
Integration of Multiplied Omics, a Step Forward in Systematic Dairy Research


Abstract
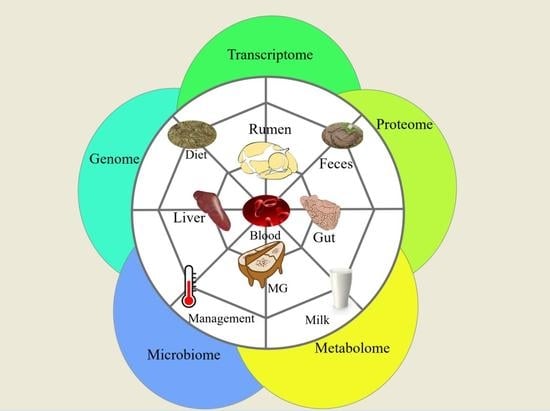
1. Introduction
2. Multi-Omics Studies in Lactation Physiology
3. Multi-Omics Methods for Reproduction Research
4. Multi-Omics Assists Feeding and Management
5. Multi-Omics Promotes Revealing Dairy Diseases
6. Conclusions and Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vailati-Riboni, M.; Palombo, V.; Loor, J.J. What Are Omics Sciences? In Periparturient Diseases of Dairy Cows, 1st ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–7. [Google Scholar]
- Anderson, N.L.; Anderson, N.G. Proteome and proteomics: New technologies, new concepts, and new words. Electrophoresis 1998, 19, 1853–1861. [Google Scholar] [CrossRef] [PubMed]
- Rinschen, M.M.; Ivanisevic, J.; Giera, M.; Siuzdak, G. Identification of bioactive metabolites using activity metabolomics. Nat. Rev. Mol. Cell Biol. 2019, 20, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Shao, W.; Huang, Z.; Tang, H.; Zhang, J.; Ding, Z.; Huang, K. MOGONET integrates multi-omics data using graph convolutional networks allowing patient classification and biomarker identification. Nat. Commun. 2021, 12, 3445. [Google Scholar] [CrossRef]
- Dimitrakopoulos, L.; Prassas, I.; Diamandis, E.P.; Charames, G.S. Onco-proteogenomics: Multi-omics level data integration for accurate phenotype prediction. Crit. Rev. Clin. Lab. Sci. 2017, 54, 414–432. [Google Scholar] [CrossRef] [PubMed]
- Malmuthuge, N.; Guan, L.L. Gut microbiome and omics: A new definition to ruminant production and health. Anim. Front. 2016, 6, 8–12. [Google Scholar] [CrossRef]
- Ametaj, B. A Systems Veterinary Approach in Understanding Transition Cow Diseases: Metabolomics. In Proceedings of the 4th International Symposium on Dairy Cow Nutrition and Milk Quality, Session 1, Advances in Fundamental Research, Beijing, China, 8–10 May 2015. [Google Scholar]
- Gu, F.F.; Liang, S.L.; Zhu, S.L.; Liu, J.X.; Sun, H.Z. Multi-omics revealed the effects of rumen-protected methionine on the nutrient profile of milk in dairy cows. Food Res. Int. 2021, 149, 110682. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.Y.; Sun, H.Z.; Wu, X.H.; Liu, J.X.; Guan, L.L. Multi-omics reveals that the rumen microbiome and its metabolome together with the host metabolome contribute to individualized dairy cow performance. Microbiome 2020, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Hess, M.; Sczyrba, A.; Egan, R.; Kim, T.W.; Chokhawala, H.; Schroth, G.; Luo, S.; Clark, D.S.; Chen, F.; Zhang, T.; et al. Metagenomic discovery of biomass-degrading genes and genomes from cow rumen. Science 2011, 331, 463–467. [Google Scholar] [CrossRef]
- Burd, N.A.; Hamer, H.M.; Pennings, B.; Pellikaan, W.F.; Senden, J.M.G.; Gijsen, A.P.; van Loon, L.J.C. Substantial Differences between Organ and Muscle Specific Tracer Incorporation Rates in a Lactating Dairy Cow. PLoS ONE 2013, 8, e68109. [Google Scholar] [CrossRef] [PubMed]
- Belanche, A.; Kingston-Smith, A.H.; Newbold, C.J. An Integrated Multi-Omics Approach Reveals the Effects of Supplementing Grass or Grass Hay with Vitamin E on the Rumen Microbiome and Its Function. Front. Microbiol. 2016, 7, 905. [Google Scholar] [CrossRef]
- Mamuad, L.L.; Lee, S.S.; Lee, S.S. Recent insight and future techniques to enhance rumen fermentation in dairy goats. Asian Austral. J. Anim. 2019, 32, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Pryce, J.E.; Wales, W.J.; de Haas, Y.; Veerkamp, R.F.; Hayes, B.J. Genomic selection for feed efficiency in dairy cattle. Animal 2014, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sigdel, A.; Abdollahi-Arpanahi, R.; Aguilar, I.; Peñagaricano, F. Whole Genome Mapping Reveals Novel Genes and Pathways Involved in Milk Production Under Heat Stress in US Holstein Cows. Front. Genet. 2019, 10, 928. [Google Scholar] [CrossRef]
- Tarekegn, G.M.; Strandberg, E.; Andonov, S.; Båge, R.; Ask-Gullstrand, P.; Rius-Vilarrasa, E.; Christensen, J.M.; Berglund, B. Single-step genome-wide association study uncovers known and novel candidate genomic regions for endocrine and classical fertility traits in Swedish Red and Holstein dairy cows. Livest. Sci. 2021, 253, 104731. [Google Scholar] [CrossRef]
- Feugang, J.M.; Rodriguez-Osorio, N.; Kaya, A.; Wang, H.; Page, G.; Ostermeier, G.C.; Topper, E.K.; Memili, E. Transcriptome analysis of bull spermatozoa: Implications for male fertility. Reprod. Biomed. Online 2010, 21, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, A.; Rincon, G.; Islas-Trejo, A.; Wickramasinghe, S.; Medrano, J.F. SNP discovery in the bovine milk transcriptome using RNA-Seq technology. Mamm. Genome 2010, 21, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Akbar, H.; Bionaz, M.; Carlson, D.B.; Rodriguez-Zas, S.L.; Everts, R.E.; Lewin, H.A.; Drackley, J.K.; Loor, J.J. Feed restriction, but not l-carnitine infusion, alters the liver transcriptome by inhibiting sterol synthesis and mitochondrial oxidative phosphorylation and increasing gluconeogenesis in mid-lactation dairy cows. J. Dairy Sci. 2013, 96, 2201–2213. [Google Scholar] [CrossRef] [PubMed]
- Fagerlind, M.; Stalhammar, H.; Olsson, B.; Klinga-Levan, K. Expression of miRNAs in Bull Spermatozoa Correlates with Fertility Rates. Reprod. Domest. Anim. 2015, 50, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liang, G.; Wang, B.; Sun, H.; Liu, J.; Guan, L.L. Systematic microRNAome profiling reveals the roles of microRNAs in milk protein metabolism and quality: Insights on low-quality forage utilization. Sci. Rep. 2016, 6, 21194. [Google Scholar] [CrossRef]
- Li, F.; Guan, L.L. Metatranscriptomic Profiling Reveals Linkages between the Active Rumen Microbiome and Feed Efficiency in Beef Cattle. Appl. Environ. Microb. 2017, 83, e00061-17. [Google Scholar] [CrossRef] [PubMed]
