

Identification of the Biosynthetic Gene Cluster of New Piperazic Acid-Containing Lipopeptides with Cytotoxic Activity in the Genome of Marine Streptomyces PHM034

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media, and Culture Conditions
2.2. Isolation of the Strain
2.3. Taxonomy
2.4. Fermentation and Extraction
2.5. Isolation and Structural Elucidation of 1, 2 and 3
2.6. Sequencing of the Genome
2.7. Bioinformatic Analysis
2.8. Construction of Disruption Plasmid and Protoplast Generation and Transformation
2.9. Bioactivity of Fermentation Broth
2.10. Citotoxic Activity Assays
2.11. Antimicrobial Activity
3. Results
3.1. Strain Identification
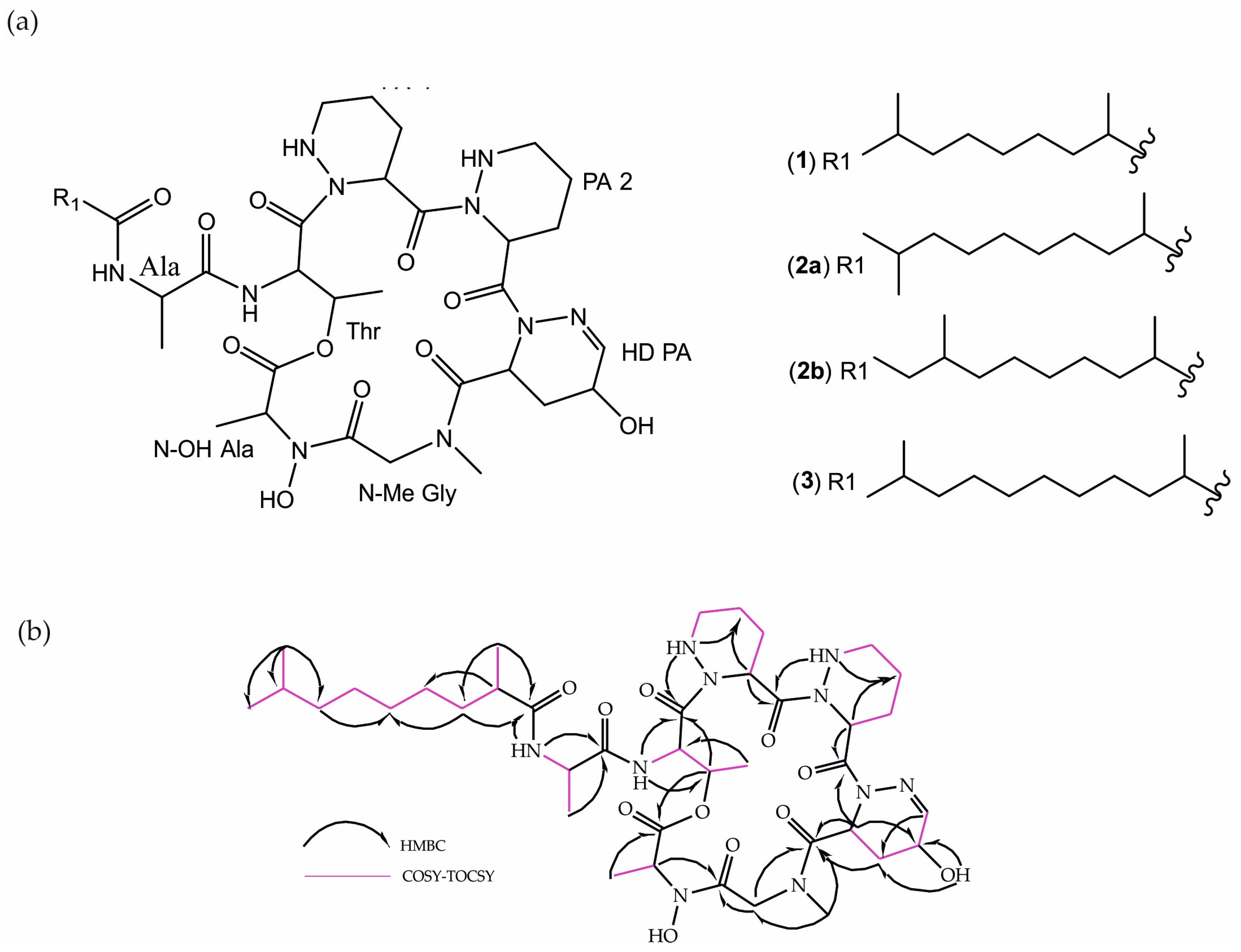
3.2. Bacteria Cultivation and Identification of Lipopeptides 1, 2, and 3
3.3. In Vitro Bioactivity
3.4. Genome Sequencing and Identification of Secondary Metabolism Gene Clusters
3.5. Inactivation of the NRPS
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Cancer Observatory: Cancer Today. Lyon: International Agency for Research on Cancer. Available online: https://www.who.int/health-topics/cancer#tab=tab_1 (accessed on 4 February 2021).
