

Transcriptomic Analysis Reveals the Response Mechanisms of Bell Pepper (Capsicum annuum) to Phosphorus Deficiency
 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. RNA Extraction and RNA Sequencing
2.3. Bioinformatic Data Analysis
2.4. Real-Time PCR Analysis of Candidate Genes
2.5. GO and KEGG Enrichment Analysis
3. Results
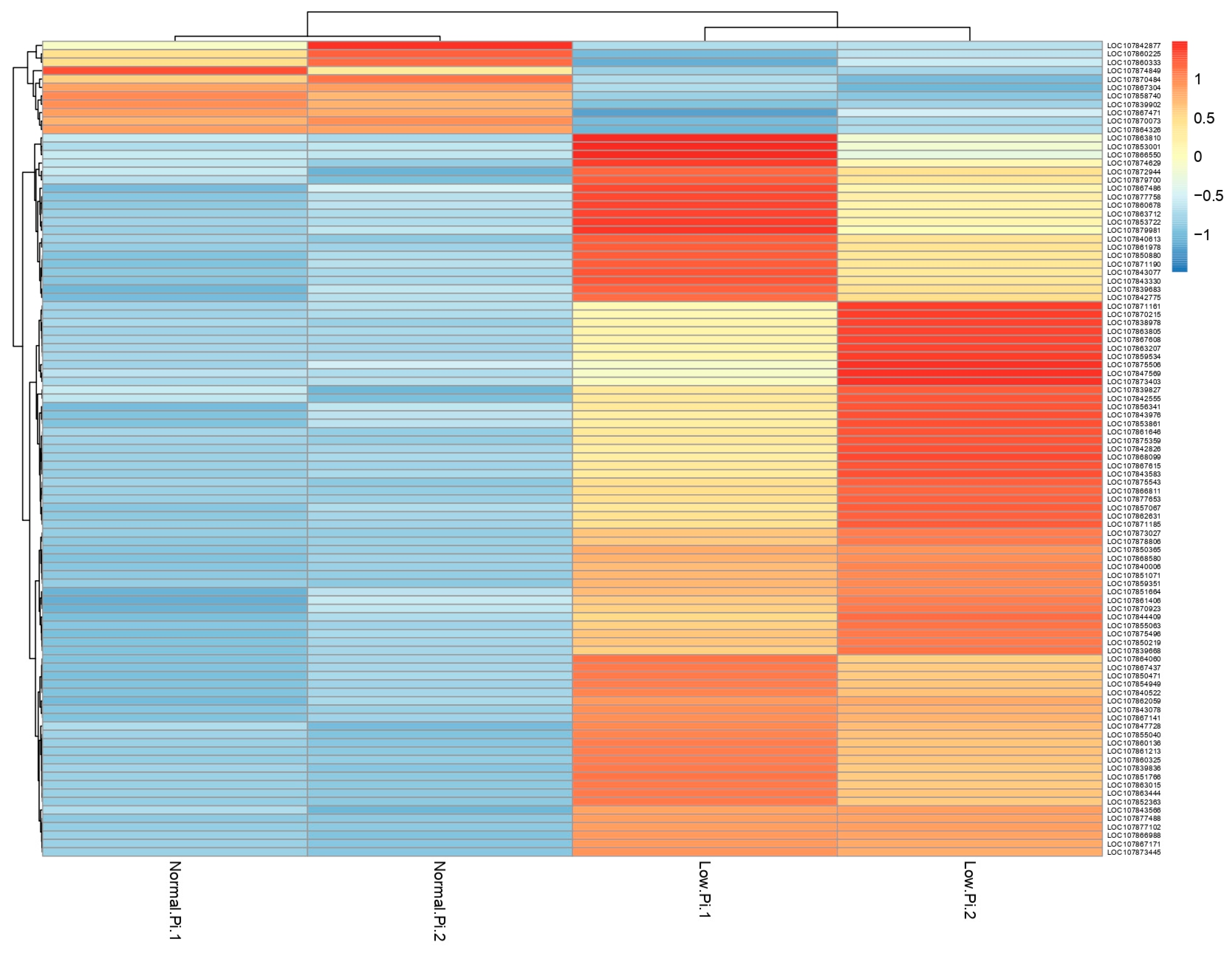
3.1. Analysis of Transcriptome Data
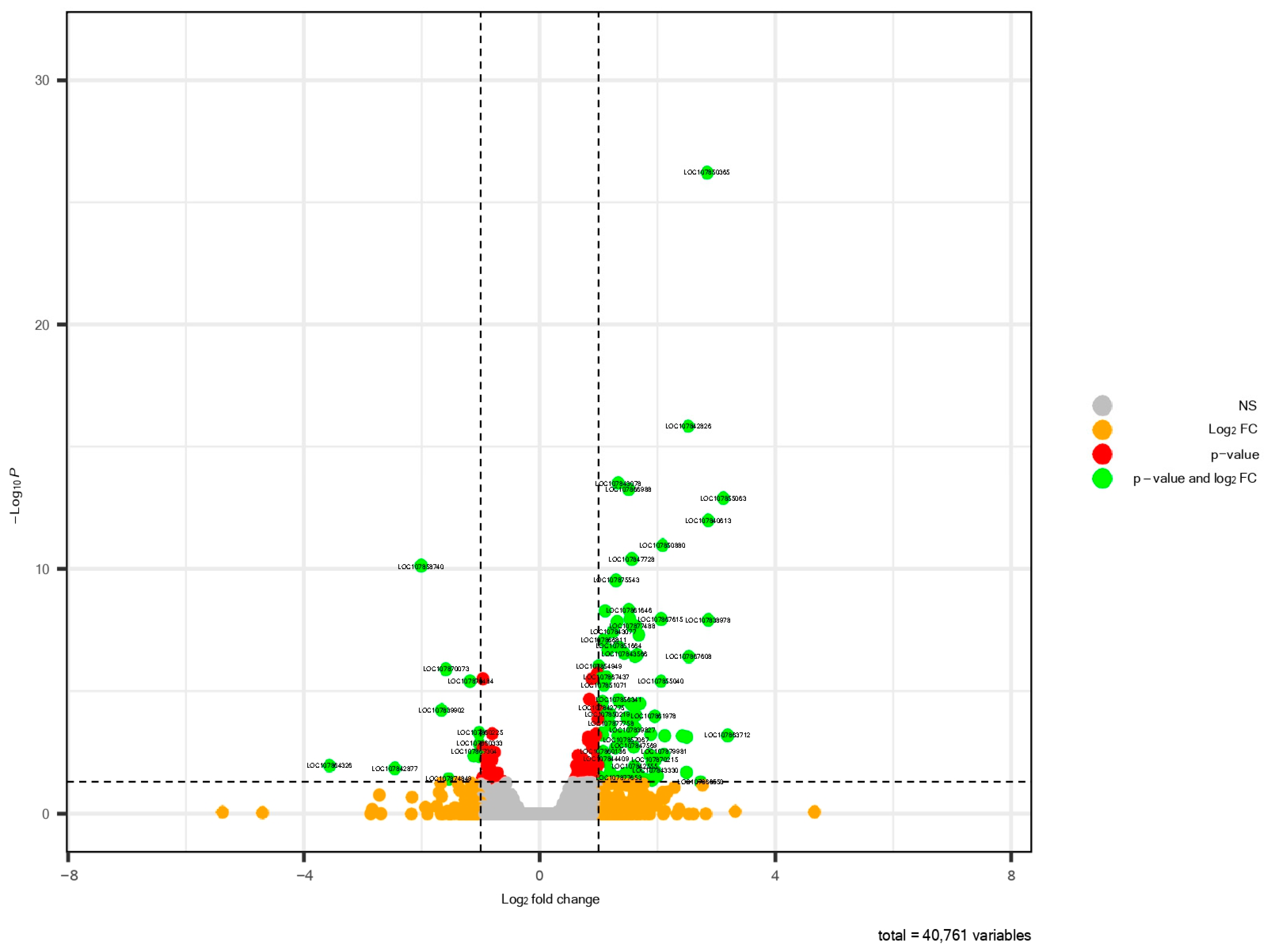
3.2. Differential Expression Analysis of Pepper Roots
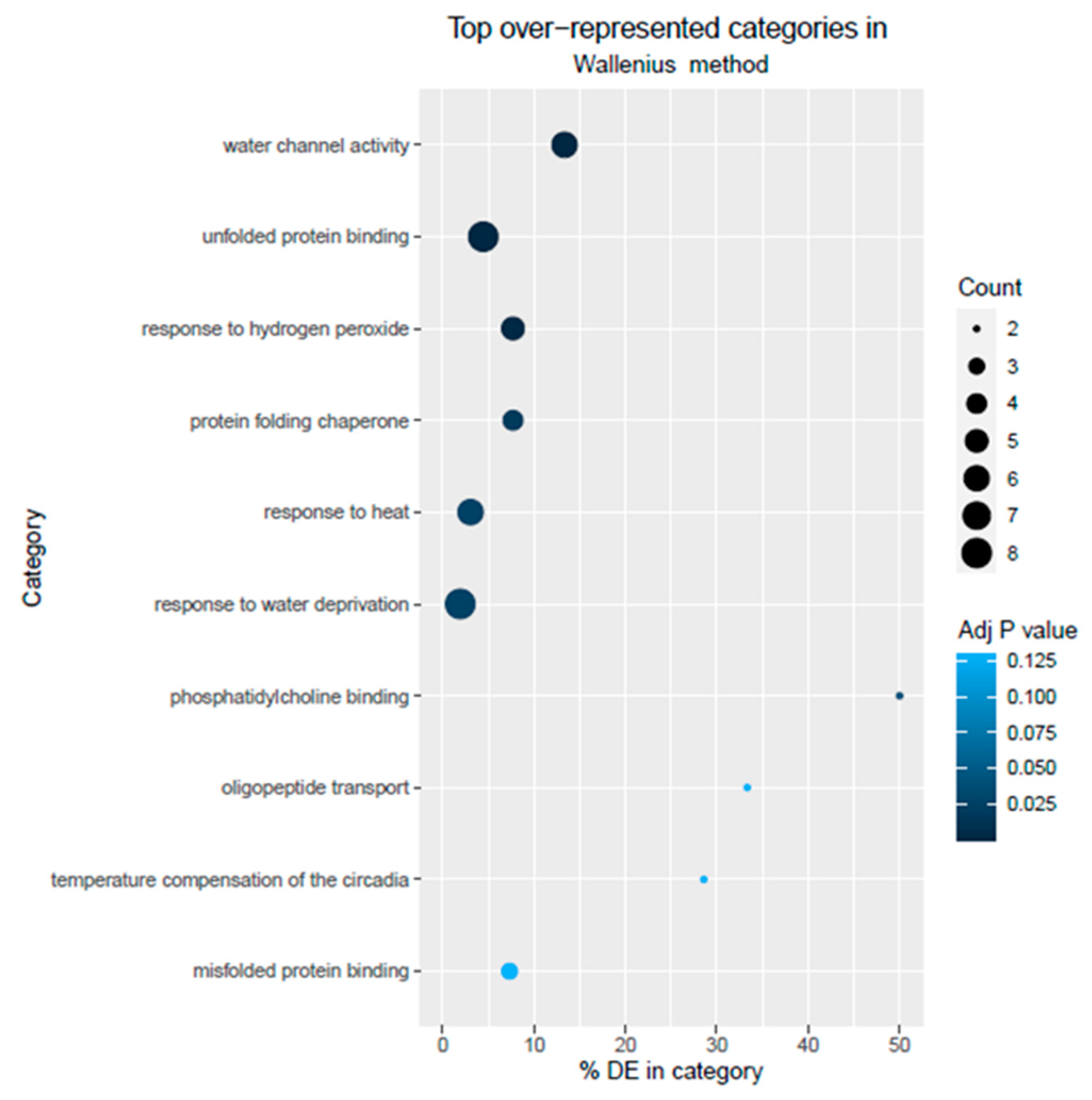
3.3. GO Term Analysis of DEGs
3.4. KEGG Pathway Analysis
3.5. DEGs Associated with Metabolism
3.6. DEGs Related to Transcription Regulation
3.7. DEGs Related to Transportation
3.8. DEGs Related to Stress Response
4. Discussion
Biological Pathways Involved with Stress Response in Pepper Roots under Low Pi
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marschner, H. Beneficial Mineral Elements. In Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 1995; pp. 405–435. [Google Scholar]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [PubMed]
- Raghothama, K.G.; Karthikeyan, A.S. Phosphate acquisition. In Plant Ecophysiology; Springer: Dordrecht, The Netherlands, 2005; pp. 37–49. [Google Scholar] [CrossRef]
- Lin, H.-J.; Gao, J.; Zhang, Z.-M.; Shen, Y.-O.; Lan, H.; Liu, L.; Xiang, K.; Zhao, M.; Zhou, S.; Zhang, Y.-Z.; et al. Transcriptional responses of maize seedling root to phosphorus starvation. Mol. Biol. Rep. 2013, 40, 5359–5379. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, L.; Müller, R.; Nielsen, T.H. Dissecting the plant transcriptome and the regulatory responses to phosphate deprivation. Physiol. Plant. 2010, 139, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Rawoof, A.; Islam, K.; Momo, J.; Ramchiary, N. Identification and expression analysis of phosphate transporter genes and metabolites in response to phosphate stress in Capsicum annuum. Environ. Exp. Bot. 2021, 190, 104597. [Google Scholar] [CrossRef]
- Hernández, G.; Ramírez, M.; Valdés-López, O.; Tesfaye, M.; Graham, M.A.; Czechowski, T.; Schlereth, A.; Wandrey, M.; Erban, A.; Cheung, F.; et al. Phosphorus Stress in Common Bean: Root Transcript and Metabolic Responses. Plant Physiol. 2007, 144, 752–767. [Google Scholar] [CrossRef]
