

The Link between Trace Metal Elements and Glucose Metabolism: Evidence from Zinc, Copper, Iron, and Manganese-Mediated Metabolic Regulation


Abstract

1. Introduction
2. Zinc and Glucose Metabolism

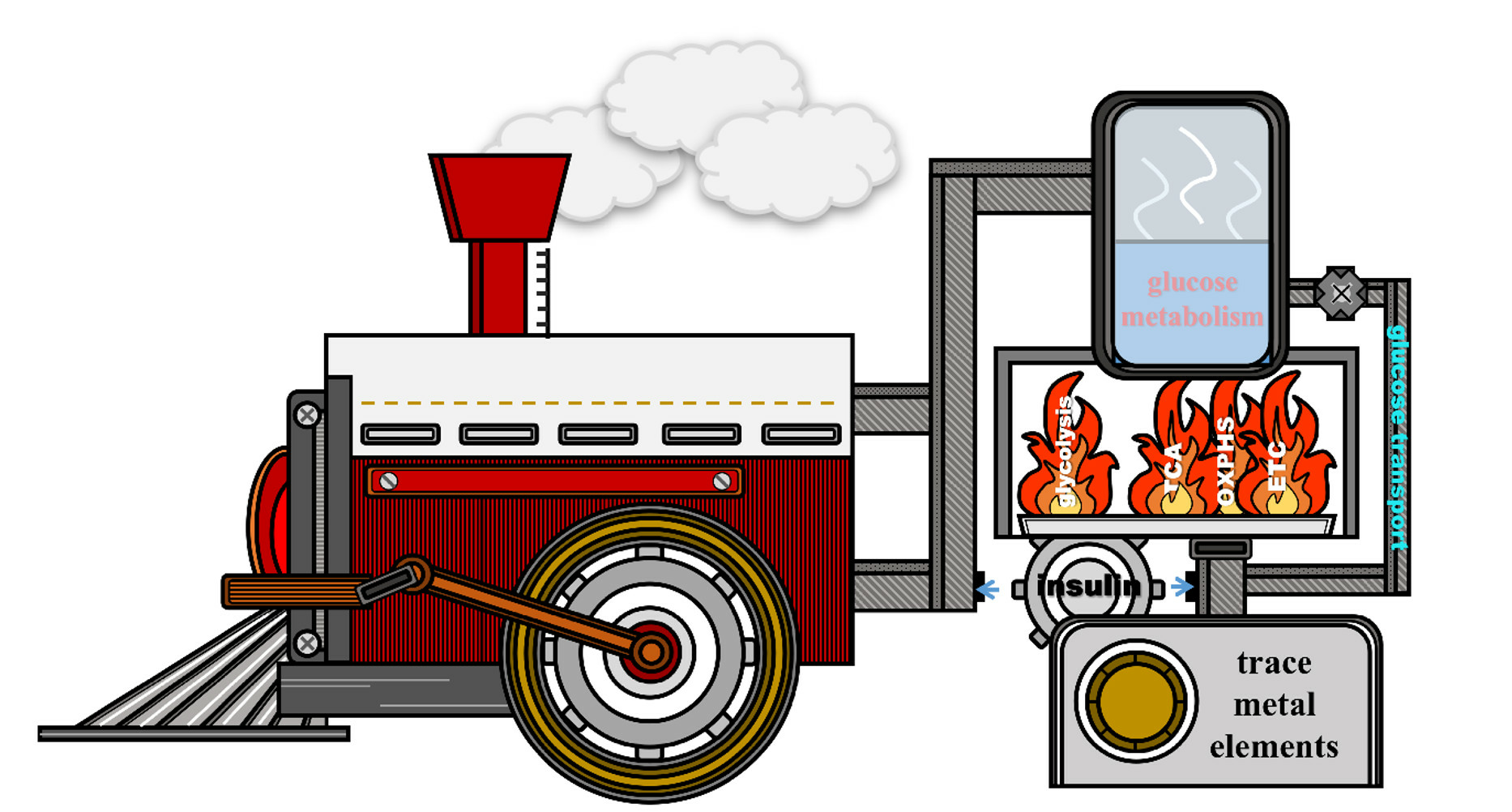{kind=link}
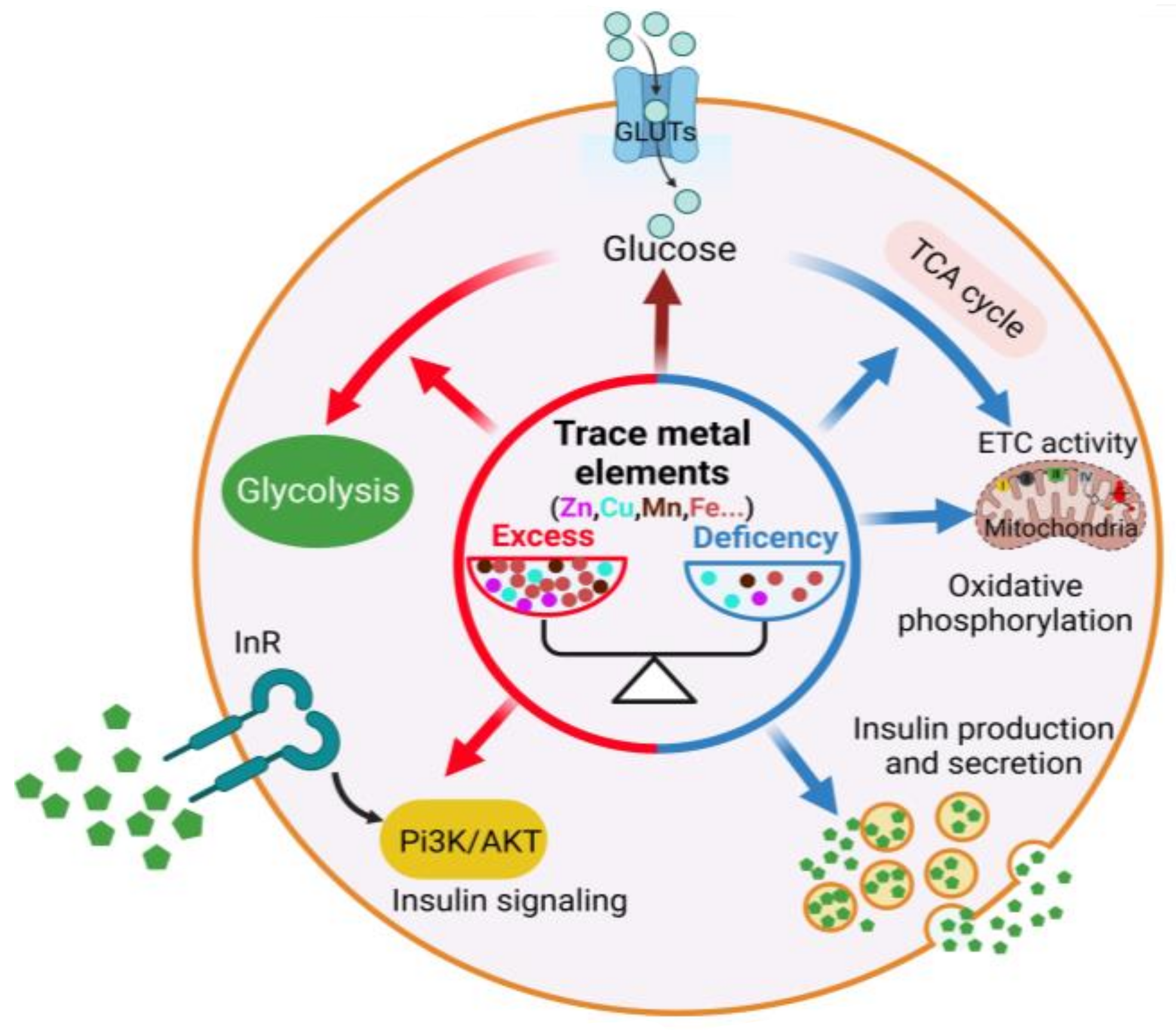{kind=link}
| Metal Elements | Association with Diabetes Mellitus | References |
|---|---|---|
| Zinc | Plasma zinc levels are decreased in diabetes. Increasing zinc levels reduces the risk of T2D 1. Low zinc levels aggravate diabetes. Zinc has curative effects on diabetes. | [33,35] |
| Copper | Plasma copper levels are increased in people with diabetes. High plasma copper levels increase the risk of T2D. Copper reduction can alleviate diabetic symptoms. | [54,55] |
| Manganese | Plasma manganese levels are reduced in T2D patients. Manganese supplementation can effectively protect against T2D. | [56,57] |
| Iron | Iron and ferritin levels are increased in diabetes. Iron overload increases the incidence of T2D. Iron depletion restores insulin sensitivity and reduces plasma glucose levels. | [58,59,60] |
3. Copper and Glucose Metabolism
4. Manganese and Glucose Metabolism
5. Iron and Glucose Metabolism
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AKT\PKB | protein kinase B |
| ATP | adenosine triphosphate |
| ASK | Apoptosis Signal Regulating Kinase |
| Cdkal1 | CDK5 Regulatory Subunit Associated Protein 1-like 1 |
| COX | cytochrome c oxidase |
| CHOP | Human Endoplasmic reticulum stress-related proteins |
| CS | citrate synthase |
| Cyt c | cytochrome c |
| ETC | electron transport chain |
| G6Pase | glucose-6-phosphatase |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| GlUT | glucose transporter |
| GS | glutamine synthetase |
| GSK | glycogen synthase kinase |
| H2O2 | hydrogen peroxide |
| HIF-1α | hypoxia inducible factor-1 |
| IDE | insulin-degrading enzymes |
| IGF-R | IGF receptor |
| InR | insulin receptor |
| IRP2 | iron regulatory protein 2 |
| KGDHC | α-ketoglutarate dehydrogenase complex |
| LDH | lactate dehydrogenase |
| MnP | Mn porphyrin |
| MnSOD | manganese superoxide dismutase |
| MT | metallothionein |
| NADH | nicotinamide adenine dinucleotide |
| NF-κB | NF-kappa B |
| OGG1 | recombinant oxoguanine glycosylase 1 |
| OXPHOS | oxidative phosphorylation |
| PCK1 | phosphoenolpyruvate carboxykinase |
| PDHC | pyruvate dehydrogenase complex |
| PDK1 | pyruvate dehydrogenase kinase 1 |
| PFK1 | phosphofructokinase 1 |
| PGK1 | phosphoglycerate kinase 1 |
| Pi3K | phosphatidylinositol3-kinase |
| PPP | pentose phosphate pathway |
| PTEN | phosphatase and tensin |
| PTPase | protein tyrosine phosphatase |
| ROS | reactive oxygen species |
| SDH | succinate dehydrogenase |
| SNAP25 | synaptosomal associated protein 25 |
| SYT7 | synaptotagmin 7 |
| T2D | type 2 diabetes |
| TCA cycle | tricarboxylic acids cycle |
| TORC1 | target of rapamycin complex 1 |
| TRARG1 | GLUT4-1 trafficking regulator 1 |
| VEGF | vascular endothelial growth factor |
| ZnT | zinc transporter |
References
- Mertz, W. The Essential Trace Elements. Science 1981, 213, 1332–1338. [Google Scholar] [CrossRef]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef]
- He, Z.; Geng, S.; Pan, Y.; Cai, C.; Wang, J.; Wang, L.; Liu, S.; Zheng, P.; Xu, X.; Hu, B. Improvement of the trace metal composition of medium for nitrite-dependent anaerobic methane oxidation bacteria: Iron (II) and copper (II) make a difference. Water Res. 2015, 85, 235–243. [Google Scholar] [CrossRef]
- Wilson, D. The role of zinc in the pathogenicity of human fungal pathogens. Adv. Appl. Microbiol. 2021, 117, 35–61. [Google Scholar]
- Islam, M.R.; Akash, S.; Jony, M.H.; Alam, M.N.; Nowrin, F.T.; Rahman, M.M.; Rauf, A.; Thiruvengadam, M. Exploring the potential function of trace elements in human health: A therapeutic perspective. Mol. Cell. Biochem. 2023, 478, 2141–2171. [Google Scholar] [CrossRef] [PubMed]
- Chasapis, C.T.; Loutsidou, A.C.; Spiliopoulou, C.A.; Stefanidou, M.E. Zinc and human health: An update. Arch. Toxicol. 2012, 86, 521–534. [Google Scholar] [CrossRef]
- Mehri, A. Trace Elements in Human Nutrition (II)—An Update. Int. J. Prev. Med. 2020, 11, 2. [Google Scholar]
- Chen, P.; Bornhorst, J.; Aschner, M. Manganese metabolism in humans. Front. Biosci. 2018, 23, 1655–1679. [Google Scholar] [CrossRef] [PubMed]
- Shribman, S.; Poujois, A.; Bandmann, O.; Czlonkowska, A.; Warner, T.T. Wilson’s disease: Update on pathogenesis, biomarkers and treatments. J. Neurol. Neurosurg. Psychiatry 2021, 92, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, A.; Gadgil, M. Trace metals in cellular metabolism and their impact on recombinant protein production. Process Biochem. 2021, 110, 251–262. [Google Scholar] [CrossRef]
- Chandel, N.S. Carbohydrate Metabolism. Cold Spring Harb. Perspect. Biol. 2021, 13, a040568. [Google Scholar] [CrossRef] [PubMed]
- Norton, L.; Shannon, C.; Gastaldelli, A.; DeFronzo, R.A. Insulin: The master regulator of glucose metabolism. Metabolism 2022, 129, 155142. [Google Scholar] [CrossRef] [PubMed]
- Susnea, I.; Weiskirchen, R. Trace metal imaging in diagnostic of hepatic metal disease. Mass Spectrom. Rev. 2016, 35, 666–686. [Google Scholar] [CrossRef] [PubMed]
- Beard, J.L. Iron biology in immune function, muscle metabolism and neuronal functioning. J. Nutr. 2001, 131, 568S–579S, discussion 580S. [Google Scholar] [CrossRef]
- Potashnik, R.; Kozlovsky, N.; Ben-Ezra, S.; Rudich, A.; Bashan, N. Regulation of glucose transport and GLUT-1 expression by iron chelators in muscle cells in culture. Am. J. Physiol. 1995, 269, E1052–E1058. [Google Scholar] [CrossRef] [PubMed]