- Crosby, D.; Bhatia, S.; Brindle, K.M.; Coussens, L.M.; Dive, C.; Emberton, M.; Esener, S.; Fitzgerald, R.C.; Gambhir, S.S.; Kuhn, P.; et al. Early detection of cancer. Science 2022, 375, eaay9040. [Google Scholar] [CrossRef] [PubMed]
- Mayo Clinic. Cancer Treatment. Available online: https://www.mayoclinic.org/tests-procedures/cancer-treatment/about/pac-20393344 (accessed on 22 September 2023).
- Goodman, L.S.; Wintrobe, M.M. Nitrogen mustard therapy; use of methyl-bis (beta-chloroethyl) amine hydrochloride and tris (beta-chloroethyl) amine hydrochloride for Hodgkin’s disease, lymphosarcoma, leukemia and certain allied and miscellaneous disorders. J. Am. Med. Assoc. 1946, 132, 126–132. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society. Chemotherapy Side Effects. Available online: https://www.cancer.org/cancer/managing-cancer/treatment-types/chemotherapy/chemotherapy-side-effects.html (accessed on 22 September 2023).
- Demain, A.L.; Fang, A. The natural functions of secondary metabolites. Adv. Biochem. Eng. Biotechnol. 2000, 69, 1–39. [Google Scholar] [PubMed]
- Ordenes-Aenishanslins, N.; Anziani-Ostuni, G.; Vargas-Reyes, M.; Alarcon, J.; Tello, A.; Perez-Donoso, J.M. Pigments from UV-resistant Antarctic bacteria as photosensitizers in dye sensitized solar cells. J. Photochem. Photobiol. B 2016, 162, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Mullis, M.M.; Rambo, I.M.; Baker, B.J.; Reese, B.K. Diversity, ecology, and prevalence of antimicrobials in nature. Front. Microbiol. 2019, 10, 2518. [Google Scholar] [CrossRef] [PubMed]
- Pishchany, G.; Kolter, R. On the possible ecological roles of antimicrobials. Mol. Microbiol. 2020, 113, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Chater, K.F. Streptomyces inside-out: A new perspective on the bacteria that provide us with antibiotics. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 761–768. [Google Scholar] [CrossRef]
- Manteca, Á.; Yagüe, P. Streptomyces as a source of antimicrobials: Novel approaches to activate cryptic secondary metabolite pathways. In Antimicrobials, Antibiotic Resistance, Antibiofilm Strategies and Activity Methods; Kırmusaoğlu, S., Ed.; IntechOpen: Rijeka, Croatia, 2019. [Google Scholar]
- Olano, C.; Mendez, C.; Salas, J.A. Antitumor compounds from marine actinomycetes. Mar. Drugs 2009, 7, 210–248. [Google Scholar] [CrossRef]
- Rao, M.P.N.; Xiao, M.; Li, W.J. Fungal and bacterial pigments: Secondary metabolites with wide applications. Front. Microbiol. 2017, 8, 1113. [Google Scholar]
- Kumar, A.; Vishwakarma, H.S.; Singh, J.; Dwivedi, S.; Kumar, M. Microbial pigments: Production and their applications in various industries. Int. J. Pharm. Chem. Biol. Sci. 2015, 5, 203–212. [Google Scholar]
- Ghareeb, M.A.; Tammam, M.A.; El-Demerdash, A.; Atanaov, A.G. Insights about clinically approved and Preclinically investigated marine natural products. Curr. Res. Biotechnol. 2020, 2, 88–102. [Google Scholar] [CrossRef]
