
Unusual Lipid Components of Legionella gormanii Membranes

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Composition of Fatty Acids of Individual Classes of Phospholipids
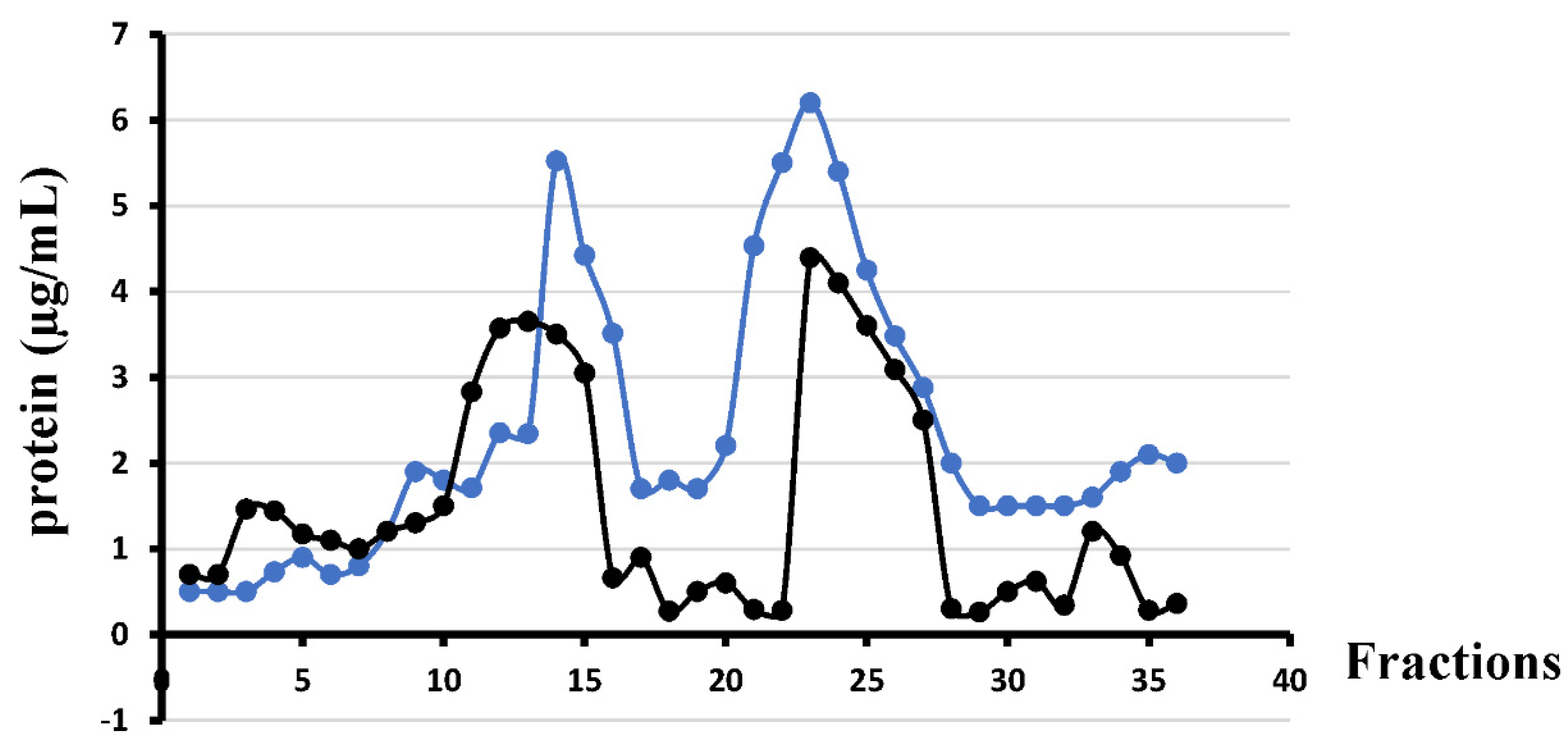
2.2. Separation of the Inner (IM) and Outer (OM) Membranes
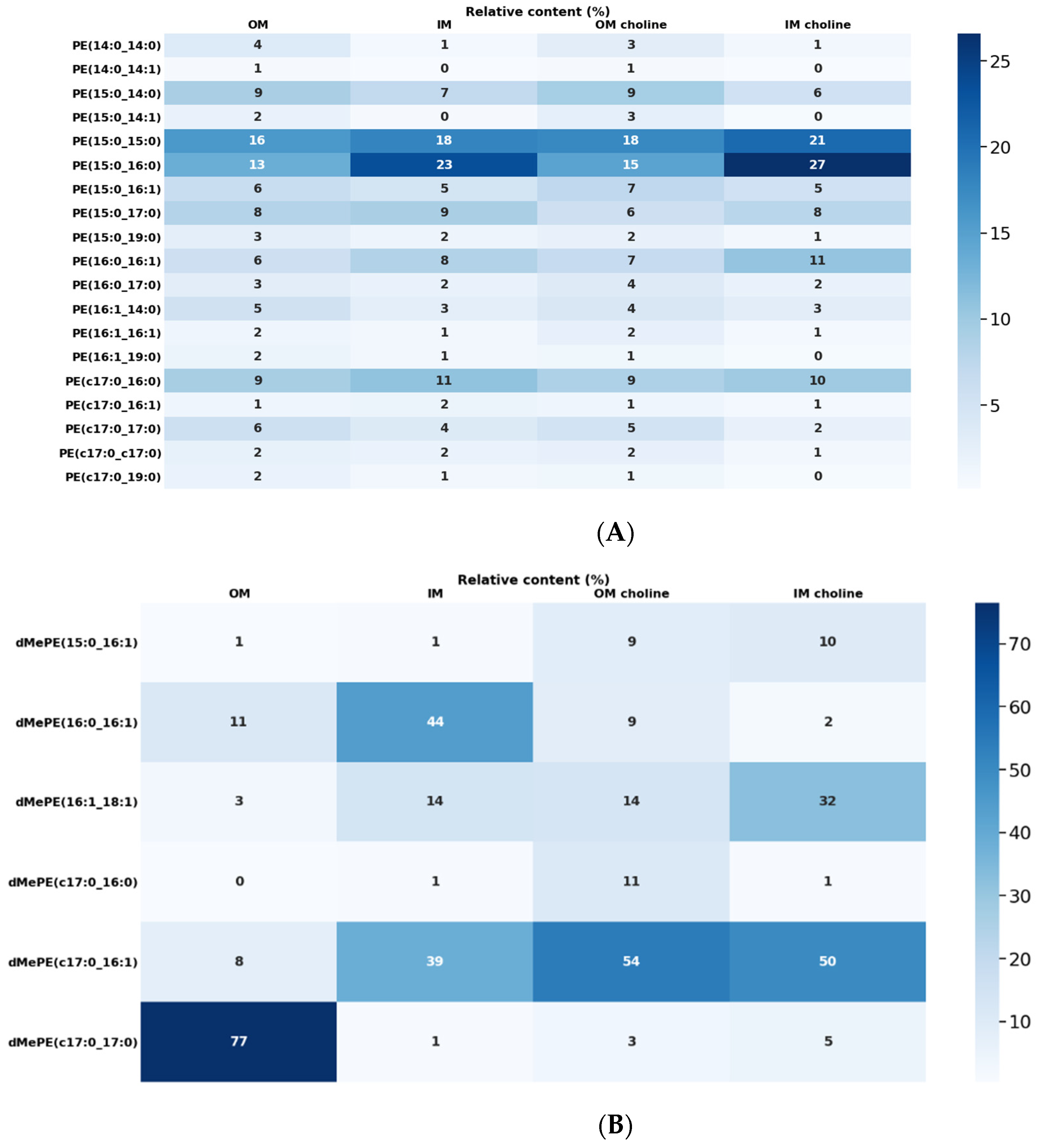
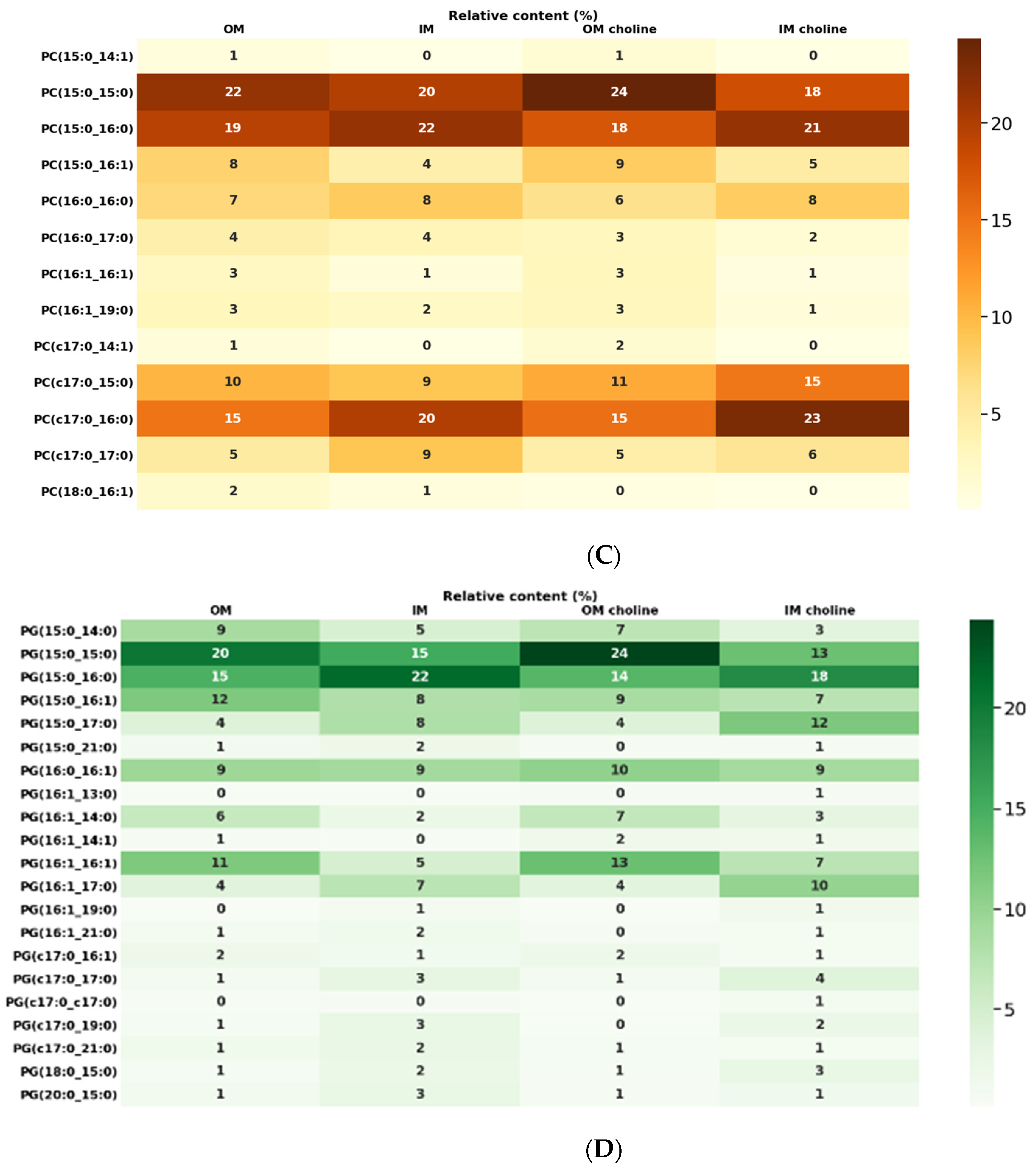
2.3. Lipidomic Analysis
2.4. Phospholipid Profile
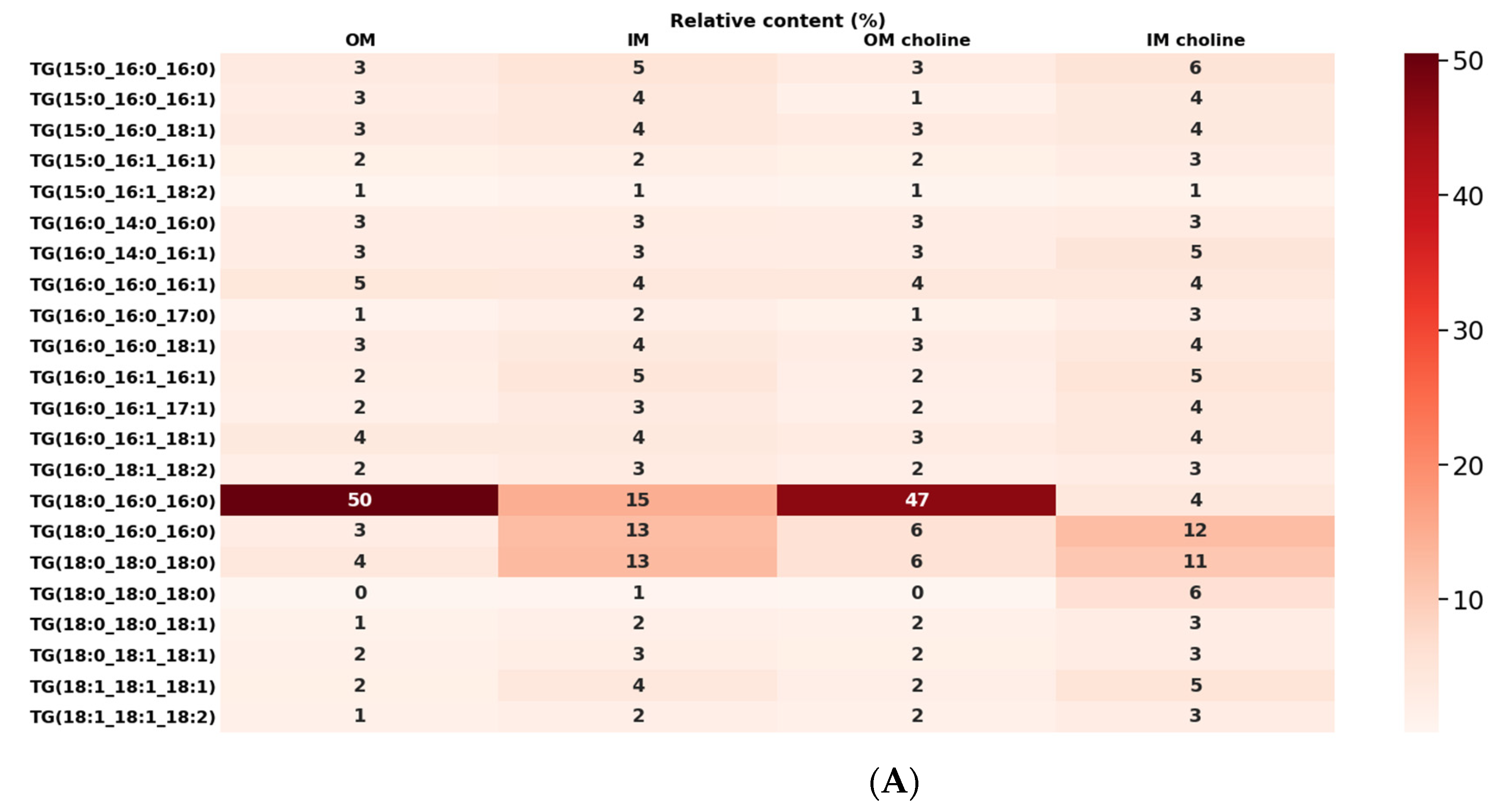
2.5. Analysis of Glycerolipids
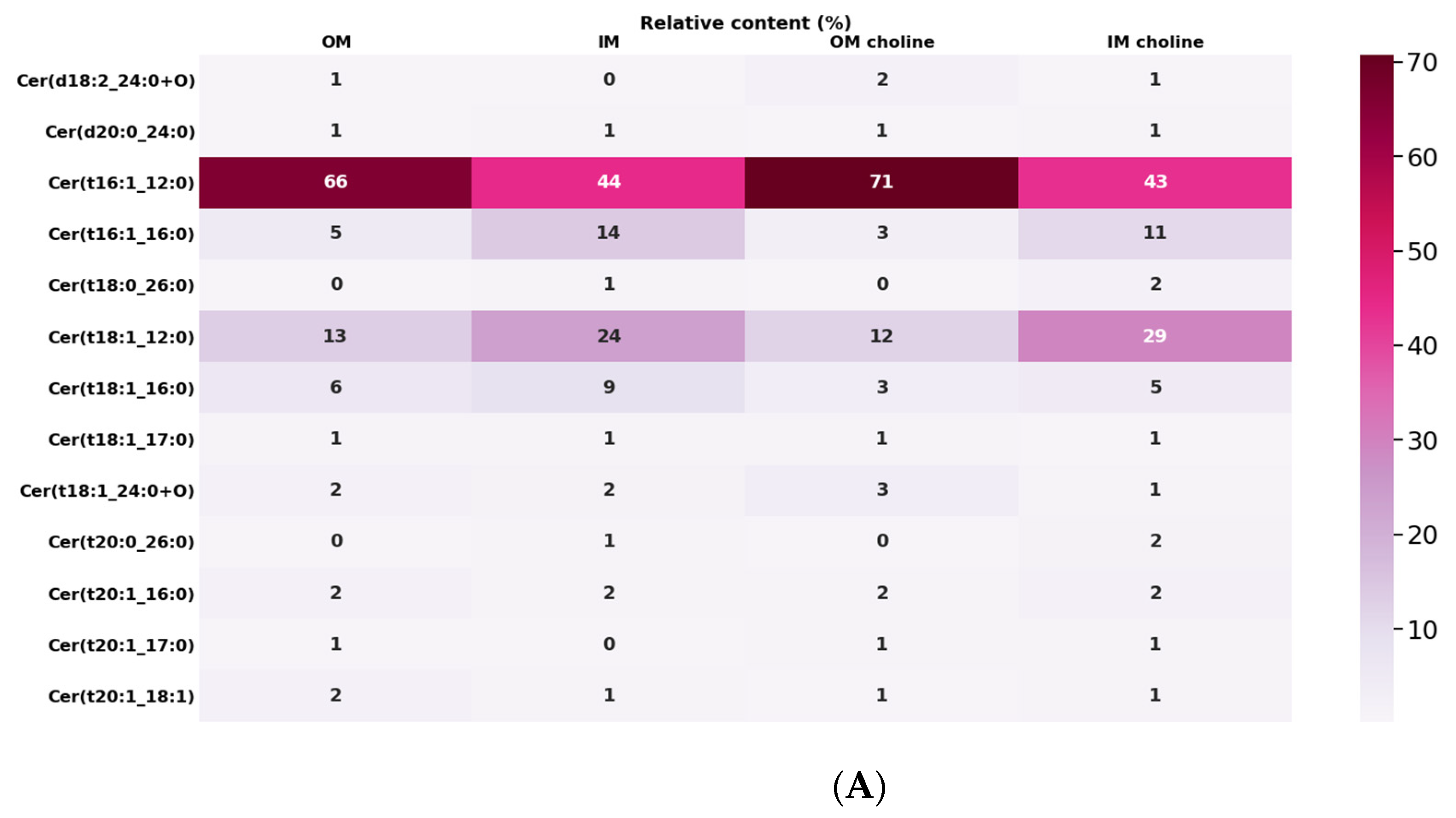
2.6. Characteristic of Ceramides
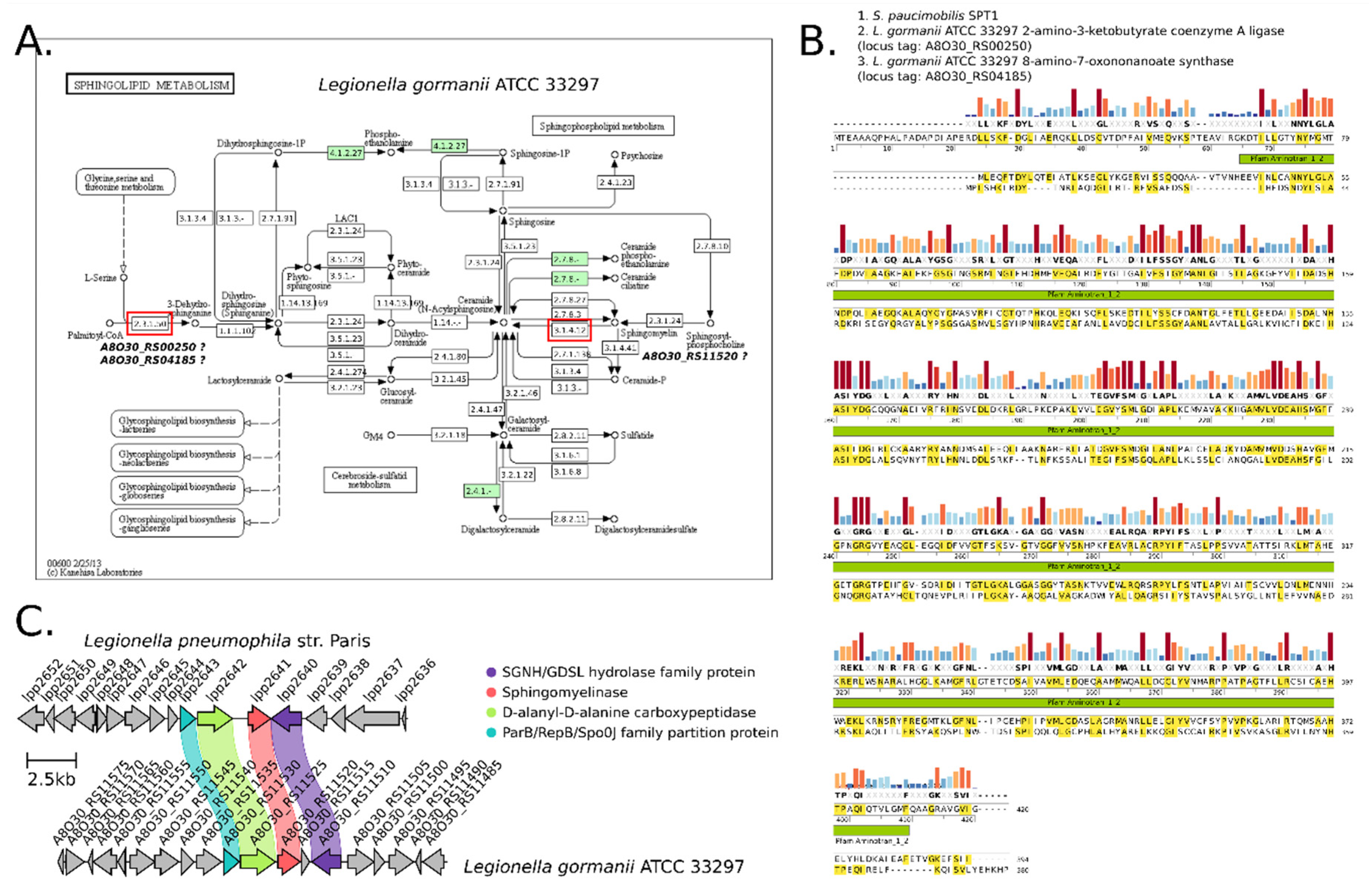
2.7. Bioinformatic Search for Ceramide Synthesis Pathways in the Genome of L. gormanii
3. Discussion
4. Materials and Methods
4.1. Strain and Growth Condition
4.2. Isolation of Phospholipids and Thin-Layer Chromatography
4.3. Preparation of Fatty Acid Methyl Esters
4.4. Gas-Liquid Chromatography and Mass Spectrometry
4.5. Separation of IM and OM in a Sucrose Density Gradient
4.6. Ultra-High Performance Liquid Chromatography/Mass Spectrometry (UHP LC-MS/MS)
4.7. Bioinformatic Analysis
4.7.1. Sequence Retrieval and Gene Annotation
4.7.2. BLASTp and PSI-BLAST Searches
4.7.3. K Number Assignment and KEGG Mapping
4.7.4. Gene Cluster Comparisons
