
Roles of Estrogens in the Healthy and Diseased Oviparous Vertebrate Liver

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Liver Sexual Dimorphism and Hepatic Sexually Dimorphic Gene Expression
3. Crosstalk between Liver and Ovary in the Production of Egg Yolk and Eggshell Proteins
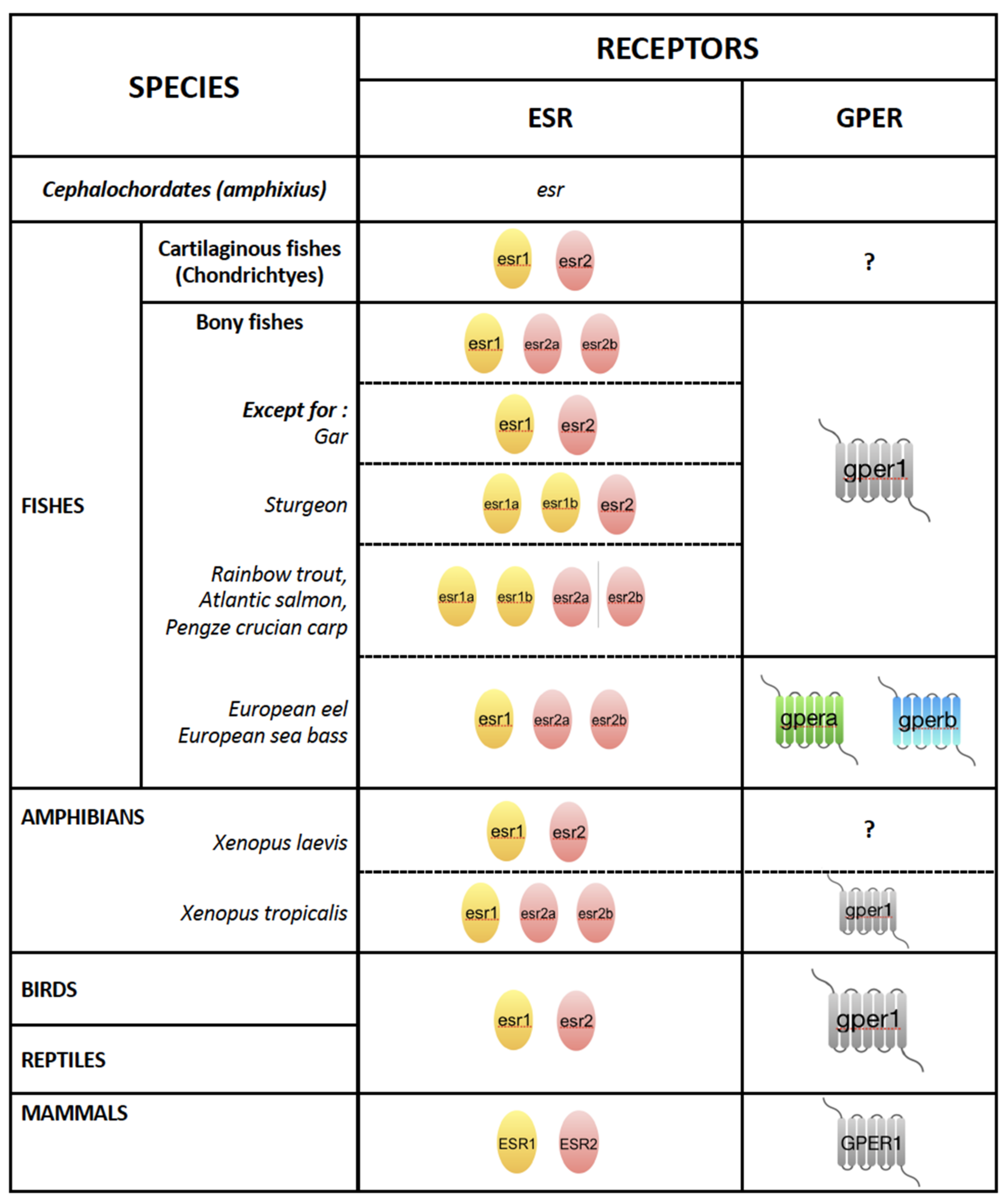
4. Estrogen Receptors in Oviparous Vertebrates and Their Hepatic Expression
5. Estrogen-Dependent Lipid and Lipoprotein Metabolisms in the Oviparous Liver
6. Hepatic Crosstalk between ESRS and Other Members of the Nuclear Receptor Superfamily
7. The Oviparous Vertebrate Liver as a Sensor of Estrogen Disrupting Compounds
8. Estrogen Associated Liver Diseases in Oviparous Vertebrates
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kietzmann, T. Liver Zonation in Health and Disease: Hypoxia and Hypoxia-Inducible Transcription Factors as Concert Masters. Int. J. Mol. Sci. 2019, 20, 2347. [Google Scholar] [CrossRef] [Green Version]
- Pastorino, P.; Prearo, M.; Pizzul, E.; Bertoli, M.; Francese, D.R.; Menconi, V.; Mugetti, D.; Bozzetta, E.; Varello, K. Hepatic Steatosis in a Bullhead (Cottus Gobio) Population from a High-Mountain Lake (Carnic Alps): Adaptation to an Extreme Ecosystem? Water 2019, 11, 2570. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Swarup, V.; Le, H.; Kumar, V. Transcriptional Signatures in Liver Reveal Metabolic Adaptations to Seasons in Migratory Blackheaded Buntings. Front. Physiol. 2018, 9, 1568. [Google Scholar] [CrossRef]
- Zhu, W.; Zhang, M.; Chang, L.; Zhu, W.; Li, C.; Xie, F.; Zhang, H.; Zhao, T.; Jiang, J. Characterizing the Composition, Metabolism and Physiological Functions of the Fatty Liver in Rana Omeimontis Tadpoles. Front. Zool. 2019, 16, 42. [Google Scholar] [CrossRef] [PubMed]
- Kietzmann, T. Metabolic Zonation of the Liver: The Oxygen Gradient Revisited. Redox Biol. 2017, 11, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Ben-Moshe, S.; Itzkovitz, S. Spatial Heterogeneity in the Mammalian Liver. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 395–410. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Martínez-Ramírez, A.S.; Borders, T.L.; Gao, F.; Sosa-Pineda, B. Metabolic and Non-Metabolic Liver Zonation Is Established Non-Synchronously and Requires Sinusoidal Wnts. eLife 2020, 9, e46206. [Google Scholar] [CrossRef] [PubMed]
- Jungermann, K.; Keitzmann, T. Zonation of Parenchymal and Nonparenchymal Metabolism in Liver. Annu. Rev. Nutr. 1996, 16, 179–203. [Google Scholar] [CrossRef]
- He, L.; Pu, W.; Liu, X.; Zhang, Z.; Han, M.; Li, Y.; Huang, X.; Han, X.; Li, Y.; Liu, K.; et al. Proliferation Tracing Reveals Regional Hepatocyte Generation in Liver Homeostasis and Repair. Science 2021, 371, eabc4346. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, Y.G.; Jia, Y.; Li, L.; Yoon, J.; Zhang, S.; Wang, Z.; Zhang, Y.; Zhu, M.; Sharma, T.; et al. Liver Homeostasis Is Maintained by Midlobular Zone 2 Hepatocytes. Science 2021, 371, eabb1625. [Google Scholar] [CrossRef] [PubMed]
- Desvergne, B.; Michalik, L.; Wahli, W. Transcriptional Regulation of Metabolism. Physiol. Rev. 2006, 86, 465–514. [Google Scholar] [CrossRef]
- Tao, L.J.; Seo, D.E.; Jackson, B.; Ivanova, N.B.; Santori, F.R. Nuclear Hormone Receptors and Their Ligands: Metabolites in Control of Transcription. Cells 2020, 9, 2606. [Google Scholar] [CrossRef] [PubMed]
- Brunmeir, R.; Xu, F. Functional Regulation of PPARs through Post-Translational Modifications. Int. J. Mol. Sci. 2018, 19, 1738. [Google Scholar] [CrossRef] [Green Version]
- Leuenberger, N.; Pradervand, S.; Wahli, W. Sumoylated PPARα Mediates Sex-Specific Gene Repression and Protects the Liver from Estrogen-Induced Toxicity in Mice. J. Clin. Investig. 2009, 119, 3138–3148. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z. Genomic Analysis of the Nuclear Receptor Family: New Insights Into Structure, Regulation, and Evolution From the Rat Genome. Genome Res. 2004, 14, 580–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lösel, R.; Wehling, M. Nongenomic Actions of Steroid Hormones. Nat. Rev. Mol. Cell Biol. 2003, 4, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schütz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The Nuclear Receptor Superfamily: The Second Decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Cai, S.-Y.; Boyer, J.L. The Role of the Retinoid Receptor, RAR/RXR Heterodimer, in Liver Physiology. Biochim. Biophys. Acta 2021, 1867, 166085. [Google Scholar] [CrossRef] [PubMed]
- Russo-Savage, L.; Schulman, I.G. Liver X Receptors and Liver Physiology. Biochim. Biophys. Acta 2021, 1867, 166121. [Google Scholar] [CrossRef]
- Cai, X.; Young, G.M.; Xie, W. The Xenobiotic Receptors PXR and CAR in Liver Physiology, an Update. Biochim. Biophys. Acta 2021, 1867, 166101. [Google Scholar] [CrossRef]
- Panzitt, K.; Wagner, M. FXR in Liver Physiology: Multiple Faces to Regulate Liver Metabolism. Biochim. Biophys. Acta 2021, 1867, 166133. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Kim, M.; Lazar, M.A. Nuclear Receptors and Transcriptional Regulation in Non-Alcoholic Fatty Liver Disease. Mol. Metab. 2020, 101119. [Google Scholar] [CrossRef] [PubMed]
- Fougerat, A.; Montagner, A.; Loiseau, N.; Guillou, H.; Wahli, W. Peroxisome Proliferator-Activated Receptors and Their Novel Ligands as Candidates for the Treatment of Non-Alcoholic Fatty Liver Disease. Cells 2020, 9, 1638. [Google Scholar] [CrossRef]
- Dixon, E.D.; Nardo, A.D.; Claudel, T.; Trauner, M. The Role of Lipid Sensing Nuclear Receptors (PPARs and LXR) and Metabolic Lipases in Obesity, Diabetes and NAFLD. Genes 2021, 12, 645. [Google Scholar] [CrossRef]
- Berthier, A.; Johanns, M.; Zummo, F.P.; Lefebvre, P.; Staels, B. PPARs in Liver Physiology. Biochim. Biophys. Acta. Mol. Basis Dis. 2021, 1867, 166097. [Google Scholar] [CrossRef]
- Lodé, T. Oviparity or Viviparity? That Is the Question…. Reprod. Biol. 2012, 12, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Wahli, W.; Dawid, I.; Ryffel, G.; Weber, R. Vitellogenesis and the Vitellogenin Gene Family. Science 1981, 212, 298–304. [Google Scholar] [CrossRef]
- Wahli, W. Evolution and Expression of Vitellogenin Genes. Trends Genet. 1988, 4, 227–232. [Google Scholar] [CrossRef]
- Litscher, E.S.; Wassarman, P.M. Egg Extracellular Coat Proteins: From Fish to Mammals. Histol. Histopathol. 2007, 22, 337–347. [Google Scholar] [CrossRef]
- Kim, K.H.; Moore, D.D. Regulation of Liver Energy Balance by the Nuclear Receptors Farnesoid X Receptor and Peroxisome Proliferator Activated Receptor α. Dig. Dis. 2017, 35, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.K.; Chatterjee, B. Sexual Dimorphism in the Liver. Annu. Rev. Physiol. 1983, 45, 37–50. [Google Scholar] [CrossRef]
