


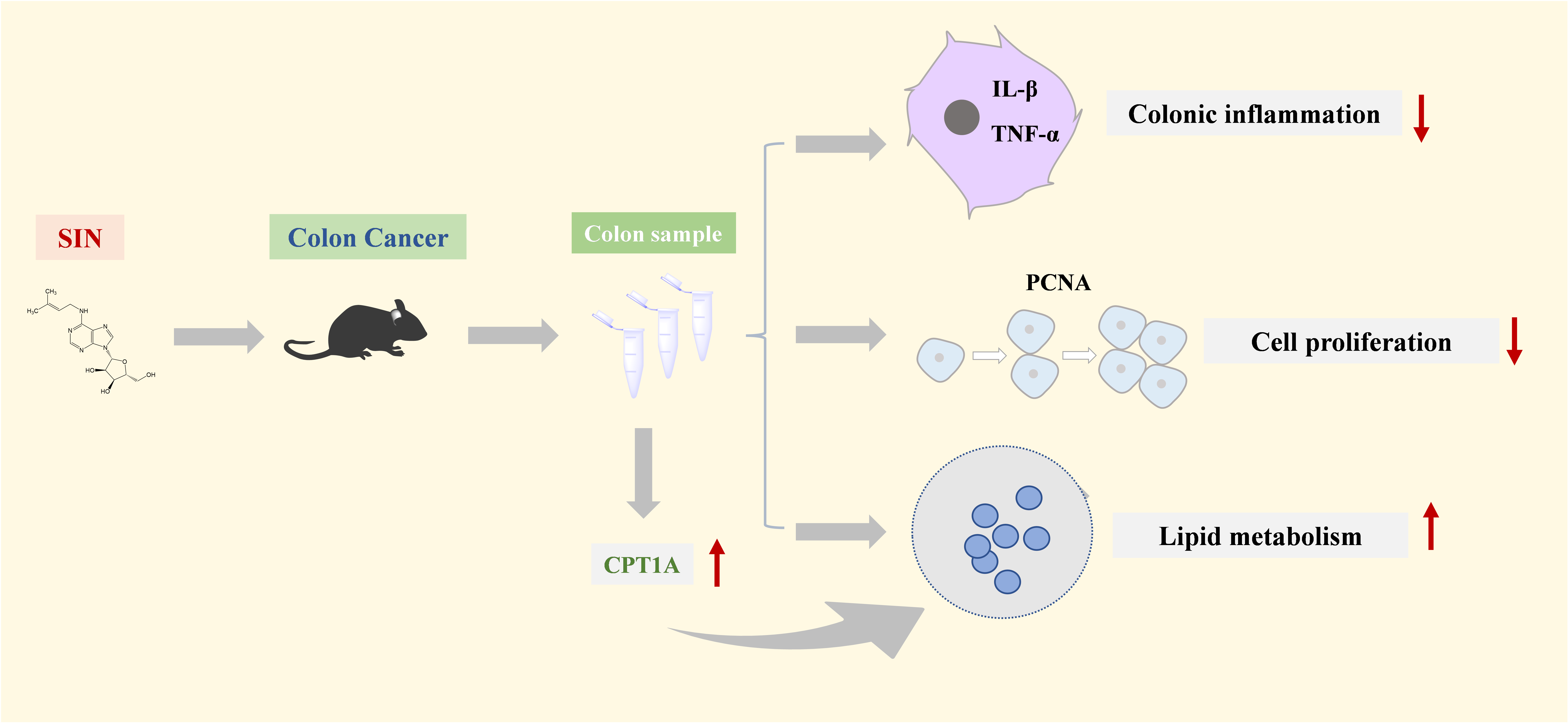
Sinomenine Ameliorates Colitis-Associated Cancer by Modulating Lipid Metabolism via Enhancing CPT1A Expression

Abstract

1. Introduction
2. Material and Methods
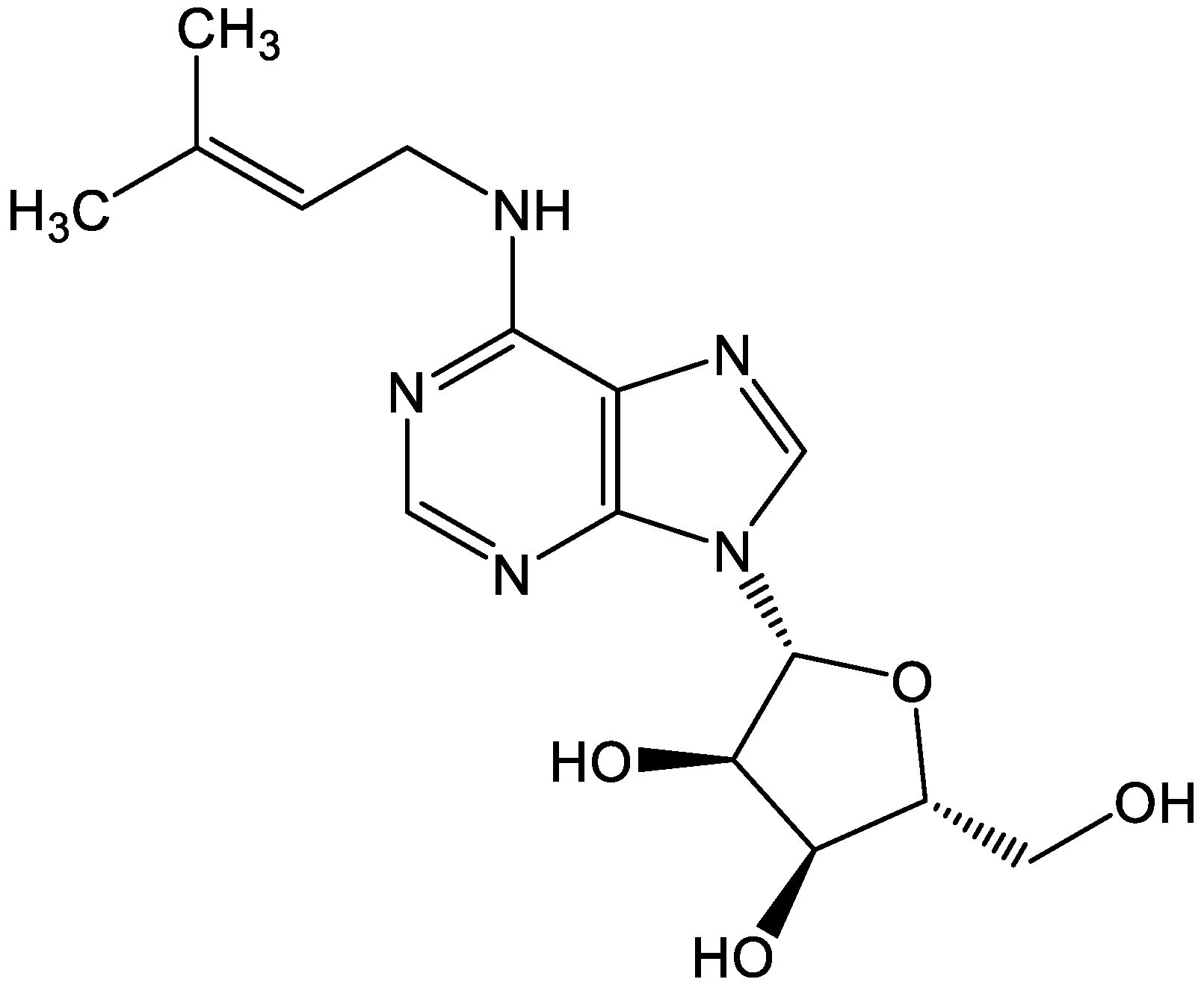
2.1. Chemicals and Reagents
2.2. Animals
2.3. Induction of CAC and Treatments
2.4. Colonic Histological Analysis
2.5. Colonic Immunohistochemical Analysis
2.6. Quantitative Real-Time PCR (qRT-PCR)
2.7. ELISA
2.8. Metabolomics Analysis of Serum Samples
2.8.1. Sample Pretreatment
2.8.2. UPLC-QTOF-MS/MS Conditions
2.8.3. Multivariate Data Analysis and Identification of the Differential Metabolites
2.9. Western Blotting
2.10. Cell Culture
2.11. MTT Assay
2.12. Statistical Analysis
3. Results
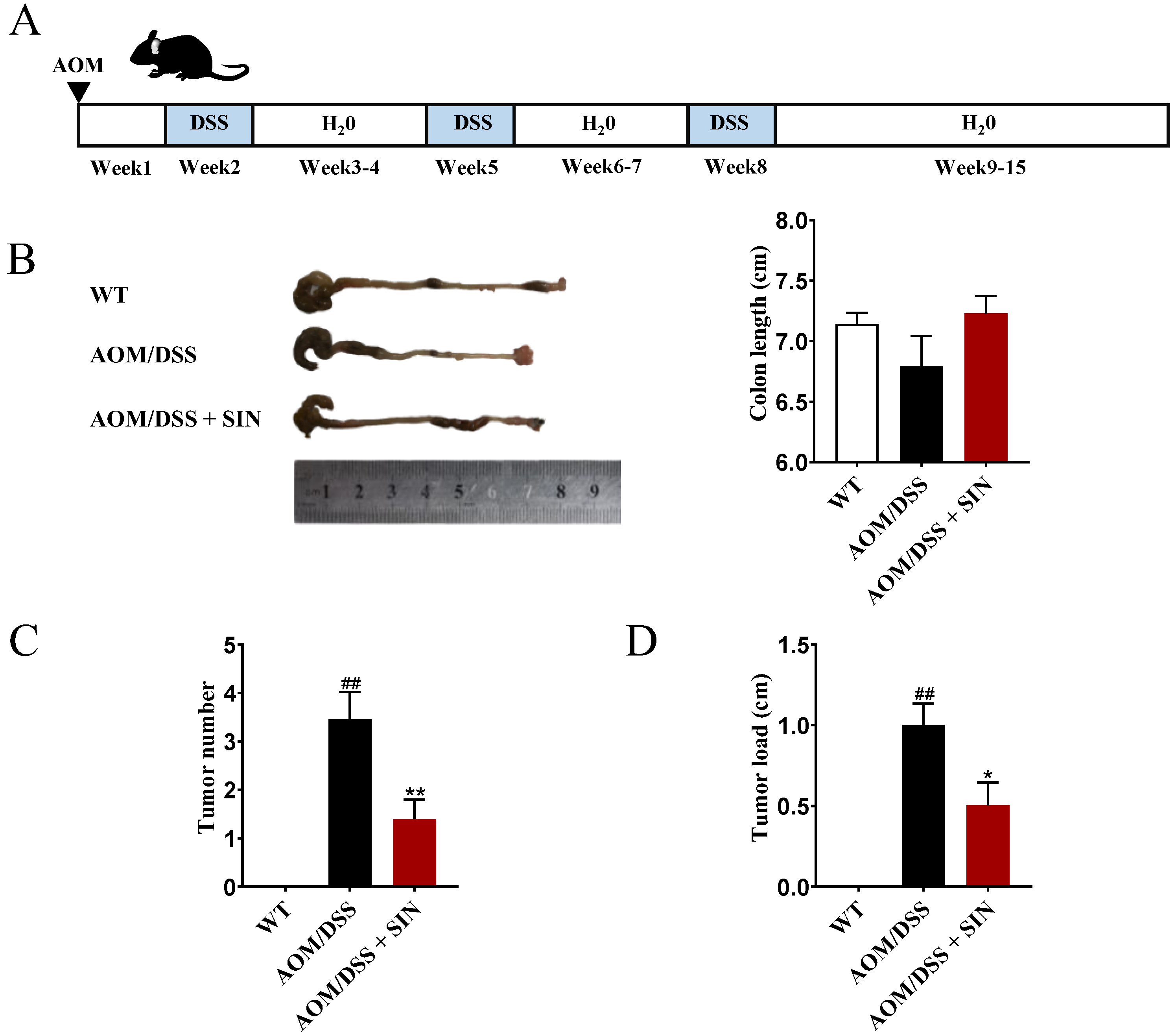
3.1. SIN Suppressed Colorectal Tumorigenesis in CAC Mice
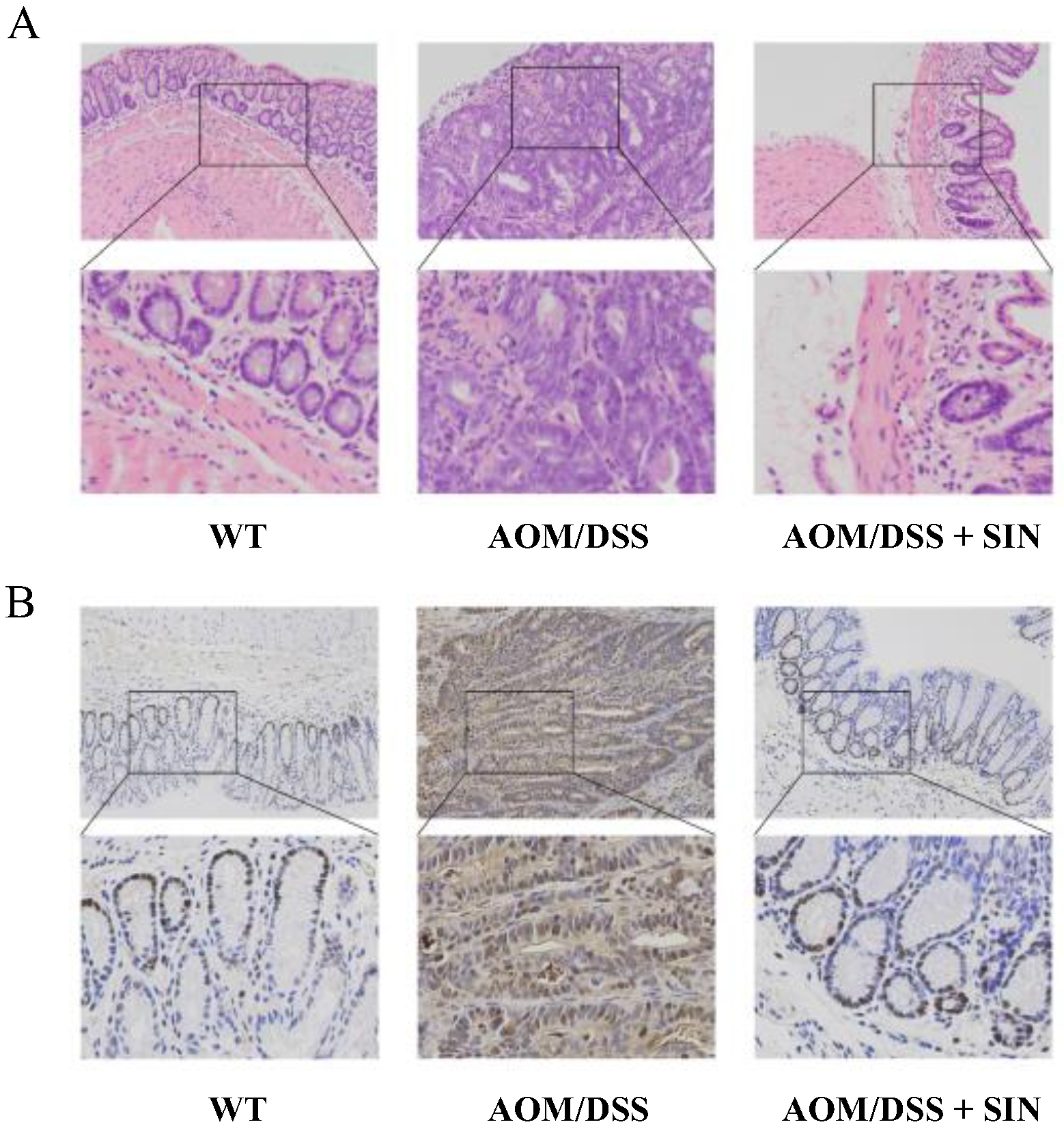
3.2. SIN Attenuated Histopathological Injury in CAC Mice
3.3. SIN Decreased the Number of PCNA+ in CAC Mice
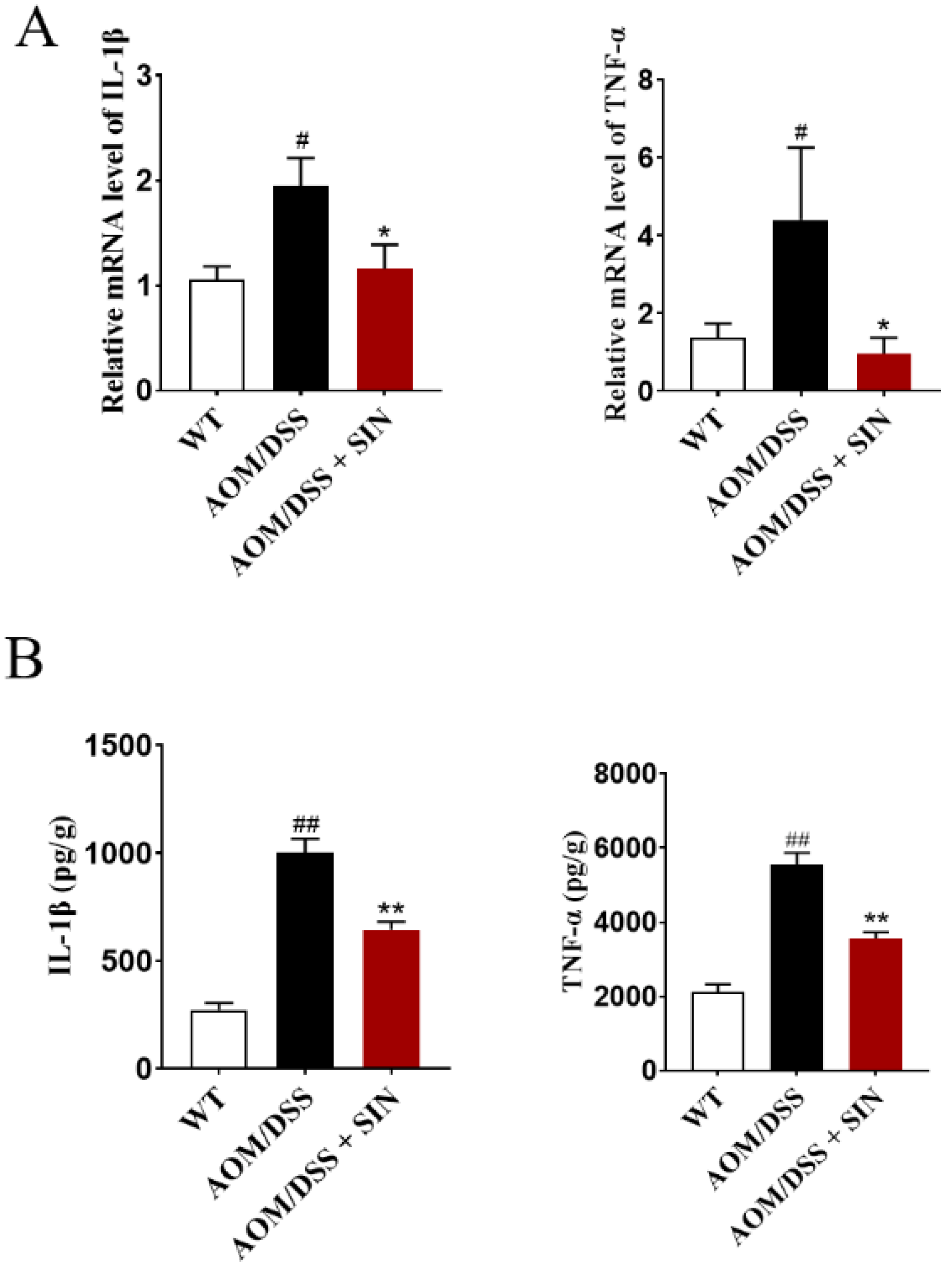
3.4. SIN Alleviated Colonic Inflammation in CAC Mice
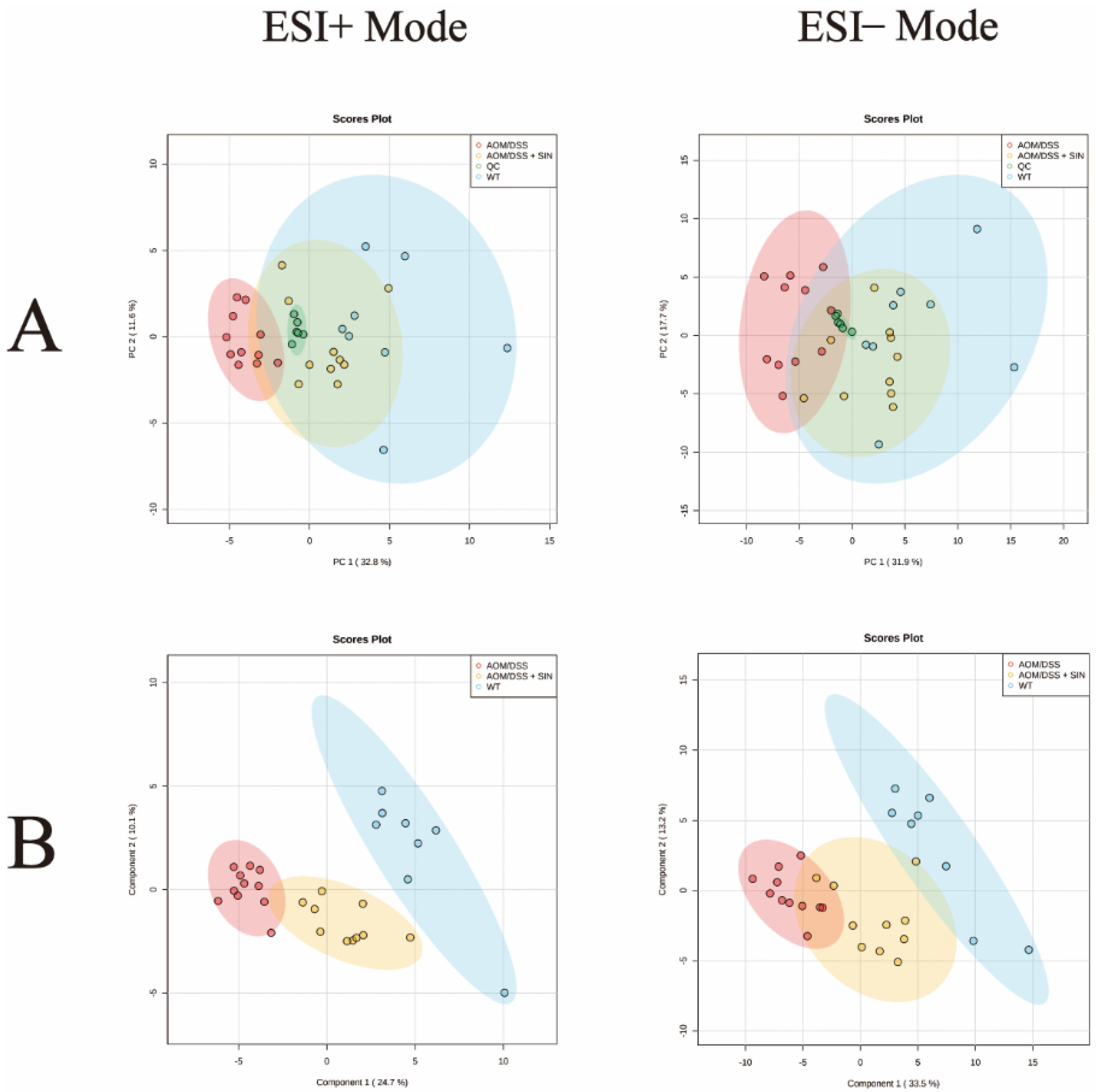
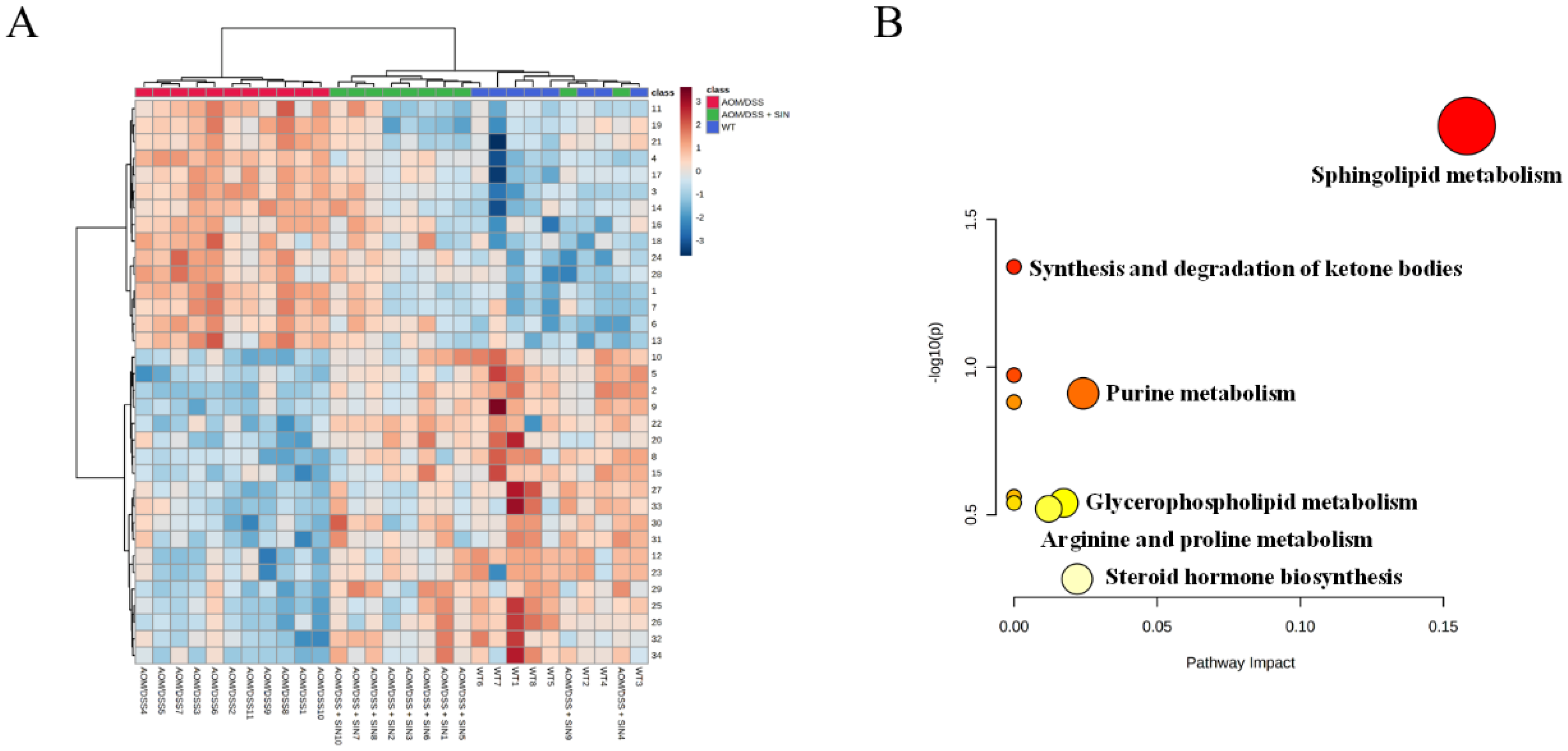
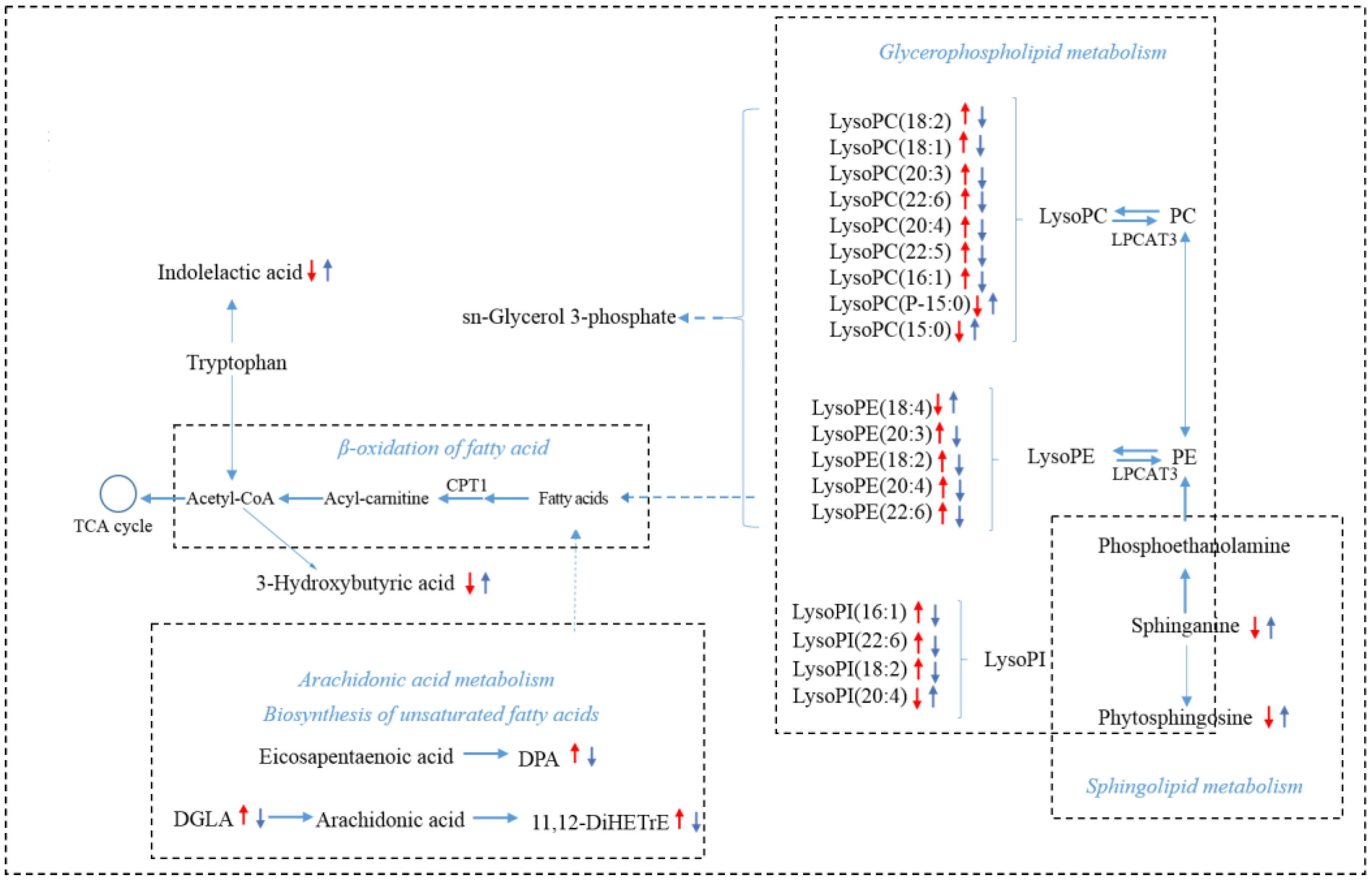
3.5. SIN Modulated Serum Metabolism in CAC Mice
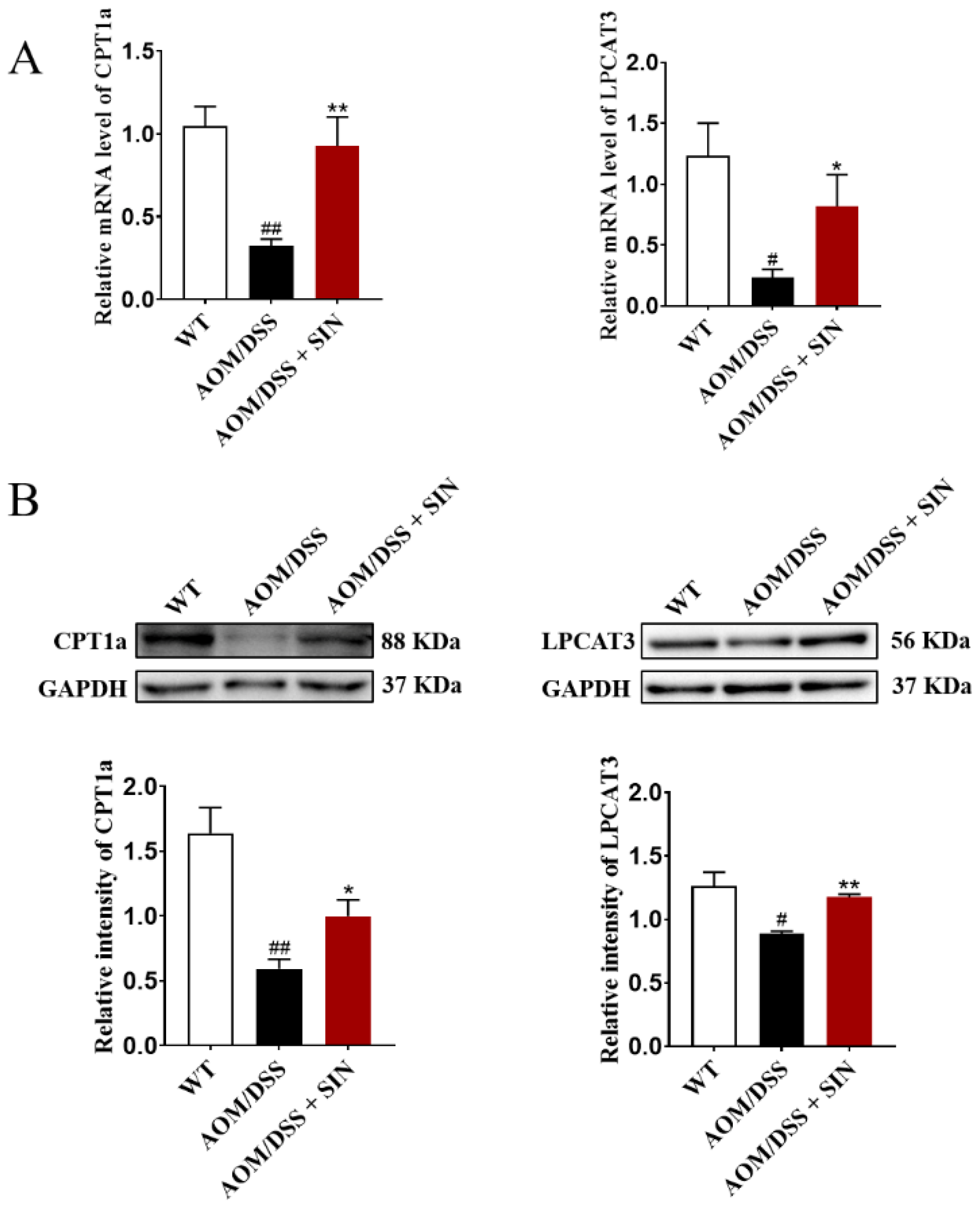
3.6. SIN Increased the Expression of CPT1A and LPCAT3 in Colon of CAC Mice
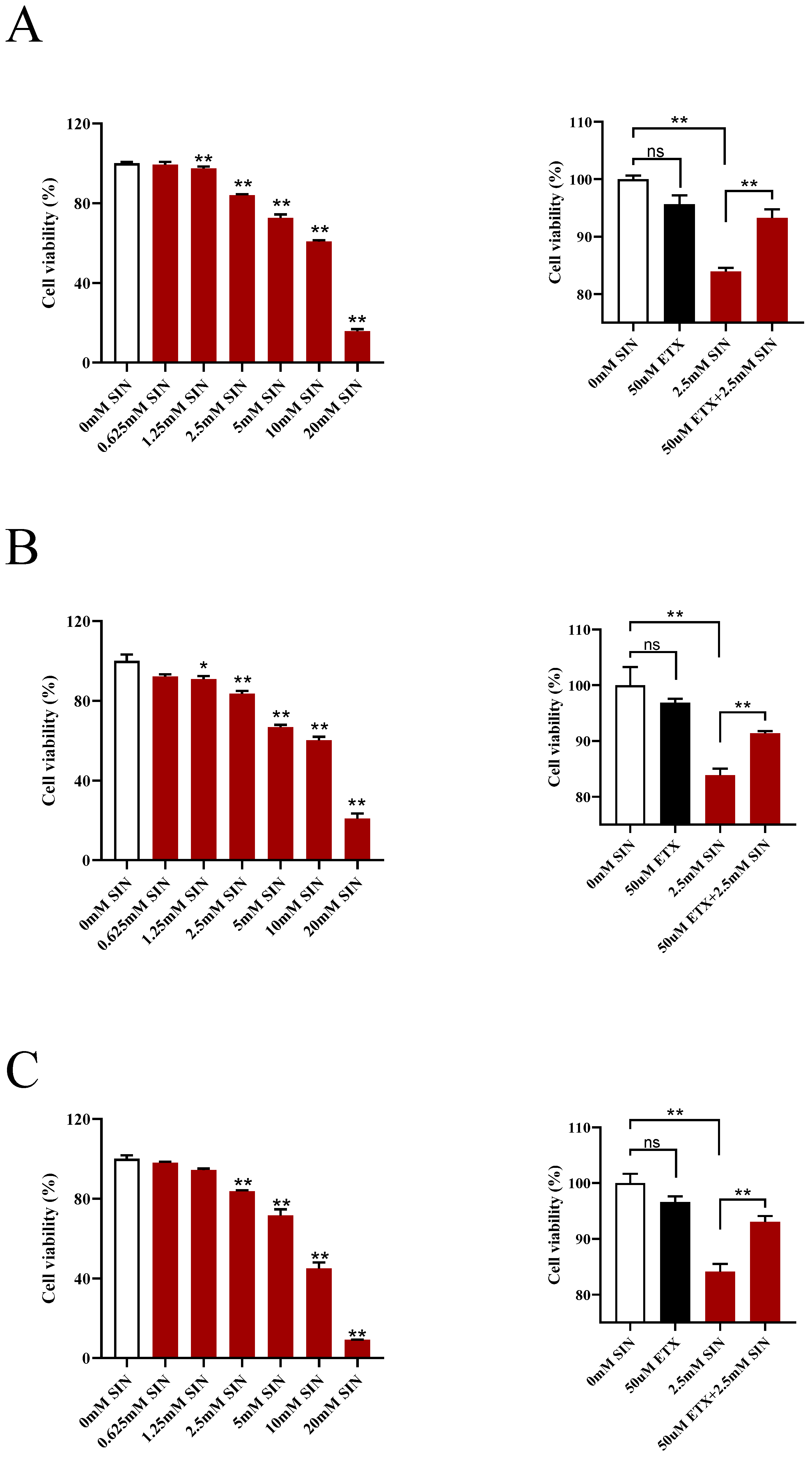
3.7. Inhibition of CPT1A Weakened the Antiproliferative Effect of SIN
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Biller, L.H.; Schrag, D. Diagnosis and treatment of metastatic colorectal cancer: A review. JAMA 2021, 325, 669–685. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef] [PubMed]