- Comtet-Marre, S.; Parisot, N.; Lepercq, P.; Chaucheyras-Durand, F.; Mosoni, P.; Peyretaillade, E.; Bayat, A.R.; Shingfield, K.J.; Peyret, P.; Forano, E. Metatranscriptomics Reveals the Active Bacterial and Eukaryotic Fibrolytic Communities in the Rumen of Dairy Cow Fed a Mixed Diet. Front. Microbiol. 2017, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Malmuthuge, N.; Steele, M.A.; Guan, L.L. Shift of hindgut microbiota and microbial short chain fatty acids profiles in dairy calves from birth to pre-weaning. FEMS Microbiol. Ecol. 2017, 94, fix179. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, D.; Wu, X.; Cai, J.; Liu, M.; Huang, X.; Wu, J.; Liu, J.; Guan, L. Effects of dietary physical or nutritional factors on morphology of rumen papillae and transcriptome changes in lactating dairy cows based on three different forage-based diets. BMC Genom. 2017, 18, 353. [Google Scholar] [CrossRef] [PubMed]
- Sollinger, A.; Tveit, A.T.; Poulsen, M.; Noel, S.J.; Bengtsson, M.; Bernhardt, J.; Hellwing, A.L.F.; Lund, P.; Riedel, K.; Schleper, C.; et al. Holistic Assessment of Rumen Microbiome Dynamics through Quantitative Metatranscriptomics Reveals Multifunctional Redundancy during Key Steps of Anaerobic Feed Degradation. Msystems 2018, 3, e00038-18. [Google Scholar] [CrossRef] [PubMed]
- Putz, E.J.; Putz, A.M.; Jeon, H.; Lippolis, J.D.; Ma, H.; Reinhardt, T.A.; Casas, E. MicroRNA profiles of dry secretions through the first three weeks of the dry period from Holstein cows. Sci. Rep. 2019, 9, 19658. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Gelsinger, S.; Edwards, A.; Riehle, C.; Koch, D. Changes in meta-transcriptome of rumen epimural microbial community and liver transcriptome in young calves with feed induced acidosis. Sci. Rep. 2019, 9, 18967. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Gelsinger, S.; Edwards, A.; Riehle, C.; Koch, D. Transcriptome analysis of rumen epithelium and meta-transcriptome analysis of rumen epimural microbial community in young calves with feed induced acidosis. Sci. Rep. 2019, 9, 4744. [Google Scholar] [CrossRef] [PubMed]
- Ogunade, I.; Pech-Cervantes, A.; Schweickart, H. Metatranscriptomic Analysis of Sub-Acute Ruminal Acidosis in Beef Cattle. Animals 2019, 9, 232. [Google Scholar] [CrossRef] [PubMed]
- Ametaj, B.N.; Zebeli, Q.; Saleem, F.; Psychogios, N.; Lewis, M.J.; Dunn, S.M.; Xia, J.G.; Wishart, D.S. Metabolomics reveals unhealthy alterations in rumen metabolism with increased proportion of cereal grain in the diet of dairy cows. Metabolomics 2010, 6, 583–594. [Google Scholar] [CrossRef]
- Zhang, R. Omics Based Approaches to Assess the Effects of Subacute Ruminal Acidosis on Rumen Microbiota Metabolism and Epithelial Function in Dairy Cows; Nanjing Agricultural University: Nanjing, China, 2015. [Google Scholar]
- Forde, N.; O’Gorman, A.; Whelan, H.; Duffy, P.; O’Hara, L.; Kelly, A.K.; Havlicek, V.; Besenfelder, U.; Brennan, L.; Lonergan, P. Lactation-induced changes in metabolic status and follicular-fluid metabolomic profile in postpartum dairy cows. Reprod. Fertil. Dev. 2016, 28, 1882. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.C.; Mudaliar, M.; Tassi, R.; McNeilly, T.N.; Burchmore, R.; Burgess, K.; Herzyk, P.; Zadoks, R.N.; Eckersall, P.D. Mastitomics, the integrated omics of bovine milk in an experimental model of Streptococcus uberis mastitis: 3. Untargeted metabolomics. Mol. Biosyst. 2016, 12, 2762–2769. [Google Scholar] [CrossRef] [PubMed]
- Alejandro, B.; Newbold, C.J.; Wanchang, L.; Pauline, R.S.; Kingston-Smith, A.H. A Systems Biology Approach Reveals Differences in the Dynamics of Colonization and Degradation of Grass vs. Hay by Rumen Microbes with Minor Effects of Vitamin E Supplementation. Front. Microbiol. 2017, 8, 1456. [Google Scholar]
- Humer, E.; Khol-Parisini, A.; Metzler-Zebeli, B.U.; Gruber, L.; Zebeli, Q. Alterations of the Lipid Metabolome in Dairy Cows Experiencing Excessive Lipolysis Early Postpartum. PLoS ONE 2016, 11, e0158633. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Wang, B.; Wang, J.; Liu, H.; Liu, J. Biomarker and pathway analyses of urine metabolomics in dairy cows when corn stover replaces alfalfa hay. J. Anim. Sci. Biotechnol. 2016, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Chen, Q.; Wang, Q.; White, R.R.; Liu, J.; Liu, H. Complementary transcriptomic and proteomic analyses reveal regulatory mechanisms of milk protein production in dairy cows consuming different forages. Sci. Rep. 2017, 7, 44234. [Google Scholar] [CrossRef] [PubMed]
- Artegoitia, V.M.; Foote, A.P.; Lewis, R.M.; Freetly, H.C. Rumen Fluid Metabolomics Analysis Associated with Feed Efficiency on Crossbred Steers. Sci. Rep. 2017, 7, 2864. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, H.; Fan, Y.; Guo, Y.; Zhang, G.; Nie, H.; Wang, F. Metabolomic profiling in umbilical venous plasma reveals effects of dietary rumen-protected arginine or N-carbamylglutamate supplementation in nutrient-restricted Hu sheep during pregnancy. Reprod. Domest. Anim. 2017, 52, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Murovec, B.; Makuc, D.; Repinc, S.K.; Prevorsek, Z.; Zavec, D.; Sket, R.; Pecnik, K.; Plavec, J.; Stres, B. 1H NMR metabolomics of microbial metabolites in the four MW agricultural biogas plant reactors: A case study of inhibition mirroring the acute rumen acidosis symptoms. J. Environ. Manag. 2018, 222, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Elolimy, A.; Alharthi, A.; Zeineldin, M.; Parys, C.; Helmbrecht, A.; Loor, J.J. Supply of Methionine During Late-Pregnancy Alters Fecal Microbiota and Metabolome in Neonatal Dairy Calves Without Changes in Daily Feed Intake. Front. Microbiol. 2019, 10, 2159. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.Z.; Zhou, M.; Wang, O.; Chen, Y.H.; Liu, J.X.; Guan, L.L. Multi-omics reveals functional genomic and metabolic mechanisms of milk production and quality in dairy cows. Bioinformatics 2020, 36, 2530–2537. [Google Scholar] [CrossRef]