- Chiou, T.-J.; Aung, K.; Lin, S.-I.; Wu, C.-C.; Chiang, S.-F.; Su, C.-l. Regulation of Phosphate Homeostasis by MicroRNA in Arabidopsis. Plant Cell 2005, 18, 412–421. [Google Scholar] [CrossRef]
- Ajmera, I.; Hodgman, T.C.; Lu, C. An integrative systems perspective on plant phosphate research. Genes 2019, 10, 139. [Google Scholar] [CrossRef]
- Ajmera, I.; Shi, J.; Giri, J.; Wu, P.; Stekel, D.J.; Lu, C.; Hodgman, T.C. Regulatory feedback response mechanisms to phosphate starvation in rice. NPJ Syst. Biol. Appl. 2018, 4, 4. [Google Scholar] [CrossRef]
- López-Bucio, J.; Hernández-Abreu, E.; Sánchez-Calderón, L.; Nieto-Jacobo, M.F.; Simpson, J.; Herrera-Estrella, L. Phosphate Availability Alters Architecture and Causes Changes in Hormone Sensitivity in the Arabidopsis Root System. Plant Physiol. 2002, 129, 244–256. [Google Scholar] [CrossRef]
- Steiner, A.A. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil 1961, 15, 134–154. [Google Scholar] [CrossRef]
- Su, S.-Z.; Wu, L.; Liu, D.; Lu, Y.-L.; Lin, H.-J.; Zhang, S.-Z.; Shen, Y.-O.; Liu, H.-L.; Zhang, Z.-M.; Rong, T.-Z.; et al. Genome-Wide Expression Profile of Maize Root Response to Phosphorus Deficiency Revealed by Deep Sequencing. J. Integr. Agric. 2014, 13, 1216–1229. [Google Scholar] [CrossRef]
- Andrews, S. FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 18 April 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Park, M.; Yeom, S.-I.; Kim, Y.-M.; Lee, J.M.; Lee, H.-A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.-T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.S.; Kim, S. Analysis of Whole Transcriptome Sequencing Data: Workflow and Software. Genom. Inform. 2015, 13, 119. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Kong, X.; Zhang, M.; De Smet, I.; Ding, Z. Designer crops: Optimal root system architecture for nutrient acquisition. Trends Biotechnol. 2014, 32, 597–598. [Google Scholar] [CrossRef]