4.7.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fields, B.S.; Benson, R.F.; Besser, R.E. Legionella and Legionnaires’ disease: 25 years of investigation. Clin. Microbiol. Rev. 2002, 15, 506–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauté, J.; Zucs, P.; de Jong, B. European Legionnaires’ Disease Surveillance Network. Legionnaires’ disease in Europe, 2009–2010. Eurosurveillance 2013, 18, 20417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, S.T.; Slow, S.; Scott-Thomas, A.; Murdoch, D.R. Legionellosis caused by non-Legionella pneumophila species, with a focus on Legionella longbeachae. Microorganisms 2021, 9, 291. [Google Scholar] [CrossRef] [PubMed]
- Griffith, M.E.; Lindquist, D.S.; Benson, R.F.; Thacker, W.L.; Brenner, D.J.; Wilkinson, H.W. First isolation of Legionella gormanii from human disease. J. Clin. Microbiol. 1988, 26, 380–381. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, D.; Chiou, C.C.; Famigilleti, R.; Lee, T.C.; Yu, V.L. Problem pathogens: Paediatric legionellosis—Implications for improved diagnosis. Lancet Infect. Dis. 2006, 6, 529–535. [Google Scholar] [CrossRef]
- Ephros, M.; Engelhard, D.; Maayan, S.; Bercovier, H.; Avital, A.; Yatsiv, I. L. gormanii pneumonia in a child with chronic granulomatous disease. Pediatr. Infect. Dis. J. 1989, 8, 726–727. [Google Scholar]
- Bender, J.; Rydzewski, K.; Broich, M.; Schunder, E.; Heuner, K.; Flieger, A. Phospholipase PlaB of Legionella pneumophila represents a novel lipase family: Protein residues essential for lipolytic activity, substrate specificity, and hemolysis. J. Biol. Chem. 2009, 284, 27185–27194. [Google Scholar] [CrossRef] [Green Version]
- Conover, G.M.; Martinez-Morales, F.; Heidtman, M.I.; Luo, Z.Q.; Tang, M.; Chen, C.; Geiger, O.; Isberg, R.R. Phosphatidylcholine synthesis required for optimal function of Legionella pneumophila virulence determinants. Cell. Microbiol. 2008, 10, 514–528. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Morales, F.; Schobert, M.; Lopez-Lara, I.M.; Geiger, O. Pathways for phosphatidylcholine biosynthesis in bacteria. Microbiology 2003, 149, 3461–3471. [Google Scholar] [CrossRef]
- Palusińska-Szysz, M.; Szuster-Ciesielska, A.; Janczarek, M.; Wdowiak-Wróbel, S.; Schiller, J.; Reszczyńska, E.; Gruszecki, W.I.; Fuchs, B. Genetic diversity of Legionella pcs and pmtA genes and the effect of utilization of choline by Legionella spp. on induction of proinflammatory cytokines. Pathog. Dis. 2019, 77, ftz065. [Google Scholar] [CrossRef]
