
Ramipril Reduces Acylcarnitines and Distinctly Increases Angiotensin-Converting Enzyme 2 Expression in Lungs of Rats

 , ,
, ,  and
and
Abstract
:
1. Introduction
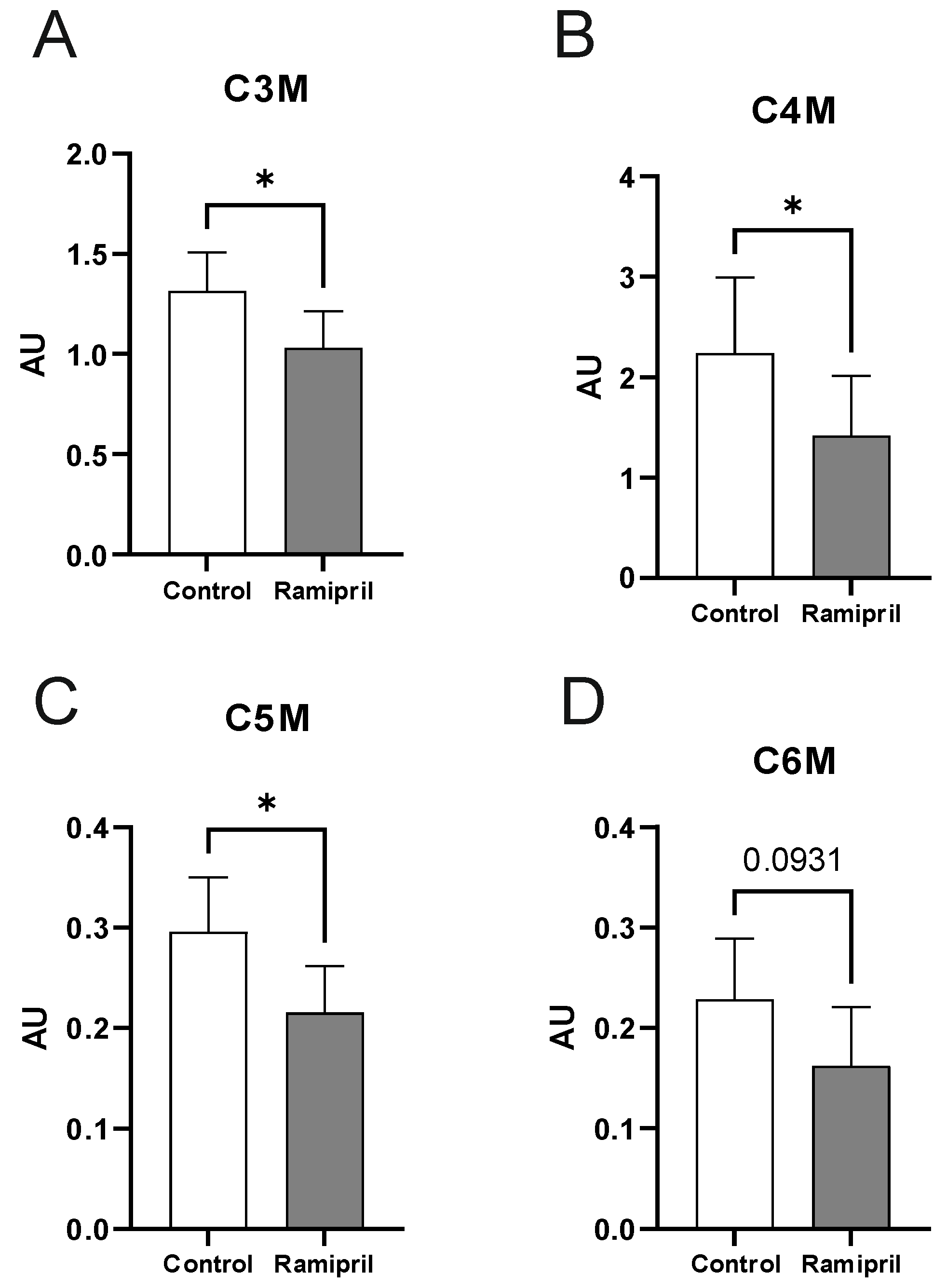
2. Results
3. Discussion
4. Material and Methods
4.1. Animals and Experimental Design
4.2. Molecular and Protein Analysis
4.3. Metabolome Analysis
4.4. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chauhan, M.; Patel, J.; Ahmad, F. StatPearls: Ramipril; StatPearls: Treasure Island, FL, USA, 2022.