- Fukunaka, A.; Fujitani, Y. Role of Zinc Homeostasis in the Pathogenesis of Diabetes and Obesity. Int. J. Mol. Sci. 2018, 19, 476. [Google Scholar] [CrossRef]
- Ruiz, L.M.; Libedinsky, A.; Elorza, A.A. Role of Copper on Mitochondrial Function and Metabolism. Front. Mol. Biosci. 2021, 8, 711227. [Google Scholar] [CrossRef]
- Li, L.; Yang, X. The Essential Element Manganese, Oxidative Stress, and Metabolic Diseases: Links and Interactions. Oxid. Med. Cell. Longev. 2018, 2018, 7580707. [Google Scholar] [CrossRef]
- Oexle, H.; Gnaiger, E.; Weiss, G. Iron-dependent changes in cellular energy metabolism: Influence on citric acid cycle and oxidative phosphorylation. Biochim. Biophys. Acta 1999, 1413, 99–107. [Google Scholar] [CrossRef]
- Wiernsperger, N.; Rapin, J. Trace elements in glucometabolic disorders: An update. Diabetol. Metab. Syndr. 2010, 2, 70. [Google Scholar] [CrossRef]
- Al-Maroof, R.A.; Al-Sharbatti, S.S. Serum zinc levels in diabetic patients and effect of zinc supplementation on glycemic control of type 2 diabetics. Saudi Med. J. 2006, 27, 344–350. [Google Scholar] [PubMed]
- Viktorínová, A.; Toserová, E.; Krizko, M.; Duracková, Z. Altered metabolism of copper, zinc, and magnesium is associated with increased levels of glycated hemoglobin in patients with diabetes mellitus. Metab. Clin. Exp. 2009, 58, 1477–1482. [Google Scholar] [CrossRef] [PubMed]
- Masood, N.; Baloch, G.H.; Ghori, R.A.; Memon, I.A.; Memon, M.A.; Memon, M.S. Serum zinc and magnesium in type-2 diabetic patients. J. Coll. Physicians Surg.-Pak. JCPSP 2009, 19, 483–486. [Google Scholar] [PubMed]
- Kazi, T.G.; Afridi, H.I.; Kazi, N.; Jamali, M.K.; Arain, M.B.; Jalbani, N.; Kandhro, G.A. Copper, chromium, manganese, iron, nickel, and zinc levels in biological samples of diabetes mellitus patients. Biol. Trace Elem. Res. 2008, 122, 1–18. [Google Scholar] [CrossRef]
- Uğurlu, V.; Binay, Ç.; Şimşek, E.; Bal, C. Cellular Trace Element Changes in Type 1 Diabetes Patients. J. Clin. Res. Pediatr. Endocrinol. 2016, 8, 180–186. [Google Scholar] [CrossRef]
- Zhou, Q.; Guo, W.; Jia, Y.; Xu, J. Comparison of Chromium and Iron Distribution in Serum and Urine among Healthy People and Prediabetes and Diabetes Patients. BioMed Res. Int. 2019, 2019, 3801639. [Google Scholar] [CrossRef] [PubMed]
- Bafaro, E.; Liu, Y.; Xu, Y.; Dempski, R.E. The emerging role of zinc transporters in cellular homeostasis and cancer. Signal Transduct. Target. Ther. 2017, 2, 17029. [Google Scholar] [CrossRef]
- Myers, S.A.; Nield, A.; Myers, M. Zinc transporters, mechanisms of action and therapeutic utility: Implications for type 2 diabetes mellitus. J. Nutr. Metab. 2012, 2012, 173712. [Google Scholar] [CrossRef]
- Roohani, N.; Hurrell, R.; Kelishadi, R.; Schulin, R. Zinc and its importance for human health: An integrative review. J. Res. Med. Sci. 2013, 18, 144–157. [Google Scholar]
- Rungby, J. Zinc, zinc transporters and diabetes. Diabetologia 2010, 53, 1549–1551. [Google Scholar] [CrossRef]
- Jansen, J.; Karges, W.; Rink, L. Zinc and diabetes—Clinical links and molecular mechanisms. J. Nutr. Biochem. 2009, 20, 399–417. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Cao, J.C.; Warthon-Medina, M.; Moran, V.H.; Arija, V.; Doepking, C.; Serra-Majem, L.; Lowe, N.M. Zinc Intake and Status and Risk of Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis. Nutrients 2019, 11, 1027. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Zhang, Z.; Liu, S.; Aluo, Z.; Zhang, L.; Yu, L.; Li, Y.; Song, Z.; Zhou, L. Zinc Supplementation Alleviates Lipid and Glucose Metabolic Disorders Induced by a High-Fat Diet. J. Agric. Food Chem. 2020, 68, 5189–5200. [Google Scholar] [CrossRef] [PubMed]
- Bandeira, V.D.S.; Pires, L.V.; Hashimoto, L.L.; Alencar, L.L.; Almondes, K.G.S.; Lottenberg, S.A.; Cozzolino, S.M.F. Association of reduced zinc status with poor glycemic control in individuals with type 2 diabetes mellitus. J. Trace Elem. Med. Biol. 2017, 44, 132–136. [Google Scholar] [CrossRef]
- Jayawardena, R.; Ranasinghe, P.; Galappatthy, P.; Malkanthi, R.; Constantine, G.; Katulanda, P. Effects of zinc supplementation on diabetes mellitus: A systematic review and meta-analysis. Diabetol. Metab. Syndr. 2012, 4, 13. [Google Scholar] [CrossRef]
- Wang, X.; Wu, W.; Zheng, W.; Fang, X.; Chen, L.; Rink, L.; Min, J.; Wang, F. Zinc supplementation improves glycemic control for diabetes prevention and management: A systematic review and meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2019, 110, 76–90. [Google Scholar] [CrossRef]
- Nygaard, S.B.; Larsen, A.; Knuhtsen, A.; Rungby, J.; Smidt, K. Effects of zinc supplementation and zinc chelation on in vitro β-cell function in INS-1E cells. BMC Res. Notes 2014, 7, 84. [Google Scholar] [CrossRef]
- Cooper-Capetini, V.; de Vasconcelos, D.A.A.; Martins, A.R.; Hirabara, S.M.; Donato, J., Jr.; Carpinelli, A.R.; Abdulkader, F. Zinc Supplementation Improves Glucose Homeostasis in High Fat-Fed Mice by Enhancing Pancreatic β-Cell Function. Nutrients 2017, 9, 1150. [Google Scholar] [CrossRef]
- Li, Y.V. Zinc and insulin in pancreatic beta-cells. Endocrine 2014, 45, 178–189. [Google Scholar] [CrossRef]
- Chimienti, F.; Devergnas, S.; Favier, A.; Seve, M. Identification and cloning of a β-cell-specific zinc transporter, ZnT-8, localized into insulin secretory granules. Diabetes 2004, 53, 2330–2337. [Google Scholar] [CrossRef]
