
Lipidomic Signatures of Changes in Adiposity: A Large Prospective Study of 5849 Adults from the Australian Diabetes, Obesity and Lifestyle Study

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Characteristics of Participants
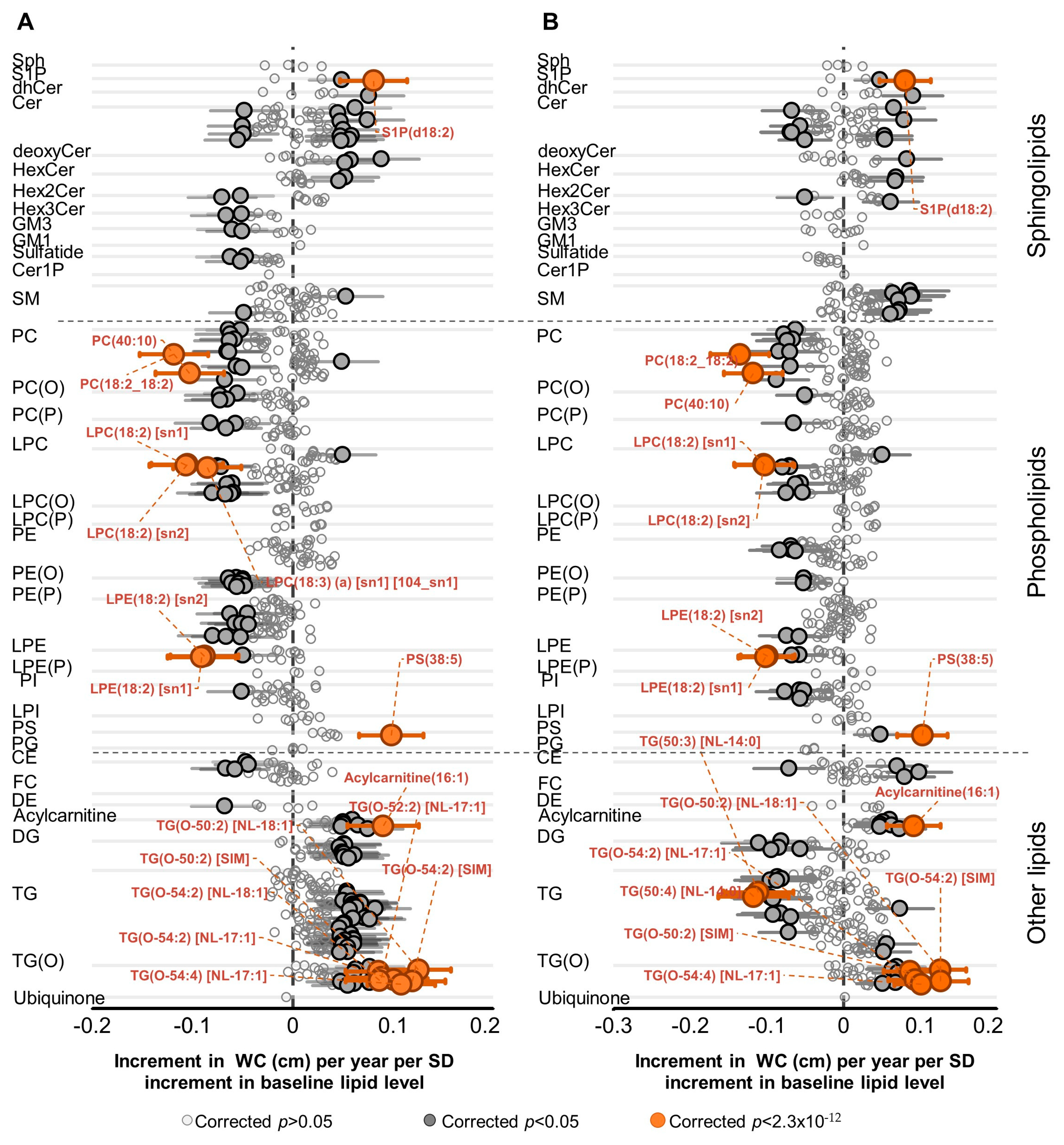
2.2. Association of Lipid Species with Change in WC
2.3. Association of Lipid Species with Change in BMI
2.4. Overlapping Associations of Lipid Species with Change in WC and Change in BMI
2.5. Multivariate Modeling to Predict Change in Waist Circumference
3. Discussion
4. Materials and Methods
4.1. Study Design and Participants
4.2. Data Collection and Laboratory Measurements
4.3. Ethics
4.4. Plasma Lipidomic Profiling
4.4.1. Lipid Extraction
4.4.2. Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS)
4.4.3. Lipid Classes/Subclasses and Species
4.4.4. Data Processing
4.5. Statistics
4.5.1. Univariate Analyses
4.5.2. Multivariate Modeling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reilly, J.J.; El-Hamdouchi, A.; Diouf, A.; Monyeki, A.; Somda, S.A. Determining the worldwide prevalence of obesity. Lancet 2018, 391, 1773–1774. [Google Scholar] [CrossRef]
- Adab, P.; Pallan, M.; Whincup, P.H. Is BMI the best measure of obesity? BMJ 2018, 360, k1274. [Google Scholar] [CrossRef] [Green Version]
- Hajian-Tilaki, K.; Heidari, B. Is waist circumference a better predictor of diabetes than body mass index or waist-to-height ratio in Iranian adults? Int. J. Prev. Med. 2015, 6, 5. [Google Scholar] [CrossRef]
- Janiszewski, P.M.; Janssen, I.; Ross, R. Does Waist Circumference Predict Diabetes and Cardiovascular Disease Beyond Commonly Evaluated Cardiometabolic Risk Factors? Diabetes Care 2007, 30, 3105–3109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias-Audibert, F.L.; Navarro, L.C.; de Oliveira, D.N.; Delafiori, J.; Melo, C.F.O.R.; Guerreiro, T.M.; Rosa, F.T.; Petenuci, D.L.; Watanabe, M.A.E.; Velloso, L.A.; et al. Combining Machine Learning and Metabolomics to Identify Weight Gain Biomarkers. Front. Bioeng. Biotechnol. 2020, 8, 6. [Google Scholar] [CrossRef]
- Lamichhane, S.; Dickens, A.M.; Sen, P.; Laurikainen, H.; Suvisaari, J.; Hyotylainen, T.; Howes, O.; Hietala, J.; Oresic, M. Association Between Circulating Lipids and Future Weight Gain in Individuals With an At-Risk Mental State and in First-Episode Psychosis. Schizophrenia Bulletin. 2021, 47, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Cirulli, E.T.; Guo, L.; Leon Swisher, C.; Shah, N.; Huang, L.; Napier, L.A.; Kirkness, E.F.; Spector, T.D.; Caskey, C.T.; Thorens, B.; et al. Profound Perturbation of the Metabolome in Obesity Is Associated with Health Risk. Cell Metab. 2019, 29, 488–500.e2. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Ren, Y.; Wang, C.; Li, L.; Zhang, L.; Wang, B.; Zhao, Y.; Han, C.; Zhang, H.; Yang, X.; et al. Association of change in waist circumference and dyslipidaemia risk: The rural Chinese cohort study. Diabetes Metab. Res. Rev. 2018, 34, e2949. [Google Scholar] [CrossRef] [PubMed]