- Ringel, A.E.; Drijvers, J.M.; Baker, G.J.; Catozzi, A.; García-Cañaveras, J.C.; Gassaway, B.M.; Miller, B.C.; Juneja, V.R.; Nguyen, T.H.; Joshi, S.; et al. Obesity shapes metabolism in the tumor microenvironment to suppress anti-tumor immunity. Cell 2020, 183, 1848–1866. [Google Scholar] [CrossRef]
- Sinha, R. Colorectal cancer. Clin. Radiol. 2021, 76, 870. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Fontana, E.; Meyers, J.; Sobrero, A.; Iveson, T.; Shields, A.F.; Taieb, J.; Yoshino, T.; Souglakos, I.; Smyth, E.C.; Lordick, F.; et al. Early-onset colorectal adenocarcinoma in the IDEA database: Treatment adherence, toxicities, and outcomes with 3 and 6 months of adjuvant fluoropyrimidine and oxaliplatin. J. Clin. Oncol. 2021, 39, 4009–4019. [Google Scholar] [CrossRef] [PubMed]
- Tsilimigras, D.I.; Brodt, P.; Clavien, P.A.; Muschel, R.J.; D’Angelica, M.I.; Endo, I.; Parks, R.W.; Doyle, M.; Santibañes, E.; Pawlik, T.M. Liver metastases. Nat. Rev. Dis. Primers 2021, 7, 27. [Google Scholar] [CrossRef]
- Gao, L.N.; Zhong, B.; Wang, Y. Mechanism underlying antitumor effects of sinomenine. Chin. J. Integr. Med. 2019, 25, 873–878. [Google Scholar] [CrossRef]
- Hong, Y.; Yang, J.; Shen, X.; Zhu, H.F.; Sun, X.X.; Wen, X.; Bian, J.; Hu, H.Y.; Yuan, L.; Tao, J.; et al. Sinomenine hydrochloride enhancement of the inhibitory effects of anti-transferrin receptor antibody-dependent on the COX-2 pathway in human hepatoma cells. Cancer Immunol. Immunother. 2013, 62, 447–454. [Google Scholar] [CrossRef]