- Zhang, H.; Tong, J.J.; Zhang, Y.H.; Xiong, B.H.; Jiang, L.S. Metabolomics reveals potential biomarkers in the rumen fluid of dairy cows with different levels of milk production. Asian-Australas. J. Anim. Sci. 2020, 33, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, B.A.; Powers, J.B.; Campagna, S.R.; Seay, T.B.; Embree, M.M.; Myer, P.R. Rumen fluid metabolomics of beef steers differing in feed efficiency. Metabolomics 2020, 16, 23. [Google Scholar] [CrossRef] [PubMed]
- Ogunade, I.M.; McCoun, M.; Idowu, M.D.; Peters, S.O. Comparative effects of two multispecies direct-fed microbial products on energy status, nutrient digestibility, and ruminal fermentation, bacterial community, and metabolome of beef steers. J. Anim. Sci. 2020, 98, skaa201. [Google Scholar] [CrossRef]
- Wang, C.; Liu, H.; Wang, C.; Liu, J.; Liu, H. Effects of Dietary Rumen-Protected Betaine on Lactation Performance and Serum Metabolites of Mid-lactation Holstein Dairy Cows. J. Agric. Food Chem. 2020, 68, 13154–13159. [Google Scholar] [CrossRef] [PubMed]
- Luke, T.D.W.; Pryce, J.E.; Elkins, A.C.; Wales, W.J.; Rochfort, S.J. Use of Large and Diverse Datasets for H-1 NMR Serum Metabolic Profiling of Early Lactation Dairy Cows. Metabolites 2020, 10, 180. [Google Scholar] [CrossRef] [PubMed]
- Lisuzzo, A.; Laghi, L.; Faillace, V.; Zhu, C.; Contiero, B.; Morgante, M.; Mazzotta, E.; Gianesella, M.; Fiore, E. Differences in the serum metabolome profile of dairy cows according to the BHB concentration revealed by proton nuclear magnetic resonance spectroscopy (1H-NMR). Sci. Rep. 2022, 12, 2525. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Sun, Z.Q.; Tu, Y.; Si, B.W.; Liu, Y.L.; Yang, L.; Luo, H.L.; Yu, Z. Untargeted metabolomic investigate milk and ruminal fluid of Holstein cows supplemented with Perilla frutescens leaf. Food Res. Int. 2021, 140, 110017. [Google Scholar] [CrossRef]
- Stergiadis, S.; Cabeza-Luna, I.; Mora-Ortiz, M.; Stewart, R.D.; Dewhurst, R.J.; Humphries, D.J.; Watson, M.; Roehe, R.; Auffret, M.D. Unravelling the Role of Rumen Microbial Communities, Genes, and Activities on Milk Fatty Acid Profile Using a Combination of Omics Approaches. Front. Microbiol. 2021, 11, 590441. [Google Scholar] [CrossRef]
- Wang, Y.; Nan, X.; Zhao, Y.; Wang, Y.; Jiang, L.; Xiong, B. Ruminal Degradation of Rumen-Protected Glucose Influences the Ruminal Microbiota and Metabolites in Early-Lactation Dairy Cows. Appl. Environ. Microbiol. 2021, 87, e01908-20. [Google Scholar] [CrossRef] [PubMed]
- Peddinti, D.; Nanduri, B.; Kaya, A.; Feugang, J.M.; Burgess, S.C.; Memili, E. Comprehensive proteomic analysis of bovine spermatozoa of varying fertility rates and identification of biomarkers associated with fertility. BMC Syst. Biol. 2008, 2, 19. [Google Scholar] [CrossRef] [PubMed]
- Ledgard, A.M.; Berg, M.C.; Mcmillan, W.H.; Smolenski, G.; Peterson, A.J. Effect of asynchronous transfer on bovine embryonic development and relationship with early cycle uterine proteome profiles. Reprod. Fertil. Dev. 2012, 24, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Saadi, H.A.S.; van Riemsdijk, E.; Dance, A.L.; Rajamanickam, G.D.; Kastelic, J.P.; Thundathil, J.C. Proteins associated with critical sperm functions and sperm head shape are differentially expressed in morphologically abnormal bovine sperm induced by scrotal insulation. J. Proteom. 2013, 82, 64–80. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Shen, J.S.; Ren, D.X.; Liu, J.X. Effects of the processing methods of corn grain and soybean meal on milk protein expression profiles in dairy cows. Animal 2015, 9, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.C.; Mullen, W.; Tassi, R.; Ramirez-Torres, A.; Mudaliar, M.; McNeilly, T.N.; Zadoks, R.N.; Burchmore, R.; Eckersall, P.D. Mastitomics, the integrated omics of bovine milk in an experimental model of Streptococcus uberis mastitis: 1. High abundance proteins, acute phase proteins and peptidomics. Mol. Biosyst. 2016, 12, 2735–2747. [Google Scholar] [CrossRef]
- Zachut, M.; Sood, P.; Livshitz, L.; Kra, G.; Levin, Y.; Moallem, U. Proteome dataset of pre-ovulatory follicular fluids from less fertile dairy cows. Data Brief 2016, 7, 1515–1518. [Google Scholar] [CrossRef][Green Version]
- Mudaliar, M.; Tassi, R.; Thomas, F.C.; McNeilly, T.N.; Weidt, S.K.; McLaughlin, M.; Wilson, D.; Burchmore, R.; Herzyk, P.; Eckersall, P.D.; et al. Mastitomics, the integrated omics of bovine milk in an experimental model of Streptococcus uberis mastitis: 2. Label-free relative quantitative proteomics. Mol. Biosyst. 2016, 12, 2748–2761. [Google Scholar] [CrossRef] [PubMed]
- Snelling, T.J.; Wallace, R.J. The rumen microbial metaproteome as revealed by SDS-PAGE. BMC Microbiol. 2017, 17, 9. [Google Scholar] [CrossRef]
- Skibiel, A.L.; Zachut, M.; do Amaral, B.C.; Levin, Y.; Dahl, G.E. Liver proteomic analysis of postpartum Holstein cows exposed to heat stress or cooling conditions during the dry period. J. Dairy Sci. 2018, 101, 705–716. [Google Scholar] [CrossRef]
- Veshkini, A.; Bonnet, M.; Vogel, L.; Trscher, A.; Sauerwein, H. liver proteomics of dairy cows supplied with essential fatty acids and conjugated linoleic acids. In Proceedings of the 71st Annual Meeting of the European Federation of Animal Science, Porto, Portugal, 31 August–4 September 2020. [Google Scholar]
- Morgavi, D.P.; Kelly, W.J.; Janssen, P.H.; Attwood, G.T. Rumen microbial (meta)genomics and its application to ruminant production. Animal 2012, 7, 184–201. [Google Scholar] [CrossRef] [PubMed]