- Arukwe, A.; Goksøyr, A. Eggshell and Egg Yolk Proteins in Fish: Hepatic Proteins for the next Generation: Oogenetic, Population, and Evolutionary Implications of Endocrine Disruption. Comp. Hepatol. 2003, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Rando, G.; Wahli, W. Sex Differences in Nuclear Receptor-Regulated Liver Metabolic Pathways. Biochim. Biophys. Acta 2011, 1812, 964–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maggi, A.; Della Torre, S. Sex, Metabolism and Health. Mol. Metab. 2018, 15, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Brie, B.; Ramirez, M.C.; De Winne, C.; Lopez Vicchi, F.; Villarruel, L.; Sorianello, E.; Catalano, P.; Ornstein, A.M.; Becu-Villalobos, D. Brain Control of Sexually Dimorphic Liver Function and Disease: The Endocrine Connection. Cell. Mol. Neurobiol. 2019, 39, 169–180. [Google Scholar] [CrossRef] [PubMed]
- De Castro Barbosa, T.; Salgueiro, R.B.; Serrano-Nascimento, C.; Amaral, F.G.; Cipolla-Neto, J.; Nunes, M.T. Molecular Basis of Growth Hormone Daily MRNA and Protein Synthesis in Rats. Life Sci. 2018, 207, 36–41. [Google Scholar] [CrossRef]
- Toews, J.N.C.; Hammond, G.L.; Viau, V. Liver at the Nexus of Rat Postnatal HPA Axis Maturation and Sexual Dimorphism. J. Endocrinol. 2021, 248, R1–R17. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.M.; Otero-Corchon, V.; Hammond, G.L.; Veldhuis, J.D.; Qi, N.; Low, M.J. Somatostatin Is Essential for the Sexual Dimorphism of GH Secretion, Corticosteroid-Binding Globulin Production, and Corticosterone Levels in Mice. Endocrinology 2015, 156, 1052–1065. [Google Scholar] [CrossRef] [Green Version]
- Jansson, J.O.; Edén, S.; Isaksson, O. Sexual Dimorphism in the Control of Growth Hormone Secretion. Endocr. Rev. 1985, 6, 128–150. [Google Scholar] [CrossRef]
- Viau, V. Functional Cross-Talk between the Hypothalamic-Pituitary-Gonadal and -Adrenal Axes. J. Neuroendocr. 2002, 14, 506–513. [Google Scholar] [CrossRef]
- Schlegel, A.; Gut, P. Metabolic Insights from Zebrafish Genetics, Physiology, and Chemical Biology. Cell. Mol. Life Sci. 2015, 72, 2249–2260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, K.; Takami, T.; Kimoto, Y.; Matsumoto, T.; Yamamoto, N.; Terai, S.; Sakaida, I. Circadian Variations in the Liver Metabolites of Medaka (Oryzias Latipes). Sci. Rep. 2016, 6, 20916. [Google Scholar] [CrossRef] [Green Version]
- Udy, G.B.; Towers, R.P.; Snell, R.G.; Wilkins, R.J.; Park, S.H.; Ram, P.A.; Waxman, D.J.; Davey, H.W. Requirement of STAT5b for Sexual Dimorphism of Body Growth Rates and Liver Gene Expression. Proc. Natl. Acad. Sci. USA 1997, 94, 7239–7244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, S.; Mei, J.; Huang, P.; Jing, J.; Li, Z.; Kang, J.; Gui, J.-F. Essential Roles of Stat5.1/Stat5b in Controlling Fish Somatic Growth. J. Genet. Genom. 2017, 44, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Xiong, S.; Kang, J.; Mei, J.; Gui, J.-F. Stat5b Regulates Sexually Dimorphic Gene Expression in Zebrafish Liver. Front. Physiol. 2018, 9, 676. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.H.; Wang, J.Y.; Zhang, G.X.; Wei, Y.; Gu, Y.P.; Yu, Y.B. Single Nucleotide Polymorphism in the STAT5b Gene Is Associated with Body Weight and Reproductive Traits of the Jinghai Yellow Chicken. Mol. Biol. Rep. 2012, 39, 4177–4183. [Google Scholar] [CrossRef]
- Davis, L.K.; Pierce, A.L.; Hiramatsu, N.; Sullivan, C.V.; Hirano, T.; Grau, E.G. Gender-Specific Expression of Multiple Estrogen Receptors, Growth Hormone Receptors, Insulin-like Growth Factors and Vitellogenins, and Effects of 17 Beta-Estradiol in the Male Tilapia (Oreochromis Mossambicus). Gen. Comp. Endocrinol. 2008, 156, 544–551. [Google Scholar] [CrossRef]
- Qiao, Q.; Le Manach, S.; Sotton, B.; Huet, H.; Duvernois-Berthet, E.; Paris, A.; Duval, C.; Ponger, L.; Marie, A.; Blond, A.; et al. Deep Sexual Dimorphism in Adult Medaka Fish Liver Highlighted by Multi-Omic Approach. Sci. Rep. 2016, 6, 32459. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, B.J.G.; Prokkola, J.M.; Audet, C.; Bernatchez, L. Sex-Specific Co-Expression Networks and Sex-Biased Gene Expression in the Salmonid Brook Charr Salvelinus Fontinalis. G3 2019, 9, 955–968. [Google Scholar] [CrossRef] [Green Version]
- Saelens, W.; Cannoodt, R.; Saeys, Y. A Comprehensive Evaluation of Module Detection Methods for Gene Expression Data. Nat. Commun. 2018, 9, 1090. [Google Scholar] [CrossRef]
- Jin, L.; Yu, J.P.; Yang, Z.J.; Merilä, J.; Liao, W.B. Modulation of Gene Expression in Liver of Hibernating Asiatic Toads (Bufo Gargarizans). Int. J. Mol. Sci. 2018, 19, 2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Park, J.Y.; Yoo, W.; Gwag, T.; Lee, J.-W.; Byun, M.-W.; Choi, I. Overcoming Muscle Atrophy in a Hibernating Mammal despite Prolonged Disuse in Dormancy: Proteomic and Molecular Assessment. J. Cell. Biochem. 2008, 104, 642–656. [Google Scholar] [CrossRef]
- Cox, R.M.; Cox, C.L.; McGlothlin, J.W.; Card, D.C.; Andrew, A.L.; Castoe, T.A. Hormonally Mediated Increases in Sex-Biased Gene Expression Accompany the Breakdown of between-Sex Genetic Correlations in a Sexually Dimorphic Lizard. Am. Nat. 2017, 189, 315–332. [Google Scholar] [CrossRef] [Green Version]
- Mank, J.E. The Transcriptional Architecture of Phenotypic Dimorphism. Nat. Ecol. Evol. 2017, 1, 0006. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.N. Vertebrate Yolk Complexes and the Functional Implications of Phosvitins and Other Subdomains in Vitellogenins1. Biol. Reprod. 2007, 76, 926–935. [Google Scholar] [CrossRef] [Green Version]
- Buisine, N.; Trichet, V.; Wolff, J. Complex Evolution of Vitellogenin Genes in Salmonid Fishes. Mol. Gen. Genom. 2002, 268, 535–542. [Google Scholar] [CrossRef]
- Jaggi, R.B.; Wyler, T.; Ryffel, G.U. Comparative Analysis of Xenopus Tropicalis and Xenopus Laevis Vitellogenin Gene Sequences. Nucleic Acids Res. 1982, 10, 1515–1533. [Google Scholar] [CrossRef] [Green Version]
- Brawand, D.; Wahli, W.; Kaessmann, H. Loss of Egg Yolk Genes in Mammals and the Origin of Lactation and Placentation. PLoS Biol. 2008, 6, e63. [Google Scholar] [CrossRef] [PubMed]
- Litscher, E.S.; Wassarman, P.M. Evolution, Structure, and Synthesis of Vertebrate Egg-Coat Proteins. Trends Dev. Biol. 2014, 8, 65–76. [Google Scholar] [PubMed]
- Wahli, W.; Wyler, T.; Weber, R.; Ryffel, G.U. Size, Complexity and Abundance of a Specific Poly(A)-Containing RNA of Liver from Male Xenopus Induced to Vitellogenin Synthesis by Estrogen. Eur. J. Biochem. 1976, 66, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Ryffel, G.U.; Wahli, W.; Weber, R. Quantitation of Vitellogenin Messenger RNA in the Liver of Male Xenopus Toads during Primary and Secondary Stimulation by Estrogen. Cell 1977, 11, 213–221. [Google Scholar] [CrossRef]