- Palusińska-Szysz, M.; Zdybicka-Barabas, A.; Pawlikowska-Pawlęga, B.; Mak, P.; Cytryńska, M. Anti-Legionella dumoffii activity of Galleria mellonella defensin and apolipophorin III. Int. J. Mol. Sci. 2012, 13, 17048–17064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmiel, E.; Palusińska-Szysz, M.; Zdybicka-Barabas, A.; Cytryńska, M.; Mak, P. The effect of Galleria mellonella hemolymph polypeptides on Legionella gormanii. Acta Biochim. Pol. 2014, 61, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Aramaki, T.; Blanc-Mathieu, R.; Endo, H.; Ohkubo, K.; Kanehisa, M.; Susumu Goto, G.; Ogata, H. KofamKOALA: KEGG Ortholog assignment based on profile HMM and adaptive score threshold. Bioinformatics 2020, 36, 2251–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yard, B.; Carter, L.; Johnson, K.A.; Overton, I.; Dorward, M.; Liu, H.; McMahon, S.; Oke, M.; Puech, D.; Barton, G.J.; et al. The structure of serine palmitoyltransferase; gateway to sphingolipid biosynthesis. J. Mol. Biol. 2007, 370, 870–886. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [Green Version]
- Rolando, M.; Buchrieser, C. A Comprehensive Review on the Manipulation of the Sphingolipid Pathway by Pathogenic Bacteria. Front. Cell Dev. Biol. 2019, 7, 168. [Google Scholar] [CrossRef]
- Gilchrist, C.L.M.; Chooi, Y.H. Clinker & clustermap.js: Automatic generation of gene cluster comparison figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef] [Green Version]
- Heaver, S.L.; Johnson, E.L.; Ley, R.E. Sphingolipids in host-microbial interactions. Curr. Opin. Microbiol. 2018, 43, 92–99. [Google Scholar] [CrossRef]
- Stoffel, W.; Dittmar, K.; Wilmes, R. Sphingolipid metabolism in Bacteroideaceae. Hoppe-Seyler’s Z. Physiol. Chem. 1975, 35, 6715–6726. [Google Scholar] [CrossRef]
- Cazalet, C.; Rusniok, C.; Brüggemann, H.; Zidane, N.; Magnier, A.; Ma, L.; Tichit, M.; Jarraud, S.; Bouchier, C.; Vandenesch, F.; et al. Evidence in the Legionella pneumophila genome for exploitation of host cell functions and high genome plasticity. Nat. Genet. 2004, 36, 1165–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolando, M.; Escoll, P.; Buchrieser, C. Legionella pneumophila restrains autophagy by modulating the host’s sphingolipid metabolism. Autophagy 2016, 12, 1053–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolando, M.; Escoll, P.; Nora, T.; Botti, J.; Boitez, V.; Bedia, C.; Daniels, C.; Abraham, G.; Stogios, P.J.; Skarina, T.; et