- Cruz, K.J.C.; de Oliveira, A.R.S.; Morais, J.B.S.; Severo, J.S.; Mendes, P.M.V.; de Sousa Melo, S.R.; de Sousa, G.S.; Marreiro, D.D.N. Zinc and Insulin Resistance: Biochemical and Molecular Aspects. Biol. Trace Elem. Res. 2018, 186, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Bellomo, E.; Massarotti, A.; Hogstrand, C.; Maret, W. Zinc ions modulate protein tyrosine phosphatase 1B activity. Metallomics 2014, 6, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Barthel, A.; Ostrakhovitch, E.A.; Walter, P.L.; Kampkötter, A.; Klotz, L.O. Stimulation of phosphoinositide 3-kinase/Akt signaling by copper and zinc ions: Mechanisms and consequences. Arch. Biochem. Biophys. 2007, 463, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Norris, D.M.; Humphrey, S.J.; Yang, P.; Cooke, K.C.; Bultitude, W.P.; Parker, B.L.; Conway, O.J.; Burchfield, J.G.; Krycer, J.R.; et al. Trafficking regulator of GLUT4-1 (TRARG1) is a GSK3 substrate. Biochem. J. 2022, 479, 1237–1256. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Di, L.J. Glycogen synthesis and beyond, a comprehensive review of GSK3 as a key regulator of metabolic pathways and a therapeutic target for treating metabolic diseases. Med. Res. Rev. 2022, 42, 946–982. [Google Scholar] [CrossRef]
- Hall, R.K.; Wang, X.L.; George, L.; Koch, S.R.; Granner, D.K. Insulin Represses Phosphoenolpyruvate Carboxykinase Gene Transcription by Causing the Rapid Disruption of an Active Transcription Complex: A Potential Epigenetic Effect. Mol. Endocrinol. 2007, 21, 550–563. [Google Scholar] [CrossRef][Green Version]
- Puigserver, P.; Rhee, J.; Donovan, J.; Walkey, C.J.; Yoon, J.C.; Oriente, F.; Kitamura, Y.; Altomonte, J.; Dong, H.; Accili, D.; et al. Insulin-regulated hepatic gluconeogenesis through FOXO1-PGC-1α interaction. Nature 2003, 423, 550–555. [Google Scholar] [CrossRef]
- Tamaki, N.; Ikeda, T.; Funatsuka, A. Zinc as activating cation for muscle glycolysis. J. Nutr. Sci. Vitaminol. 1983, 29, 655–662. [Google Scholar] [CrossRef]
- Rofe, A.M.; Philcox, J.C.; Coyle, P. Activation of glycolysis by zinc is diminished in hepatocytes from metallothionein-null mice. Biol. Trace Elem. Res. 2000, 75, 87–97. [Google Scholar] [CrossRef]
- Gupta, S.K.; Maggon, K.K.; Venkitasubramanian, T.A. Effect of Zinc on tricarboxylic acid cycle intermediates and enzymes in relation to aflatoxin biosynthesis. J. Gen. Microbiol. 1977, 99, 43–48. [Google Scholar] [CrossRef]
- Yang, X.; Wang, H.; Huang, C.; He, X.; Xu, W.; Luo, Y.; Huang, K. Zinc enhances the cellular energy supply to improve cell motility and restore impaired energetic metabolism in a toxic environment induced by OTA. Sci. Rep. 2017, 7, 14669. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sheng, M.; Wang, J.; Teng, T.; Sun, Y.; Yang, Q.; Xu, Z. Zinc improves mitochondrial respiratory function and prevents mitochondrial ROS generation at reperfusion by phosphorylating STAT3 at Ser(727). J. Mol. Cell. Cardiol. 2018, 118, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Gale, J.R.; Reynolds, I.J.; Weiss, J.H.; Aizenman, E. The Multifaceted Roles of Zinc in Neuronal Mitochondrial Dysfunction. Biomedicines 2021, 9, 489. [Google Scholar] [CrossRef] [PubMed]
- Walter, R.M., Jr.; Uriu-Hare, J.Y.; Olin, K.L.; Oster, M.H.; Anawalt, B.D.; Critchfield, J.W.; Keen, C.L. Copper, zinc, manganese, and magnesium status and complications of diabetes mellitus. Diabetes Care 1991, 14, 1050–1056. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Wang, X.; Li, S.; Zhu, Y.; Chen, S.; Li, P.; Luo, C.; Huang, Y.; Li, X.; Hu, X.; et al. Interactions between plasma copper concentrations and SOD1 gene polymorphism for impaired glucose regulation and type 2 diabetes. Redox Biol. 2019, 24, 101172. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.S.; Kim, S.J.; Yoon, H.E.; Chung, J.H.; Chung, S.; Park, C.W.; Chang, Y.S.; Shin, S.J. Association of blood manganese level with diabetes and renal dysfunction: A cross-sectional study of the Korean general population. BMC Endocr. Disord. 2014, 14, 24. [Google Scholar] [CrossRef]
- Shan, Z.; Chen, S.; Sun, T.; Luo, C.; Guo, Y.; Yu, X.; Yang, W.; Hu, F.B.; Liu, L. U-Shaped Association between Plasma Manganese Levels and Type 2 Diabetes. Environ. Health Perspect. 2016, 124, 1876–1881. [Google Scholar] [CrossRef]
- Hart, P.C.; Mao, M.; de Abreu, A.L.; Ansenberger-Fricano, K.; Ekoue, D.N.; Ganini, D.; Kajdacsy-Balla, A.; Diamond, A.M.; Minshall, R.D.; Consolaro, M.E.; et al. MnSOD upregulation sustains the Warburg effect via mitochondrial ROS and AMPK-dependent signalling in cancer. Nat. Commun. 2015, 6, 6053. [Google Scholar] [CrossRef]
- Klempa, K.L.; Willis, W.T.; Chengson, R.; Dallman, P.R.; Brooks, G.A. Iron deficiency decreases gluconeogenesis in isolated rat hepatocytes. J. Appl. Physiol. 1989, 67, 1868–1872. [Google Scholar] [CrossRef]
- Fernández-Real, J.M.; López-Bermejo, A.; Ricart, W. Cross-talk between iron metabolism and diabetes. Diabetes 2002, 51, 2348–2354. [Google Scholar] [CrossRef]
- Vetchý, M. Biological role of copper as an essential trace element in the human organism. Ceska Slov. Farm. 2018, 67, 143–153. [Google Scholar] [PubMed]
- Xue, Q.; Kang, R.; Klionsky, D.J.; Tang, D.; Liu, J.; Chen, X. Copper metabolism in cell death and autophagy. Autophagy 2023, 19, 2175–2195. [Google Scholar] [CrossRef] [PubMed]
- Nargund, S.; Qiu, J.; Goudar, C.T. Elucidating the role of copper in CHO cell energy metabolism using (13)C metabolic flux analysis. Biotechnol. Prog. 2015, 31, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Gouw, A.M.; LaGory, E.L.; Guo, S.; Attarwala, N.; Tang, Y.; Qi, J.; Chen, Y.S.; Gao, Z.; Casey, K.M.; et al. Mitochondrial copper depletion suppresses triple-negative breast cancer in mice. Nat. Biotechnol. 2021, 39, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Bustos, R.I.; Jensen, E.L.; Ruiz, L.M.; Rivera, S.; Ruiz, S.; Simon, F.; Riedel, C.; Ferrick, D.; Elorza, A.A. Copper deficiency alters cell bioenergetics and induces mitochondrial fusion through up-regulation of MFN2 and OPA1 in erythropoietic cells. Biochem. Biophys. Res. Commun. 2013, 437, 426–432. [Google Scholar] [CrossRef]