- Corthésy, B.; Hipskind, R.; Theulaz, I.; Wahli, W. Estrogen-Dependent in Vitro Transcription from the Vitellogenin Promoter in Liver Nuclear Extracts. Science 1988, 239, 1137–1139. [Google Scholar] [CrossRef]
- Corthésy, B.; Claret, F.X.; Wahli, W. Estrogen Receptor Level Determines Sex-Specific in Vitro Transcription from the Xenopus Vitellogenin Promoter. Proc. Natl. Acad. Sci. USA 1990, 87, 7878–7882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verderame, M.; Limatola, E. Molecular Identification of Estrogen Receptors (ERα and ERβ) and Their Differential Expression during VTG Synthesis in the Liver of Lizard Podarcis Sicula. Gen. Comp. Endocrinol. 2010, 168, 231–238. [Google Scholar] [CrossRef]
- Baker, M.E. Steroid Receptors and Vertebrate Evolution. Mol. Cell. Endocrinol. 2019, 496, 110526. [Google Scholar] [CrossRef] [Green Version]
- Bondesson, M.; Hao, R.; Lin, C.-Y.; Williams, C.; Gustafsson, J.-Å. Estrogen Receptor Signaling during Vertebrate Development. Biochim. Biophys. Acta 2015, 1849, 142–151. [Google Scholar] [CrossRef] [Green Version]
- Lecroisey, C.; Laudet, V.; Schubert, M. The Cephalochordate Amphioxus: A Key to Reveal the Secrets of Nuclear Receptor Evolution. Brief. Funct. Genom. 2012, 11, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Ogino, Y.; Tohyama, S.; Kohno, S.; Toyota, K.; Yamada, G.; Yatsu, R.; Kobayashi, T.; Tatarazako, N.; Sato, T.; Matsubara, H.; et al. Functional Distinctions Associated with the Diversity of Sex Steroid Hormone Receptors ESR and AR. J. Steroid Biochem. Mol. Biol. 2018, 184, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Cabas, I.; Chaves-Pozo, E.; Mulero, V.; García-Ayala, A. Role of Estrogens in Fish Immunity with Special Emphasis on GPER1. Dev. Comp. Immunol. 2018, 89, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Schaaf, M.J.M. Nuclear Receptor Research in Zebrafish. J. Mol. Endocrinol. 2017, 59, R65–R76. [Google Scholar] [CrossRef] [Green Version]
- Coumailleau, P.; Pellegrini, E.; Adrio, F.; Diotel, N.; Cano-Nicolau, J.; Nasri, A.; Vaillant, C.; Kah, O. Aromatase, Estrogen Receptors and Brain Development in Fish and Amphibians. Biochim. Biophys. Acta 2015, 1849, 152–162. [Google Scholar] [CrossRef]
- Menuet, A.; Pellegrini, E.; Anglade, I.; Blaise, O.; Laudet, V.; Kah, O.; Pakdel, F. Molecular Characterization of Three Estrogen Receptor Forms in Zebrafish: Binding Characteristics, Transactivation Properties, and Tissue Distributions1. Biol. Reprod. 2002, 66, 1881–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takase, M.; Iguchi, T. Molecular Cloning of Two Isoforms of Xenopus (Silurana) Tropicalis Estrogen Receptor MRNA and Their Expression during Development. Biochim. Biophys. Acta 2007, 1769, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Morini, M.; Peñaranda, D.S.; Vílchez, M.C.; Nourizadeh-Lillabadi, R.; Lafont, A.-G.; Dufour, S.; Asturiano, J.F.; Weltzien, F.-A.; Pérez, L. Nuclear and Membrane Progestin Receptors in the European Eel: Characterization and Expression in Vivo through Spermatogenesis. Comp. Biochem. Physiol. Pert A 2017, 207, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Pakdel, F.; Féon, S.; Le Gac, F.; Le Menn, F.; Valotaire, Y. In Vivo Estrogen Induction of Hepatic Estrogen Receptor MRNA and Correlation with Vitellogenin MRNA in Rainbow Trout. Mol. Cell. Endocrinol. 1991, 75, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Mushirobira, Y.; Niida, M.; Hotta, T.; Fujinami, Y.; Soyano, K. Hepatic Expression Profiles of Three Subtypes of Vitellogenin and Estrogen Receptor during Vitellogenesis in Cultured Female Yellowtail. Gen. Comp. Endocrinol. 2020, 299, 113612. [Google Scholar] [CrossRef]
- Le Dréan, Y.; Lazennec, G.; Kern, L.; Saligaut, D.; Pakdel, F.; Valotaire, Y. Characterization of an Estrogen-Responsive Element Implicated in Regulation of the Rainbow Trout Estrogen Receptor Gene. J. Mol. Endocrinol. 1995, 15, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Menuet, A.; Le Page, Y.; Torres, O.; Kern, L.; Kah, O.; Pakdel, F. Analysis of the Estrogen Regulation of the Zebrafish Estrogen Receptor (ER) Reveals Distinct Effects of ERalpha, ERbeta1 and ERbeta2. J. Mol. Endocrinol. 2004, 32, 975–986. [Google Scholar] [CrossRef] [Green Version]
- Boyce-Derricott, J.; Nagler, J.J.; Cloud, J.G. Variation among Rainbow Trout (Oncorhynchus Mykiss) Estrogen Receptor Isoform 3’ Untranslated Regions and the Effect of 17beta-Estradiol on MRNA Stability in Hepatocyte Culture. DNA Cell Biol. 2010, 29, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Lassiter, C.S.; Kelley, B.; Linney, E. Genomic Structure and Embryonic Expression of Estrogen Receptor Beta a (ERbetaa) in Zebrafish (Danio Rerio). Gene 2002, 299, 141–151. [Google Scholar] [CrossRef]
- Nelson, E.R.; Habibi, H.R. Estrogen Receptor Function and Regulation in Fish and Other Vertebrates. Gen. Comp. Endocrinol. 2013, 192, 15–24. [Google Scholar] [CrossRef]
- MacKay, M.E.; Raelson, J.; Lazier, C.B. Up-Regulation of Estrogen Receptor MRNA and Estrogen Receptor Activity by Estradiol in Liver of Rainbow Trout and Other Teleostean Fish. Comp. Biochem. Physiol. Part C 1996, 115, 201–209. [Google Scholar] [CrossRef]
- Marlatt, V.L.; Martyniuk, C.J.; Zhang, D.; Xiong, H.; Watt, J.; Xia, X.; Moon, T.; Trudeau, V.L. Auto-Regulation of Estrogen Receptor Subtypes and Gene Expression Profiling of 17beta-Estradiol Action in the Neuroendocrine Axis of Male Goldfish. Mol. Cell. Endocrinol. 2008, 283, 38–48. [Google Scholar] [CrossRef]
- Marlatt, V.L.; Lakoff, J.; Crump, K.; Martyniuk, C.J.; Watt, J.; Jewell, L.; Atkinson, S.; Blais, J.M.; Sherry, J.; Moon, T.W.; et al. Sex- and Tissue-Specific Effects of Waterborne Estrogen on Estrogen Receptor Subtypes and E2-Mediated Gene Expression in the Reproductive Axis of Goldfish. Comp. Biochem. Physiol. Part A 2010, 156, 92–101. [Google Scholar] [CrossRef]
- Dahlman-Wright, K.; Cavailles, V.; Fuqua, S.A.; Jordan, V.C.; Katzenellenbogen, J.A.; Korach, K.S.; Maggi, A.; Muramatsu, M.; Parker, M.G.; Gustafsson, J.-A. International Union of Pharmacology. LXIV. Estrogen Receptors. Pharm. Rev. 2006, 58, 773–781. [Google Scholar] [CrossRef] [Green Version]
- Walker, P.; Germond, J.E.; Brown-Luedi, M.; Givel, F.; Wahli, W. Sequence Homologies in the Region Preceding the Transcription Initiation Site of the Liver Estrogen-Responsive Vitellogenin and Apo-VLDLII Genes. Nucleic Acids Res. 1984, 12, 8611–8626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seiler-Tuyns, A.; Walker, P.; Martinez, E.; Mérillat, A.M.; Givel, F.; Wahli, W. Identification of Estrogen-Responsive DNA Sequences by Transient Expression Experiments in a Human Breast Cancer Cell Line. Nucleic Acids Res. 1986, 14, 8755–8770. [Google Scholar] [CrossRef] [PubMed]