- Wan, H.; Yuan, W.; Ruan, M.; Ye, Q.; Wang, R.; Li, Z.; Zhou, G.; Yao, Z.; Zhao, J.; Liu, S.; et al. Identification of reference genes for reverse transcription quantitative real-time PCR normalization in pepper (Capsicum annuum L.). Biochem. Biophys. Res. Commun. 2011, 416, 24–30. [Google Scholar] [CrossRef]
- Kochian, L.V.; Piñeros, M.A.; Liu, J.; Magalhaes, J.V. Plant Adaptation to Acid Soils: The Molecular Basis for Crop Aluminum Resistance. Annu. Rev. Plant Biol. 2015, 66, 571–598. [Google Scholar] [CrossRef]
- Li, L.; Liu, C.; Lian, X. Gene expression profiles in rice roots under low phosphorus stress. Plant Mol. Biol. 2009, 72, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Péret, B.; Clément, M.; Nussaume, L.; Desnos, T. Root developmental adaptation to phosphate starvation: Better safe than sorry. Trends Plant Sci. 2011, 16, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Datta, R.; Kumar, D.; Sultana, A.; Hazra, S.; Bhattacharyya, D.; Chattopadhyay, S. Glutathione regulates ACC synthase transcription via WRKY33 and ACC oxidase by modulating mRNA stability to induce ethylene synthesis during stress. Plant Physiol. 2015, 169, 2963–2981. [Google Scholar] [CrossRef] [PubMed]
- Houben, M.; Van de Poel, B. 1-Aminocyclopropane-1-carboxylic acid oxidase (ACO): The enzyme that makes the plant hormone ethylene. Front. Plant Sci. 2019, 10, 695. [Google Scholar] [CrossRef]
- Borch, K.; Bouma, T.J.; Lynch, J.P.; Brown, K.M. Ethylene: A regulator of root architectural responses to soil phosphorus availability. Plant Cell Environ. 1999, 22, 425–431. [Google Scholar] [CrossRef]
- Růžička, K.; Ljung, K.; Vanneste, S.; Podhorská, R.; Beeckman, T.; Friml, J.; Benková, E. Ethylene Regulates Root Growth through Effects on Auxin Biosynthesis and Transport-Dependent Auxin Distribution. Plant Cell 2007, 19, 2197–2212. [Google Scholar] [CrossRef]
- Chen, L.; Song, Y.; Li, S.; Zhang, L.; Zou, C.; Yu, D. The role of WRKY transcription factors in plant abiotic stresses. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2012, 1819, 120–128. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A. The role of aquaporins during plant abiotic stress responses. In Plant Life Under Changing Environment; Elsevier: Amsterdam, The Netherlands, 2020; pp. 643–661. [Google Scholar] [CrossRef]