- Würtz, P.; Wang, Q.; Kangas, A.J.; Richmond, R.C.; Skarp, J.; Tiainen, M.; Tynkkynen, T.; Soininen, P.; Havulinna, A.S.; Kaakinen, M.; et al. Metabolic signatures of adiposity in young adults: Mendelian randomization analysis and effects of weight change. PLoS Med. 2014, 11, e1001765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäntyselkä, P.; Kautiainen, H.; Saltevo, J.; Würtz, P.; Soininen, P.; Kangas, A.J.; Ala-Korpela, M.; Vanhala, M. Weight change and lipoprotein particle concentration and particle size: A cohort study with 6.5-year follow-up. Atherosclerosis 2012, 223, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, D.W.; Zimmet, P.Z.; Welborn, T.A.; Cameron, A.J.; Shaw, J.; de Courten, M.; Jolley, D.; McCarty, D.J. The Australian Diabetes, Obesity and Lifestyle Study (AusDiab)—methods and response rates. Diabetes Res. Clin. Pract. 2002, 57, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Willinger, C.M.; Keefe, J.; Liu, J.; Fernández-Ortiz, A.; Ibáñez, B.; Peñalvo, J.; Adourian, A.; Chen, G.; Corella, D.; et al. Lipidomic profiling identifies signatures of metabolic risk. EBioMedicine 2020, 51, 102520. [Google Scholar] [CrossRef] [Green Version]
- Meikle, P.J.; Wong, G.; Barlow, C.K.; Weir, J.M.; Greeve, M.A.; MacIntosh, G.L.; Almasy, L.; Comuzzie, A.G.; Mahaney, M.C.; Kowalczyk, A.; et al. Plasma Lipid Profiling Shows Similar Associations with Prediabetes and Type 2 Diabetes. PLoS ONE 2013, 8, e74341. [Google Scholar]
- Khan, A.A.; Mundra, P.A.; Straznicky, N.E.; Nestel, P.J.; Wong, G.; Tan, R.; Huynh, K.; Ng, T.W.; Mellett, N.A.; Weir, J.M.; et al. Weight Loss and Exercise Alter the High-Density Lipoprotein Lipidome and Improve High-Density Lipoprotein Functionality in Metabolic Syndrome. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 438–447. [Google Scholar] [CrossRef] [Green Version]
- Ahola-Olli, A.V.; Mustelin, L.; Kalimeri, M.; Kettunen, J.; Jokelainen, J.; Auvinen, J.; Puukka, K.; Havulinna, A.S.; Lehtimäki, T.; Kähönen, M.; et al. Circulating metabolites and the risk of type 2 diabetes: A prospective study of 11,896 young adults from four Finnish cohorts. Diabetologia 2019, 62, 2298–2309. [Google Scholar] [CrossRef] [Green Version]
- Mihalik, S.J.; Goodpaster, B.H.; Kelley, D.E.; Chace, D.H.; Vockley, J.; Toledo, F.G.S.; DeLany, J.P. Increased levels of plasma acylcarnitines in obesity and type 2 diabetes and identification of a marker of glucolipotoxicity. Obesity 2010, 18, 1695–1700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanamas, S.K.; Shaw, J.E.; Backholer, K.; Magliano, D.J.; Peeters, A. Twelve-year weight change, waist circumference change and incident obesity: The Australian diabetes, obesity and lifestyle study. Obesity 2014, 22, 1538–1545. [Google Scholar] [CrossRef] [PubMed]