al. Legionella pneumophila S1P-lyase targets host sphingolipid metabolism and restrains autophagy. Proc. Natl. Acad. Sci. USA 2016, 113, 1901–1906. [Google Scholar] [CrossRef] [Green Version]
- Bligh, E.G.; Dyer, J.W. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Andersson, B.A. Mass spectrometry of fatty acid pyrrolidides. Prog. Chem. Fats Other Lipids 1978, 16, 279–308. [Google Scholar] [CrossRef]
- Osborn, M.J.; Gander, J.E.; Parisi, E.; Garson, J. Mechanism of assembly of the outer membrane of Salmonella typhimurium. J. Biol. Chem. 1972, 247, 3962–3972. [Google Scholar] [CrossRef]
- Reusch, V.M.; Burger, M.M. Distribution of marker enzymes between mesosomal and protoplast membranes. J. Biol. Chem. 1974, 249, 5337–5345. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding data and analysis capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef] [Green Version]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, 457–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative Content [%] | ||||||
|---|---|---|---|---|---|---|
| Retention Time | Fatty Acid | PE + mMePE | dMePE | PC | PG | CL |
| 9.67 | i14:0 | 0.5 ± 0.2 | 0 | 2.5 ± 0.2 | 0 | tr |
| 10.49 | n14:0 | tr | 1± 0.2 | 0.5 ± 0.2 | 1 ± 0.4 | 1 ± 0.2 |
| 11.81 | i15:0 | tr | 0 | tr | 0.5 ± 0.2 | tr |
| 11.97 | a15:0 | 19 ± 2 | 1 ± 0.3 | 8 ± 2 | 17 ± 2 | 10 ± 0.5 |
| 12.33 | 9 *-15:1 | 0 | 0 | 0 | 0 | 0.6 ± 0.2 |
| 12.62 | n15:0 | 3 ± 0.3 | 1 ± 0.5 | 2 ± 0.2 | 2 ± 0.4 | 2 ± 0.4 |
| 13.90 | i16:0 | 22 ± 0.5 | 1 ± 0.2 | 12 ± 1 | 19 ± 1 | 15 ± 4 |
| 14.25 | cis 9 *-16:1 | 1 ± 0.2 | 1 ± 0.3 | 0.7 ± 0.1 | 1 ± 0.1 | 6 ± 0.7 |
| 14.43 | trans 9 *-16:1 | 2 ± 0.4 | 0 | 1 ± 0.2 | 1 ± 0.6 | 0.5 ± 0.2 |
| 14.66 | n16:0 | 17 ± 0.4 | 25 ± 3 | 18 ± 2 | 17 ± 1 | 14 ± 2 |
| 15.92 | i17:0 | 1 ± 0.1 | 0 | 1± 0.2 | 2 ± 0.4 | 1 ± 0.2 |
| 16.09 | a17:0 | 7 ± 0.5 | 3 ± 0.5 | 5.5 ± 0.8 | 9 ± 0.1 | 7 ± 0.6 |
| 16.467 | c17:0 | 18 ± 1 | 19 ± 2 | 22 ± 2 | 10 ± 1 | 15 ± 1 |
| 16.673 | n17:0 | 3 ± 0.3 | 2 ± 0.2 | 4 ± 0.5 | 3 ± 0.4 | 3 ± 0.2 |
| 18.531 | cis 9 *-18:1 | 0 | 2 ± 0.5 | 0 | tr | 5 ± 0.6 |
| 18.810 | n18:0 | 6 ± 1 | 40 ± 1.5 | 16 ± 1.5 | 13 ± 0.3 | 14 ± 3 |
| 19.690 | i19:0 | 0 | 0 | 2 ± 0 | 1 ± 0.3 | 0.5 ± 0.1 |
| 20.362 | n19:0 | 0.5 ± 0.1 | 0.5 ± 0.2 | 2 ± 0.4 | 2 ± 0.2 | 1 ± 0.2 |
| 20.584 | i20:0 | 0 | 0 | 0 | 0 | 1 ± 0.3 |
| 21.675 | 20:1 | 0 | 0 | 0 | 0 | 1 ± 0.1 |
| 22.100 | n20:0 | tr | 0 | 0.8 ± 0.2 | 0.7 ± 0.4 | 0.9 ± 0.2 |