- Tavsan, Z.; Ayar Kayali, H. The effect of iron and copper as an essential nutrient on mitochondrial electron transport system and lipid peroxidation in Trichoderma harzianum. Appl. Biochem. Biotechnol. 2013, 170, 1665–1675. [Google Scholar] [CrossRef]
- Lai, J.C.; Blass, J.P. Neurotoxic effects of copper: Inhibition of glycolysis and glycolytic enzymes. Neurochem. Res. 1984, 9, 1699–1710. [Google Scholar] [CrossRef]
- Lauer, M.M.; de Oliveira, C.B.; Yano, N.L.; Bianchini, A. Copper effects on key metabolic enzymes and mitochondrial membrane potential in gills of the estuarine crab Neohelice granulata at different salinities. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2012, 156, 140–147. [Google Scholar] [CrossRef]
- Gebhard, S.; Ronimus, R.S.; Morgan, H.W. Inhibition of phosphofructokinases by copper(II). FEMS Microbiol. Lett. 2001, 197, 105–109. [Google Scholar] [CrossRef][Green Version]
- Tavsan, Z.; Ayar Kayali, H. The Variations of Glycolysis and TCA Cycle Intermediate Levels Grown in Iron and Copper Mediums of Trichoderma harzianum. Appl. Biochem. Biotechnol. 2015, 176, 76–85. [Google Scholar] [CrossRef]
- Li, X.; Qin, Y.; Kong, L.; Yan, X.; Zhang, W.; Martyniuk, C.J.; Wang, X.; Yan, B. Metabolomic and bioenergetic responses of human hepatocellular carcinoma cells following exposure to commercial copper hydroxide nanopesticide. Environ. Sci. Nano 2022, 9, 589–605. [Google Scholar] [CrossRef]
- Tsvetkov, P.; Coy, S.; Petrova, B.; Dreishpoon, M.; Verma, A.; Abdusamad, M.; Rossen, J.; Joesch-Cohen, L.; Humeidi, R.; Spangler, R.D.; et al. Copper induces cell death by targeting lipoylated TCA cycle proteins. Science 2022, 375, 1254–1261. [Google Scholar] [CrossRef]
- Wazir, S.M.; Ghobrial, I. Copper deficiency, a new triad: Anemia, leucopenia, and myeloneuropathy. J. Community Hosp. Intern. Med. Perspect. 2017, 7, 265–268. [Google Scholar] [CrossRef]
- D’Angelo, G. Copper deficiency mimicking myelodysplastic syndrome. Blood Res. 2016, 51, 217–219. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.M.; Kennedy, F.S.; Green, B.G. The effect of iron substrate on mitochondrial haem synthesis in copper deficiency. Br. J. Nutr. 1985, 53, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Ming, J.; Sana, S.; Deng, X. Identification of copper-related biomarkers and potential molecule mechanism in diabetic nephropathy. Front. Endocrinol. 2022, 13, 978601. [Google Scholar] [CrossRef]
- Cunningham, J.; Leffell, M.; Mearkle, P.; Harmatz, P. Elevated plasma ceruloplasmin in insulin-dependent diabetes mellitus: Evidence for increased oxidative stress as a variable complication. Metabolism 1995, 44, 996–999. [Google Scholar] [CrossRef] [PubMed]
- Cooper, G.J.; Chan, Y.K.; Dissanayake, A.M.; Leahy, F.E.; Keogh, G.F.; Frampton, C.M.; Gamble, G.D.; Brunton, D.H.; Baker, J.R.; Poppitt, S.D. Demonstration of a hyperglycemia-driven pathogenic abnormality of copper homeostasis in diabetes and its reversibility by selective chelation: Quantitative comparisons between the biology of copper and eight other nutritionally essential elements in normal and diabetic individuals. Diabetes 2005, 54, 1468–1476. [Google Scholar] [CrossRef]
- Qiu, Q.; Zhang, F.; Zhu, W.; Wu, J.; Liang, M. Copper in Diabetes Mellitus: A Meta-Analysis and Systematic Review of Plasma and Serum Studies. Biol. Trace Elem. Res. 2017, 177, 53–63. [Google Scholar] [CrossRef]
- Tanaka, A.; Kaneto, H.; Miyatsuka, T.; Yamamoto, K.; Yoshiuchi, K.; Yamasaki, Y.; Shimomura, I.; Matsuoka, T.A.; Matsuhisa, M. Role of copper ion in the pathogenesis of type 2 diabetes. Endocr. J. 2009, 56, 699–706. [Google Scholar] [CrossRef]
- Hamann, I.; Petroll, K.; Grimm, L.; Hartwig, A.; Klotz, L.O. Insulin-like modulation of Akt/FoxO signaling by copper ions is independent of insulin receptor. Arch. Biochem. Biophys. 2014, 558, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Ostrakhovitch, E.A.; Lordnejad, M.R.; Schliess, F.; Sies, H.; Klotz, L.O. Copper ions strongly activate the phosphoinositide-3-kinase/Akt pathway independent of the generation of reactive oxygen species. Arch. Biochem. Biophys. 2002, 397, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Schmoll, D.; Walker, K.S.; Alessi, D.R.; Grempler, R.; Burchell, A.; Guo, S.; Walther, R.; Unterman, T.G. Regulation of glucose-6-phosphatase gene expression by protein kinase Bα and the forkhead transcription factor FKHR. Evidence for insulin response unit-dependent and -independent effects of insulin on promoter activity. J. Biol. Chem. 2000, 275, 36324–36333. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Cho, H.; Ryu, S.E.; Choi, M.U. Effects of metal ions on the activity of protein tyrosine phosphatase VHR: Highly potent and reversible oxidative inactivation by Cu2+ ion. Arch. Biochem. Biophys. 2000, 382, 72–80. [Google Scholar] [CrossRef]
- Goldstein, B.J.; Ahmad, F.; Ding, W.; Li, P.M.; Zhang, W.R. Regulation of the insulin signalling pathway by cellular protein-tyrosine phosphatases. Mol. Cell. Biochem. 1998, 182, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.B.; Guo, L.; Liu, Y.; Zhou, S.R.; Liu, Y.; Dou, X.; Du, S.Y.; Ding, M.; Peng, W.Q.; Qian, S.W.; et al. Synthesis of cytochrome c oxidase 1 (SCO1) inhibits insulin sensitivity by decreasing copper levels in adipocytes. Biochem. Biophys. Res. Commun. 2017, 491, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Tolbert, M.E.; Kamalu, J.A.; Draper, G.D. Effects of cadmium, zinc, copper and manganese on hepatic parenchymal cell gluconeogenesis. J. Environ. Sci. Health B 1981, 16, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Grasso, G.; Salomone, F.; Tundo, G.R.; Pappalardo, G.; Ciaccio, C.; Spoto, G.; Pietropaolo, A.; Coletta, M.; Rizzarelli, E. Metal ions affect insulin-degrading enzyme activity. J. Inorg. Biochem. 2012, 117, 351–358. [Google Scholar] [CrossRef][Green Version]