- Yao, Z.F.; Liang, C.Y.; Zhang, Q.; Chen, Z.J.; Xiao, B.X.; Tian, J.; Liao, H. SPX1 is an important component in the phosphorus signalling network of common bean regulating root growth and phosphorus homeostasis. J. Exp. Bot. 2014, 65, 3299–3310. [Google Scholar] [CrossRef]
- Liu, N.; Shang, W.; Li, C.; Jia, L.; Wang, X.; Xing, G.; Zheng, W. Evolution of the SPX gene family in plants and its role in the response mechanism to phosphorus stress. Open Biol. 2018, 8, 170231. [Google Scholar] [CrossRef]
- Zhou, J.; Hu, Q.; Xiao, X.; Yao, D.; Ge, S.; Ye, J.; Li, H.; Cai, R.; Liu, R.; Meng, F.; et al. Mechanism of phosphate sensing and signaling revealed by rice SPX1-PHR2 complex structure. Nat. Commun. 2021, 12, 7040. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Xu, C.; Fan, W.; Zhang, H.; Hou, J.; Yang, A.; Zhang, K. Phosphoproteome and proteome analyses reveal low-phosphate mediated plasticity of root developmental and metabolic regulation in maize (Zea mays L.). Plant Physiol. Biochem. 2014, 83, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Aidoo, M.K.; Sherman, T.; Lazarovitch, N.; Fait, A.; Rachmilevitch, S. A bell pepper cultivar tolerant to chilling enhanced nitrogen allocation and stress-related metabolite accumulation in the roots in response to low root-zone temperature. Physiol. Plant. 2017, 161, 196–210. [Google Scholar] [CrossRef] [PubMed]
- Mandadi, K.K.; Misra, A.; Ren, S.; McKnight, T.D. BT2, a BTB protein, mediates multiple responses to nutrients, stresses, and hormones in Arabidopsis. Plant Physiol. 2009, 150, 1930–1939. [Google Scholar] [CrossRef]
- Rohman, M.M.; Islam, M.R.; Monsur, M.B.; Amiruzzaman, M.; Fujita, M.; Hasanuzzaman, M. Trehalose protects maize plants from salt stress and phosphorus deficiency. Plants 2019, 8, 568. [Google Scholar] [CrossRef]
- Aluko, O.O.; Li, C.; Wang, Q.; Liu, H. Sucrose utilization for improved crop yields: A review article. Int. J. Mol. Sci. 2021, 22, 4704. [Google Scholar] [CrossRef]