- Klein-Hitpass, L.; Schorpp, M.; Wagner, U.; Ryffel, G.U. An Estrogen-Responsive Element Derived from the 5’ Flanking Region of the Xenopus Vitellogenin A2 Gene Functions in Transfected Human Cells. Cell 1986, 46, 1053–1061. [Google Scholar] [CrossRef]
- Batistuzzo de Medeiros, S.R.; Krey, G.; Hihi, A.K.; Wahli, W. Functional Interactions between the Estrogen Receptor and the Transcription Activator Sp1 Regulate the Estrogen-Dependent Transcriptional Activity of the Vitellogenin A1 Io Promoter. J. Biol. Chem. 1997, 272, 18250–18260. [Google Scholar] [CrossRef] [Green Version]
- Cardinaux, J.R.; Chapel, S.; Wahli, W. Complex Organization of CTF/NF-I, C/EBP, and HNF3 Binding Sites within the Promoter of the Liver-Specific Vitellogenin Gene. J. Biol. Chem. 1994, 269, 32947–32956. [Google Scholar] [CrossRef]
- Tsai-Pflugfelder, M.; Gasser, S.M.; Wahli, W. Functional Interaction between the Estrogen Receptor and CTF1: Analysis of the Vitellogenin Gene B1 Promoter in Yeast. Mol. Endocrinol. 1998, 12, 1525–1541. [Google Scholar] [CrossRef] [PubMed]
- Robyr, D.; Gegonne, A.; Wolffe, A.P.; Wahli, W. Determinants of Vitellogenin B1 Promoter Architecture. J. Biol. Chem. 2000, 275, 28291–28300. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, C.K.; Navarro-Martin, L.; Neufeld, M.; Basak, A.; Trudeau, V.L. Early Expression of Aromatase and the Membrane Estrogen Receptor GPER in Neuromasts Reveals a Role for Estrogens in the Development of the Frog Lateral Line System. Gen. Comp. Endocrinol. 2014, 205, 242–250. [Google Scholar] [CrossRef]
- Pinto, P.I.S.; Andrade, A.R.; Estêvão, M.D.; Alvarado, M.V.; Felip, A.; Power, D.M. Duplicated Membrane Estrogen Receptors in the European Sea Bass (Dicentrarchus Labrax): Phylogeny, Expression and Regulation throughout the Reproductive Cycle. J. Steroid Biochem. Mol. Biol. 2018, 178, 234–242. [Google Scholar] [CrossRef]
- Thomas, P.; Alyea, R.; Pang, Y.; Peyton, C.; Dong, J.; Berg, A.H. Conserved Estrogen Binding and Signaling Functions of the G Protein-Coupled Estrogen Receptor 1 (GPER) in Mammals and Fish. Steroids 2010, 75, 595–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dovey, J.L.; Vasudevan, N. Does GPER1 Play a Role in Sexual Dimorphism? Front. Endocrinol. 2020, 11, 595895. [Google Scholar] [CrossRef]
- Qie, Y.; Qin, W.; Zhao, K.; Liu, C.; Zhao, L.; Guo, L.-H. Environmental Estrogens and Their Biological Effects through GPER Mediated Signal Pathways. Environ. Pollut. 2021, 278, 116826. [Google Scholar] [CrossRef]
- Gohar, E.Y. G Protein-Coupled Estrogen Receptor 1 as a Novel Regulator of Blood Pressure. Am. J. Physiol. Ren. Physiol. 2020, 319, F612–F617. [Google Scholar] [CrossRef]
- Trenti, A.; Tedesco, S.; Boscaro, C.; Trevisi, L.; Bolego, C.; Cignarella, A. Estrogen, Angiogenesis, Immunity and Cell Metabolism: Solving the Puzzle. Int. J. Mol. Sci. 2018, 19, 859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjimarkou, M.M.; Vasudevan, N. GPER1/GPR30 in the Brain: Crosstalk with Classical Estrogen Receptors and Implications for Behavior. J. Steroid Biochem. Mol. Biol. 2018, 176, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Dufour, C.R.; Giguère, V. ERRα as a Bridge Between Transcription and Function: Role in Liver Metabolism and Disease. Front. Endocrinol. 2019, 10, 206. [Google Scholar] [CrossRef] [Green Version]
- Bardet, P.-L.; Obrecht-Pflumio, S.; Thisse, C.; Laudet, V.; Thisse, B.; Vanacker, J.-M. Cloning and Developmental Expression of Five Estrogen-Receptor Related Genes in the Zebrafish. Dev. Genes Evol. 2004, 214, 240–249. [Google Scholar] [CrossRef]
- Bardet, P.-L.; Horard, B.; Laudet, V.; Vanacker, J.-M. The ERRalpha Orphan Nuclear Receptor Controls Morphogenetic Movements during Zebrafish Gastrulation. Dev. Biol. 2005, 281, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarrant, A.M.; Greytak, S.R.; Callard, G.V.; Hahn, M.E. Estrogen Receptor-Related Receptors in the Killifish Fundulus Heteroclitus: Diversity, Expression, and Estrogen Responsiveness. J. Mol. Endocrinol. 2006, 37, 105–120. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.; Shi, H. Sex Hormones and Their Receptors Regulate Liver Energy Homeostasis. Int. J. Endocrinol. 2015, 2015, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, R.R.; Zangeronimo, M.G.; Pereira, L.J.; Rodrigues, P.B.; Gomide, E.M. Lipoprotein Metabolism in Poultry. World’s Poult. Sci. J. 2011, 67, 431–440. [Google Scholar] [CrossRef]
- Duggan, A.; Paolucci, M.; Tercyak, A.; Gigliotti, M.; Small, D.; Callard, I. Seasonal Variation in Plasma Lipids, Lipoproteins, Apolipoprotein A-I and Vitellogenin in the Freshwater Turtle, Chrysemys Picta. Comp. Biochem. Physiol. Part A 2001, 130, 253–269. [Google Scholar] [CrossRef]
- Price, E.R. The Physiology of Lipid Storage and Use in Reptiles. Biol. Rev. 2017, 92, 1406–1426. [Google Scholar] [CrossRef]
- Levi, L.; Pekarski, I.; Gutman, E.; Fortina, P.; Hyslop, T.; Biran, J.; Levavi-Sivan, B.; Lubzens, E. Revealing Genes Associated with Vitellogenesis in the Liver of the Zebrafish (Danio Rerio) by Transcriptome Profiling. BMC Genom. 2009, 10, 141. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, T.; Xu, C.; Wang, D.; Ren, J.; Li, Y.; Tian, Y.; Wang, Y.; Jiao, Y.; Kang, X.; et al. Transcriptome Profile of Liver at Different Physiological Stages Reveals Potential Mode for Lipid Metabolism in Laying Hens. BMC Genom. 2015, 16, 763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermier, D.; Catheline, D.; Legrand, P. Relationship between Hepatic Fatty Acid Desaturation and Lipid Secretion in the Estrogenized Chicken. Comp. Biochem. Physiol. Part A 1996, 115, 259–264. [Google Scholar] [CrossRef]
- Yang, X.; Rodriguez, M.L.; Leonard, A.; Sun, L.; Fischer, K.A.; Wang, Y.; Ritterhoff, J.; Zhao, L.; Kolwicz, S.C.; Pabon, L.; et al. Fatty Acids Enhance the Maturation of Cardiomyocytes Derived from Human Pluripotent Stem Cells. Stem Cell Rep. 2019, 13, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Xu, N.; Zheng, H.; Tian, W.; Li, H.; Li, Z.; Wang, Y.; Tian, Y.; Kang, X.; Liu, X. Expression of Thyroid Hormone Responsive SPOT 14 Gene Is Regulated by Estrogen in Chicken (Gallus Gallus). Sci. Rep. 2017, 7, 10243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Gu, Z.; Yang, L.; Tian, Y.; Kang, X.; Liu, X. Transcriptome Profile Analysis Reveals an Estrogen Induced LncRNA Associated with Lipid Metabolism and Carcass Traits in Chickens (Gallus Gallus). Cell. Physiol. Biochem. 2018, 50, 1638–1658. [Google Scholar] [CrossRef]