- Huang, M.; Lu, J.; Ding, J. Natural Products in Cancer Therapy: Past, Present and Future. Nat. Prod. Bioprospecting 2021, 11, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Fan, W.; Zhao, Y.; Deng, Z.; Feng, Y. Probing and Engineering the Fatty Acyl Substrate Selectivity of Starter Condensation Domains of Nonribosomal Peptide Synthetases in Lipopeptide Biosynthesis. Biotechnol. J. 2020, 15, 1900175. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Chavez, C.; Benaud, N.; Ferrari, B.C. The ecological roles of microbial lipopeptides: Where are we going? Comput. Struct. Biotechnol. J. 2021, 19, 1400–1413. [Google Scholar] [CrossRef] [PubMed]
- Cimermancic, P.; Medema, M.H.; Claesen, J.; Kurita, K.; Brown, L.C.W.; Mavrommatis, K.; Pati, A.; Godfrey, P.A.; Koehrsen, M.; Clardy, J.; et al. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhang, R.; Wang, D.; Dong, W.; Wang, Z.; Zhai, Y.; Han, W.; Yin, X.; Tian, J.; Wei, J.; et al. Genomic and Metabolite Profiling Reveal a Novel Streptomyces Strain, QHH-9511, from the Qinghai-Tibet Plateau. Microbiol. Spectr. 2023, 11, e02764-22. [Google Scholar] [CrossRef] [PubMed]
- Wietz, M.; Mansson, M.; Vynne, N.G.; Gram, L. Small-molecule antibiotics from marine bacteria and strategies to prevent rediscovery of known compounds. In Marine Microbiology: Bioactive Compounds and Biotechnological Applications; Kim, S.-C., Ed.; Wiley-Vch: Weinheim, Germany, 2013. [Google Scholar]
- Yang, Z.; He, J.; Wei, X.; Ju, J.; Ma, J. Exploration and genome mining of natural products from marine Streptomyces. Appl. Microbiol. Biotechnol. 2020, 104, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Li, Y.L.; Zhao, F.C. Secondary metabolites from polar organisms. Mar. Drugs 2017, 15, 28. [Google Scholar] [CrossRef]
- Rangseekaew, P.; Pathom-Aree, W. Cave actinobacteria as producers of bioactive metabolites. Front. Microbiol. 2019, 10, 387. [Google Scholar] [CrossRef]
- de la Calle, F. Marine microbiome as source of natural products. Microb. Biotechnol. 2017, 10, 1293–1296. [Google Scholar] [CrossRef]
- Ceniceros, A.; Cuervo, L.; Mendez, C.; Salas, J.A.; Olano, C.; Malmierca, M.G. A Multidisciplinary Approach to Unraveling the Natural Product Biosynthetic Potential of a Streptomyces Strain Collection Isolated from Leaf-Cutting Ants. Microorganisms 2021, 9, 2225. [Google Scholar] [CrossRef] [PubMed]
- Santos-Beneit, F.; Ceniceros, A.; Nikolaou, A.; Salas, J.A.; Gutierrez-Merino, J. Identification of Antimicrobial Compounds in Two Streptomyces sp. Strains Isolated From Beehives. Front. Microbiol. 2022, 13, 742168. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Lee, G.; Hwang, G.J.; Heo, K.T.; Lee, J.K.; Jang, J.; Hwang, B.Y.; Jang, J.; Cho, Y.; Hong, Y. Rubiflavin G, photorubiflavin G, and photorubiflavin E: Novel pluramycin derivatives from Streptomyces sp. W2061 and their anticancer activity against breast cancer cells. J Antibiot. 2023, 76, 585–591. [Google Scholar] [CrossRef]