- Jain, A.; Poling, M.D.; Karthikeyan, A.S.; Blakeslee, J.J.; Peer, W.A.; Titapiwatanakun, B.; Murphy, A.S.; Raghothama, K.G. Differential effects of sucrose and auxin on localized phosphate deficiency-induced modulation of different traits of root system architecture in Arabidopsis. Plant Physiol. 2007, 144, 232–247. [Google Scholar] [CrossRef]
- Mo, X.; Zhang, M.; Liang, C.; Cai, L.; Tian, J. Integration of metabolome and transcriptome analyses highlights soybean roots responding to phosphorus deficiency by modulating phosphorylated metabolite processes. Plant Physiol. Biochem. 2019, 139, 697–706. [Google Scholar] [CrossRef]
- Lemoine, R. Sucrose transporters in plants: Update on function and structure. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1465, 246–262. [Google Scholar] [CrossRef]
- Williams, D.L.H. Synthesis, properties and reactions of S-nitrosothiols. In Nitrosation Reactions and the Chemistry of Nitric Oxide; Elsevier: Amsterdam, The Netherlands, 2004; pp. 137–160. [Google Scholar] [CrossRef]
- Gómez-García, M.d.R.; Ochoa-Alejo, N. Predominant role of the L-galactose pathway in l-ascorbic acid biosynthesis in fruits and leaves of the Capsicum annuum L. chili pepper. Braz. J. Bot. 2015, 39, 157–168. [Google Scholar] [CrossRef]
- Fitzpatrick, T.B.; Chapman, L.M. The importance of thiamine (vitamin B1) in plant health: From crop yield to biofortification. J. Biol. Chem. 2020, 295, 12002–12013. [Google Scholar] [CrossRef] [PubMed]
- Yusof, Z.N.B. Thiamine and its role in protection against stress in plants (enhancement in thiamine content for nutritional quality improvement). In Nutritional Quality Improvement in Plants; Springer: Cham, Switzerland, 2019; pp. 177–186. [Google Scholar]
- Subki, A.; Abidin, A.A.Z.; Yusof, Z.N.B. The role of thiamine in plants and current perspectives in crop improvement. B Group Vitam.-Curr. Uses Perspect. 2018, 5, 33–44. [Google Scholar]
- Salazar-Gutiérrez, D.; Villicaña-Torres, M.C.; Mendívil, A.C.; Heredia, J.B.; Lightbourn-Rojas, L.A.; León-Félix, J. Transcriptomic Analysis of Pepper Roots (Capsicum annuum) under Phosphorus Deficiency. Research Square 2022, Re-searchSquare:rs.3.rs-1859606, Version 1. Available online: https://www.researchsquare.com/article/rs-1859606/v1 (accessed on 13 July 2023).