- Tapio, I.; Fischer, D.; Blasco, L.; Tapio, M.; Wallace, R.J.; Bayat, A.R.; Ventto, L.; Kahala, M.; Negussie, E.; Shingfield, K.J.; et al. Taxon abundance, diversity, co-occurrence and network analysis of the ruminal microbiota in response to dietary changes in dairy cows. PLoS ONE 2017, 12, e0180260. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Ruiz, D.R.; Abecia, L.; Newbold, C.J. Manipulating rumen microbiome and fermentation through interventions during early life: A review. Front. Microbiol. 2015, 6, 1133. [Google Scholar] [CrossRef] [PubMed]
- Huws, S.A.; Creevey, C.J.; Oyama, L.B.; Mizrahi, I.; Denman, S.E.; Popova, M.; Munoz-Tamayo, R.; Forano, E.; Waters, S.M.; Hess, M.; et al. Addressing Global Ruminant Agricultural Challenges Through Understanding the Rumen Microbiome: Past, Present, and Future. Front. Microbiol. 2018, 9, 2161. [Google Scholar] [CrossRef]
- Zhang, Y.; Choi, S.H.; Nogoy, K.M.; Liang, S. Review: The development of the gastrointestinal tract microbiota and intervention in neonatal ruminants. Animal 2021, 15, 100316. [Google Scholar] [CrossRef] [PubMed]
- Mavrommatis, A.; Skliros, D.; Flemetakis, E.; Tsiplakou, E. Changes in the Rumen Bacteriome Structure and Enzymatic Activities of Goats in Response to Dietary Supplementation with Schizochytrium spp. Microorganisms 2021, 9, 1528. [Google Scholar] [CrossRef]
- Shabat, S.K.B.; Sasson, G.; Doron-Faigenboim, A.; Durman, T.; Yaacoby, S.; Berg Miller, M.E.; White, B.A.; Shterzer, N.; Mizrahi, I. Specific microbiome-dependent mechanisms underlie the energy harvest efficiency of ruminants. ISME J. 2016, 10, 2958–2972. [Google Scholar] [CrossRef]
- Gruninger, R.J.; Ribeiro, G.O.; Cameron, A.; McAllister, T.A. Invited review: Application of meta-omics to understand the dynamic nature of the rumen microbiome and how it responds to diet in ruminants. Animal 2019, 13, 1843–1854. [Google Scholar] [CrossRef] [PubMed]
- Alves, K.L.G.C.; Granja-Salcedo, Y.T.; Messana, J.D.; de Souza, V.C.; Ganga, M.J.G.; Colovate, P.H.D.; Kishi, L.T.; Berchielli, T.T. Rumen bacterial diversity in relation to nitrogen retention in beef cattle. Anaerobe 2021, 67, 102316. [Google Scholar] [CrossRef] [PubMed]
- Guyader, J.; Eugene, M.; Noziere, P.; Morgavi, D.P.; Doreau, M.; Martin, C. Influence of rumen protozoa on methane emission in ruminants: A meta-analysis approach. Animal 2014, 8, 1816–1825. [Google Scholar] [CrossRef] [PubMed]
- Newbold, C.J.; de la Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The Role of Ciliate Protozoa in the Rumen. Front. Microbiol. 2015, 6, 1313. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, J.; Tamminga, S. Simulation of the Effects of Diet on the Contribution of Rumen Protozoa to Degradation of Fiber in the Rumen. Br. J. Nutr. 1995, 74, 617–634. [Google Scholar] [CrossRef] [PubMed]
- Belanche, A.; de la Fuente, G.; Newbold, C.J. Effect of progressive inoculation of fauna-free sheep with holotrich protozoa and total-fauna on rumen fermentation, microbial diversity and methane emissions. FEMS Microbiol. Ecol. 2015, 91, fiu026. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.W. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation. J. Anim. Sci. 1995, 2804–2819. [Google Scholar] [CrossRef]
- Drackley, J.K. Biology of dairy cows during the transition period: The final frontier? J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef]
- Fiore, E.; Blasi, F.; Morgante, M.; Cossignani, L.; Badon, T.; Gianesella, M.; Contiero, B.; Berlanda, M. Changes of milk fatty acid composition in four lipid classes as biomarkers for the diagnosis of bovine ketosis using bioanalytical Thin Layer Chromatography and Gas Chromatographic techniques (TLC-GC). J. Pharmaceut. Biomed. 2020, 188, 113372. [Google Scholar] [CrossRef] [PubMed]
- Fiore, E.; Lisuzzo, A.; Tessari, R.; Spissu, N.; Moscati, L.; Morgante, M.; Gianesella, M.; Badon, T.; Mazzotta, E.; Berlanda, M.; et al. Milk Fatty Acids Composition Changes According to β-Hydroxybutyrate Concentrations in Ewes during Early Lactation. Animals 2021, 11, 1371. [Google Scholar] [CrossRef] [PubMed]
- Lisuzzo, A.; Bonelli, F.; Sgorbini, M.; Nocera, I.; Cento, G.; Mazzotta, E.; Turini, L.; Martini, M.; Salari, F.; Morgante, M.; et al. Differences of the Plasma Total Lipid Fraction from Pre-Foaling to Post-Foaling Period in Donkeys. Animals 2022, 12, 304. [Google Scholar] [CrossRef]
- Bobe, G.; Young, J.W.; Beitz, D.C. Invited review: Pathology, etiology, prevention, and treatment of fatty liver in dairy cows. J. Dairy Sci. 2004, 87, 3105–3124. [Google Scholar] [CrossRef]
- Ametaj, B.; Ametaj, B.N.; Herold. Periparturient Diseases of Dairy Cows; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Gerspach, C.; Imhasly, S.; Gubler, M.; Naegeli, H.; Ruetten, M.; Laczko, E. Altered plasma lipidome profile of dairy cows with fatty liver disease. Res. Vet. Sci. 2017, 110, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Carlson, D.B.; McFadden, J.W.; D’Angelo, A.; Woodworth, J.C.; Drackley, J.K. Dietary L-carnitine affects periparturient nutrient metabolism and lactation in multiparous cows. J. Dairy Sci. 2007, 90, 3422–3441. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Zinder, M.; Donthu, R.; Larkin, D.M.; Kumar, C.G.; Rodriguez-Zas, S.L.; Andropolis, K.E.; Oliveira, R.; Lewin, H.A. Multisite haplotype on cattle chromosome 3 is associated with quantitative trait locus effects on lactation traits. Physiol. Genom. 2011, 43, 1185–1197. [Google Scholar] [CrossRef] [PubMed]