- Meng, J.; Xiao, G.; Zhang, J.; He, X.; Ou, M.; Bi, J.; Yang, R.; Di, W.; Wang, Z.; Li, Z.; et al. Renin-angiotensin system inhibitors improve the clinical outcomes of COVID-19 patients with hypertension. Emerg. Microbes Infect. 2020, 9, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Gheblawi, M.; Oudit, G.Y. Angiotensin Converting Enzyme 2: A Double-Edged Sword. Circulation 2020, 142, 426–428. [Google Scholar] [CrossRef] [PubMed]
- Danser, A.H.J.; Epstein, M.; Batlle, D. Renin-Angiotensin System Blockers and the COVID-19 Pandemic: At Present There Is No Evidence to Abandon Renin-Angiotensin System Blockers. Hypertension 2020, 75, 1382–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrario, C.M. Cardiac remodelling and RAS inhibition. Ther. Adv. Cardiovasc. Dis. 2016, 10, 162–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrario, C.M.; Jessup, J.; Chappell, M.C.; Averill, D.B.; Brosnihan, K.B.; Tallant, E.A.; Diz, D.I.; Gallagher, P.E. Effect of angiotensin-converting enzyme inhibition and angiotensin II receptor blockers on cardiac angiotensin-converting enzyme 2. Circulation 2005, 111, 2605–2610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertanto, D.M.; Wiratama, B.S.; Sutanto, H.; Wungu, C.D.K. Immunomodulation as a Potent COVID-19 Pharmacotherapy: Past, Present and Future. J. Inflamm. Res. 2021, 14, 3419–3428. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.L.C.; Lely, A.T.; Navis, G.J.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Batlle, D.; Jose Soler, M.; Ye, M. ACE2 and diabetes: ACE of ACEs? Diabetes 2010, 59, 2994–2996. [Google Scholar] [CrossRef] [Green Version]
- Tikellis, C.; Bialkowski, K.; Pete, J.; Sheehy, K.; Su, Q.; Johnston, C.; Cooper, M.E.; Thomas, M.C. ACE2 deficiency modifies renoprotection afforded by ACE inhibition in experimental diabetes. Diabetes 2008, 57, 1018–1025. [Google Scholar] [CrossRef] [Green Version]
- Koshy, A.N.; Murphy, A.C.; Farouque, O.; Ramchand, J.; Burrell, L.M.; Yudi, M.B. Renin-angiotensin system inhibition and risk of infection and mortality in COVID-19: A systematic review and meta-analysis. Intern. Med. J. 2020, 50, 1468–1474. [Google Scholar] [CrossRef] [PubMed]
- Vaduganathan, M.; Vardeny, O.; Michel, T.; McMurray, J.J.V.; Pfeffer, M.A.; Solomon, S.D. Renin-Angiotensin-Aldosterone System Inhibitors in Patients with COVID-19. N. Engl. J. Med. 2020, 382, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- Bindom, S.M.; Hans, C.P.; Xia, H.; Boulares, A.H.; Lazartigues, E. Angiotensin I-converting enzyme type 2 (ACE2) gene therapy improves glycemic control in diabetic mice. Diabetes 2010, 59, 2540–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tocci, G.; Paneni, F.; Palano, F.; Sciarretta, S.; Ferrucci, A.; Kurtz, T.; Mancia, G.; Volpe, M. Angiotensin-converting enzyme inhibitors, angiotensin II receptor blockers and diabetes: A meta-analysis of placebo-controlled clinical trials. Am. J. Hypertens. 2011, 24, 582–590. [Google Scholar] [CrossRef] [Green Version]