- Pearcey, S.M.; de Castro, J.M. Food intake and meal patterns of weight-stable and weight-gaining persons. Am. J. Clin. Nutr. 2002, 76, 107–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambring, A.; Johansson, M.; Axelsen, M.; Gan, L.; Strandvik, B.; Friberg, P. Mediterranean-inspired diet lowers the ratio of serum phospholipid n–6 to n–3 fatty acids, the number of leukocytes and platelets, and vascular endothelial growth factor in healthy subjects. Am. J. Clin. Nutr. 2006, 83, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.E.; Xanthakis, V.; Moore, L.L.; Vasan, R.S.; Jacques, P.F. Cumulative sugar-sweetened beverage consumption is associated with higher concentrations of circulating ceramides in the Framingham Offspring Cohort. Am. J. Clin. Nutr. 2020, 111, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Shih, C.W.; Hauser, M.E.; Aronica, L.; Rigdon, J.; Gardner, C.D. Changes in blood lipid concentrations associated with changes in intake of dietary saturated fat in the context of a healthy low-carbohydrate weight-loss diet: A secondary analysis of the Diet Intervention Examining the Factors Interacting with Treatment Success (DIETFITS) trial. Am. J. Clin. Nutr. 2019, 109, 433–441. [Google Scholar]
- Arner, P.; Andersson, D.P.; Bäckdahl, J.; Dahlman, I.; Rydén, M. Weight Gain and Impaired Glucose Metabolism in Women Are Predicted by Inefficient Subcutaneous Fat Cell Lipolysis. Cell Metab. 2018, 28, 45–54.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyene, H.B.; Olshansky, G.; AA, T.S.; Giles, C.; Huynh, K.; Cinel, M.; Mellett, N.A.; Cadby, G.; Hung, J.; Hui, J.; et al. High-coverage plasma lipidomics reveals novel sex-specific lipidomic fingerprints of age and BMI: Evidence from two large population cohort studies. PLoS Biol. 2020, 18, e3000870. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [Green Version]
- Bajzer, M.; Seeley, R.J. Physiology: Obesity and gut flora. Nature 2006, 444, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Forouhi, N.G.; Imamura, F.; Sharp, S.J.; Koulman, A.; Schulze, M.B.; Zheng, J.; Ye, Z.; Sluijs, I.; Guevara, M.; Huerta, J.M.; et al. Association of Plasma Phospholipid n-3 and n-6 Polyunsaturated Fatty Acids with Type 2 Diabetes: The EPIC-InterAct Case-Cohort Study. PLOS Med. 2016, 13, e1002094. [Google Scholar] [CrossRef] [Green Version]
- Zong, G.; Liu, G.; Willett, W.C.; Wanders, A.J.; Alssema, M.; Zock, P.L.; Hu, F.B.; Sun, Q. Associations Between Linoleic Acid Intake and Incident Type 2 Diabetes Among U.S. Men and Women. Diabetes Care 2019, 42, 1406–1413. [Google Scholar] [CrossRef]
- Demine, S.; Reddy, N.; Renard, P.; Raes, M.; Arnould, T. Unraveling biochemical pathways affected by mitochondrial dysfunctions using metabolomic approaches. Metabolites 2014, 4, 831–878. [Google Scholar] [CrossRef] [Green Version]
- Kelley, D.E.; He, J.; Menshikova, E.V.; Ritov, V.B. Dysfunction of mitochondria in human skeletal muscle in type 2 diabetes. Diabetes 2002, 51, 2944–2950. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.; Yoo, H.J.; Kim, M.; Kim, M.; Lee, J.H. Metabolomics identifies increases in the acylcarnitine profiles in the plasma of overweight subjects in response to mild weight loss: A randomized, controlled design study. Lipids Health Dis. 2018, 17, 237. [Google Scholar] [CrossRef] [Green Version]
