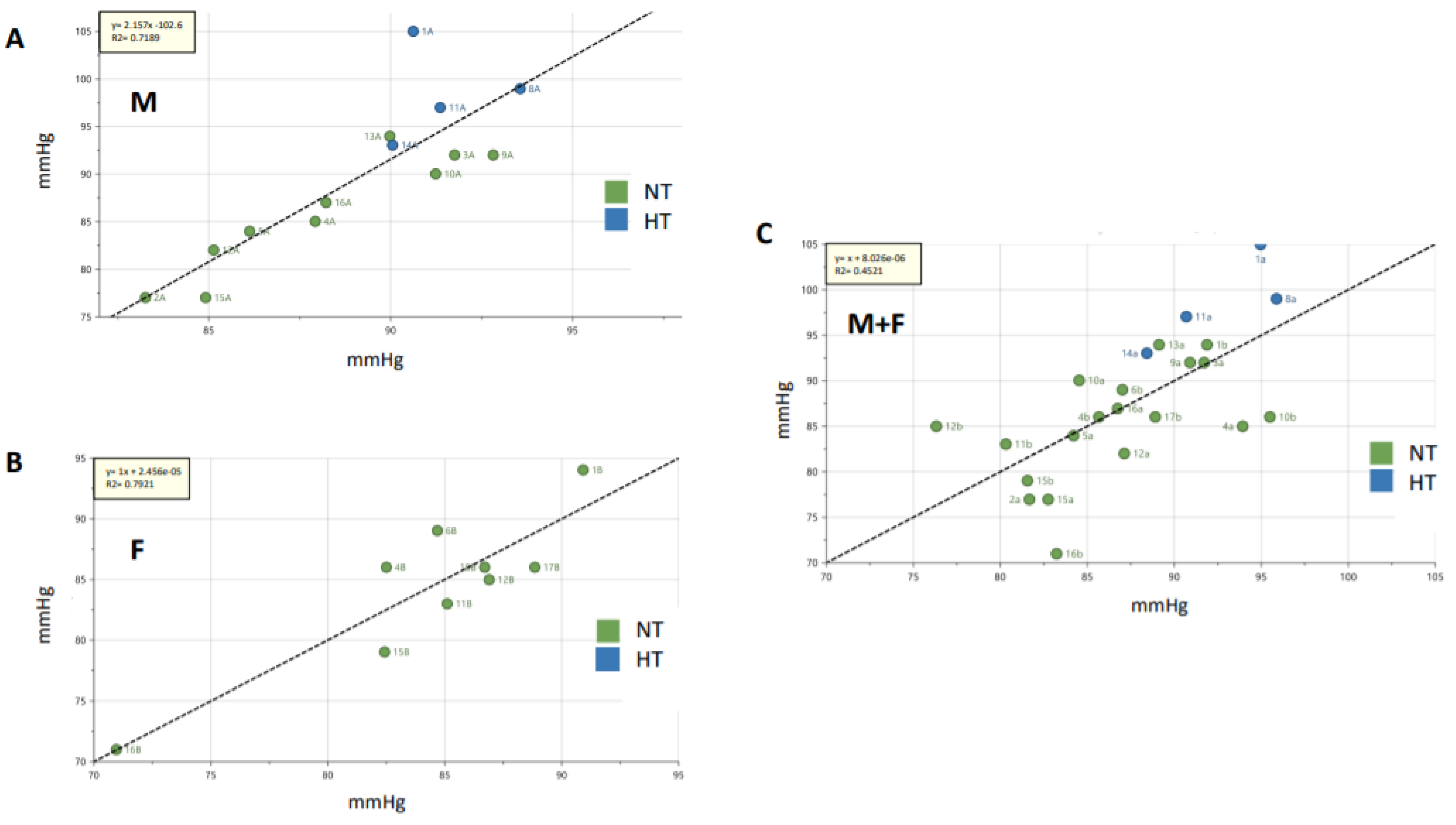
2. Results
2.1. Clinical Characteristics of the Three Cohorts
2.2. Anaerobic Culture of Fecal Bacteria
2.3. GM Characterization by 16S Amplicon Sequencing
2.4. Untargeted Metabolomics: Multivariate Analyses of the Entire Stool Metabolome
2.5. Metabolomics: Univariate Analyses of the Relative Quantification of the Three Main SCFA Levels
3. Discussion
4. Materials and Methods
- -
- Do you eat white or whole wheat bread, rice, and pasta? (White/Whole wheat/Both)
- -
- Do you eat yogurt? (Yes/Never)
- -
- Do you follow a vegetarian diet? (Yes/No)
- -
- Do you eat fruits and vegetables daily? (Yes/No)
- -
- What kind of fats do you eat? (Butter/Vegetable oil/Both)
- -
- Do you consume sugar or sweeteners? (Sugar/Sweetener/Both)
- -
- Do you use salt for cooking? (Yes/Never).
Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Vermali, R.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut Dysbiosis Is Linked to Hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Q.; Gu, Y.; Li, X.; Yang, W.; Jia, L.; Chen, C.; Han, X.; Huang, Y.; Zhao, L.; Li, P.; et al. Alterations of the Gut Microbiome in Hypertension. Front. Cell. Infect. Microbiol. 2017, 7, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Goel, R.; Kumar, A.; Qi, Y.; Lobaton, G.; Hosaka, K.; Mohammed, M.; Handberg, E.M.; Richards, E.M.; Pepine, C.J.; et al. Imbalance of gut microbiome and intestinal epithelial barrier dysfunction in patients with high blood pressure. Clin. Sci. 2018, 132, 701–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijers, B.; Jouret, F.; Evenepoel, P. Linking gut microbiota to cardiovascular disease and hypertension: Lessons from chronic kidney disease. Pharmacol. Res. 2018, 133, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Butel, M.-J. Probiotics, gut microbiota and health. Méd. Mal. Infect. 2014, 44, 1–8. [Google Scholar] [CrossRef]
- Alonso, V.R.; Guarner, F. Linking the gut microbiota to human health. Br. J. Nutr. 2013, 109, S21–S26. [Google Scholar] [CrossRef] [Green Version]
- Raizada, M.K.; Joe, B.; Bryan, N.S.; Chang, E.B.; Dewhirst, F.E.; Borisy, G.G.; Galis, Z.S.; Henderson, W.; Jose, P.A.; Ketchum, C.J.; et al. Report of the National Heart, Lung, and Blood Institute Working Group on the Role of Microbiota in Blood Pressure Regulation. Hypertension 2017, 70, 479–485. [Google Scholar] [CrossRef]
- Van Hul, M.; Geurts, L.; Plovier, H.; Druart, C.; Everard, A.; Ståhlman, M.; Rhimi, M.; Chira, K.; Teissedre, P.-L.; Delzenne, N.M.; et al. Reduced obesity, diabetes, and steatosis upon cinnamon and grape pomace are associated with changes in gut microbiota and markers of gut barrier. Am. J. Physiol. Metab. 2018, 314, E334–E352. [Google Scholar] [CrossRef]
- Duparc, T.; Plovier, H.; Marrachelli, V.G.; Van Hul, M.; Essaghir, A.; Ståhlman, M.; Matamoros, S.; Geurts, L.; Pardo-Tendero, M.M.; Druart, C.; et al. Hepatocyte MyD88 affects bile acids, gut microbiota and metabolome contributing to regulate glucose and lipid metabolism. Gut 2016, 66, 620–632. [Google Scholar] [CrossRef] [Green Version]
- Genest, J. Progress in Hypertension Research. Hypertension 2001, 38, E13–E18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzesinski, J.-M.; Saint-Remy, A. Essential hypertension, a complex trait. Rev. Med. Liege 2012, 67, 279–285. [Google Scholar]
- Davidson, M.B.; Hix, J.K.; Vidt, D.G.; Brotman, D.J. Association of Impaired Diurnal Blood Pressure Variation with a Subsequent Decline in Glomerular Filtration Rate. Arch. Intern. Med. 2006, 166, 846–852. [Google Scholar] [CrossRef] [Green Version]
- Zweiker, R.; Eber, B.; Schumacher, M.; Toplak, H.; Klein, W. “Non-dipping” related to cardiovascular events in essential hypertensive patients. Acta Med. Austriaca 1994, 21, 86–89. [Google Scholar]