- Huang, Y.; Hu, W.; Huang, S.; Chu, J.; Liang, Y.; Tao, Z.; Wang, G.; Zhuang, J.; Zhang, Z.; Zhou, X.; et al. Taxonomy and anticancer potential of Streptomyces niphimycinicus sp. nov. against nasopharyngeal carcinoma cells. Appl. Microbiol. Biotechnol. 2023, 107, 6325–6338. [Google Scholar] [CrossRef] [PubMed]
- Kouroshnia, A.; Zeinali, S.; Irani, S.; Sadeghi, A. Induction of apoptosis and cell cycle arrest in colorectal cancer cells by novel anticancer metabolites of Streptomyces sp. 801. Cancer Cell Int. 2022, 22, 235. [Google Scholar] [CrossRef] [PubMed]
- Aftab, U.; Zechel, D.L.; Sajid, I. Antitumor compounds from Streptomyces sp. KML-2, isolated from Khewra salt mines, Pakistan. Biol. Res. 2015, 48, 58. [Google Scholar] [CrossRef] [PubMed]
- Morgan, K.D.; Andersen, R.J.; Ryan, K.S. Piperazic acid-containing natural products: Structures and biosynthesis. Nat. Prod. Rep. 2019, 36, 1628–1653. [Google Scholar] [CrossRef]
- Dembitsky, V.M. Hydrobiological Aspects of Fatty Acids: Unique, Rare, and Unusual Fatty Acids Incorporated into Linear and Cyclic Lipopeptides and Their Biological Activity. Hydrobiology 2022, 1, 331–432. [Google Scholar] [CrossRef]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Bierman, M.; Logan, R.; O’Brien, K.; Seno, E.T.; Rao, R.N.; Schoner, B.E. Plasmid cloning vectors for the conjugal transfer of DNA from Escherichia coli to Streptomyces spp. Gene 1992, 116, 43–49. [Google Scholar] [CrossRef]
- Clarridge, J.E., III. Impact of 16S rRNA gene sequence analysis for identification of bacteria on clinical microbiology and infectious diseases. Clin. Microbiol. Rev. 2004, 17, 840–862. [Google Scholar] [CrossRef]
- Cook, A.E.; Meyers, P.R. Rapid identification of filamentous actinomycetes to the genus level using genus-specific 16S rRNA gene restriction fragment patterns. Int. J. Syst. Evol. Microbiol. 2003, 53, 1907–1915. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef]
- Skinnider, M.A.; Merwin, N.J.; Johnston, C.W.; Magarvey, N.A. PRISM 3: Expanded prediction of natural product chemical structures from microbial genomes. Nucleic Acids Res. 2017, 45, W49–W54. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Soares, J.; Greninger, P.; Edelman, E.J.; Lightfoot, H.; Forbes, S.; Bindal, N.; Beare, D.; Smith, J.A.; Thompson, I.R.; et al. Genomics of Drug Sensitivity in Cancer (GDSC): A resource for therapeutic biomarker discovery in cancer cells. Nucleic Acids Res. 2013, 41, 955. [Google Scholar] [CrossRef]
- Shoemaker, R.H. The NCI60 human tumour cell line anticancer drug screen. Nat. Rev. Cancer 2006, 6, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Goker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.H.; Tang, Y. Adding the lipo to lipopeptides: Do more with less. Chem. Biol. 2010, 17, 791–793. [Google Scholar] [CrossRef]
- Rausch, C.; Hoof, I.; Weber, T.; Wohlleben, W.; Huson, D.H. Phylogenetic analysis of condensation domains in NRPS sheds light on their functional evolution. BMC Evol. Biol. 2007, 7, 78. [Google Scholar] [CrossRef]