- Xie, T.; Ren, H.Y.; Lin, H.Q.; Mao, J.P.; Zhu, T.; Wang, S.D.; Ye, Z.M. Sinomenine prevents metastasis of human osteosarcoma cells via S phase arrest and suppression of tumor-related neovascularization and osteolysis through the CXCR4-STAT3 pathway. Int. J. Oncol. 2016, 48, 2098–2112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Zou, B.H.; Tan, Y.H.; Su, J.B.; Wang, Y.Y.; Xu, J.L.; Tao, L.; Zhou, H.; Liu, L.; Li, X.J. Sinomenine inhibits osteolysis in breast cancer by reducing IL-8/CXCR1 and c-Fos/NFATc1 signaling. Pharmacol. Res. 2019, 142, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.H.; Hung, J.H.; Liao, Y.C.; Tsai, S.T.; Wu, M.J.; Chen, P.S. Sinomenine inhibits migration and invasion of human lung cancer cell through downregulating expression of miR-21 and MMPs. Int. J. Mol. Sci. 2020, 21, 3080. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Feng, Q.H.; Li, M.J.; Su, J.Q.; Wang, P.Y.; Wang, X.; Yin, Y.C.; Wang, X.; Zhao, M.D. Sinomenine suppresses development of hepatocellular carcinoma cells via inhibiting MARCH1 and AMPK/STAT3 signaling pathway. Front. Mol. Biosci. 2021, 8, 684262. [Google Scholar] [CrossRef]
- Zheng, X.J.; Li, W.; Xu, H.L.; Liu, J.Y.; Ren, L.W.; Yang, Y.H.; Li, S.; Wang, J.H.; Ji, T.F.; Du, G.H. Sinomenine ester derivative inhibits glioblastoma by inducing mitochondria-dependent apoptosis and autophagy by PI3K/AKT/mTOR and AMPK/mTOR pathway. Acta Pharm. Sin. B 2021, 11, 3465–3480. [Google Scholar] [CrossRef]
- Zhang, J.X.; Yang, Z.R.; Wu, D.D.; Song, J.; Guo, X.F.; Wang, J.; Dong, W.G. Suppressive effect of sinomenine combined with 5-fluorouracil on colon carcinoma cell growth. Asian Pac. J. Cancer Prev. 2014, 15, 6737–6743. [Google Scholar] [CrossRef]
- Yang, H.B.; Yin, P.H.; Shi, Z.; Ma, Y.C.; Zhao, C.G.; Zheng, J.; Chen, T. Sinomenine, a COX-2 inhibitor, induces cell cycle arrest and inhibits growth of human colon carcinoma cells in vitro and in vivo. Oncol. Lett. 2016, 11, 411–418. [Google Scholar] [CrossRef]