- Colombani, C.; Legarra, A.; Fritz, S.; Guillaume, F.; Croiseau, P.; Ducrocq, V.; Robert-Granie, C. Application of Bayesian least absolute shrinkage and selection operator (LASSO) and BayesCpi methods for genomic selection in French Holstein and Montbeliarde breeds. J. Dairy Sci. 2013, 96, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Roche, J.R.; Burke, C.R.; Crookenden, M.A.; Heiser, A.; Loor, J.L.; Meier, S.; Mitchell, M.D.; Phyn, C.V.C.; Turner, S.A. Fertility and the transition dairy cow. Reprod. Fertil. Dev. 2018, 30, 85. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, S. Assessing the Association of the Level of Milk Production with Reproductive Performance in Dairy Cattle. J. Reprod. Dev. 2010, 56, S1–S7. [Google Scholar] [CrossRef] [PubMed]
- Aranciaga, N.; Morton, J.D.; Berg, D.K.; Gathercole, J.L. Proteomics and metabolomics in cow fertility: A systematic review. Reproduction 2020, 160, 639–658. [Google Scholar] [CrossRef] [PubMed]
- Lima, F.S.; Silvestre, F.T.; Penagaricano, F.; Thatcher, W.W. Early genomic prediction of daughter pregnancy rate is associated with improved fertility outcomes in Holstein dairy cows. J. Dairy Sci. 2019, 102, 114. [Google Scholar]
- Pacheco, H.A.; Rezende, F.M.; Peñagaricano, F. Gene mapping and genomic prediction of bull fertility using sex chromosome markers. J. Dairy Sci. 2020, 103, 3304–3311. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Kwon, W.S.; Oh, S.A.; Pang, M.G. Fertility-Related Proteomic Profiling Bull Spermatozoa Separated by Percoll. J. Proteome Res. 2012, 11, 4162–4168. [Google Scholar] [CrossRef]
- Soggiu, A.; Piras, C.; Hussein, H.A.; De Canio, M.; Gaviraghi, A.; Galli, A.; Urbani, A.; Bonizzia, L.; Roncada, P. Unravelling the bull fertility proteome. Mol. Biosyst. 2013, 9, 1188–1195. [Google Scholar] [CrossRef]
- Govindaraju, A.; Dogan, S.; Rodriguez-Osorio, N.; Grant, K.; Kaya, A.; Memili, E. Delivering value from sperm proteomics for fertility. Cell Tissue Res. 2012, 349, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Govindaraju, A.; Uzun, A.; Robertson, L.; Atli, M.O.; Kaya, A.; Topper, E.; Crate, E.A.; Padbury, J.; Perkins, A.; Memili, E. Dynamics of microRNAs in bull spermatozoa. Reprod. Biol. Endocrin. 2012, 10, 82. [Google Scholar] [CrossRef]
- Gilbert, R.O. Symposium review: Mechanisms of disruption of fertility by infectious diseases of the reproductive tract. J. Dairy Sci. 2019, 102, 3754–3765. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, S.; Elia, G.; O’Boyle, P.; Dunn, M.; Morris, D. Composition of the bovine uterine proteome is associated with stage of cycle and concentration of systemic progesterone. Proteomics 2013, 13, 3333–3353. [Google Scholar] [CrossRef] [PubMed]
- Forde, N.; Simintiras, C.A.; Sturmey, R.; Mamo, S.; Kelly, A.K.; Spencer, T.E.; Bazer, F.W.; Lonergan, P. Amino Acids in the Uterine Luminal Fluid Reflects the Temporal Changes in Transporter Expression in the Endometrium and Conceptus during Early Pregnancy in Cattle. PLoS ONE 2014, 9, e100010. [Google Scholar] [CrossRef] [PubMed]
- Macklon, N.S.; van der Gaast, M.H.; Hamilton, A.; Fauser, B.C.J.M.; Gludice, L.C. The impact of ovarian stimulation with recombinant FSH in combination with GnRH antagonist on the endometrial transcriptome in the window of implantation. Reprod. Sci. 2008, 15, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, T.M.; Goncalves, R.F.; Melo, C.F.O.R.; de Oliveira, D.N.; Lima, E.D.; Visintin, J.A.; de Achilles, M.A.; Catharino, R.R. A Metabolomic Overview of Follicular Fluid in Cows. Front. Vet. Sci. 2018, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Brito, L.F.; Oliveira, H.R.; Houlahan, K.; Fonseca, P.A.S.; Lam, S.; Butty, A.M.; Seymour, D.J.; Vargas, G.; Chud, T.C.S.; Silva, F.F.; et al. Genetic mechanisms underlying feed utilization and implementation of genomic selection for improved feed efficiency in dairy cattle. Can. J. Anim. Sci. 2020, 100, 587–604. [Google Scholar] [CrossRef]
- Grummer, R.R. Impact of Changes in Organic Nutrient Metabolism on Feeding the Transition Dairy-Cow. J. Anim. Sci. 1995, 73, 2820–2833. [Google Scholar] [CrossRef] [PubMed]
- Redfern, E.A.; Sinclair, L.A.; Robinson, P.A. Dairy cow health and management in the transition period: The need to understand the human dimension. Res. Vet. Sci. 2021, 137, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Grummer, R.R.; Rastani, R.R. Why Reevaluate Dry Period Length? J. Dairy Sci. 2004, 87, E77–E85. [Google Scholar] [CrossRef]
- Verweij, J.J.; Koets, A.P.; Eisenberg, S.W.F. Effect of continuous milking on immunoglobulin concentrations in bovine colostrum. Vet. Immunol. Immunopathol. 2014, 160, 225–229. [Google Scholar] [CrossRef]
- Sun, H.-Z.; Wang, D.-M.; Wang, B.; Wang, J.-K.; Liu, H.-Y.; Guan, L.L.; Liu, J.-X. Metabolomics of Four Biofluids from Dairy Cows: Potential Biomarkers for Milk Production and Quality. J. Proteome Res. 2015, 14, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Veshkini, A.; Hammon, H.M.; Sauerwein, H.; Tröscher, A.; Viala, D.; Delosière, M.; Ceciliani, F.; Déjean, S.; Bonnet, M. Longitudinal liver proteome profiling in dairy cows during the transition from gestation to lactation: Investigating metabolic adaptations and their interactions with fatty acids supplementation via repeated measurements ANOVA-simultaneous component analy. J. Proteom. 2022, 252, 104435. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Chang, G.T. A Literature Review on the Delactation Effect of Malt. Adv. Integr. Med. 2019, 6, S110–S111. [Google Scholar] [CrossRef]
- Lobos, N.E.; Wattiaux, M.A.; Broderick, G.A. Effect of rumen-protected lysine supplementation of diets based on corn protein fed to lactating dairy cows. J. Dairy Sci. 2021, 104, 6620–6632. [Google Scholar] [CrossRef] [PubMed]