- Bartolomaeus, H.; Markó, L.; Wilck, N.; Luft, F.C.; Forslund, S.K.; Muller, M.N. Precarious Symbiosis Between Host and Microbiome in Cardiovascular Health. Hypertension 2019, 73, 926–935. [Google Scholar] [CrossRef]
- Miyamoto, J.; Kasubuchi, M.; Nakajima, A.; Irie, J.; Itoh, H.; Kimura, I. The role of short-chain fatty acid on blood pressure regulation. Curr. Opin. Nephrol. Hypertens. 2016, 25, 379–383. [Google Scholar] [CrossRef] [PubMed]
- De La Cuesta-Zuluaga, J.; Mueller, N.T.; Álvarez-Quintero, R.; Velásquez-Mejía, E.P.; Sierra, J.A.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Higher Fecal Short-Chain Fatty Acid Levels Are Associated with Gut Microbiome Dysbiosis, Obesity, Hypertension and Cardiometabolic Disease Risk Factors. Nutrients 2018, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Pluznick, J.L. Microbial Short-Chain Fatty Acids and Blood Pressure Regulation. Curr. Hypertens. Rep. 2017, 19, 25. [Google Scholar] [CrossRef] [Green Version]
- Bugaut, M. Occurrence, absorption and metabolism of short chain fatty acids in the digestive tract of mammals. Comp. Biochem. Physiol. Part B Comp. Biochem. 1987, 86, 439–472. [Google Scholar] [CrossRef]
- Pluznick, J.L. Gut microbiota in renal physiology: Focus on short-chain fatty acids and their receptors. Kidney Int. 2016, 90, 1191–1198. [Google Scholar] [CrossRef] [Green Version]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; Reyes-Gavilán, C.G.D.L.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, N.E.; Kotarsky, K.; Owman, C.; Olde, B. Identification of a free fatty acid receptor, FFA2R, expressed on leukocytes and activated by short-chain fatty acids. Biochem. Biophys. Res. Commun. 2003, 303, 1047–1052. [Google Scholar] [CrossRef]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The Orphan G Protein-coupled Receptors GPR41 and GPR43 Are Activated by Propionate and Other Short Chain Carboxylic Acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef] [Green Version]
- Huart, J.; Leenders, J.; Taminiau, B.; Descy, J.; Saint-Remy, A.; Daube, G.; Krzesinski, J.-M.; Melin, P.; De Tullio, P.; Jouret, F. Gut Microbiota and Fecal Levels of Short-Chain Fatty Acids Differ Upon 24-Hour Blood Pressure Levels in Men. Hypertension 2019, 74, 1005–1013. [Google Scholar] [CrossRef]
- Sun, S.; Lulla, A.; Sioda, M.; Winglee, K.; Wu, M.C.; Jacobs, D.R.; Shikany, J.M.; Lloyd-Jones, D.M.; Launer, L.J.; Fodor, A.A.; et al. Gut Microbiota Composition and Blood Pressure. Hypertension 2019, 73, 998–1006. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhaar, B.J.H.; Collard, D.; Prodan, A.; Levels, J.H.M.; Zwinderman, A.H.; Bäckhed, F.; Vogt, L.; Peters, M.J.L.; Muller, M.; Nieuwdorp, M.; et al. Associations between gut microbiota, faecal short-chain fatty acids, and blood pressure across ethnic groups: The HELIUS study. Eur. Hear. J. 2020, 41, 4259–4267. [Google Scholar] [CrossRef] [PubMed]