- Yachida, S.; Mizutani, S.; Shiroma, H.; Shiba, S.; Nakajima, T.; Sakamoto, T.; Watanabe, H.; Masuda, K.; Nishimoto, Y.; Kubo, M.; et al. Metagenomic and metabolomic analyses reveal distinct stage-specific phenotypes of the gut microbiota in colorectal cancer. Nat. Med. 2019, 25, 968–976. [Google Scholar] [CrossRef]
- Vantaku, V.; Dong, J.R.; Ambati, C.R.; Perera, D.; Donepudi, S.R.; Amara, C.S.; Putluri, V.; Ravi, S.S.; Robertson, M.J.; Piyarathna, D.W.B.P.; et al. Multi-omics integration analysis robustly predicts high-grade patient survival and identifies CPT1B effect on fatty acid metabolism in bladder cancer. Clin. Cancer Res. 2019, 25, 3689–3701. [Google Scholar] [CrossRef]
- Husain, A.; Chiu, Y.T.; Sze, K.M.; Ho, D.W.; Tsui, Y.M.; Suarez, E.M.; Zhang, V.X.; Chan, L.K.; Lee, E.; Lee, J.M.; et al. Ephrin-A3/EphA2 axis regulates cellular metabolic plasticity to enhance cancer stemness in hypoxic hepatocellular carcinoma. J. Hepatol. 2022, S0168–S8278, 00125–00128. [Google Scholar] [CrossRef]
- Jia, W.; Xie, G.X.; Jia, W.P. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 111–128. [Google Scholar] [CrossRef]
- Ecker, J.; Benedetti, E.; Kindt, A.S.D.; Höring, M.; Perl, M.; Machmüller, A.C.; Sichler, A.; Plagge, J.; Wang, Y.T.; Zeissig, S.; et al. The colorectal cancer lipidome: Identification of a robust tumor-specific lipid species signature. Gastroenterology 2021, 161, 910–923. [Google Scholar] [CrossRef] [PubMed]
- Qiao, S.M.; Lv, C.J.; Tao, Y.; Miao, Y.M.; Zhu, Y.R.; Zhang, W.J.; Sun, D.D.; Yun, X.M.; Xia, Y.F.; Wei, Z.F.; et al. Arctigenin disrupts NLRP3 inflammasome assembly in colonic macrophages via downregulating fatty acid oxidation to prevent colitis-associated cancer. Cancer Lett. 2020, 491, 162–179. [Google Scholar] [CrossRef]