- Mavrommatis, A.; Mitsiopoulou, C.; Christodoulou, C.; Kariampa, P.; Simoni, M.; Righi, F.; Tsiplakou, E. Effects of Supplementing Rumen-Protected Methionine and Lysine on Milk Performance and Oxidative Status of Dairy Ewes. Antioxidants 2021, 10, 654. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.Q.; Jiang, B.B.; Wang, H.R. Growth performance, meat quality and lipid metabolism in finishing lambs fed diets containing rumen-unprotected and rumen-protected betaine. Ital. J. Anim. Sci. 2021, 20, 2041–2050. [Google Scholar] [CrossRef]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2019, 10, 115–132. [Google Scholar] [CrossRef]
- Ogunade, I.; Schweickart, H.; McCoun, M.; Cannon, K.; McManus, C. Integrating 16S rRNA Sequencing and LC-MS-Based Metabolomics to Evaluate the Effects of Live Yeast on Rumen Function in Beef Cattle. Animals 2019, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Mamuad, L.L.; Kim, S.H.; Biswas, A.A.; Yu, Z.; Cho, K.-K.; Kim, S.-B.; Lee, K.; Lee, S.S. Rumen fermentation and microbial community composition influenced by live Enterococcus faecium supplementation. AMB Express 2019, 9, 123. [Google Scholar] [CrossRef]
- Choudhury, P.K.; Sirohi, S.K.; Puniya, A.K.; Chaudhary, P.P. Harnessing the Diversity of Rumen Microbes using Molecular Approaches. In Livestock Green House Gases: Emission and Options for Mitigation; Satish Serial Publishing House: Delhi, India, 2012. [Google Scholar]
- Xu, Q.B.; Qiao, Q.Q.; Gao, Y.; Hou, J.X.; Hu, M.Y.; Du, Y.F.; Zhao, K.; Li, X. Gut Microbiota and Their Role in Health and Metabolic Disease of Dairy Cow. Front. Nutr. 2021, 8, 701511. [Google Scholar] [CrossRef]
- Krause, K.M.; Oetzel, G.R. Understanding and preventing subacute ruminal acidosis in dairy herds: A review. Anim. Feed. Sci. Technol. 2006, 126, 215–236. [Google Scholar] [CrossRef]
- Oetzel, G.R. Understanding the Impact of Subclinical Ketosis. In Proceedings of the 2012 Cornell Nutrition Conference for Feed Manufacturers; University of Wisconsin: Madison, WI, USA, 2012; pp. 15–26. [Google Scholar]
- Luo, Z.Z.; Shen, L.H.; Jiang, J.; Huang, Y.X.; Bai, L.P.; Yu, S.M.; Yao, X.P.; Ren, Z.H.; Yang, Y.X.; Cao, S.Z. Plasma metabolite changes in dairy cows during parturition identified using untargeted metabolomics. J. Dairy Sci. 2019, 102, 4639–4650. [Google Scholar] [CrossRef]
- Piazzolla, V.A.; Mangia, A. Noninvasive Diagnosis of NAFLD and NASH. Cells 2020, 9, 1005. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Zhu, Y.; Xiao, J.; Qian, B.; You, L.; Zhang, Y.; Yu, S.; Zong, X.; Cao, S. Serum adipokines play different roles in type I and II ketosis. Asian-Australas. J. Anim. Sci. 2019, 33, 1930. [Google Scholar] [CrossRef] [PubMed]
- Heringstad, B.; Chang, Y.M.; Gianola, D.; Klemetsdal, G. Genetic analysis of clinical mastitis, milk fever, ketosis, and retained placenta in three lactations of Norwegian Red cows. J. Dairy Sci. 2005, 88, 3273–3281. [Google Scholar] [CrossRef]
- Tetens, J.; Heuer, C.; Heyer, I.; Klein, M.S.; Gronwald, W.; Junge, W.; Oefner, P.J.; Thaller, G.; Krattenmacher, N. Polymorphisms within the APOBR gene are highly associated with milk levels of prognostic ketosis biomarkers in dairy cows. Physiol. Genom. 2015, 47, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Loor, J.J.; Everts, R.E.; Bionaz, M.; Dann, H.M.; Morin, D.E.; Oliveira, R.; Rodriguez-Zas, S.L.; Drackley, J.K.; Lewin, H.A. Nutrition-induced ketosis alters metabolic and signaling gene networks in liver of periparturient dairy cows. Physiol. Genom. 2007, 32, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, K. Systems Physiology and Nutrition in Dairy Cattle: Applications of Omics and Bioinformatics to Better Understand the Hepatic Metabolomics and Transcriptomics Adaptations in Transition Dairy Cows; University of Illinois at Urbana-Champaign: Champaign, IL, USA, 2017. [Google Scholar]
- Akbar, H.; Batistel, F.; Drackley, J.K.; Loor, J.J. Alterations in Hepatic FGF21, Co-Regulated Genes, and Upstream Metabolic Genes in Response to Nutrition, Ketosis and Inflammation in Peripartal Holstein Cows. PLoS ONE 2015, 10, e0139963. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Sun, L.-W.; Xia, C.; Zhang, H.-Y.; Zheng, J.-S.; Wang, J.-S. 1H-Nuclear Magnetic Resonance-Based Plasma Metabolic Profiling of Dairy Cows with Fatty Liver. Asian-Australas. J. Anim. Sci. 2015, 29, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Soares, R.A.N.; Vargas, G.; Muniz, M.M.M.; Soares, M.A.M.; Cánovas, A.; Schenkel, F.; Squires, E.J. Differential gene expression in dairy cows under negative energy balance and ketosis: A systematic review and meta-analysis. J. Dairy Sci. 2021, 104, 602–615. [Google Scholar] [CrossRef]
- Chella Krishnan, K.; Kurt, Z.; Barrere-Cain, R.; Sabir, S.; Das, A.; Floyd, R.; Vergnes, L.; Zhao, Y.; Che, N.; Charugundla, S.; et al. Integration of Multi-omics Data from Mouse Diversity Panel Highlights Mitochondrial Dysfunction in Non-alcoholic Fatty Liver Disease. Cell Syst. 2018, 6, 103–115.e107. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhu, K.-L.; Chen, Y.-Y.; Yang, W.; Xia, C.; Zhang, H.-Y.; Wu, L.; Shen, T.-Y.; Yu, H.; Xu, Q.-S.; et al. Isolation Identification and Bioinformatics of Differences Protein in Plasma of Cows Suffer from Fatty Liver with SELDI-TOF-MS Techniques. Sci. Agric. Sin. 2016, 49, 1585–1598. [Google Scholar]
- Wu, Z.L.; Chen, S.Y.; Hu, S.Q.; Jia, X.B.; Wang, J.; Lai, S.J. Metabolomic and Proteomic Profiles Associated With Ketosis in Dairy Cows. Front. Genet. 2020, 11, 551587. [Google Scholar] [CrossRef]
- Seegers, H.; Fourichon, C.; Beaudeau, F. Production effects related to mastitis and mastitis economics in dairy cattle herds. Vet. Res. 2003, 34, 475–491. [Google Scholar] [CrossRef] [PubMed]
- Jain, N.C. Common Mammary Pathogens and Factors in Infection and Mastitis. J. Dairy Sci. 1979, 62, 128–134. [Google Scholar] [CrossRef]