- Bellia, F.; Lanza, V.; Ahmed, I.M.M.; Garcia-Vinuales, S.; Veiss, E.; Arizzi, M.; Calcagno, D.; Milardi, D.; Grasso, G. Site directed mutagenesis of insulin-degrading enzyme allows singling out the molecular basis of peptidase versus E1-like activity: The role of metal ions. Metallomics 2019, 11, 278–281. [Google Scholar] [CrossRef]
- Grasso, G.; Pietropaolo, A.; Spoto, G.; Pappalardo, G.; Tundo, G.R.; Ciaccio, C.; Coletta, M.; Rizzarelli, E. Copper(I) and copper(II) inhibit Aβ peptides proteolysis by insulin-degrading enzyme differently: Implications for metallostasis alteration in Alzheimer’s disease. Chemistry 2011, 17, 2752–2762. [Google Scholar] [CrossRef]
- Maianti, J.P.; McFedries, A.; Foda, Z.H.; Kleiner, R.E.; Du, X.Q.; Leissring, M.A.; Tang, W.J.; Charron, M.J.; Seeliger, M.A.; Saghatelian, A.; et al. Anti-diabetic activity of insulin-degrading enzyme inhibitors mediated by multiple hormones. Nature 2014, 511, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Yuk, I.H.; Zhang, J.D.; Ebeling, M.; Berrera, M.; Gomez, N.; Werz, S.; Meiringer, C.; Shao, Z.; Swanberg, J.C.; Lee, K.H.; et al. Effects of copper on CHO cells: Insights from gene expression analyses. Biotechnol. Prog. 2014, 30, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, W.; Kang, Y.J. Copper affects the binding of HIF-1α to the critical motifs of its target genes. Metallomics 2019, 11, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Ishida, S.; Andreux, P.; Poitry-Yamate, C.; Auwerx, J.; Hanahan, D. Bioavailable copper modulates oxidative phosphorylation and growth of tumors. Proc. Natl. Acad. Sci. USA 2013, 110, 19507–19512. [Google Scholar] [CrossRef] [PubMed]
- Ramchandani, D.; Berisa, M.; Tavarez, D.A.; Li, Z.; Miele, M.; Bai, Y.; Lee, S.B.; Ban, Y.; Dephoure, N.; Hendrickson, R.C.; et al. Copper depletion modulates mitochondrial oxidative phosphorylation to impair triple negative breast cancer metastasis. Nat. Commun. 2021, 12, 7311. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.L.; Gonzalez-Ibanez, A.M.; Mendoza, P.; Ruiz, L.M.; Riedel, C.A.; Simon, F.; Schuringa, J.J.; Elorza, A.A. Copper deficiency-induced anemia is caused by a mitochondrial metabolic reprograming in erythropoietic cells. Metallomics 2019, 11, 282–290. [Google Scholar] [CrossRef]
- Ruiz, L.M.; Jensen, E.L.; Rossel, Y.; Puas, G.I.; Gonzalez-Ibanez, A.M.; Bustos, R.I.; Ferrick, D.A.; Elorza, A.A. Non-cytotoxic copper overload boosts mitochondrial energy metabolism to modulate cell proliferation and differentiation in the human erythroleukemic cell line K562. Mitochondrion 2016, 29, 18–30. [Google Scholar] [CrossRef]
- Rozenberg, J.M.; Kamynina, M.; Sorokin, M.; Zolotovskaia, M.; Koroleva, E.; Kremenchutckaya, K.; Gudkov, A.; Buzdin, A.; Borisov, N. The Role of the Metabolism of Zinc and Manganese Ions in Human Cancerogenesis. Biomedicines 2022, 10, 1072. [Google Scholar] [CrossRef]
- Hurley, L.S.; Keen, C.L.; Baly, D.L. Manganese deficiency and toxicity: Effects on carbohydrate metabolism in the rat. Neurotoxicology 1984, 5, 97–104. [Google Scholar]
- Flores, C.R.; Puga, M.P.; Wrobel, K.; Garay Sevilla, M.E.; Wrobel, K. Trace elements status in diabetes mellitus type 2: Possible role of the interaction between molybdenum and copper in the progress of typical complications. Diabetes Res. Clin. Pract. 2011, 91, 333–341. [Google Scholar] [CrossRef]
- Bresciani, G.; Cruz, I.B.; de Paz, J.A.; Cuevas, M.J.; González-Gallego, J. The MnSOD Ala16Val SNP: Relevance to human diseases and interaction with environmental factors. Free Radic. Res. 2013, 47, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Jouihan, H.A.; Cooksey, R.C.; Jones, D.; Kim, H.J.; Winge, D.R.; McClain, D.A. Manganese supplementation protects against diet-induced diabetes in wild type mice by enhancing insulin secretion. Endocrinology 2013, 154, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Burlet, E.; Jain, S.K. Manganese supplementation reduces high glucose-induced monocyte adhesion to endothelial cells and endothelial dysfunction in Zucker diabetic fatty rats. J. Biol. Chem. 2013, 288, 6409–6416. [Google Scholar] [CrossRef] [PubMed]
- Baly, D.L.; Curry, D.L.; Keen, C.L.; Hurley, L.S. Effect of manganese deficiency on insulin secretion and carbohydrate homeostasis in rats. J. Nutr. 1984, 114, 1438–1446. [Google Scholar] [CrossRef] [PubMed]
- Baly, D.L.; Curry, D.L.; Keen, C.L.; Hurley, L.S. Dynamics of insulin and glucagon release in rats: Influence of dietary manganese. Endocrinology 1985, 116, 1734–1740. [Google Scholar] [CrossRef]
- Bryan, M.R.; Uhouse, M.A.; Nordham, K.D.; Joshi, P.; Rose, D.I.R.; O’Brien, M.T.; Aschner, M.; Bowman, A.B. Phosphatidylinositol 3 kinase (PI3K) modulates manganese homeostasis and manganese-induced cell signaling in a murine striatal cell line. Neurotoxicology 2018, 64, 185–194. [Google Scholar] [CrossRef]
- Bryan, M.R.; Nordham, K.D.; Rose, D.I.R.; O’Brien, M.T.; Joshi, P.; Foshage, A.M.; Gonçalves, F.M.; Nitin, R.; Uhouse, M.A.; Aschner, M.; et al. Manganese Acts upon Insulin/IGF Receptors to Phosphorylate AKT and Increase Glucose Uptake in Huntington’s Disease Cells. Mol. Neurobiol. 2020, 57, 1570–1593. [Google Scholar] [CrossRef]
- Ueda, M.; Robinson, F.W.; Smith, M.M.; Kono, T. Effects of divalent cations on the regulation of insulin-sensitive glucose transport and cAMP phosphodiesterase in adipocytes. Insulin-like effects of divalent cations. J. Biol. Chem. 1984, 259, 9520–9525. [Google Scholar] [CrossRef]