- McDonald, J.G.; Ejsing, C.S.; Kopczynski, D.; Holčapek, M.; Aoki, J.; Arita, M.; Arita, M.; Baker, E.S.; Bertrand-Michel, J.; Bowden, J.A.; et al. Introducing the Lipidomics Minimal Reporting Checklist. Nat. Metab. 2022, 8, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Popivanova, B.K.; Kitamura, K.; Wu, Y.; Kondo, T.; Kagaya, T.; Kaneko, S.; Oshima, M.; Fujii, C.; Mukaida, N. Blocking TNF-alpha in mice reduces colorectal carcinogenesis associated with chronic colitis. J. Clin. Investig. 2008, 118, 560–570. [Google Scholar] [PubMed]
- Ahechu, P.; Zozaya, G.; Martí, P.; Hernández-Lizoáin, J.L.; Baixauli, J.; Unamuno, X.; Frühbeck, G.; Catalán, V. NLRP3 inflammasome: A possible link between obesity-associated low-grade chronic inflammation and colorectal cancer development. Front. Immunol. 2018, 9, 2918. [Google Scholar] [CrossRef]
- Wang, B.; Rong, X.; Palladino, E.N.D.; Wang, J.F.; Fogelman, A.M.; Martín, M.G.; Alrefai, W.A.; Ford, D.A.; Tontonoz, P. Phospholipid remodeling and cholesterol availability regulate intestinal stemness and tumorigenesis. Cell Stem Cell 2018, 22, 206–220. [Google Scholar] [CrossRef]
- Peng, S.; Chen, D.; Cai, J.; Yuan, Z.; Huang, B.; Li, Y.; Wang, H.; Luo, Q.; Kuang, Y.; Liang, W.; et al. Enhancing cancer-associated fibroblast fatty acid catabolism within a metabolically challenging tumor microenvironment drives colon cancer peritoneal metastasis. Mol. Oncol. 2021, 15, 1391–1411. [Google Scholar] [CrossRef]
- Ni, T.; He, Z.H.; Dai, Y.Y.; Yao, J.Y.; Guo, Q.L.; Wei, L.B. Oroxylin A suppresses the development and growth of colorectal cancer through reprogram of HIF1α-modulated fatty acid metabolism. Cell Death Dis. 2017, 8, e2865. [Google Scholar] [CrossRef]