- Bustillo-Avendaño, E.; Serrano-Ron, L.; Moreno-Risueno, M.A. The root clock as a signal integrator system: Ensuring balance for survival. Front. Plant Sci. 2022, 13, 886700. [Google Scholar] [CrossRef] [PubMed]
- Voß, U.; Wilson, M.H.; Kenobi, K.; Gould, P.D.; Robertson, F.C.; Peer, W.A.; Lucas, M.; Swarup, K.; Casimiro, I.; Holman, T.J.; et al. The circadian clock rephases during lateral root organ initiation in Arabidopsis thaliana. Nat. Commun. 2015, 6, 7641. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ying, Y.; Secco, D.; Wang, C.; Narsai, R.; Whelan, J.; Shou, H. Molecular interaction between PHO2 and GIGANTEA reveals a newcrosstalk between flowering time and phosphate homeostasis in Oryza sativa. Plant Cell Environ. 2017, 40, 1487–1499. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Su, X.; Wang, Y.; Yang, W.; Pan, Y.; Su, C.; Zhang, X. Genome-wide identification and expression analysis of the BTB domain-containing protein gene family in tomato. Genes Genom. 2018, 40, 1–15. [Google Scholar] [CrossRef]
- Kelley, D.R. E3 ubiquitin ligases: Key regulators of hormone signaling in plants. Mol. Cell. Proteom. 2018, 17, 1047–1054. [Google Scholar] [CrossRef]
- Pan, W.; Wu, Y.; Xie, Q. Regulation of ubiquitination is central to the phosphate starvation response. Trends Plant Sci. 2019, 24, 755–769. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Gene_ID | Gene_Symbol | Log2_FC | P-adj |
|---|---|---|---|
| LOC107864326 | glycine-rich RNA-binding protein 2, mitochondrial-like | −3.57005391 | 0.0107 |
| LOC107842877 | ribulose bisphosphate carboxylase/oxygenase activase 1, chloroplastic | −2.46310915 | 0.0138 |
| LOC107858740 | cyclic dof factor 3-like | −2.01495454 | 0.0000 |
| LOC107839902 | protein EXORDIUM-like 5 | −1.66993147 | 0.0001 |
| LOC107870073 | magnesium-chelatase subunit ChlH, chloroplastic | −1.59432771 | 0.0000 |
| LOC107874849 | ethylene-responsive transcription factor ERF003 | −1.54356028 | 0.0374 |
| LOC107870484 | protein RSI-1 | −1.18048035 | 0.0000 |
| LOC107867304 | uncharacterized LOC107867304 | −1.11502756 | 0.0042 |
| LOC107860225 | lysine-rich arabinogalactan protein 18 | −1.02547235 | 0.0005 |
| LOC107860333 | l-ascorbate oxidase homolog | −1.02069936 | 0.0014 |
| LOC107867471 | protein PHOSPHATE-INDUCED 1 | −1.01901492 | 0.0045 |
| LOC107863712 | uncharacterized LOC107863712 | 1.00184462 | 0.0000 |
| LOC107855063 | protein GIGANTEA-like | 1.0094648 | 0.0006 |
| LOC107838978 | heat shock factor protein HSF30 | 1.02050388 | 0.0424 |
| LOC107840613 | phosphoprotein ECPP44 | 1.02935247 | 0.0323 |
| LOC107850365 | protein GIGANTEA-like | 1.0360357 | 0.0000 |
| LOC107866550 | anthocyanidin 3-O-glucosyltransferase 2 | 1.04703371 | 0.0000 |
| LOC107867608 | small heat shock protein, chloroplastic | 1.04765427 | 0.0066 |
| LOC107842826 | ultraviolet-B receptor UVR8-like | 1.05281042 | 0.0005 |
| LOC107853001 | uncharacterized acetyltransferase At3g50280-like | 1.05288687 | 0.0416 |
| LOC107873403 | heat shock protein 83 | 1.06575196 | 0.0034 |
| LOC107839668 | dnaJ homolog subfamily B member 6-B | 1.06607451 | 0.0499 |
| LOC107863207 | heat shock protein 83-like | 1.06753615 | 0.0000 |
| LOC107879981 | uncharacterized LOC107879981 | 1.06935719 | 0.0006 |
| LOC107850880 | 1-aminocyclopropane-1-carboxylate oxidase 1 | 1.07409809 | 0.0028 |