- Baly, D.L.; Schneiderman, J.S.; Garcia-Welsh, A.L. Effect of manganese deficiency on insulin binding, glucose transport and metabolism in rat adipocytes. J. Nutr. 1990, 120, 1075–1079. [Google Scholar] [CrossRef]
- Nicastro, R.; Gaillard, H.; Zarzuela, L.; Péli-Gulli, M.P.; Fernández-García, E.; Tomé, M.; García-Rodríguez, N.; Durán, R.V.; De Virgilio, C.; Wellinger, R.E. Manganese is a physiologically relevant TORC1 activator in yeast and mammals. eLife 2022, 11, e80497. [Google Scholar] [CrossRef]
- Mao, Z.; Zhang, W. Role of mTOR in Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2018, 19, 2043. [Google Scholar] [CrossRef]
- Malthankar, G.V.; White, B.K.; Bhushan, A.; Daniels, C.K.; Rodnick, K.J.; Lai, J.C.K. Differential lowering by manganese treatment of activities of glycolytic and tricarboxylic acid (TCA) cycle enzymes investigated in neuroblastoma and astrocytoma cells is associated with manganese-induced cell death. Neurochem. Res. 2004, 29, 709–717. [Google Scholar] [CrossRef]
- Wimhurst, J.M.; Manchester, K.L. Effects of manganese on the activity of glycolytic and gluconeogenic enzymes in the perfused rat liver. FEBS Lett. 1973, 29, 201–203. [Google Scholar] [CrossRef]
- Delmastro-Greenwood, M.M.; Votyakova, T.; Goetzman, E.; Marre, M.L.; Previte, D.M.; Tovmasyan, A.; Batinic-Haberle, I.; Trucco, M.M.; Piganelli, J.D. Mn porphyrin regulation of aerobic glycolysis: Implications on the activation of diabetogenic immune cells. Antioxid. Redox Signal. 2013, 19, 1902–1915. [Google Scholar] [CrossRef] [PubMed]
- Zwingmann, C.; Leibfritz, D.; Hazell, A.S. Energy metabolism in astrocytes and neurons treated with manganese: Relation among cell-specific energy failure, glucose metabolism, and intercellular trafficking using multinuclear NMR-spectroscopic analysis. J. Cereb. Blood Flow Metab. 2003, 23, 756–771. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Xu, Z.; Gao, J.; Xu, B.; Deng, Y. In vitro effect of manganese chloride exposure on energy metabolism and oxidative damage of mitochondria isolated from rat brain. Environ. Toxicol. Pharmacol. 2008, 26, 232–236. [Google Scholar] [CrossRef]
- Kaur, G.; Kumar, V.; Arora, A.; Tomar, A.; Ashish; Sur, R.; Dutta, D. Affected energy metabolism under manganese stress governs cellular toxicity. Sci. Rep. 2017, 7, 11645. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS Signaling: The New Wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef]
- Lee, S.; Tak, E.; Lee, J.; Rashid, M.A.; Murphy, M.P.; Ha, J.; Kim, S.S. Mitochondrial H2O2 generated from electron transport chain complex I stimulates muscle differentiation. Cell Res. 2011, 21, 817–834. [Google Scholar] [CrossRef] [PubMed]
- Raffield, L.M.; Louie, T.; Sofer, T.; Jain, D.; Ipp, E.; Taylor, K.D.; Papanicolaou, G.J.; Avilés-Santa, L.; Lange, L.A.; Laurie, C.C.; et al. Genome-wide association study of iron traits and relation to diabetes in the Hispanic Community Health Study/Study of Latinos (HCHS/SOL): Potential genomic intersection of iron and glucose regulation? Hum. Mol. Genet. 2017, 26, 1966–1978. [Google Scholar] [CrossRef]
- Simcox, J.A.; McClain, D.A. Iron and diabetes risk. Cell Metab. 2013, 17, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Real, J.M.; Manco, M. Effects of iron overload on chronic metabolic diseases. Lancet Diabetes Endocrinol. 2014, 2, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Sun, L.; Tan, Y.; Wang, G.; Lin, X.; Cai, L. Role of iron deficiency and overload in the pathogenesis of diabetes and diabetic complications. Curr. Med. Chem. 2009, 16, 113–129. [Google Scholar] [CrossRef]
- Kranz, R.G.; Richard-Fogal, C.; Taylor, J.S.; Frawley, E.R. Cytochrome c biogenesis: Mechanisms for covalent modifications and trafficking of heme and for heme-iron redox control. Microbiol. Mol. Biol. Rev. 2009, 73, 510–528. [Google Scholar] [CrossRef]
- Hagler, L.; Askew, E.W.; Neville, J.R.; Mellick, P.W.; Coppes, R.I., Jr.; Lowder, J.F., Jr. Influence of dietary iron deficiency on hemoglobin, myoglobin, their respective reductases, and skeletal muscle mitochondrial respiration. Am. J. Clin. Nutr. 1981, 34, 2169–2177. [Google Scholar] [CrossRef] [PubMed]
- Frise, M.C.; Holdsworth, D.A.; Johnson, A.W.; Chung, Y.J.; Curtis, M.K.; Cox, P.J.; Clarke, K.; Tyler, D.J.; Roberts, D.J.; Ratcliffe, P.J.; et al. Abnormal whole-body energy metabolism in iron-deficient humans despite preserved skeletal muscle oxidative phosphorylation. Sci. Rep. 2022, 12, 998. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.J.; Swietach, P.; Curtis, M.K.; Ball, V.; Robbins, P.A.; Lakhal-Littleton, S. Iron-Deficiency Anemia Results in Transcriptional and Metabolic Remodeling in the Heart Toward a Glycolytic Phenotype. Front. Cardiovasc. Med. 2020, 7, 616920. [Google Scholar] [CrossRef]
- Stugiewicz, M.; Tkaczyszyn, M.; Kasztura, M.; Banasiak, W.; Ponikowski, P.; Jankowska, E.A. The influence of iron deficiency on the functioning of skeletal muscles: Experimental evidence and clinical implications. Eur. J. Heart Fail. 2016, 18, 762–773. [Google Scholar] [CrossRef]
- Quail, E.A.; Yeoh, G.C. The effect of iron status on glyceraldehyde 3-phosphate dehydrogenase expression in rat liver. FEBS Lett. 1995, 359, 126–128. [Google Scholar] [CrossRef][Green Version]
- Li, J.; Jia, L.; Ma, W.; Feng, Y.; Yu, H.; Du, H. Dietary iron modulates hepatic glucose homeostasis via regulating gluconeogenesis. J. Nutr. Biochem. 2022, 109, 109104. [Google Scholar] [CrossRef]
- Ohira, Y.; Chen, C.S.; Hegenauer, J.; Saltman, P. Adaptations of lactate metabolism in iron-deficient rats. Proc. Soc. Exp. Biol. Med. 1983, 173, 213–216. [Google Scholar] [CrossRef]