- Tong, B.; Yu, J.T.; Wang, T.; Dou, Y.N.; Wu, X.; Kong, L.Y.; Dai, Y.; Xia, Y.F. Sinomenine suppresses collagen-induced arthritis by reciprocal modulation of regulatory T cells and Th17 cells in gut-associated lymphoid tissues. Mol. Immunol. 2015, 65, 94–103. [Google Scholar] [CrossRef]
- Song, W.; Yang, X.T.; Wang, W.Q.; Wang, Z.; Wu, J.; Huang, F.J. Sinomenine ameliorates septic acute lung injury in mice by modulating gut homeostasis via aryl hydrocarbon receptor/Nrf2 pathway. Eur. J. Pharmacol. 2021, 912, 174581. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, S.; Gu, W.X.; Sun, X.; Wang, L.X.; Tang, L.M. Sinomenine hydrochloride ameliorates dextran sulfate sodium-induced colitis in mice by modulating the gut microbiota composition whilst suppressing the activation of the NLRP3 inflammasome. Exp. Ther. Med. 2021, 22, 1287. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Xia, B.; Guo, Q.S.; Zhang, L.; Wang, F.; Jiang, L.; Wang, Z.L.; Zhang, Y.X.; Li, C. Sinomenine attenuates 2, 4, 6-trinitrobenzene sulfonic acid-induced colitis in mice. Int. Immunopharmacol. 2007, 7, 604–611. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequences |
|---|---|---|
| IL-1β | Forward | 5′- TCGCAGCAGCACATCAACAAGAG -3′ |
| Reverse | 5′- AGGTCCACGGGAAAGACACAGG -3′ | |
| TNF-α | Forward | 5′- GCCTCTTCTCATTCCTGCTTGTGG -3′ |
| Reverse | 5′- GTGGTTTGTGAGTGTGAGGGTCTG -3′ | |
| CPT1A | Forward | 5′- GATGTGGACCTGCATTCCTT -3′ |
| Reverse | 5′- TCCTTGTAATGTGCGAGCTG -3′ | |
| LPCAT3 | Forward | 5′- GCTGCGGCTCATCTTCTCCATC -3′ |
| Reverse | 5′- AGGAACTGAAGCACGACACATAGC -3′ | |
| GAPDH | Forward | 5′- GGTTGTCTCCTGCGACTTCA -3′ |
| Reverse | 5′- TGGTCCAGGGTTTCTTACTCC -3′ |