- Tan, W.; Zheng, H.; Wang, D.; Tian, F.; Li, H.; Liu, X. Expression Characteristics and Regulatory Mechanism of Apela Gene in Liver of Chicken (Gallus Gallus). PLoS ONE 2020, 15, e0238784. [Google Scholar] [CrossRef]
- Di Croce, L.; Bruscalupi, G.; Trentalance, A. Independent Responsiveness of Frog Liver Low-Density Lipoprotein Receptor and HMGCoA Reductase to Estrogen Treatment. Pflug. Arch. 1997, 435, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Sellers, J.A.; Hou, L.; Schoenberg, D.R.; Batistuzzo de Medeiros, S.R.; Wahli, W.; Shelness, G.S. Microsomal Triglyceride Transfer Protein Promotes the Secretion of Xenopus Laevis Vitellogenin A1. J. Biol. Chem. 2005, 280, 13902–13905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivessa, N.E.; Rehberg, E.; Kienzle, B.; Seif, F.; Hermann, R.; Hermann, M.; Schneider, W.J.; Gordon, D.A. Molecular Cloning, Expression, and Hormonal Regulation of the Chicken Microsomal Triglyceride Transfer Protein. Gene 2013, 523, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Walzem, R.L.; Hansen, R.J.; Williams, D.L.; Hamilton, R.L. Estrogen Induction of VLDLy Assembly in Egg-Laying Hens. J. Nutr. 1999, 129, 467S–472S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rustaeus, S.; Lindberg, K.; Stillemark, P.; Claesson, C.; Asp, L.; Larsson, T.; Borén, J.; Olofsson, S.O. Assembly of Very Low Density Lipoprotein: A Two-Step Process of Apolipoprotein B Core Lipidation. J. Nutr. 1999, 129, 463S–466S. [Google Scholar] [CrossRef] [PubMed]
- Salvante, K.G.; Lin, G.; Walzem, R.L.; Williams, T.D. Characterization of Very-Low Density Lipoprotein Particle Diameter Dynamics in Relation to Egg Production in a Passerine Bird. J. Exp. Biol. 2007, 210, 1064–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Tang, H.; He, J.; Wu, X.; Wang, L.; Liu, X.; Lin, H. Interaction of Nuclear ERs and GPER in Vitellogenesis in Zebrafish. J. Steroid Biochem. Mol. Biol. 2019, 189, 10–18. [Google Scholar] [CrossRef]
- Segars, J.H.; Marks, M.S.; Hirschfeld, S.; Driggers, P.H.; Martinez, E.; Grippo, J.F.; Brown, M.; Wahli, W.; Ozato, K. Inhibition of Estrogen-Responsive Gene Activation by the Retinoid X Receptor Beta: Evidence for Multiple Inhibitory Pathways. Mol. Cell. Biol. 1993, 13, 2258–2268. [Google Scholar] [CrossRef] [Green Version]
- Schippers, I.J.; Kloppenburg, M.; Snippe, L.; Ab, G. 9-Cis-Retinoic Acid Represses Estrogen-Induced Expression of the Very Low Density Apolipoprotein II Gene. Mol. Cell. Endocrinol. 1994, 105, 175–182. [Google Scholar] [CrossRef]
- Nuñez, S.B.; Medin, J.A.; Braissant, O.; Kemp, L.; Wahli, W.; Ozato, K.; Segars, J.H. Retinoid X Receptor and Peroxisome Proliferator-Activated Receptor Activate an Estrogen Responsive Gene Independent of the Estrogen Receptor. Mol. Cell. Endocrinol. 1997, 127, 27–40. [Google Scholar] [CrossRef]
- Keller, H.; Givel, F.; Perroud, M.; Wahli, W. Signaling Cross-Talk between Peroxisome Proliferator-Activated Receptor/Retinoid X Receptor and Estrogen Receptor through Estrogen Response Elements. Mol. Endocrinol. 1995, 9, 794–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabelo, E.M.; Baker, B.S.; Tata, J.R. Interplay between Thyroid Hormone and Estrogen in Modulating Expression of Their Receptor and Vitellogenin Genes during Xenopus Metamorphosis. Mech. Dev. 1994, 45, 49–57. [Google Scholar] [CrossRef]
- Nelson, E.R.; Habibi, H.R. Thyroid Hormone Regulates Vitellogenin by Inducing Estrogen Receptor Alpha in the Goldfish Liver. Mol. Cell. Endocrinol. 2016, 436, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Hammouche, S.B.; Remana, S.; Exbrayat, J.-M. Immunolocalization of Hepatic Estrogen and Progesterone Receptors in the Female Lizard Uromastyx Acanthinura. Comptes Rendus Biol. 2012, 335, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Slater, E.P.; Redeuilh, G.; Beato, M. Hormonal Regulation of Vitellogenin Genes: An Estrogen-Responsive Element in the Xenopus A2 Gene and a Multihormonal Regulatory Region in the Chicken II Gene. Mol. Endocrinol. 1991, 5, 386–396. [Google Scholar] [CrossRef] [Green Version]
- Lethimonier, C.; Flouriot, G.; Valotaire, Y.; Kah, O.; Ducouret, B. Transcriptional Interference between Glucocorticoid Receptor and Estradiol Receptor Mediates the Inhibitory Effect of Cortisol on Fish Vitellogenesis. Biol. Reprod. 2000, 62, 1763–1771. [Google Scholar] [CrossRef] [Green Version]
- Karmakar, S.; Jin, Y.; Nagaich, A.K. Interaction of Glucocorticoid Receptor (GR) with Estrogen Receptor (ER) α and Activator Protein 1 (AP1) in Dexamethasone-Mediated Interference of ERα Activity. J. Biol. Chem. 2013, 288, 24020–24034. [Google Scholar] [CrossRef] [Green Version]
- Toporova, L.; Balaguer, P. Nuclear Receptors Are the Major Targets of Endocrine Disrupting Chemicals. Mol. Cell. Endocrinol. 2020, 502, 110665. [Google Scholar] [CrossRef]
- Le Magueresse-Battistoni, B. Endocrine Disrupting Chemicals and Metabolic Disorders in the Liver: What If We Also Looked at the Female Side? Chemosphere 2021, 268, 129212. [Google Scholar] [CrossRef]
- Delfosse, V.; Grimaldi, M.; Cavaillès, V.; Balaguer, P.; Bourguet, W. Structural and Functional Profiling of Environmental Ligands for Estrogen Receptors. Environ. Health Perspect. 2014, 122, 1306–1313. [Google Scholar] [CrossRef] [Green Version]
- Sumpter, J.P.; Jobling, S. Vitellogenesis as a Biomarker for Estrogenic Contamination of the Aquatic Environment. Environ. Health Perspect. 1995, 103 (Suppl. 7), 173–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, J.; Pan, Z.; Zhang, Y.; Zhang, X.; Tian, H.; Wang, W.; Ru, S. Distribution of Vitellogenin in Japanese Flounder (Paralichthys Olivaceus) for Biomarker Analysis of Marine Environmental Estrogens. Aquat. Toxicol. 2019, 216, 105321. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Lu, L.; Li, Y.; Wei, Y.; Cheng, Y.; Zhang, X.; Tian, H.; Wang, W.; Ru, S. Genotoxic Biomarkers and Histological Changes in Marine Medaka (Oryzias Melastigma) Exposed to 17α-Ethynylestradiol and 17β-Trenbolone. Mar. Pollut. Bull. 2020, 150, 110601. [Google Scholar] [CrossRef] [PubMed]
- Amano, H.; Uno, S.; Koyama, J.; Hiramatsu, N.; Todo, T.; Hara, A. Development of Specific Enzyme-Linked Immunosorbent Assays for Multiple Vitellogenins in Marbled Sole, Pleuronectes Yokohamae. Gen. Comp. Endocrinol. 2019, 281, 67–72. [Google Scholar] [CrossRef]
- Amano, H.; Kotake, A.; Hiramatsu, N.; Fujita, T.; Todo, T.; Aoki, J.; Soyano, K.; Kagawa, H.; Hara, A. Development of Specific Chemiluminescent Immunoassays for Three Subtypes of Vitellogenin in Grey Mullet (Mugil Cephalus). Gen. Comp. Endocrinol. 2019, 271, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Martyniuk, C.J.; Feswick, A.; Munkittrick, K.R.; Dreier, D.A.; Denslow, N.D. Twenty Years of Transcriptomics, 17alpha-Ethinylestradiol, and Fish. Gen. Comp. Endocrinol. 2020, 286, 113325. [Google Scholar] [CrossRef]