- Bier, A.; Braun, T.; Khasbab, R.; Di Segni, A.; Grossman, E.; Haberman, Y.; Leibowitz, A. A High Salt Diet Modulates the Gut Microbiota and Short Chain Fatty Acids Production in a Salt-Sensitive Hypertension Rat Model. Nutrients 2018, 10, 1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, S.; Mandal, J.; Cheng, X.; Galla, S.; Hindupur, A.; Saha, P.; Yeoh, B.S.; Mell, B.; Yeo, J.-Y.; Vijay-Kumar, M.; et al. Diurnal Timing Dependent Alterations in Gut Microbial Composition Are Synchronously Linked to Salt-Sensitive Hypertension and Renal Damage. Hypertension 2020, 76, 59–72. [Google Scholar] [CrossRef]
- Huart, J.; Cirillo, A.; Saint-Remy, A.; Krzesinski, J.-M.; de Tullio, P.; Jouret, F. The faecal abundance of short chain fatty acids is increased in men with a non-dipping blood pressure profile. Acta Cardiol. 2021, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Hasan, A.U.; Nishiyama, A.; Kobori, H. Altered Circadian Timing System-Mediated Non-Dipping Pattern of Blood Pressure and Associated Cardiovascular Disorders in Metabolic and Kidney Diseases. Int. J. Mol. Sci. 2018, 19, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, R. Regulation of circadian blood pressure: From mice to astronauts. Curr. Opin. Nephrol. Hypertens. 2010, 19, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paschos, G.K.; FitzGerald, G.A. Circadian Clocks and Metabolism: Implications for Microbiome and Aging. Trends Genet. 2017, 33, 760–769. [Google Scholar] [CrossRef]
- Voigt, R.M.; Forsyth, C.B.; Green, S.J.; Mutlu, E.; Engen, P.; Vitaterna, M.H.; Turek, F.W.; Keshavarzian, A. Circadian Disorganization Alters Intestinal Microbiota. PLoS ONE 2014, 9, e97500. [Google Scholar] [CrossRef] [PubMed]
- Leone, V.; Gibbons, S.M.; Martinez, K.; Hutchison, A.L.; Huang, E.Y.; Cham, C.M.; Pierre, J.F.; Heneghan, A.F.; Nadimpalli, A.; Hubert, N.; et al. Effects of Diurnal Variation of Gut Microbes and High-Fat Feeding on Host Circadian Clock Function and Metabolism. Cell Host Microbe 2015, 17, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Mukherji, A.; Kobiita, A.; Ye, T.; Chambon, P. Homeostasis in Intestinal Epithelium Is Orchestrated by the Circadian Clock and Microbiota Cues Transduced by TLRs. Cell 2013, 153, 812–827. [Google Scholar] [CrossRef] [Green Version]
- Thaiss, C.A.; Zeevi, D.; Levy, M.; Zilberman-Schapira, G.; Suez, J.; Tengeler, A.C.; Abramson, L.; Katz, M.N.; Korem, T.; Zmora, N.; et al. Transkingdom Control of Microbiota Diurnal Oscillations Promotes Metabolic Homeostasis. Cell 2014, 159, 514–529. [Google Scholar] [CrossRef] [Green Version]
- Pickering, T.G.; Kario, K. Nocturnal non-dipping: What does it augur? Curr. Opin. Nephrol. Hypertens. 2001, 10, 611–616. [Google Scholar] [CrossRef]
- Pioli, M.R.; Ritter, A.M.V.; De Faria, A.P.; Modolo, R. White coat syndrome and its variations: Differences and clinical impact. Integr. Blood Press. Control 2018, 11, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Bo, Y.; Kwok, K.-O.; Chung, V.C.-H.; Yu, C.-P.; Tsoi, K.K.-F.; Wong, S.Y.-S.; Lee, E.K.-P. Short-term reproducibility of ambulatory blood pressure measurements: A systematic review and meta-analysis of 35 observational studies. J. Hypertens. 2020, 38, 2095–2109. [Google Scholar] [CrossRef]