- Aguer, C.; McCoin, C.S.; Knotts, T.A.; Thrush, A.B.; Ono-Moore, K.; McPherson, R.; Dent, R.; Hwang, D.H.; Adams, S.H.; Harper, M.-E. Acylcarnitines: Potential implications for skeletal muscle insulin resistance. FASEB J. 2015, 29, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Guasch-Ferré, M.; Ruiz-Canela, M.; Li, J.; Zheng, Y.; Bulló, M.; Wang, D.D.; Toledo, E.; Clish, C.; Corella, D.; Estruch, R.; et al. Plasma Acylcarnitines and Risk of Type 2 Diabetes in a Mediterranean Population at High Cardiovascular Risk. J. Clin. Endocrinol. Metab. 2019, 104, 1508–1519. [Google Scholar] [CrossRef] [PubMed]
- Gunther, S.H.; Khoo, C.M.; Tai, E.-S.; Sim, X.; Kovalik, J.-P.; Ching, J.; Lee, J.J.; van Dam, R.M. Serum acylcarnitines and amino acids and risk of type 2 diabetes in a multiethnic Asian population. BMJ Open Diabetes Res. Care 2020, 8, e001315. [Google Scholar] [CrossRef]
- Deniau, A.-L.; Mosset, P.; Le Bot, D.; Legrand, A.B. Which alkylglycerols from shark liver oil have anti-tumour activities? Biochimie 2011, 93, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Deniau, A.-L.; Mosset, P.; Pédrono, F.; Mitre, R.; Le Bot, D.; Legrand, A.B. Multiple beneficial health effects of natural alkylglycerols from shark liver oil. Mar. Drugs 2010, 8, 2175–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathew, M.; Mathew, S.; Kumar, K.N.A.; Anandan, R. Analgesic and anti-inflammatory activities of liver oils of four shark species from indian. J. Food Lipids 2008, 15, 470–487. [Google Scholar] [CrossRef]
- Park, H.; He, A.; Tan, M.; Johnson, J.M.; Dean, J.M.; Pietka, T.A.; Chen, Y.; Zhang, X.; Hsu, F.-F.; Razani, B.; et al. Peroxisome-derived lipids regulate adipose thermogenesis by mediating cold-induced mitochondrial fission. J. Clin. Investig. 2019, 129, 694–711. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Dilbaz, S.; Coßmann, J.; Hoang, A.C.; Diedrich, V.; Herwig, A.; Harauma, A.; Hoshi, Y.; Moriguchi, T.; Landgraf, K.; et al. Breast milk alkylglycerols sustain beige adipocytes through adipose tissue macrophages. J. Clin. Investig. 2019, 129, 2485–2499. [Google Scholar] [CrossRef] [Green Version]
- Karastergiou, K.; Smith, S.R.; Greenberg, A.S.; Fried, S.K. Sex differences in human adipose tissues—The biology of pear shape. Biol. Sex Differ. 2012, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Ralston, J.C.; Zulyniak, M.A.; Nielsen, D.E.; Clarke, S.; Badawi, A.; El-Sohemy, A.; Ma, D.W.L.; Mutch, D.M. Ethnic- and sex-specific associations between plasma fatty acids and markers of insulin resistance in healthy young adults. Nutr. Metab. 2013, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Rauschert, S.; Uhl, O.; Koletzko, B.; Mori, T.A.; Beilin, L.J.; Oddy, W.H.; Hellmuth, C. Sex differences in the association of phospholipids with components of the metabolic syndrome in young adults. Biol. Sex Differ. 