| No. | RT (min) | Formula | Identification | Detected m/z | Change Trend (AOM/DSS vs. WT) | Change Trend (AOM/DSS + SIN vs. AOM/DSS) |
|---|---|---|---|---|---|---|
| 1 | 10.42 | C25H47O12P | LysoPI (16:1) | 571.2961 | ↑ *** | ↓ *** |
| 2 | 6.62 | C18H39NO3 | Phytosphingosine | 318.2997 | ↓ *** | ↑ *** |
| 3 | 10.2 | C26H50NO7P | LysoPC (18:2) | 520.3393 | ↑ *** | ↓ ** |
| 4 | 12.23 | C26H52NO7P | LysoPC (18:1) | 522.3552 | ↑ *** | ↓ *** |
| 5 | 8.99 | C23H40NO7P | LysoPE (18:4) | 474.2596 | ↓ *** | ↑ ** |
| 6 | 21.52 | C20H34O2 | Dihomo-gamma-linolenic acid | 307.262 | ↑ *** | ↓ * |
| 7 | 10.23 | C30H50NO7P | LysoPC (22:6) | 568.3392 | ↑ *** | ↓ ** |
| 8 | 6.43 | C21H30O4 | 21-Deoxycortisol | 347.2217 | ↓ *** | ↑ ** |
| 9 | 8 | C18H39NO2 | Sphinganine | 302.3051 | ↓ *** | ↑ *** |
| 10 | 0.49 | C4H9N3O2 | Creatine | 132.0766 | ↓ *** | ↑ ** |
| 11 | 10.23 | C28H50NO7P | LysoPC (20:4) | 544.339 | ↑ *** | ↓ ** |
| 12 | 0.82 | C10H12N4O5 | Inosine | 269.089 | ↓ *** | ↑ ** |
| 13 | 10.75 | C25H46NO7P | LysoPE (20:3) | 504.3049 | ↑ *** | ↓ ** |
| 14 | 9.87 | C23H44NO7P | LysoPE (18:2) | 478.2923 | ↑ *** | ↓ ** |
| 15 | 0.51 | C8H17NO2 | 2-Aminocaprylic acid | 160.1333 | ↓ *** | ↑ ** |
| 16 | 21.01 | C22H34O2 | Docosapentaenoic acid | 331.263 | ↑ *** | ↓ * |
| 17 | 11.11 | C30H52NO7P | LysoPC (22:5) | 570.3522 | ↑ *** | ↓ ** |
| 18 | 11.38 | C28H52NO7P | LysoPC (20:3) | 546.3551 | ↑ ** | ↓ * |
| 19 | 9.96 | C25H44NO7P | LysoPE (20:4) | 502.2921 | ↑ ** | ↓ ** |
| 20 | 4.23 | C10H11NO3 | Phenylacetylglycine | 194.0805 | ↓ ** | ↑ ** |
| 21 | 9.98 | C27H44NO7P | LysoPE (22:6) | 526.2919 | ↑ ** | ↓ ** |
| 22 | 20.52 | C29H49O12P | LysoPI (20:4) | 621.3089 | ↓ * | ↑ *** |
| 23 | 0.82 | C5H4N4O | Hypoxanthine | 137.0467 | ↓ * | ↑ ** |