- Haug, K.; Cochrane, K.; Nainala, V.C.; Williams, M.; Chang, J.; Jayaseelan, K.V.; O’Donovan, C. MetaboLights: A resource evolving in response to the needs of its scientific community. Nucleic Acids Res. 2019, 48, D440–D444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancia, G.; Fagard, R.; Narkiewicz, K.; Redon, J.; Zanchetti, A.; Böhm, M.; Christiaens, T.; Cifkova, R.; De Backer, G.; Dominiczak, A.; et al. 2013 ESH/ESC Guidelines for the management of arterial hypertension: The task force for the management of arterial hypertension of the European Society of Hypertension (ESH) and of the European Society of Cardiology (ESC). Eur. Heart J. 2013, 34, 2159–2219. [Google Scholar]
- Fagard, R.H. Dipping pattern of nocturnal blood pressure in patients with hypertension. Expert Rev. Cardiovasc. Ther. 2009, 7, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Cardona, S.; Eck, A.; Cassellas, M.; Gallart, M.; Alastrue, C.; Dore, J.; Azpiroz, F.; Roca, J.; Guarner, F.; Manichanh, C. Storage conditions of intestinal microbiota matter in metagenomic analysis. BMC Microbiol. 2012, 12, 158. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, C.; Taminiau, B.; Korsak, N.; Avesani, V.; Van Broeck, J.; Brach, P.; Delmée, M.; Daube, G. Longitudinal survey of Clostridium difficile presence and gut microbiota composition in a Belgian nursing home. BMC Microbiol. 2016, 16, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Martin, A.P. Phylogenetic Approaches for Describing and Comparing the Diversity of Microbial Communities. Appl. Environ. Microbiol. 2002, 68, 3673–3682. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2020 Male and Female Cohorts | Normotension | Hypertension | p Value |
|---|---|---|---|
| N | 19 | 7 | |
| Age (years) | 49.2 ± 13.7 | 52.5 ± 10.5 | 0.469 |
| Female (N) | 9 | 1 | |
| BMI (kg/m2) | 23.3 ± 2.5 | 25 ± 2.9 | 0.132 |
| Smokers (%, N) | 5% (1) | 14.3% (1) | 0.949 |
| Alcohol (glass/week) | 3.7 ± 6 | 6 ± 7 | 0.193 |
| Family HT (%, N) | 37% (7) | 57% (4) | 0.629 |
| Diabetes (%, N) | 0 | 14.3% (1) | 0.595 |
| CV history (%, N) | 16% (3) | 29% (2) | 0.862 |
| GE history (%, N) | 36.8 (7) | 14.3 (1) | 0.531 |
| 24 h Systolic BP (mmHg) | 113 ± 7 | 127 ± 4 | 0.0005 |
| 24 h Diastolic BP (mmHg) | 70 ± 6 | 81 ± 4 | 0.0004 |
| 24 h Mean BP (mmHg) | 82 ± 6 | 98 ± 5 | 0.0005 |
| Anti-HT treatment (%, N) | 0 | 28.6% (2) | ns |
| Non-dippers (%, N) | 58% (11) | 57% (4) | 0.679 |
| 2020 Male and Female Cohorts | Dippers | Non-Dippers | p Value |
|---|---|---|---|
| N | 11 | 13 | |
| Age (years) | 48.8 ± 11.4 | 51.9 ± 14.6 | 0.643 |
| Female (N) | 3 | 6 | |
| BMI (kg/m2) | 23.6 ± 2.5 | 23.5 ± 3 | 0.907 |
| Smokers (%, N) | 9% (1) | 0 | ns |
| Alcohol (glass/week) | 6 ± 8 | 3.4 ± 4.8 | 0.417 |
| Family HT (%, N) | 63.6% (7) | 23% (3) | 0.111 |
| Diabetes (%, N) | 0 | 0 | ns |
| CV history (%, N) | 0 | 23% (3) | 0.278 |
| GE history (%, N) | 36.4% (4) | 30.8% (4) | 0.884 |
| 24 h Systolic BP (mmHg) | 119 ± 9 | 114 ± 9 | 0.131 |
| 24 h Diastolic BP (mmHg) | 74 ± 7 | 71 ± 8 | 0.417 |
| 24 h Mean BP (mmHg) | 90 ± 8 | 87 ± 9 | 0.602 |