| 23.740 | 21:0 | tr | 0 | 2 ± 0.8 | 0.8 ± 0.1 | 1 ± 0.2 |
| 25.346 | 22:0 | 0 | 1.5 ± 0.1 | 0 | 0 | tr |
| 28.303 | 24:0 | 0 | 2 ± 0.2 | 0 | 0 | 0.5 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chmiel, E.; Galuska, C.E.; Koper, P.; Kowalczyk, B.; Urbanik-Sypniewska, T.; Palusińska-Szysz, M.; Fuchs, B. Unusual Lipid Components of Legionella gormanii Membranes. Metabolites 2022, 12, 418. https://doi.org/10.3390/metabo12050418
Chmiel E, Galuska CE, Koper P, Kowalczyk B, Urbanik-Sypniewska T, Palusińska-Szysz M, Fuchs B. Unusual Lipid Components of Legionella gormanii Membranes. Metabolites. 2022; 12(5):418. https://doi.org/10.3390/metabo12050418
Chicago/Turabian StyleChmiel, Elżbieta, Christina E. Galuska, Piotr Koper, Bożena Kowalczyk, Teresa Urbanik-Sypniewska, Marta Palusińska-Szysz, and Beate Fuchs. 2022. "Unusual Lipid Components of Legionella gormanii Membranes" Metabolites 12, no. 5: 418. https://doi.org/10.3390/metabo12050418
APA StyleChmiel, E., Galuska, C. E., Koper, P., Kowalczyk, B., Urbanik-Sypniewska, T., Palusińska-Szysz, M., & Fuchs, B. (2022). Unusual Lipid Components of Legionella gormanii Membranes. Metabolites, 12(5), 418. https://doi.org/10.3390/metabo12050418