- Wellnitz, O.; Bruckmaier, R.M. The innate immune response of the bovine mammary gland to bacterial infection. Vet. J. 2012, 192, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Ruegg, P.L. A 100-Year Review: Mastitis detection, management, and prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [PubMed]
- Giger-Reverdin, S. Recent advances in the understanding of subacute ruminal acidosis (SARA) in goats, with focus on the link to feeding behaviour. Small Rumin. Res. 2018, 163, 24–28. [Google Scholar] [CrossRef]
- Dill-McFarland, K.A.; Breaker, J.D.; Suen, G. Microbial succession in the gastrointestinal tract of dairy cows from 2 weeks to first lactation. Sci. Rep. 2017, 7, 40864. [Google Scholar] [CrossRef]
- Qi, W.P.; Mu, Y.Y.; Zhang, T.; Zhang, J.Y.; Mao, S.Y. Plasma biochemical indexes and metabolomics profile changes of dairy cows with subacute ruminal acidosis. Acta Pratacult. Sin. 2021, 30, 141–150. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| First Author | Omics Applied | Year | Techniques | Targeted/Non-Targeted | Outcome | Reference |
|---|---|---|---|---|---|---|
| Pryce | Genomics | 2014 | - | - | Residual feed intake could be used as a breeding trait | [14] |
| Sigdel | Whole Genomic Mapping | 2019 | At least three different genomic regions on BTA5, BTA14, and BTA15 are strongly associated with milk production under heat stress | [15] | ||
| Tarekegn | Genomics | 2021 | - | - | Fertility-involved SNPs are different in Swedish red and Holstein cows | [16] |
| Feugang | Spermatozoa transcriptomics | 2010 | - | - | CD36 molecule decreased in low fertility bulls | [17] |
| Canovas | Milk transcriptomic | 2010 | - | - | Over 33,000 SNPs involved in lactation process | [18] |
| Akbar | Liver transcriptomic | 2013 | - | - | Feed restriction but not L-carnitine increased expression of GPX3,PC, PDK4, SAA3, and ADIPOR2 | [19] |
| Fagerlind | Spermatozoa transcriptomics | 2015 | - | - | Mir-502-5p, mir-1249, mir-320a, mir-34c-3p, mir-19b-3p, mir-27a-5p and mir-148b-3p expressed differently with fertility | [20] |
| Wang | miRNA Transcriptomic | 2016 | - | - | MiRNAs expressed in these five tissues play roles in regulating the transportation of AA for downstream milk production | [21] |
| Li | Rumen Meta transcriptomics | 2017 | - | - | Carbohydrate active enzymes are related to feed efficiency | [22] |
| Comtet-Marre | Rumen Meta transcriptomics | 2017 | - | - | Cellobiose-phosphorylase, amylase, hemicellulases, cellulases, pectinase, and oligosaccharidases are main carbohydrate active enzymes | [23] |
| Song | Hind gut microbiome and meta-transcriptomic | 2017 | 454 sequencing | - | SCFA alters the hindgut microbiome and their transcripts | [24] |
| Wang | Rumen Transcriptomic | 2017 | - | - | Expression of proliferation and apoptotic processes (BAG3, HLA-DQA1, and UGT2B17) related protein changes with different forages | [25] |
| Sollinger | Rumen meta transcriptomic | 2018 | - | - | Methyl-reducing but not CO2-reducing methanogens were positively correlated with methane emissions. Methanosphaera is the dominating methanol-reducing methanogen. | [26] |
| Putz | miRNA Transcriptomic | 2019 | - | - | 46 miRNAs changes in the transition period | [27] |
| Li | Rumen meta-transcriptomic, liver transcriptomic | 2019 | - | - | 627 gene involved in cell signaling and morphogenesis expressed differentially during acidosis | [28] |
| Li | Rumen meta-transcriptomic. Rumen epithelial transcriptomic | 2019 | - | - | Acidosis affected the expression of lipid metabolism involved genes | [29] |
| Ogunade | Metatranscriptomic | 2019 | - | - | Carbohydrate, amino acid, energy, vitamin and co-factor metabolism pathways, and bacterial biofilm formation pathways changes in the ruminal acidosis | [30] |
| Ametaj | Rumen metabolomic | 2010 | 1H-NMR, GC-MS | Non-targeted | Over 30% proportion of barley grain diet increased potentially toxic metabolites | [31] |
| Zhang | Metabolomic, transcriptomic | 2015 | GC-MS | Non-targeted | Ruminal xanthine, hypoxanthine and uracil, biogenic amines, ethanolamine, glutaric acid, and amino acids concentrations elevated in the acidosis | [32] |
| Forde | Follicular-fluid metabolomic | 2016 | GC-MS | Non-targeted | Follicular-fluid of dry cows have higher tyrosine, phenylalanine and valine and fatty acids heneicosanoic acid and docosahexaenoic acid concentrations | [33] |
| Thomas | Milk metabolomics | 2016 | LC-MS | Non-targeted | Metabolites relevant to carbohydrate and nucleotide decrease after infection | [34] |
| Alejandro | Ruminal microbiome & metabolomic | 2016 | LC-MS | Non-targeted | Vitamin E changes rumen microbiome and enhances dry matter degradation | [12,35] |
| Humer | Serum metabolomic | 2016 | LC-ESI | Non-targeted | Excessive sphingolipids and phospholipids degradation is related to decreased insulin sensitivity in transition cows | [36] |
| Sun | Urine metabolomic | 2016 | GC-TOF/MS | Non-targeted | Hippuric acid and N-methyl-glutamic concentrations are significantly different between alfalfa hay fed and corn stover fed cows | [37] |
| Dai | Milk transcriptomic and proteomic | 2017 | LC-MS | iTRAQ labelling | Rice stover inhibits protein synthesis of dairy cows | [38] |
| Artegoitia | Rumen Fluid Metabolomic | 2017 | LC-MS | Non-targeted | linoleic and alpha-linolenic metabolism are correlated to daily growth | [39] |
| Sun | Umbilical blood metabolomic | 2017 | 1H-NMR | Non-targeted | Rumen-protected arginine supplementation altered metabolic pathways of amino acid, carbohydrate and energy, lipids and oxidative stress metabolism of pregnancy cows | [40] |
| Murovec | Metabolomics | 2018 | 1H-NMR | Non-targeted | Simulated an in vitro acidosis rumen model | [41] |
| Elolimy | Fecal metabolomic | 2019 | LC-MS | Non-targeted | Rumen-protected methionine supplementation onlate-pregnancy cows enhanced endogenous antibiotics synthesis, also hindgut functionality and health of their calves | [42] |