| ND Systolic BP ratio | 0.85 ± 0.03 | 0.93 ± 0.03 | <0.0001 |
| All Patients | 2015 Male Cohort | 2020 Male Cohort | p Value |
|---|---|---|---|
| N | 16 | 16 | |
| Age (years) | 47.6 ± 12.2 | 52.3 ± 12.4 | 0.0004 |
| BMI (kg/m2) | 23.8 ± 2.4 | 24.6 ± 2.6 | 0.214 |
| Smokers (%, N) | 12.5 (2) | 6.2 (1) | ns |
| Alcohol (glass/week) | 4.8 ± 3.5 | 6.4 ± 7 | 0.272 |
| Family HT (%, N) | 50 (8) | 50 (8) | 0.479 |
| Diabetes (%, N) | 0 | 6.2 (1) | ns |
| CV history (%, N) | 12.5 (2) | 18.7 (3) | ns |
| GE history (%, N) | 31.2 (5) | 31.2 (5) | ns |
| Anti-HT treatment (%, N) | 6.2 (1) | 6.2 (1) | ns |
| Non-dippers (%, N) | 31.2 (5) | 50 (8) | 0.449 |
| Patients with Change in BP Status | 2015 Male Cohort | 2020 Male Cohort | p Value |
| N | 6 | 6 | |
| Age (years) | 42.6 ± 13.6 | 47.1 ± 13.2 | 0.027 |
| BMI (kg/m2) | 22.6 ± 1.7 | 23.5 ± 1.7 | 0.345 |
| Smokers (% N) | 33 (2) | 16.6 (1) | ns |
| Alcohol (glass/week) | 4.7 ± 1.4 | 7.3 ± 7 | 0.345 |
| Family HT (%, N) | 66 (4) | 66 (4) | ns |
| Diabetes (%) | 0 | 0 | ns |
| CV history (%) | 0 | 0 | ns |
| GE history (%, N) | 16.6 (1) | 16.6 (1) | ns |
| Anti-HT treatment (%) | 0 | 0 | ns |
| Non-dippers (%, N) | 50 (3) | 33 (2) | ns |
| (A) NT vs. HT | (B) Dippers vs. Non-Dippers | ||||
|---|---|---|---|---|---|
| X-Variable in ppm | VIP | Feature ID | X-Variable in ppm | VIP | Feature ID |
| 0.91 | 1.51 | Butyrate | 0.91 | 1.1 | Butyrate |
| 3.39 | 1.48 | matrix | 3.84 | 1.1 | matrix |
| 0.89 | 1.47 | Butyrate | 3.94 | 1.09 | matrix |
| 2.99 | 1.43 | matrix | 2.09 | 1.09 | matrix |
| 1.55 | 1.43 | Butyrate | 2.14 | 1.09 | Propionate |
| 2.14 | 1.39 | Propionate | 1.5 | 1.08 | Butyrate |
| 3.94 | 1.38 | matrix | 0.89 | 1.08 | Butyrate |
| 4.19 | 1.29 | matrix | 1.04 | 1.08 | Propionate |
| 0.54 | 1.25 | matrix | 2.04 | 1.08 | matrix |
| 0.64 | 1.24 | matrix | 1.06 | 1.08 | Propionate |
| 1.03 | 1.24 | Propionate | 1.93 | 1.06 | Acetate |
| 1.93 | 1.2 | Acetate | 1.74 | 1.06 | matrix |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huart, J.; Cirillo, A.; Taminiau, B.; Descy, J.; Saint-Remy, A.; Daube, G.; Krzesinski, J.-M.; Melin, P.; de Tullio, P.; Jouret, F. Human Stool Metabolome Differs upon 24 h Blood Pressure Levels and Blood Pressure Dipping Status: A Prospective Longitudinal Study. Metabolites 2021, 11, 282. https://doi.org/10.3390/metabo11050282
Huart J, Cirillo A, Taminiau B, Descy J, Saint-Remy A, Daube G, Krzesinski J-M, Melin P, de Tullio P, Jouret F. Human Stool Metabolome Differs upon 24 h Blood Pressure Levels and Blood Pressure Dipping Status: A Prospective Longitudinal Study. Metabolites. 2021; 11(5):282. https://doi.org/10.3390/metabo11050282
Chicago/Turabian StyleHuart, Justine, Arianna Cirillo, Bernard Taminiau, Julie Descy, Annie Saint-Remy, Georges Daube, Jean-Marie Krzesinski, Pierrette Melin, Pascal de Tullio, and François Jouret. 2021. "Human Stool Metabolome Differs upon 24 h Blood Pressure Levels and Blood Pressure Dipping Status: A Prospective Longitudinal Study" Metabolites 11, no. 5: 282. https://doi.org/10.3390/metabo11050282