2017, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- González-Granillo, M.; Helguero, L.A.; Alves, E.; Archer, A.; Savva, C.; Pedrelli, M.; Ahmed, O.; Li, X.; Domingues, M.R.; Parini, P.; et al. Sex-specific lipid molecular signatures in obesity-associated metabolic dysfunctions revealed by lipidomic characterization in ob/ob mouse. Biol. Sex Differ. 2019, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Hoene, M.; Li, J.; Häring, H.-U.; Weigert, C.; Xu, G.; Lehmann, R. The lipid profile of brown adipose tissue is sex-specific in mice. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2014, 1841, 1563–1570. [Google Scholar] [CrossRef] [Green Version]
- Bairey Merz, C.N.; Ramineni, T.; Leong, D. Sex-specific risk factors for cardiovascular disease in women-making cardiovascular disease real. Curr. Opin. Cardiol. 2018, 33, 500–505. [Google Scholar] [CrossRef]
- Vaccarezza, M.; Papa, V.; Milani, D.; Gonelli, A.; Secchiero, P.; Zauli, G.; Gemmati, D.; Tisato, V. Sex/Gender-Specific Imbalance in CVD: Could Physical Activity Help to Improve Clinical Outcome Targeting CVD Molecular Mechanisms in Women? Int. J. Mol. Sci. 2020, 21, 1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horst, R.T.; van den Munckhof, I.C.L.; Schraa, K.; Aguirre-Gamboa, R.; Jaeger, M.; Smeekens, S.P.; Brand, T.; Lemmers, H.; Dijkstra, H.; Galesloot, T.E.; et al. Sex-Specific Regulation of Inflammation and Metabolic Syndrome in Obesity. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1787–1800. [Google Scholar] [CrossRef] [PubMed]
- Steffen, A.; Sorensen, T.I.; Knuppel, S.; Travier, N.; Sanchez, M.J.; Huerta, J.M.; Quiros, J.R.; Ardanaz, E.; Dorronsoro, M.; Teucher, B.; et al. Development and validation of a risk score predicting substantial weight gain over 5 years in middle-aged European men and women. PLoS ONE 2013, 8, e67429. [Google Scholar]
- Ross, R.; Neeland, I.J.; Yamashita, S.; Shai, I.; Seidell, J.; Magni, P.; Santos, R.D.; Arsenault, B.; Cuevas, A.; Hu, F.B.; et al. Waist circumference as a vital sign in clinical practice: A Consensus Statement from the IAS and ICCR Working Group on Visceral Obesity. Nat. Rev. Endocrinol. 2020, 16, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Katzmarzyk, P.T.; Janssen, I.; Ross, R.; Church, T.S.; Blair, S.N. The Importance of Waist Circumference in the Definition of Metabolic Syndrome. Prospect. Anal. Mortal. Men 2006, 29, 404–409. [Google Scholar]
- Williams, E.D.; Tapp, R.J.; Magliano, D.J.; Shaw, J.E.; Zimmet, P.Z.; Oldenburg, B.F. Health behaviours, socioeconomic status and diabetes incidence: The Australian Diabetes Obesity and Lifestyle Study (AusDiab). Diabetologia 2010, 53, 2538–2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Australian Institute of Health and Wellfare (AIHW). The Active Australia Survey: A Guide and Manual for Implementation, Analysis and Reporting; AIHW: Canberra, Australia, 2003.