- Rojas-Hucks, S.; Gutleb, A.C.; González, C.M.; Contal, S.; Mehennaoui, K.; Jacobs, A.; Witters, H.E.; Pulgar, J. Xenopus Laevis as a Bioindicator of Endocrine Disruptors in the Region of Central Chile. Arch. Environ. Contam. Toxicol. 2019, 77, 390–408. [Google Scholar] [CrossRef]
- McArdle, M.E.; Freeman, E.L.; Staveley, J.P.; Ortego, L.S.; Coady, K.K.; Weltje, L.; Weyers, A.; Wheeler, J.R.; Bone, A.J. Critical Review of Read-Across Potential in Testing for Endocrine-Related Effects in Vertebrate Ecological Receptors. Environ. Toxicol. Chem. 2020, 39, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.C.; Wheeler, J.R. A Critical Review of Histopathological Findings Associated with Endocrine and Non-Endocrine Hepatic Toxicity in Fish Models. Aquat. Toxicol. 2018, 197, 60–78. [Google Scholar] [CrossRef] [PubMed]
- De Lapuente, J.; Lourenço, J.; Mendo, S.A.; Borràs, M.; Martins, M.G.; Costa, P.M.; Pacheco, M. The Comet Assay and Its Applications in the Field of Ecotoxicology: A Mature Tool That Continues to Expand Its Perspectives. Front. Genet. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ru, S.; Wang, J.; Wang, D.; Zhao, L.; Teng, H.; Dai, Z.; Zhang, W.; Yang, A. Ultrasensitive Label-Free Electrochemical Immunosensors for Detecting Marine Medaka (Oryzias Melastigma) Vitellogenin Based on Novel Cu2O–BSA Nanoparticles and Anti-Lipovitellin Monoclonal Antibody. Sens. Actuators B 2021, 345, 130358. [Google Scholar] [CrossRef]
- Verderame, M.; Limatola, E. Interferences of an Environmental Pollutant with Estrogen-like Action in the Male Reproductive System of the Terrestrial Vertebrate Podarcis Sicula. Gen. Comp. Endocrinol. 2015, 213, 9–15. [Google Scholar] [CrossRef]
- Marsili, L.; Casini, S.; Mori, G.; Ancora, S.; Bianchi, N.; D’Agostino, A.; Ferraro, M.; Fossi, M.C. The Italian Wall Lizard (Podarcis Sicula) as a Bioindicator of Oil Field Activity. Sci. Total Environ. 2009, 407, 3597–3604. [Google Scholar] [CrossRef] [PubMed]
- Verderame, M.; Scudiero, R. How Glyphosate Impairs Liver Condition in the Field Lizard Podarcis Siculus (Rafinesque-Schmaltz, 1810): Histological and Molecular Evidence. BioMed Res. Int. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Verderame, M.; Limatola, E.; Scudiero, R. Estrogenic Contamination by Manure Fertilizer in Organic Farming: A Case Study with the Lizard Podarcis Sicula. Ecotoxicology 2016, 25, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garmshausen, J.; Kloas, W.; Hoffmann, F. 17α-Ethinylestradiol Can Disrupt Hemoglobin Catabolism in Amphibians. Comp. Biochem. Physiol. Part C 2015, 171, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Ankley, G.T.; LaLone, C.A.; Gray, L.E.; Villeneuve, D.L.; Hornung, M.W. Evaluation of the Scientific Underpinnings for Identifying Estrogenic Chemicals in Nonmammalian Taxa Using Mammalian Test Systems: Prioritizing Potential Estrogenic Chemicals. Environ. Toxicol. Chem. 2016, 35, 2806–2816. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.K.; Kim, J.S.; Ahn, H.J.; Hwang, J.H.; Kim, J.M.; Lee, H.T.; An, B.K.; Kang, C.W. Changes in Hepatic Lipid Parameters and Hepatic Messenger Ribonucleic Acid Expression Following Estradiol Administration in Laying Hens (Gallus Domesticus). Poult. Sci. 2010, 89, 2660–2667. [Google Scholar] [CrossRef]
- Trott, K.A.; Giannitti, F.; Rimoldi, G.; Hill, A.; Woods, L.; Barr, B.; Anderson, M.; Mete, A. Fatty Liver Hemorrhagic Syndrome in the Backyard Chicken: A Retrospective Histopathologic Case Series. Vet. Pathol. 2014, 51, 787–795. [Google Scholar] [CrossRef]
- Miao, Y.F.; Gao, X.N.; Xu, D.N.; Li, M.C.; Gao, Z.S.; Tang, Z.H.; Mhlambi, N.H.; Wang, W.J.; Fan, W.T.; Shi, X.Z.; et al. Protective Effect of the New Prepared Atractylodes Macrocephala Koidz Polysaccharide on Fatty Liver Hemorrhagic Syndrome in Laying Hens. Poult. Sci. 2021, 100, 938–948. [Google Scholar] [CrossRef]
- Shini, A.; Shini, S.; Bryden, W.L. Fatty Liver Haemorrhagic Syndrome Occurrence in Laying Hens: Impact of Production System. Avian Pathol. 2019, 48, 25–34. [Google Scholar] [CrossRef]
- Liang, M.J.; Wang, Z.P.; Xu, L.; Leng, L.; Wang, S.Z.; Luan, P.; Cao, Z.P.; Li, Y.M.; Li, H. Estimating the Genetic Parameters for Liver Fat Traits in Broiler Lines Divergently Selected for Abdominal Fat. Genet. Mol. Res. 2015, 14, 9646–9654. [Google Scholar] [CrossRef] [PubMed]
- Polin, D.; Wolford, J.H. Role of Estrogen as a Cause of Fatty Liver Hemorrhagic Syndrome. J. Nutr. 1977, 107, 873–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haghighi-Rad, F.; Polin, D. The Relationship of Plasma Estradiol and Progesterone Levels to the Fatty Liver Hemorrhagic Syndrome in Laying Hens. Poult. Sci. 1981, 60, 2278–2283. [Google Scholar] [CrossRef]
- Shini, S.; Shini, A.; Bryden, W.L. Unravelling Fatty Liver Haemorrhagic Syndrome: 1. Oestrogen and Inflammation. Avian Pathol. 2020, 49, 87–98. [Google Scholar] [CrossRef]
- Dong, X.; Tong, J. Different Susceptibility to Fatty Liver-Haemorrhagic Syndrome in Young and Older Layers and the Interaction on Blood LDL-C Levels between Oestradiols and High Energy-Low Protein Diets. Br. Poult. Sci. 2019, 60, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zeng, Q.; Li, F.; Fang, H.; Zhou, Z.; Jiang, T.; Yin, C.; Wei, Q.; Wang, Y.; Ruan, J.; et al. Dysregulated H3K27 Acetylation Is Implicated in Fatty Liver Hemorrhagic Syndrome in Chickens. Front. Genet. 2021, 11, 574167. [Google Scholar] [CrossRef]
- Tan, X.; Liu, R.; Zhang, Y.; Wang, X.; Wang, J.; Wang, H.; Zhao, G.; Zheng, M.; Wen, J. Integrated Analysis of the Methylome and Transcriptome of Chickens with Fatty Liver Hemorrhagic Syndrome. BMC Genom. 2021, 22, 8. [Google Scholar] [CrossRef]
- Montagner, A.; Polizzi, A.; Fouché, E.; Ducheix, S.; Lippi, Y.; Lasserre, F.; Barquissau, V.; Régnier, M.; Lukowicz, C.; Benhamed, F.; et al. Liver PPARα Is Crucial for Whole-Body Fatty Acid Homeostasis and Is Protective against NAFLD. Gut 2016, 65, 1202–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smati, S.; Polizzi, A.; Fougerat, A.; Ellero-Simatos, S.; Blum, Y.; Lippi, Y.; Régnier, M.; Laroyenne, A.; Huillet, M.; Arif, M.; et al. Integrative Study of Diet-Induced Mouse Models of NAFLD Identifies PPARα as a Sexually Dimorphic Drug Target. Gut 2021, gutjnl-2020-323323. [Google Scholar] [CrossRef] [PubMed]