- Henderson, S.A.; Dallman, P.R.; Brooks, G.A. Glucose turnover and oxidation are increased in the iron-deficient anemic rat. Am. J. Physiol. 1986, 250, E414–E421. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, Q.; Yang, Y.; Ma, L. Iron metabolism and type 2 diabetes mellitus: A meta-analysis and systematic review. J. Diabetes Investig. 2020, 11, 946–955. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Fang, A.; Yu, S.; Shen, X.; Li, K. Dietary Nonheme, Heme, and Total Iron Intake and the Risk of Diabetes in Adults: Results from the China Health and Nutrition Survey. Diabetes Care 2020, 43, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Downes, K.; Howson, J.M.; Nutland, S.; Stevens, H.E.; Walker, N.M.; Todd, J.A. Evidence of association with type 1 diabetes in the SLC11A1 gene region. BMC Med. Genet. 2011, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- Kavian, Z.; Sargazi, S.; Majidpour, M.; Sarhadi, M.; Saravani, R.; Shahraki, M.; Mirinejad, S.; Heidari Nia, M.; Piri, M. Association of SLC11A1 polymorphisms with anthropometric and biochemical parameters describing Type 2 Diabetes Mellitus. Sci. Rep. 2023, 13, 6195. [Google Scholar] [CrossRef]
- Tilbrook, L. Cross talk between iron metabolism and diabetes. Ann. Clin. Biochem. 2004, 41, 255. [Google Scholar]
- Dandona, P.; Hussain, M.A.; Varghese, Z.; Politis, D.; Flynn, D.M.; Hoffbrand, A.V. Insulin resistance and iron overload. Ann. Clin. Biochem. 1983, 20, 77–79. [Google Scholar] [CrossRef]
- Hua, N.W.; Stoohs, R.A.; Facchini, F.S. Low iron status and enhanced insulin sensitivity in lacto-ovo vegetarians. Br. J. Nutr. 2001, 86, 515–519. [Google Scholar] [CrossRef]
- Messner, D.J.; Rhieu, B.H.; Kowdley, K.V. Iron overload causes oxidative stress and impaired insulin signaling in AML-12 hepatocytes. Dig. Dis. Sci. 2013, 58, 1899–1908. [Google Scholar] [CrossRef]
- Fargion, S.; Dongiovanni, P.; Guzzo, A.; Colombo, S.; Valenti, L.; Fracanzani, A.L. Iron and insulin resistance. Aliment. Pharmacol. Ther. 2005, 22 (Suppl. 2), 61–63. [Google Scholar] [CrossRef]
- Dongiovanni, P.; Valenti, L.; Ludovica Fracanzani, A.; Gatti, S.; Cairo, G.; Fargion, S. Iron depletion by deferoxamine up-regulates glucose uptake and insulin signaling in hepatoma cells and in rat liver. Am. J. Pathol. 2008, 172, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Mehdad, A.; Campos, N.A.; Arruda, S.F.; Siqueira, E.M. Iron Deprivation May Enhance Insulin Receptor and Glut4 Transcription in Skeletal Muscle of Adult Rats. J. Nutr. Health Aging 2015, 19, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Ma, Y.; Shi, M.; Huang, M.; Xin, J.; Ci, S.; Chen, M.; Jiang, T.; Hu, Z.; He, L.; et al. Excessive iron inhibits insulin secretion via perturbing transcriptional regulation of SYT7 by OGG1. Cell. Mol. Life Sci. 2023, 80, 159. [Google Scholar] [CrossRef]
- Blesia, V.; Patel, V.B.; Al-Obaidi, H.; Renshaw, D.; Zariwala, M.G. Excessive Iron Induces Oxidative Stress Promoting Cellular Perturbations and Insulin Secretory Dysfunction in MIN6 Beta Cells. Cells 2021, 10, 1141. [Google Scholar] [CrossRef]
- Li, W.; Feng, Q.; Wang, C.; Yin, Z.; Li, X.; Li, L. LncXIST Facilitates Iron Overload and Iron Overload-Induced Islet Beta Cell Injury in Type 2 Diabetes through miR-130a-3p/ALK2 Axis. Comput. Intell. Neurosci. 2022, 2022, 6390812. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Mo, M.Q.; Zhong, J.; Li, Z.; Li, G.; Liang, Y. Iron overload induces islet β cell ferroptosis by activating ASK1/P-P38/CHOP signaling pathway. PeerJ 2023, 11, e15206. [Google Scholar] [CrossRef]
- Hansen, J.B.; Tonnesen, M.F.; Madsen, A.N.; Hagedorn, P.H.; Friberg, J.; Grunnet, L.G.; Heller, R.S.; Nielsen, A.; Størling, J.; Baeyens, L.; et al. Divalent metal transporter 1 regulates iron-mediated ROS and pancreatic β cell fate in response to cytokines. Cell Metab. 2012, 16, 449–461. [Google Scholar] [CrossRef]
- Santos, M.; Anderson, C.P.; Neschen, S.; Zumbrennen-Bullough, K.B.; Romney, S.J.; Kahle-Stephan, M.; Rathkolb, B.; Gailus-Durner, V.; Fuchs, H.; Wolf, E.; et al. Irp2 regulates insulin production through iron-mediated Cdkal1-catalyzed tRNA modification. Nat. Commun. 2020, 11, 296. [Google Scholar] [CrossRef]
- Ashok, A.; Singh, N. Prion protein modulates glucose homeostasis by altering intracellular iron. Sci. Rep. 2018, 8, 6556. [Google Scholar] [CrossRef]
- Higashida, K.; Takeuchi, N.; Inoue, S.; Hashimoto, T.; Nakai, N. Iron deficiency attenuates catecholamine-stimulated lipolysis via downregulation of lipolysis-related proteins and glucose utilization in 3T3-L1 adipocytes. Mol. Med. Rep. 2020, 21, 1383–1389. [Google Scholar] [CrossRef]
- Fillebeen, C.; Lam, N.H.; Chow, S.; Botta, A.; Sweeney, G.; Pantopoulos, K. Regulatory Connections between Iron and Glucose Metabolism. Int. J. Mol. Sci. 2020, 21, 7773. [Google Scholar] [CrossRef]
- De Meyts, P. The Insulin Receptor and Its Signal Transduction Network. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Chang, L.; Chiang, S.H.; Saltiel, A.R. Insulin signaling and the regulation of glucose transport. Mol. Med. 2004, 10, 65–71. [Google Scholar] [CrossRef] [PubMed]
- De Romaña, D.L.; Olivares, M.; Uauy, R.; Araya, M. Risks and benefits of copper in light of new insights of copper homeostasis. J. Trace Elem. Med. Biol. 2011, 25, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Dallinger, R. Invertebrate organisms as biological indicators of heavy metal pollution. Appl. Biochem. Biotechnol. 1994, 48, 27–31. [Google Scholar] [CrossRef]