| Ogunade | ruminal fluid Metabolomics | 2019 | LC-MS | Non-targeted | Live yeast supplementation increased the concentrations of 4-cyclohexanedione and glucopyranoside and decreased the concentrations of threonic acid, xanthosine, deoxycholic acid, lauroyl carnitine, methoxybenzoic acid, and pentadecylbenzoic acid | [26] |
| Sun | Metabolomic, transcriptomic, | 2020 | GC | Non-targeted | Propionate, glucose, and amino acid concentration decreased in feeding with low-quality corp. Hippuric acid is the biomarker of corn stover fed cow | [43] |
| Zhang | rumen fluid metabolomic | 2020 | LC-MS | Non-targeted | Metabolites involved in protein digestion and absorption, ABC transporters, and unsaturated fatty acid biosynthesis pathways are correlated with milk yield | [44] |
| Clemmons | Rumen Fluid Metabolomic | 2020 | LC-MS | Non-targeted | Metabolites involved in amino acid and lipid metabolism are related to feeding efficiency | [45] |
| Xue | Rumen Metagenomics and meta-metabolomics | 2020 | GC-MS | Non-targeted | Rumen microbial composition, functions, and metabolites, and the serum metabolites are contributed to milk protein yield | [9] |
| Ogunade | Ruminal microbiome &metabolomic | 2020 | LC-MS | Non-targeted | DFMs alter rumen metabolites pattern and microbiome | [46] |
| Wang | Serum metabolomic | 2020 | GC−TOF/MS | Non-targeted | Rumen-Protected Betaine alters arginine synthesis and proline degradation and cyanoamino acid synthesis, promotes milk production | [47] |
| Luke | Serum metabolomic | 2020 | 1H-NMR | Non-targeted | Quantified the relationship between NMR spectra and concentrations of the current gold standard serum biomarker of energy balance, beta-hydroxybutyrate | [48] |
| Lisuzzo | Serum metabolomic | 2022 | 1H-NMR | Non-targeted | Correlations between serum ketone levels and milk lipid components in cows | [49] |
| Wang | Milk and rumen metabolomic | 2021 | UPLC-qTOF-MS | Non-targeted | Supplementation of perilla frutescens leaf could alter the ruminal metabolic profiles and milk synthesis through regulation of the pathways of pyrimidine metabolism and biosynthesis of unsaturated fatty acids | [50] |
| Gu | Milk Transcriptomic Metabolomic | 2021 | LC-MS/MS | Non-targeted | Rumen-protected methionine supplement increased α-ketoglutaric acid concentration, and related to rumen Thermoactinomyces, Asteroleplasma and Saccharofermentan abundance | [8] |
| Stergiadis | Rumen lipidomic, metabolomics, and microbiome | 2021 | GC (lipidomic), NMR (metabolomic) | Non-targeted | Cows with high milk fatty acid have higher butyrate, propionate and tyrosine and lower concentrations of xanthine and hypoxanthine concentrations | [51] |
| Wang | Rumen microbiome & metabolomic | 2021 | UPLC-QTOF/MS | Non-targeted | Rumen-protected glucose increased bacterial richness and diversity, also acetate, propionate, butyrate, and total volatile fatty acid in the rumen | [52] |
| Peddinti | Spermatozoa Proteomics | 2008 | DDF-2-LC-MS | Non-targeted | High-fertility bull and higher protein expression in energy metabolism, cell communication, spermatogenesis, and cell motility | [53] |
| Ledgard | Uterine luminal proteomics | 2012 | 2-DE-MS | Non-targeted | Phosphoserine aminotransferase 1, purine nucleoside phosphorylase, and aldose reductase expression are related to the embryo growth environment | [54] |
| Saadi | Sperm proteomics | 2013 | LC-MS/MS | Non-targeted | Proteins involved in sperm capacitation, sperm–egg interaction, and sperm cytoskeletal structure were decreased in pyriform sperm, whereas proteins regulating antioxidant activity, apoptosis, and metabolic activity increased | [55] |
| Li | Milk proteomic | 2015 | 2-DE- MALDI-TOF/TOF-MS | Process method of corn influences milk proteome pattern | [56] | |
| Thomas | Milk peptidomics | 2016 | LC-MS/MS | Non--targeted | The abundance of caseins, beta-lactoglobulin, and alpha-lactalbumin to albumin, lactoferrin, and IgG shifted during the infection | [57] |
| Zachut | follicular fluids proteomics | 2016 | LC-MS | Non-targeted | Protein relevant to follicular function expressed differently in less fertility cows | [58] |
| Mudaliar | Milk proteomics | 2016 | LC-MS | Non-targeted | Antimicrobial peptides concentration elevates in the acute phase of mastitis | [59] |
| Snelling | Rumen metaproteomic | 2017 | 2-DE-LC-MS | Non-targeted | 2D-PAGE reveals key structural proteins and enzymes in the rumen microbial community | [60] |
| Skibiel | Liver proteomics | 2018 | nano-UPLC | Non-targeted | Oxidative phosphorylation, mitochondrial dysfunction, farnesoid X receptor/retinoid X receptor (FXR/RXR) activation, and the methylmalonyl pathway changes in the heat stress | [61] |
| Veshkini | Liver proteomic | 2020 | LC-MS/MS | Non-targeted | EFA and CLA status in transition cows had an impact on energy, lipid and vitamin metabolisms, and oxidative stress balance | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Bu, D.; Ma, L. Integration of Multiplied Omics, a Step Forward in Systematic Dairy Research. Metabolites 2022, 12, 225. https://doi.org/10.3390/metabo12030225
Zhu Y, Bu D, Ma L. Integration of Multiplied Omics, a Step Forward in Systematic Dairy Research. Metabolites. 2022; 12(3):225. https://doi.org/10.3390/metabo12030225
Chicago/Turabian StyleZhu, Yingkun, Dengpan Bu, and Lu Ma. 2022. "Integration of Multiplied Omics, a Step Forward in Systematic Dairy Research" Metabolites 12, no. 3: 225. https://doi.org/10.3390/metabo12030225
APA StyleZhu, Y., Bu, D., & Ma, L. (2022). Integration of Multiplied Omics, a Step Forward in Systematic Dairy Research. Metabolites, 12(3), 225. https://doi.org/10.3390/metabo12030225