- Della Torre, S. Non-Alcoholic Fatty Liver Disease as a Canonical Example of Metabolic Inflammatory-Based Liver Disease Showing a Sex-Specific Prevalence: Relevance of Estrogen Signaling. Front. Endocrinol. 2020, 11, 572490. [Google Scholar] [CrossRef]
- Hermier, D.; Rousselot-Pailley, D.; Peresson, R.; Sellier, N. Influence of Orotic Acid and Estrogen on Hepatic Lipid Storage and Secretion in the Goose Susceptible to Liver Steatosis. Biochim. Biophys. Acta 1994, 1211, 97–106. [Google Scholar] [CrossRef]
- Turola, E.; Petta, S.; Vanni, E.; Milosa, F.; Valenti, L.; Critelli, R.; Miele, L.; Maccio, L.; Calvaruso, V.; Fracanzani, A.L.; et al. Ovarian Senescence Increases Liver Fibrosis in Humans and Zebrafish with Steatosis. Dis. Models Mech. 2015, dmm.019950. [Google Scholar] [CrossRef] [Green Version]
- Ballestri, S.; Nascimbeni, F.; Baldelli, E.; Marrazzo, A.; Romagnoli, D.; Lonardo, A. NAFLD as a Sexual Dimorphic Disease: Role of Gender and Reproductive Status in the Development and Progression of Nonalcoholic Fatty Liver Disease and Inherent Cardiovascular Risk. Adv. Ther. 2017, 34, 1291–1326. [Google Scholar] [CrossRef]
- Asaoka, Y.; Terai, S.; Sakaida, I.; Nishina, H. The Expanding Role of Fish Models in Understanding Non-Alcoholic Fatty Liver Disease. Dis. Models Mech. 2014, 7, 409. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, K.; Takami, T.; Nagatomo, T.; Fukui, Y.; Hoshida, H.; Saeki, I.; Matsumoto, T.; Hidaka, I.; Yamamoto, N.; Sakaida, I. Usefulness of Adult Medaka Fish as a Model for the Evaluation of Alcoholic Fatty Liver. Alcohol 2019, 77, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Terai, S.; Oishi, T.; Kuwashiro, S.; Fujisawa, K.; Yamamoto, N.; Fujita, Y.; Hamamoto, Y.; Furutani-Seiki, M.; Nishina, H.; et al. Medaka as a Model for Human Nonalcoholic Steatohepatitis. Dis. Models Mech. 2010, 3, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Katoch, S.; Patial, V. Zebrafish: An Emerging Model System to Study Liver Diseases and Related Drug Discovery. J. Appl. Toxicol. 2021, 41, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Knowland, J.; Theulaz, I.; Wright, C.V.; Wahli, W. Injection of Partially Purified Estrogen Receptor Protein from Xenopus Liver Nuclei into Oocytes Activates the Silent Vitellogenin Locus. Proc. Natl. Acad. Sci. USA 1984, 81, 5777–5781. [Google Scholar] [CrossRef] [Green Version]
- Green, S.; Kumar, V.; Theulaz, I.; Wahli, W.; Chambon, P. The N-Terminal DNA-Binding “zinc Finger” of the Oestrogen and Glucocorticoid Receptors Determines Target Gene Specificity. EMBO J. 1988, 7, 3037–3044. [Google Scholar] [CrossRef]
- Wong, V.W.-S.; Adams, L.A.; de Lédinghen, V.; Wong, G.L.-H.; Sookoian, S. Noninvasive Biomarkers in NAFLD and NASH—Current Progress and Future Promise. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.T.; Shulman, G.I. Nonalcoholic Fatty Liver Disease as a Nexus of Metabolic and Hepatic Diseases. Cell. Metab. 2018, 27, 22–41. [Google Scholar] [CrossRef] [Green Version]
- Younossi, Z.; Tacke, F.; Arrese, M.; Chander Sharma, B.; Mostafa, I.; Bugianesi, E.; Wai-Sun Wong, V.; Yilmaz, Y.; George, J.; Fan, J.; et al. Global Perspectives on Nonalcoholic Fatty Liver Disease and Nonalcoholic Steatohepatitis. Hepatology 2019, 69, 2672–2682. [Google Scholar] [CrossRef] [Green Version]
- Ruggieri, A.; Gagliardi, M.C.; Anticoli, S. Sex-Dependent Outcome of Hepatitis B and C Viruses Infections: Synergy of Sex Hormones and Immune Responses? Front. Immunol. 2018, 9, 2302. [Google Scholar] [CrossRef] [Green Version]
- Lonardo, A.; Nascimbeni, F.; Ballestri, S.; Fairweather, D.; Win, S.; Than, T.A.; Abdelmalek, M.F.; Suzuki, A. Sex Differences in Nonalcoholic Fatty Liver Disease: State of the Art and Identification of Research Gaps. Hepatology 2019, 70, 1457–1469. [Google Scholar] [CrossRef]
- Grossmann, M.; Wierman, M.E.; Angus, P.; Handelsman, D.J. Reproductive Endocrinology of Nonalcoholic Fatty Liver Disease. Endocr. Rev. 2019, 40, 417–446. [Google Scholar] [CrossRef] [PubMed]
- Molnár, S.; Szőllősi, L. Sustainability and Quality Aspects of Different Table Egg Production Systems: A Literature Review. Sustainability 2020, 12, 7884. [Google Scholar] [CrossRef]
- Iannotti, L.L.; Lutter, C.K.; Bunn, D.A.; Stewart, C.P. Eggs: The Uncracked Potential for Improving Maternal and Young Child Nutrition among the World’s Poor. Nutr. Rev. 2014, 72, 355–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutter, C.K.; Iannotti, L.L.; Stewart, C.P. The Potential of a Simple Egg to Improve Maternal and Child Nutrition. Matern. Child Nutr. 2018, 14 (Suppl. 3), e12678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wake, M.H. Modes of Reproduction Verts: Hermaphroditism, Viviparity, Oviparity, Ovoviviparity: (General Definition with Examples). In Encyclopedia of Reproduction; Elsevier: Amsterdam, The Netherlands, 2018; pp. 18–22. ISBN 978-0-12-815145-7. [Google Scholar]
- Regnault, C.; Usal, M.; Veyrenc, S.; Couturier, K.; Batandier, C.; Bulteau, A.-L.; Lejon, D.; Sapin, A.; Combourieu, B.; Chetiveaux, M.; et al. Unexpected Metabolic Disorders Induced by Endocrine Disruptors in Xenopus Tropicalis Provide New Lead for Understanding Amphibian Decline. Proc. Natl. Acad. Sci. USA 2018, 115, E4416–E4425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giang, P.T.; Sakalli, S.; Fedorova, G.; Tilami, S.K.; Bakal, T.; Najmanova, L.; Grabicova, K.; Kolarova, J.; Sampels, S.; Zamaratskaia, G.; et al. Biomarker Response, Health Indicators, and Intestinal Microbiome Composition in Wild Brown Trout (Salmo Trutta m. Fario L.) Exposed to a Sewage Treatment Plant Effluent-Dominated Stream. Sci. Total Environ. 2018, 625, 1494–1509. [Google Scholar] [CrossRef]
- Viganò, L.; Loizeau, J.-L.; Mandich, A.; Mascolo, G. Medium- and Long-Term Effects of Estrogenic Contaminants on the Middle River Po Fish Community as Reconstructed from a Sediment Core. Arch. Environ. Contam. Toxicol. 2016, 71, 454–472. [Google Scholar] [CrossRef] [PubMed]
- Prado, P.S.; Pinheiro, A.P.B.; Bazzoli, N.; Rizzo, E. Reproductive Biomarkers Responses Induced by Xenoestrogens in the Characid Fish Astyanax Fasciatus Inhabiting a South American Reservoir: An Integrated Field and Laboratory Approach. Environ. Res. 2014, 131, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Urbatzka, R.; Rocha, E.; Reis, B.; Cruzeiro, C.; Monteiro, R.A.F.; Rocha, M.J. Effects of Ethinylestradiol and of an Environmentally Relevant Mixture of Xenoestrogens on Steroidogenic Gene Expression and Specific Transcription Factors in Zebrafish. Environ. Pollut. 2012, 164, 28–35. [Google Scholar] [CrossRef]
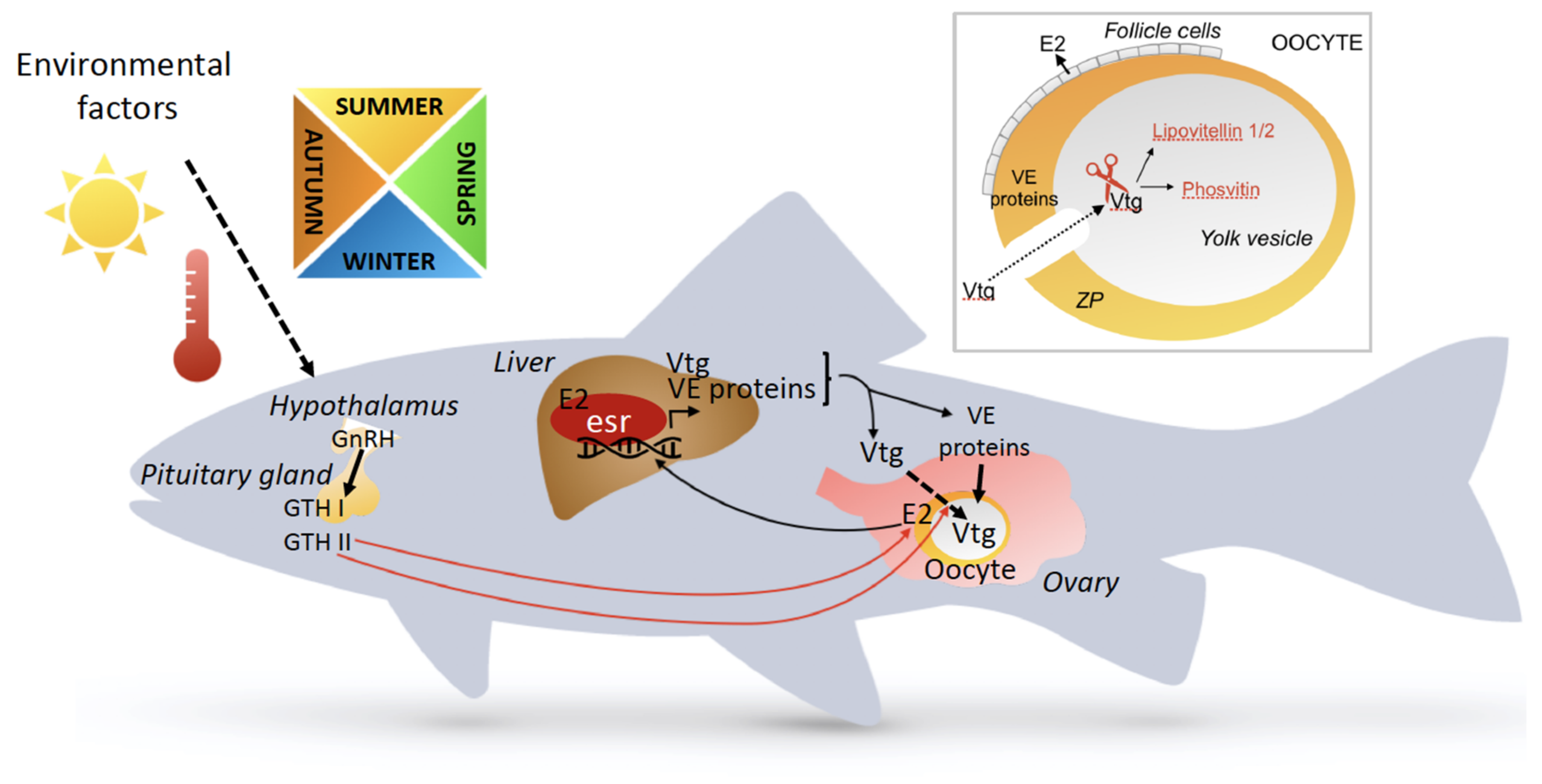
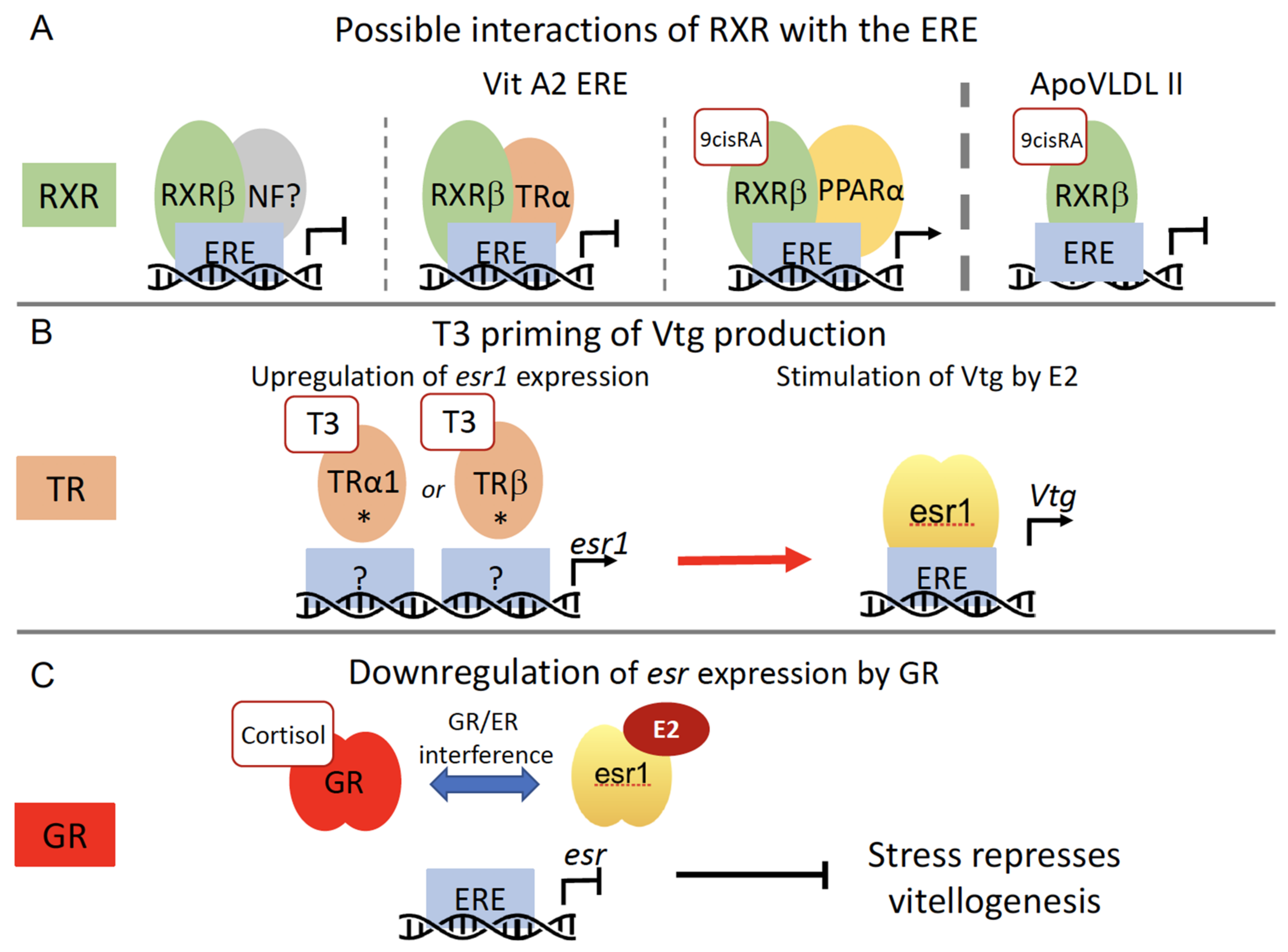
 transport;
transport;  stimulation.
transport; stimulation.
stimulation.
transport; stimulation.