- Anuradha, S.; Dunstan, D.W.; Healy, G.N.; Shaw, J.E.; Zimmet, P.Z.; Wong, T.Y.; Owen, N. Physical Activity, Television Viewing Time, and Retinal Vascular Caliber. Med. Sci. Sports Exerc. 2011, 43, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Ireland, P.; Jolley, D.; Giles, G.; O’Dea, K.; Powles, J.; Rutishauser, I.; Wahlqvist, M.L.; Williams, J. Development of the Melbourne FFQ: A food frequency questionnaire for use in an Australian prospective study involving an ethnically diverse cohort. Asia Pac. J. Clin. Nutr. 1994, 3, 19–31. [Google Scholar]
- Briganti, E.M.; Shaw, J.E.; Chadban, S.J.; Zimmet, P.Z.; Welborn, T.A.; McNeil, J.J.; Atkins, R.C. Untreated hypertension among Australian adults: The 1999–2000 Australian Diabetes, Obesity and Lifestyle Study (AusDiab). Med. J. Aust. 2003, 179, 135–139. [Google Scholar] [CrossRef]
- Alshehry, Z.H.; Barlow, C.K.; Weir, J.M.; Zhou, Y.; McConville, M.J.; Meikle, P.J. An Efficient Single Phase Method for the Extraction of Plasma Lipids. Metabolites 2015, 5, 389–403. [Google Scholar] [CrossRef]
- Beyene, H.B.; Hamley, S.; Giles, C.; Huynh, K.; Smith, A.; Cinel, M.; Mellet, N.A.; Morales-Scholz, M.G.; Kloosterman, D.; Howlett, K.F.; et al. Mapping the Associations of the Plasma Lipidome with Insulin Resistance and Response to an Oral Glucose Tolerance Test. J. Clin. Endocrinol. Metab. 2020, 105, e1041–e1055. [Google Scholar] [CrossRef]
- Weir, J.M.; Wong, G.; Barlow, C.K.; Greeve, M.A.; Kowalczyk, A.; Almasy, L.; Comuzzie, A.G.; Mahaney, M.C.; Jowett, J.B.; Shaw, J.; et al. Plasma lipid profiling in a large population-based cohort. J. Lipid Res. 2013, 54, 2898–2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Society. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | n | Annualized BMI Change (kg/m2) (SD) | ^p Value | Annualized WC Change (cm) (SD) | ^p Value |
|---|---|---|---|---|---|
| Overall | 5849 | 0.16 (0.41) | 0.43 (1.30) | ||
| Sex | |||||
| Men | 2653 | 0.14 (0.38) | 3.0 × 10−4 | 0.33 (1.51) | 3.5 × 10−8 |
| Women | 3196 | 0.18 (0.40) | 0.51 (1.40) | ||
| Age group | |||||
| ≥55 | 2241 | 0.09 (0.34) | 2.2 × 10−16 | 0.29 (1.32) | 1.0 × 10−10 |
| <55 | 3608 | 0.20 (0.41) | 0.51 (1.26) | ||
| Education | |||||
| High school and below | 2178 | 0.17 (0.37) | 7.5 × 10−1 | 0.46 (1.31) | 3.9 × 10−1 |
| Certificate and diploma | 2544 | 0.16 (0.39) | 0.41 (1.30) | ||
| Bachelor’s degree and above | 1129 | 0.16 (0.37) | 0.42 (1.27) | ||
| BMI category * | |||||
| Normal | 2207 | 0.18 (0.30) | 8.2 × 10−3 | 0.49 (1.19) | 5.2 × 10−3 |
| Overweight | 2397 | 0.15 (0.38) | 0.39 (1.29) | ||
| Obese | 1245 | 0.16 (0.52) | 0.37 (1.47) | ||
| WC category # | |||||
| Low risk | 2336 | 0.18 (0.30) | 5.5 × 10−3 | 0.67 (1.12) | 6.5 × 10−34 |
| Moderate risk | 1529 | 0.15 (0.36) | 0.36 (1.23) | ||
| High risk | 2015 | 0.15 (0.49) | 0.19 (1.47) | ||
| Smoking | |||||
| Current smoker | 664 | 0.22 (0.55) | 2.9 × 10−5 | 0.54 (1.39) | 8.2 × 10−4 |
| Ex-smoker | 1694 | 0.13 (0.39) | 0.33 (1.29) | ||