| LOC107867615 | small heat shock protein, chloroplastic-like | 1.08351024 | 0.0000 |
| LOC107855040 | ethylene-responsive transcription factor ERF054 | 1.0896119 | 0.0000 |
| LOC107877102 | putative elongation of fatty acids protein | 1.10035372 | 0.0350 |
| LOC107870215 | heat shock cognate 70 kDa protein 2-like | 1.10550088 | 0.0001 |
| LOC107843330 | phosphatidylinositol 4-phosphate 5-kinase 9 | 1.11209538 | 0.0000 |
| LOC107861978 | probable WRKY transcription factor 40 | 1.12111699 | 0.0055 |
| LOC107875506 | 17.8 kDa class I heat shock protein | 1.12586035 | 0.0075 |
| LOC107859351 | dnaJ protein homolog 1 | 1.13098799 | 0.0000 |
| LOC107853722 | UTP—glucose-1-phosphate uridylyltransferase | 1.14931399 | 0.0001 |
| LOC107853861 | multiprotein-bridging factor 1c | 1.15469284 | 0.0000 |
| LOC107870923 | probable pectinesterase/pectinesterase inhibitor 12 | 1.19701313 | 0.0202 |
| LOC107839683 | universal stress protein A-like protein | 1.1972449 | 0.0001 |
| LOC107862631 | BTB/POZ and TAZ domain-containing protein 1 | 1.20135206 | 0.0002 |
| LOC107861406 | uncharacterized LOC107861406 | 1.24467066 | 0.0000 |
| LOC107863805 | heat stress transcription factor A-7a-like | 1.28616959 | 0.0020 |
| LOC107861213 | uncharacterized LOC107861213 | 1.29308663 | 0.0000 |
| LOC107872944 | probable aquaporin TIP-type RB7-5A | 1.31615269 | 0.0000 |
| LOC107868099 | E3 ubiquitin-protein ligase MPSR1 | 1.32749478 | 0.0006 |
| LOC107864060 | ethylene-responsive transcription factor ERF071-like | 1.33103614 | 0.0000 |
| LOC107842555 | probable glutathione peroxidase 5 | 1.33549714 | 0.0322 |
| LOC107847569 | dnaJ protein homolog | 1.33669919 | 0.0000 |
| LOC107877488 | zinc finger protein CONSTANS-LIKE 5 | 1.34047692 | 0.0000 |
| LOC107839827 | protein NRT1/PTR FAMILY 4.3 | 1.35779871 | 0.0000 |
| LOC107847728 | serine carboxypeptidase-like | 1.36618804 | 0.0006 |
| LOC107871190 | probable aquaporin PIP1-2 | 1.43489879 | 0.0000 |
| LOC107875496 | serine/arginine-rich splicing factor SR45a-like | 1.45771513 | 0.0011 |
| LOC107859534 | protein HEAT-STRESS-ASSOCIATED 32 | 1.45968575 | 0.0216 |
| LOC107861646 | dnaJ homolog subfamily B member 6 | 1.48123286 | 0.0001 |
| LOC107866988 | uncharacterized LOC107866988 | 1.48592502 | 0.0001 |
| LOC107878806 | nuclear transcription factor Y subunit B-3-like | 1.50678499 | 0.0000 |
| LOC107840522 | uncharacterized protein LOC107840522 | 1.51255887 | 0.0000 |
| LOC107863810 | SPX domain-containing protein 1 | 1.53228982 | 0.0249 |
| LOC107857067 | protein DELAY OF GERMINATION 1 | 1.53235936 | 0.0000 |
| LOC107843566 | probably aquaporin PIP2-4 | 1.5539283 | 0.0000 |
| LOC107843583 | Capsicum annuum contig77344 | 1.55946121 | 0.0000 |
| LOC107867171 | ethylene-responsive transcription factor ERF010-like | 1.56569983 | 0.0004 |
| LOC107851664 | ferredoxin, root R-B2-like | 1.56750124 | 0.0000 |
| LOC107856341 | thiamine thiazole synthase 1, chloroplastic | 1.59743992 | 0.0018 |
| LOC107877653 | temperature-induced lipocalin-1 | 1.60746717 | 0.0112 |
| LOC107843078 | aquaporin PIP2-1 | 1.61611701 | 0.0003 |
| LOC107871161 | alkaline/neutral invertase A, mitochondrial-like | 1.61885367 | 0.0000 |
| LOC107873027 | alcohol dehydrogenase 1 | 1.62957467 | 0.0001 |
| LOC107875543 | heat shock cognate 70 kDa protein 1 | 1.63024785 | 0.0000 |
| LOC107871185 | uncharacterized LOC107871185 | 1.65724777 | 0.0000 |