- Abuissa, H.; Jones, P.G.; Marso, S.P.; O’Keefe, J.H. Angiotensin-converting enzyme inhibitors or angiotensin receptor blockers for prevention of type 2 diabetes: A meta-analysis of randomized clinical trials. J. Am. Coll. Cardiol. 2005, 46, 821–826. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Karakiulakis, G.; Roth, M. Are patients with hypertension and diabetes mellitus at increased risk for COVID-19 infection? Lancet Respir. Med. 2020, 8, e21. [Google Scholar] [CrossRef]
- Garg, S.; Kim, L.; Whitaker, M.; O’Halloran, A.; Cummings, C.; Holstein, R.; Prill, M.; Chai, S.J.; Kirley, P.D.; Alden, N.B.; et al. Hospitalization Rates and Characteristics of Patients Hospitalized with Laboratory-Confirmed Coronavirus Disease 2019—COVID-NET, 14 States, March 1-30, 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 458–464. [Google Scholar] [CrossRef]
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Vogt, L.; Klöting, I. Model-based prediction of diabetes incidence in BB/OK rats. Diabete Metab. 1993, 19, 183–187. [Google Scholar]
- Klöting, N.; Wilke, B.; Klöting, I. Triplet repeat in the Repin1 3′-untranslated region on rat chromosome 4 correlates with facets of the metabolic syndrome. Diabetes. Metab. Res. Rev. 2007, 23, 406–410. [Google Scholar] [CrossRef]
- Sarganas, G.; Knopf, H.; Grams, D.; Neuhauser, H.K. Trends in Antihypertensive Medication Use and Blood Pressure Control Among Adults with Hypertension in Germany. Am. J. Hypertens. 2016, 29, 104–113. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.R.; La George De Rosa, M.V.; Rosania, G.R.; Stringer, K.A. L-Carnitine and Acylcarnitines: Mitochondrial Biomarkers for Precision Medicine. Metabolites 2021, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Schooneman, M.G.; Vaz, F.M.; Houten, S.M.; Soeters, M.R. Acylcarnitines: Reflecting or inflicting insulin resistance? Diabetes 2013, 62, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zeng, X.; Ren, M.; Mao, X.; Qiao, S. Novel metabolic and physiological functions of branched chain amino acids: A review. J. Anim. Sci. Biotechnol. 2017, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Soeters, M.R.; Sauerwein, H.P.; Duran, M.; Wanders, R.J.; Ackermans, M.T.; Fliers, E.; Houten, S.M.; Serlie, M.J. Muscle acylcarnitines during short-term fasting in lean healthy men. Clin. Sci. 2009, 116, 585–592. [Google Scholar] [CrossRef] [Green Version]
- Soeters, M.R.; Serlie, M.J.; Sauerwein, H.P.; Duran, M.; Ruiter, J.P.; Kulik, W.; Ackermans, M.T.; Minkler, P.E.; Hoppel, C.L.; Wanders, R.J.A.; et al. Characterization of D-3-hydroxybutyrylcarnitine (ketocarnitine): An identified ketosis-induced metabolite. Metabolism 2012, 61, 966–973. [Google Scholar] [CrossRef]
- Kien, C.L.; Everingham, K.I.; Stevens, D.R.; Fukagawa, N.K.; Muoio, D.M. Short-term effects of dietary fatty acids on muscle lipid composition and serum acylcarnitine profile in human subjects. Obesity 2011, 19, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Mihalik, S.J.; Goodpaster, B.H.; Kelley, D.E.; Chace, D.H.; Vockley, J.; Toledo, F.G.; DeLany, J.P. Increased Levels of Plasma Acylcarnitines in Obesity and Type 2 Diabetes and Identification of a Marker of Glucolipotoxicity. Obesity 2010, 18, 1695–1700. [Google Scholar] [CrossRef] [Green Version]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [Green Version]