| Non-smoker | 3401 | 0.17 (0.37) | 0.45 (1.28) | ||
| TV viewing time (minutes per week) | |||||
| Tertile 1 (less than 420) | 1960 | 0.20 (0.36) | 8.4 × 10−7 | 0.51 (1.28) | 3.0 × 10−4 |
| Tertile 2 (420–900) | 1938 | 0.15 (0.37) | 0.41 (1.26) | ||
| Tertile 3 (>900) | 1929 | 0.14 (0.42) | 0.36 (1.34) | ||
| Diabetes | |||||
| Yes | 327 | 0.06 (0.41) | 4.7 × 10−6 | 0.33 (1.24) | 8.0 × 10−2 |
| No | 5522 | 0.17 (0.38) | 0.44 (1.30) | ||
| Exercise status based on exercise time (min/week) | |||||
| Sedentary (zero min) | 909 | 0.17 (0.41) | 5.3 × 10−1 | 0.39 (1.28) | 5.5 × 10−1 |
| Insufficient (0‒150) | 1793 | 0.17 (0.39) | 0.45 (1.29) | ||
| Sufficient (over 150min) | 3127 | 0.16 (0.38) | 0.43 (1.27) | ||
| Total energy intake (KJ/day) | |||||
| Tertile 1 (<6430.5) | 1836 | 0.17 (0.40) | 8.0 × 10−2 | 0.51 (1.35) | 4.2 × 10−4 |
| Tertile 2 (6430.5–8671) | 1901 | 0.17 (0.39) | 0.42 (1.27) | ||
| Tertile 3 (>8671) | 1900 | 0.15 (0.37) | 0.36 (1.27) | ||
| Cholesterol (mmol/L) | |||||
| High (≥5.5) | 3292 | 0.14 (0.40) | 2.4 × 10−9 | 0.36 (1.28) | 8.3 × 10−7 |
| Low (<5.5) | 2589 | 0.19 (0.40) | 0.52 (1.27) | ||
| Triglycerides (mmol/L) | |||||
| High (≥2.0) | 1265 | 0.13 (0.40) | 6.2 × 10−5 | 0.33 (1.27) | 1.7 × 10−3 |
| Low (<2.0) | 4616 | 0.18 (0.39) | 0.46 (1.31) | ||
| HDL-C (mmol/L) | |||||
| High (≥1.0) | 5288 | 0.17 (0.40) | 6.7 × 10−1 | 0.44 (1.30) | 1.9 × 10−1 |
| Low (<1.0) | 592 | 0.16 (0.39) | 0.36 (1.26) | ||
| HbA1C (%) | |||||
| High (≥6.5) | 219 | 0.06 (0.39) | 1.0 × 10−4 | 0.29 (1.19) | 9.8 × 10−2 |
| Low (<6.5) | 5630 | 0.17 (0.39) | 0.43 (1.30) | ||
| HOMA2-B (%) | |||||
| Tertile 1 (<110.5) | 1743 | 0.14 (0.34) | 6.8 × 10−1 | 0.36 (1.21) | 1.5 × 10−1 |
| Tertile 2 (110.5–139.4) | 1743 | 0.15 (0.37) | 0.43 (1.31) | ||
| Tertile 3 (>139.4) | 1743 | 0.15 (0.41) | 0.44 (1.30) | ||
| Quintiles of WC Change Score | Age (Years) | Baseline WC cm Mean (SD) | Follow up WC cm Mean (SD) | Change in WC % (SD) | Change of >5% WC n (Relative Risk) | Risk > 5% WC Change Relative to Q1 (Odds Ratio, 95% CI) |
|---|---|---|---|---|---|---|
| Model 1 (Base model 1) | ||||||
| Q1 (N = 531) | 59.5 (12.3) | 109.5 (10.0) | 110.3 (11.9) | 0.7 (5.9) | 119 (0.22) | 1.0 (reference) |
| Q2 (N = 531) | 55.3 (12.0) | 100.8 (7.0) | 101.9 (8.7) | 1.1 (5.6) | 132 (0.25) | 1.1 (0.9–1.5) |
| Q3 (N = 531) | 52.3 (11.3) | 97.1 (6.6) | 98.4 (8.3) | 1.4 (5.6) | 131 (0.25) | 1.1 (0.8–1.5) |
| Q4 (N = 530) | 49.5 (10.6) | 92.2 (6.8) | 94.2 (8.6) | 2.3 (6.2) | 152 (0.29) | 1.4 (1.1–1.8) * |
| Q5 (N = 530) | 42.4 (10.9) | 86.5 (7.4) | 89.5 (8.7) | 3.3 (6.2) | 202 (0.38) | 2.1 (1.6–2.8) * |
| Model 2 (Model 1 + lipidomic score) | ||||||
| Q1 (N = 531) | 56.8 (12.2) | 107.0 (10.5) | 107.0 (12.5) | 0.19 (5.6) | 110 (0.26) | 1.0 (reference) |
| Q2 (N = 531) | 55.0 (12.2) | 101.0 (8.7) | 102.5 (10.0) | 1.2 (5.8) | 138 (0.26) | 1.3 (1.1–1.8) * |
| Q3 (N = 531) | 53.0 (12.3) | 97.0 (8.2) | 98.4 (9.9) | 1.4 (5.7) | 133 (0.25) | 1.3 (1.1–1.7) * |
| Q4 (N = 530) | 50.1 (11.8) | 93.1 (7.8) | 95.4 (9.2) | 2.5 (6.1) | 157 (0.30) | 1.6 (1.2–2.1) * |