| LOC107843077 | aquaporin PIP2-1-like | 1.67589881 | 0.0245 |
| LOC107877758 | hemoglobin-2 | 1.68358102 | 0.0000 |
| LOC107860678 | titin | 1.70150059 | 0.0000 |
| LOC107873445 | phosphatidylinositol transfer protein 1 | 1.78702 | 0.0387 |
| LOC107875359 | peptidyl-prolyl cis-trans isomerase FKBP62 | 1.86888069 | 0.0038 |
| LOC107850219 | two-component response regulator-like PRR37 | 1.8876322 | 0.0006 |
| LOC107867437 | protein NRT1/PTR FAMILY 6.3 | 1.90637911 | 0.0416 |
| LOC107867141 | flavonoid 3′-monooxygenase CYP75B137-like | 1.90863477 | 0.0077 |
| LOC107844409 | putative MO25-like protein At5g47540 | 1.95369314 | 0.0001 |
| LOC107840006 | actin-100 | 1.96013972 | 0.0179 |
| LOC107839836 | chloride channel protein CLC-c | 1.96864421 | 0.0059 |
| LOC107879700 | cysteine-rich repeat secretory protein 38 | 1.99554005 | 0.0278 |
| LOC107851071 | cell wall/vacuolar inhibitor of fructosidase 1 | 2.05281355 | 0.0000 |
| LOC107866811 | phosphoprotein ECPP44-like | 2.05439524 | 0.0000 |
| LOC107843976 | activator of 90 kDa heat shock protein ATPase homolog | 2.0786611 | 0.0000 |
| LOC107874629 | probable aquaporin TIP1-2 | 2.10954455 | 0.0037 |
| LOC107863444 | alcohol dehydrogenase-like 4 | 2.12386301 | 0.0006 |
| LOC107863015 | uncharacterized protein At5g23160 | 2.42136983 | 0.0007 |
| LOC107860136 | uncharacterized LOC107860136 | 2.49214334 | 0.0200 |
| LOC107851766 | UDP-glucosyltransferase 29 | 2.4953006 | 0.0007 |
| LOC107860325 | peroxidase 10 | 2.51720866 | 0.0000 |
| LOC107852363 | probable trehalose-phosphate phosphatase J | 2.52472223 | 0.0000 |
| LOC107842775 | protein ENHANCED PSEUDOMONAS SUSCEPTIBILITY 1 | 2.73066046 | 0.0491 |
| LOC107868580 | serine/threonine-protein kinase SAPK3 | 2.83899314 | 0.0000 |
| LOC107867486 | l-lactate dehydrogenase B | 2.85451109 | 0.0000 |
| LOC107850471 | stearoyl-[acyl-carrier-protein] 9-desaturase, chloroplastic | 2.85691431 | 0.0000 |
| LOC107862059 | uncharacterized LOC107862059 | 3.11160163 | 0.0000 |
| LOC107854949 | early endosome antigen 1 | 3.18361572 | 0.0006 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar-Gutiérrez, D.; Cruz-Mendívil, A.; Villicaña, C.; Heredia, J.B.; Lightbourn-Rojas, L.A.; León-Félix, J. Transcriptomic Analysis Reveals the Response Mechanisms of Bell Pepper (Capsicum annuum) to Phosphorus Deficiency. Metabolites 2023, 13, 1078. https://doi.org/10.3390/metabo13101078
Salazar-Gutiérrez D, Cruz-Mendívil A, Villicaña C, Heredia JB, Lightbourn-Rojas LA, León-Félix J. Transcriptomic Analysis Reveals the Response Mechanisms of Bell Pepper (Capsicum annuum) to Phosphorus Deficiency. Metabolites. 2023; 13(10):1078. https://doi.org/10.3390/metabo13101078
Chicago/Turabian StyleSalazar-Gutiérrez, Daizha, Abraham Cruz-Mendívil, Claudia Villicaña, José Basilio Heredia, Luis Alberto Lightbourn-Rojas, and Josefina León-Félix. 2023. "Transcriptomic Analysis Reveals the Response Mechanisms of Bell Pepper (Capsicum annuum) to Phosphorus Deficiency" Metabolites 13, no. 10: 1078. https://doi.org/10.3390/metabo13101078
APA StyleSalazar-Gutiérrez, D., Cruz-Mendívil, A., Villicaña, C., Heredia, J. B., Lightbourn-Rojas, L. A., & León-Félix, J. (2023). Transcriptomic Analysis Reveals the Response Mechanisms of Bell Pepper (Capsicum annuum) to Phosphorus Deficiency. Metabolites, 13(10), 1078. https://doi.org/10.3390/metabo13101078