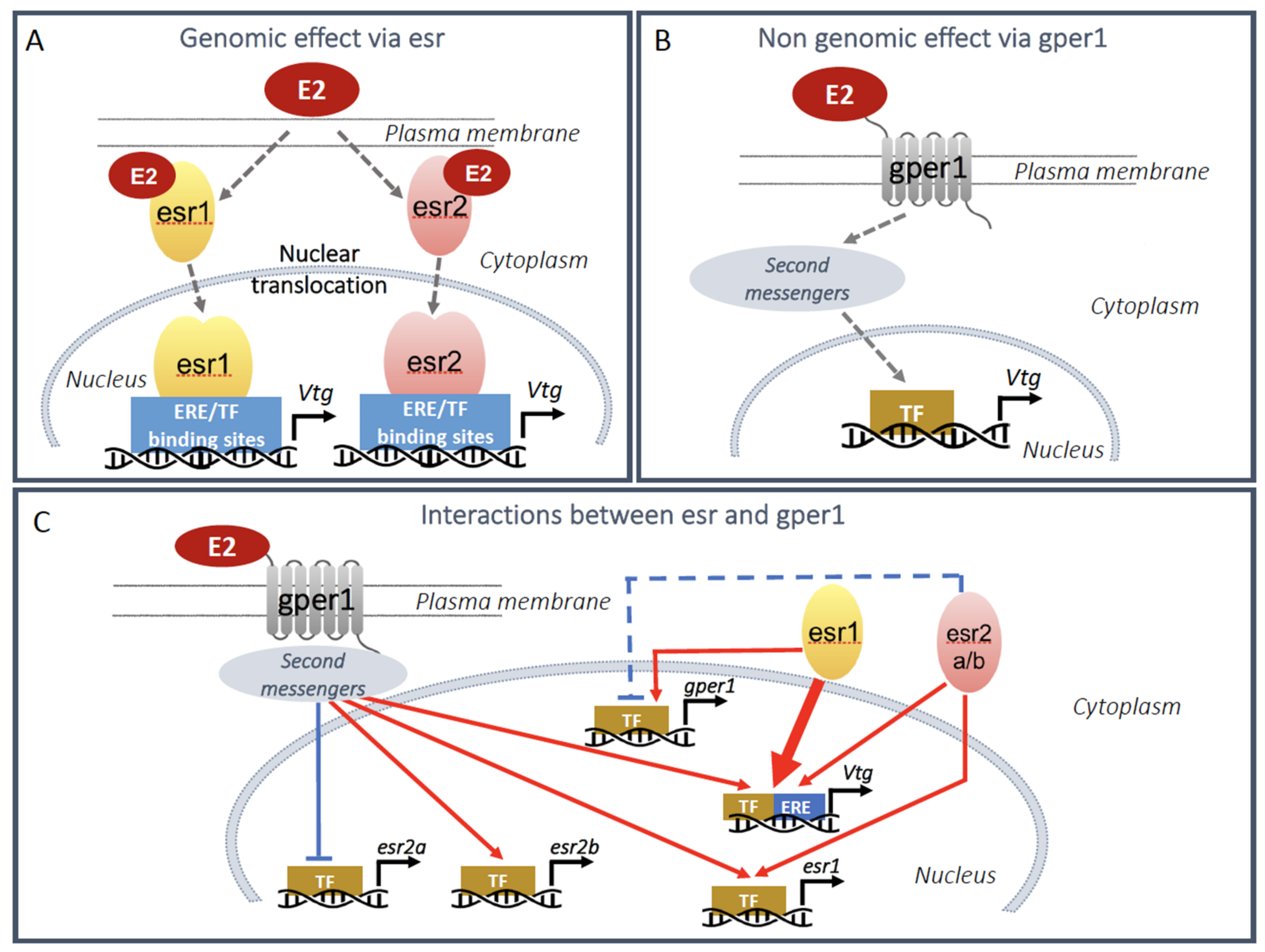
 inhibition;
inhibition;  stimulation. The blue dashed line indicates an effect to be confirmed.
inhibition; stimulation. The blue dashed line indicates an effect to be confirmed.
stimulation. The blue dashed line indicates an effect to be confirmed.
inhibition; stimulation. The blue dashed line indicates an effect to be confirmed.

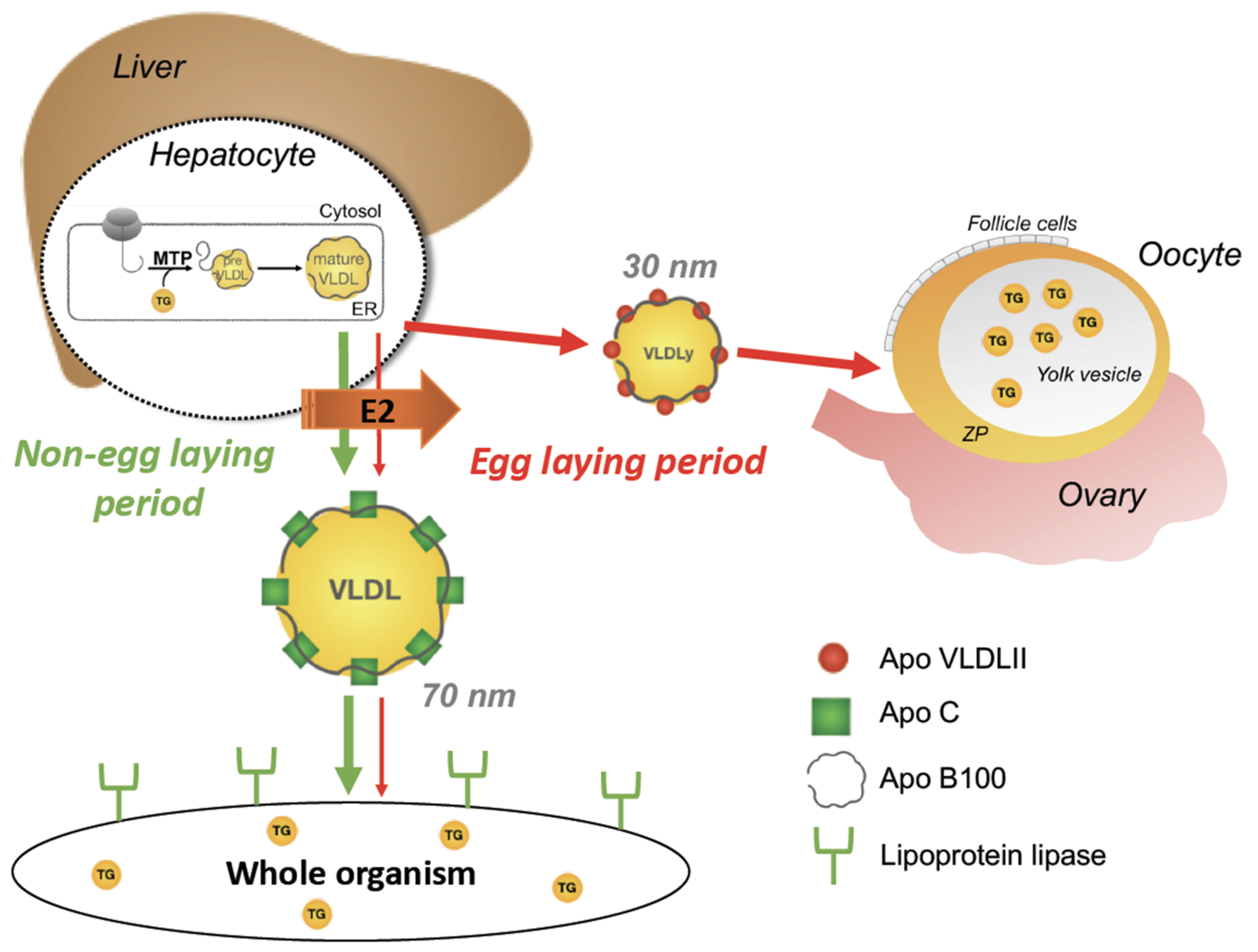
 inhibition;
inhibition;  stimulation.
inhibition; stimulation.
stimulation.
inhibition; stimulation.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tramunt, B.; Montagner, A.; Tan, N.S.; Gourdy, P.; Rémignon, H.; Wahli, W. Roles of Estrogens in the Healthy and Diseased Oviparous Vertebrate Liver. Metabolites 2021, 11, 502. https://doi.org/10.3390/metabo11080502
Tramunt B, Montagner A, Tan NS, Gourdy P, Rémignon H, Wahli W. Roles of Estrogens in the Healthy and Diseased Oviparous Vertebrate Liver. Metabolites. 2021; 11(8):502. https://doi.org/10.3390/metabo11080502
Chicago/Turabian StyleTramunt, Blandine, Alexandra Montagner, Nguan Soon Tan, Pierre Gourdy, Hervé Rémignon, and Walter Wahli. 2021. "Roles of Estrogens in the Healthy and Diseased Oviparous Vertebrate Liver" Metabolites 11, no. 8: 502. https://doi.org/10.3390/metabo11080502
APA StyleTramunt, B., Montagner, A., Tan, N. S., Gourdy, P., Rémignon, H., & Wahli, W. (2021). Roles of Estrogens in the Healthy and Diseased Oviparous Vertebrate Liver. Metabolites, 11(8), 502. https://doi.org/10.3390/metabo11080502