| Q5 (N = 530) | 44.1 (11.5) | 88.0 (8.5) | 91.1 (9.5) | 3.6 (6.0) | 198 (0.37) | 2.3 (1.7–3.0) * |
| Quintiles of WC Change Score | Age (Years) | Baseline WC cm Mean (SD) | Follow up WC cm Mean (SD) | Change in WC (%) Mean (SD) | Change of >5% WC n (Relative Risk) | Risk > 5% WC Change Relative to Q1 (Odds Ratio, 95% CI) |
|---|---|---|---|---|---|---|
| Model 1 (Base model 1) | ||||||
| Q1 (N = 640) | 58.6 (12.1) | 103.0 (9.9) | 103.3 (12.2) | 0.3 (7.7) | 130 (0.24) | 1.0 (reference) |
| Q2 (N = 639) | 55.5 (11.8) | 90.2 (6.3) | 91.8 (9.1) | 1.8 (7.8) | 88 (0.25) | 1.6 (1.2–2.1) * |
| Q3 (N = 639) | 52.9 (11.2) | 83.1 (6.1) | 86.2 (9.2) | 3.7 (8.2) | 157 (0.36) | 2.3 (1.8–3.0) * |
| Q4 (N = 639) | 47.7 (10.3) | 77.5 (5.5) | 80.7 (7.8) | 4.3 (8.1) | 312 (0.42) | 2.7 (2.1–3.4) * |
| Q5 (N = 639) | 41.2 (9.0) | 70.9 (5.0) | 75.4 (7.3) | 6.5 (8.3) | 564 (0.50) | 3.7 (2.9–4.8) * |
| Model 2 (Model 1 + lipidomic score) | ||||||
| Q1 (N = 640) | 57.8 (11.9) | 99.9 (11.8) | 99.7 (13.7) | -0.23 (7.6) | 131 (0.20) | 1.0 (reference) |
| Q2 (N = 639) | 54.0 (12.4) | 89.4 (9.4) | 91.0 (11.4) | 1.8 (7.7) | 204 (0.32) | 1.8 (1.4–2.3) * |
| Q3 (N = 639) | 51.9 (11.8) | 83.1 (9.0) | 85.7 (11.1) | 3.1 (7.7) | 254 (0.40) | 2.6 (2.0–3.3) * |
| Q4 (N = 639) | 48.0 (11.2) | 79.0 (7.6) | 82.8 (10.0) | 4.8 (7.9) | 301 (0.47) | 3.5 (2.7–4.5) * |
| Q5 (N = 639) | 44.2 (10.6) | 73.2 (7.2) | 78.2 (9.0) | 7.2 (8.5) | 561 (0.56) | 5.4 (3.8–6.6) * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beyene, H.B.; Olshansky, G.; Giles, C.; Huynh, K.; Cinel, M.; Mellett, N.A.; Smith, A.A.T.; Shaw, J.E.; Magliano, D.J.; Meikle, P.J. Lipidomic Signatures of Changes in Adiposity: A Large Prospective Study of 5849 Adults from the Australian Diabetes, Obesity and Lifestyle Study. Metabolites 2021, 11, 646. https://doi.org/10.3390/metabo11090646
Beyene HB, Olshansky G, Giles C, Huynh K, Cinel M, Mellett NA, Smith AAT, Shaw JE, Magliano DJ, Meikle PJ. Lipidomic Signatures of Changes in Adiposity: A Large Prospective Study of 5849 Adults from the Australian Diabetes, Obesity and Lifestyle Study. Metabolites. 2021; 11(9):646. https://doi.org/10.3390/metabo11090646
Chicago/Turabian StyleBeyene, Habtamu B., Gavriel Olshansky, Corey Giles, Kevin Huynh, Michelle Cinel, Natalie A. Mellett, Adam Alexander T. Smith, Jonathan E. Shaw, Dianna J. Magliano, and Peter J. Meikle. 2021. "Lipidomic Signatures of Changes in Adiposity: A Large Prospective Study of 5849 Adults from the Australian Diabetes, Obesity and Lifestyle Study" Metabolites 11, no. 9: 646. https://doi.org/10.3390/metabo11090646
APA StyleBeyene, H. B., Olshansky, G., Giles, C., Huynh, K., Cinel, M., Mellett, N. A., Smith, A. A. T., Shaw, J. E., Magliano, D. J., & Meikle, P. J. (2021). Lipidomic Signatures of Changes in Adiposity: A Large Prospective Study of 5849 Adults from the Australian Diabetes, Obesity and Lifestyle Study. Metabolites, 11(9), 646. https://doi.org/10.3390/metabo11090646