| 24 | 10.08 | C31H49O12P | LysoPI (22:6) | 643.2888 | ↑ *** | ↓ * |
| 25 | 4.6 | C8H15NO3 | Acetyl-DL-Leucine | 172.0972 | ↓ *** | ↑ * |
| 26 | 4.65 | C11H13NO3 | Acetylphenylalanine | 206.0823 | ↓ *** | ↑ * |
| 27 | 4.065 | C6H12O3 | Leucinic acid | 131.0715 | ↓ *** | ↑ * |
| 28 | 9.95 | C27H49O12P | LysoPI (18:2) | 595.2885 | ↑ ** | ↓ * |
| 29 | 4.72 | C7H8O4S | p-Cresol sulfate | 187.0071 | ↓ ** | ↑ ** |
| 30 | 4.78 | C11H11NO3 | Indolelactic acid | 204.0668 | ↓ ** | ↑ * |
| 31 | 0.91 | C4H8O3 | 3-Hydroxybutyric acid | 103.0403 | ↓ ** | ↑ ** |
| 32 | 0.52 | C4H6N4O3 | Allantoin | 157.0369 | ↓ ** | ↑ ** |
| 33 | 4.14 | C8H7NO4S | Indoxyl sulfate | 212.0011 | ↓ * | ↑ * |
| 34 | 2.1152 | C5H10O3 | 2-Hydroxyvalerate | 117.0561 | ↓ * | ↑ * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Huang, D.; Dai, Y.; Xia, Y.-F. Sinomenine Ameliorates Colitis-Associated Cancer by Modulating Lipid Metabolism via Enhancing CPT1A Expression. Metabolites 2022, 12, 946. https://doi.org/10.3390/metabo12100946
Zhang J, Huang D, Dai Y, Xia Y-F. Sinomenine Ameliorates Colitis-Associated Cancer by Modulating Lipid Metabolism via Enhancing CPT1A Expression. Metabolites. 2022; 12(10):946. https://doi.org/10.3390/metabo12100946
Chicago/Turabian StyleZhang, Jing, Dan Huang, Yue Dai, and Yu-Feng Xia. 2022. "Sinomenine Ameliorates Colitis-Associated Cancer by Modulating Lipid Metabolism via Enhancing CPT1A Expression" Metabolites 12, no. 10: 946. https://doi.org/10.3390/metabo12100946
APA StyleZhang, J., Huang, D., Dai, Y., & Xia, Y.-F. (2022). Sinomenine Ameliorates Colitis-Associated Cancer by Modulating Lipid Metabolism via Enhancing CPT1A Expression. Metabolites, 12(10), 946. https://doi.org/10.3390/metabo12100946