- Ayon, N.J.; Sharma, A.D.; Gutheil, W.G. LC-MS/MS-Based Separation and Quantification of Marfey’s Reagent Derivatized Proteinogenic Amino Acid DL-Stereoisomers. J. Am. Soc. Mass Spectrom. 2019, 30, 448–458. [Google Scholar] [CrossRef]
- Gray, N.; Zia, R.; King, A.; Patel, V.C.; Wendon, J.; McPhail, M.J.W.; Coen, M.; Plumb, R.S.; Wilson, I.D.; Nicholson, J.K. High-Speed Quantitative UPLC-MS Analysis of Multiple Amines in Human Plasma and Serum via Precolumn Derivatization with 6-Aminoquinolyl-N-hydroxysuccinimidyl Carbamate: Application to Acetaminophen-Induced Liver Failure. Anal. Chem. 2017, 89, 2478–2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesselbarth, N.; Kunath, A.; Kern, M.; Gericke, M.; Mejhert, N.; Rydén, M.; Stumvoll, M.; Blüher, M.; Klöting, N. Repin1 deficiency in adipose tissue improves whole-body insulin sensitivity, and lipid metabolism. Int. J. Obes. 2017, 41, 1815–1823. [Google Scholar] [CrossRef] [PubMed]
- Klöting, N.; Koch, L.; Wunderlich, T.; Kern, M.; Ruschke, K.; Krone, W.; Brüning, J.C.; Blüher, M. Autocrine IGF-1 action in adipocytes controls systemic IGF-1 concentrations and growth. Diabetes 2008, 57, 2074–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brauer, R.; Leichtle, A.B.; Fiedler, G.M.; Thiery, J.; Ceglarek, U. Preanalytical standardization of amino acid and acylcarnitine metabolite profiling in human blood using tandem mass spectrometry. Metabolomics 2011, 7, 344–352. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Controls (n = 6) | Ramipril-Treated (n = 6) | ||||
|---|---|---|---|---|---|
| Pre | Post | Pre | Post | ||
| Body weight (g) | 360 ± 18 | 368 ± 17 | 338 ± 18 | 341 ± 18 | n.s. |
| HbA1c (%) | 4.2 ± 0.1 | 4.2 ± 0.1 | 4.2 ± 0.2 | 4.4 ± 0.3 | n.s. |
| Adiposity Index (AI) | 1.7 ±0.4 | 2.0 ± 0.3 | n.s. | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosacka, J.; Berger, C.; Ceglarek, U.; Hoffmann, A.; Blüher, M.; Klöting, N. Ramipril Reduces Acylcarnitines and Distinctly Increases Angiotensin-Converting Enzyme 2 Expression in Lungs of Rats. Metabolites 2022, 12, 293. https://doi.org/10.3390/metabo12040293
Kosacka J, Berger C, Ceglarek U, Hoffmann A, Blüher M, Klöting N. Ramipril Reduces Acylcarnitines and Distinctly Increases Angiotensin-Converting Enzyme 2 Expression in Lungs of Rats. Metabolites. 2022; 12(4):293. https://doi.org/10.3390/metabo12040293
Chicago/Turabian StyleKosacka, Joanna, Claudia Berger, Uta Ceglarek, Anne Hoffmann, Matthias Blüher, and Nora Klöting. 2022. "Ramipril Reduces Acylcarnitines and Distinctly Increases Angiotensin-Converting Enzyme 2 Expression in Lungs of Rats" Metabolites 12, no. 4: 293. https://doi.org/10.3390/metabo12040293
APA StyleKosacka, J., Berger, C., Ceglarek, U., Hoffmann, A., Blüher, M., & Klöting, N. (2022). Ramipril Reduces Acylcarnitines and Distinctly Increases Angiotensin-Converting Enzyme 2 Expression in Lungs of Rats. Metabolites, 12(4), 293. https://doi.org/10.3390/metabo12040293