- Michalke, B.; Fernsebner, K. New insights into manganese toxicity and speciation. J. Trace Elem. Med. Biol. 2014, 28, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Gong, D.; Choong, S.; Xu, H.; Chan, Y.; Chen, X.; Fitzpatrick, S.; Glyn-Jones, S.; Zhang, S.; Nakamura, T. Copper (II)-selective chelation improves function and antioxidant defences in cardiovascular tissues of rats as a model of diabetes: Comparisons between triethylenetetramine and three less copper-selective transition-metal-targeted treatments. Diabetologia 2010, 53, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Lu, J.; Chen, X.; Reddy, S.; Crossman, D.; Glyn-Jones, S.; Choong, Y.-S.; Kennedy, J.; Barry, B.; Zhang, S. A copper (II)-selective chelator ameliorates diabetes-evoked renal fibrosis and albuminuria, and suppresses pathogenic TGF-β activation in the kidneys of rats used as a model of diabetes. Diabetologia 2008, 51, 1741–1751. [Google Scholar] [CrossRef]
- Song, E.; Vu, V.; Varin, T.V.; Botta, A.; Marette, A.; Sweeney, G. Copper fabric improves the metabolic profile of obese mice: Potential role of the gut microbiota. Basic Clin. Pharmacol. Toxicol. 2022, 131, 355–363. [Google Scholar] [CrossRef]
- Sun, Q.; Van Dam, R.M.; Willett, W.C.; Hu, F.B. Prospective study of zinc intake and risk of type 2 diabetes in women. Diabetes Care 2009, 32, 629–634. [Google Scholar] [CrossRef]
- Laouali, N.; MacDonald, C.-J.; Shah, S.; El Fatouhi, D.; Mancini, F.R.; Fagherazzi, G.; Boutron-Ruault, M.-C. Dietary copper/zinc ratio and type 2 diabetes risk in women: The E3N cohort study. Nutrients 2021, 13, 2502. [Google Scholar] [CrossRef] [PubMed]
- Gunasekara, P.; Hettiarachchi, M.; Liyanage, C.; Lekamwasam, S. Effects of zinc and multimineral vitamin supplementation on glycemic and lipid control in adult diabetes. Diabetes Metab. Syndr. Obes. Targets Ther. 2011, 4, 53–60. [Google Scholar]
- Zhu, K.; Nie, S.; Li, C.; Huang, J.; Hu, X.; Li, W.; Gong, D.; Xie, M. Antidiabetic and pancreas-protective effects of zinc threoninate chelate in diabetic rats may be associated with its antioxidative stress ability. Biol. Trace Elem. Res. 2013, 153, 291–298. [Google Scholar] [CrossRef]
- Karatug, A.; Kaptan, E.; Bolkent, S.; Mutlu, O.; Yanardag, R. Alterations in kidney tissue following zinc supplementation to STZ-induced diabetic rats. J. Trace Elem. Med. Biol. 2013, 27, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Cutler, P. Deferoxamine therapy in high-ferritin diabetes. Diabetes 1989, 38, 1207–1210. [Google Scholar] [CrossRef] [PubMed]
- Cooksey, R.C.; Jones, D.; Gabrielsen, S.; Huang, J.; Simcox, J.A.; Luo, B.; Soesanto, Y.; Rienhoff, H.; Dale Abel, E.; McClain, D.A. Dietary iron restriction or iron chelation protects from diabetes and loss of β-cell function in the obese (ob/ob lep−/−) mouse. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E1236–E1243. [Google Scholar] [CrossRef]
- Tajima, S.; Ikeda, Y.; Sawada, K.; Yamano, N.; Horinouchi, Y.; Kihira, Y.; Ishizawa, K.; Izawa-Ishizawa, Y.; Kawazoe, K.; Tomita, S. Iron reduction by deferoxamine leads to amelioration of adiposity via the regulation of oxidative stress and inflammation in obese and type 2 diabetes KKAy mice. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E77–E86. [Google Scholar] [CrossRef]
- Jirakittidul, P.; Sirichotiyakul, S.; Ruengorn, C.; Siripenpong, S.; Imruetaicharoenchok, A.; Wiriyasirivaj, B. Iron supplementation in non-anemic pregnancy and risk of developing gestational diabetes mellitus. J. Endocrinol. Metab. 2019, 8, 139–143. [Google Scholar] [CrossRef]
- Burlet, E.; Jain, S.K. Manganese supplementation increases adiponectin and lowers ICAM-1 and creatinine blood levels in Zucker type 2 diabetic rats, and downregulates ICAM-1 by upregulating adiponectin multimerization protein (DsbA-L) in endothelial cells. Mol. Cell. Biochem. 2017, 429, 1–10. [Google Scholar] [CrossRef]
- Du, S.; Wu, X.; Han, T.; Duan, W.; Liu, L.; Qi, J.; Niu, Y.; Na, L.; Sun, C. Dietary manganese and type 2 diabetes mellitus: Two prospective cohort studies in China. Diabetologia 2018, 61, 1985–1995. [Google Scholar] [CrossRef]

| Sample | Trace Metal Elements | Concentration | References | |
|---|---|---|---|---|
| Healthy People | Diabetic Patients | |||
| Plasma | Zinc | 12.2–16.9 μmol/L | 10.2–16.3 μmol/L | [21,22,23] |
| Copper | 14.6–19.8 μmol/L | 16.4–21.5 μmol/L | [22] | |
| Manganese | 0.77–1.18 μmol/L | 0.62–0.93 μmol/L | [22,23,24,25] | |
| Iron | Male: 11–31 μmol/L, Female: 9–30 μmol/L | Male: 15–33 μmol/L Female: 12–32 μmol/L | [24,26] | |
| Urine | Zinc | 300–600 μg/24 h 1 | 100–300 μg/24 h | [24] |
| Copper | 15–60 μg/24 h | 30–100 μg/24 h | [24] | |
| Manganese | 1–8 μg/24 h | 2–15 μg/24 h | [24,25] | |
| Iron | 0.2–1.1 μmol/24 h | 0.5–3.5 μmol/24 h | [26] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Shao, Y.; Yan, K.; Yao, T.; Liu, L.; Sun, F.; Wu, J.; Huang, Y. The Link between Trace Metal Elements and Glucose Metabolism: Evidence from Zinc, Copper, Iron, and Manganese-Mediated Metabolic Regulation. Metabolites 2023, 13, 1048. https://doi.org/10.3390/metabo13101048
Sun Z, Shao Y, Yan K, Yao T, Liu L, Sun F, Wu J, Huang Y. The Link between Trace Metal Elements and Glucose Metabolism: Evidence from Zinc, Copper, Iron, and Manganese-Mediated Metabolic Regulation. Metabolites. 2023; 13(10):1048. https://doi.org/10.3390/metabo13101048
Chicago/Turabian StyleSun, Zhendong, Yuzhuo Shao, Kunhao Yan, Tianzhao Yao, Lulu Liu, Feifei Sun, Jiarui Wu, and Yunpeng Huang. 2023. "The Link between Trace Metal Elements and Glucose Metabolism: Evidence from Zinc, Copper, Iron, and Manganese-Mediated Metabolic Regulation" Metabolites 13, no. 10: 1048. https://doi.org/10.3390/metabo13101048
APA StyleSun, Z., Shao, Y., Yan, K., Yao, T., Liu, L., Sun, F., Wu, J., & Huang, Y. (2023). The Link between Trace Metal Elements and Glucose Metabolism: Evidence from Zinc, Copper, Iron, and Manganese-Mediated Metabolic Regulation. Metabolites, 13(10), 1048. https://doi.org/10.3390/metabo13101048