- Li, Y.; Tahlan, K.; Bignell, D.R.D. Functional Cross-Talk of MbtH-Like Proteins During Thaxtomin Biosynthesis in the Potato Common Scab Pathogen Streptomyces scabiei. Front. Microbiol. 2020, 11, 585456. [Google Scholar] [CrossRef]
- Zwahlen, R.D.; Pohl, C.; Bovenberg, R.A.L.; Driessen, A.J.M. Bacterial MbtH-like Proteins Stimulate Nonribosomal Peptide Synthetase-Derived Secondary Metabolism in Filamentous Fungi. ACS Synth. Biol. 2019, 8, 1776–1787. [Google Scholar] [CrossRef]
- Singh, G.M.; Vaillancourt, F.H.; Yin, J.; Walsh, C.T. Characterization of SyrC, an aminoacyltransferase shuttling threonyl and chlorothreonyl residues in the syringomycin biosynthetic assembly line. Chem. Biol. 2007, 14, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Escribano, J.P.; Alt, S.; Bibb, M.J. Next Generation Sequencing of Actinobacteria for the Discovery of Novel Natural Products. Mar. Drugs 2016, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Baltz, R.H. Genome mining for drug discovery: Cyclic lipopeptides related to daptomycin. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab020. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 68, 394–424. [Google Scholar] [CrossRef]
- Rajappa, S.J.; Pinninti, R.; Are, C. Neoadjuvant therapy for pancreas cancer: Global perspective and optimal care pathways in low to middle-income countries. J. Surg. Oncol. 2021, 123, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society. Cancer Statistics Center. Available online: http://cancerstatisticscenter.cancer.org (accessed on 4 February 2021).
- Choi, Y.J.; Byun, Y.; Kang, J.S.; Kim, H.S.; Han, Y.; Kim, H.; Kwon, W.; Oh, D.Y.; Paik, W.H.; Lee, S.H.; et al. Comparison of Clinical Outcomes of Borderline Resectable Pancreatic Cancer According to the Neoadjuvant Chemo-Regimens: Gemcitabine versus FOLFIRINOX. Gut Liver 2020, 15, 466. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tao, X.; Zou, A.; Yang, S.; Zhang, L.; Mu, B. Effect of the microbial lipopeptide on tumor cell lines: Apoptosis induced by disturbing the fatty acid composition of cell membrane. Protein Cell 2010, 1, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Navarro-Munoz, J.C.; Terlouw, B.R.; van der Hooft, J.J.J.; van Santen, J.A.; Tracanna, V.; Duran, H.G.S.; Andreu, V.P.; et al. MIBiG 2.0: A repository for biosynthetic gene clusters of known function. Nucleic Acids Res. 2020, 48, D454–D458. [Google Scholar] [CrossRef]



{kind=link}
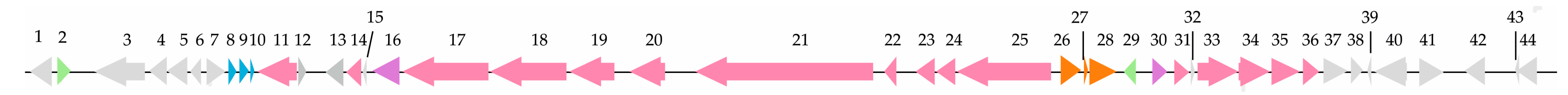{kind=link}
| Strain Name | Details | Resistance | Reference |
|---|---|---|---|
| Streptomyces tuirus PHM034 | Wild type strain | - | This work |
| Streptomyces tuirus PM13 Dis7 and Streptomyces tuirus PM13 Dis8 | Disruption of the biosynthetic cluster of PM130392, PM130393 and PM140293 by insertion of the vector pOJ260 [35] | ApraR | This work |
| Escherichia coli DH5α | F-80dlacZ M15 (lacZYA-argF) U169 recA1 endA1 hsdR17(rk−, mk+) phoAsupE44λ-thi-1 gyrA96 relA1 | - | Commercial strain |
| Primer Name | Sequence 5′➔3′ |
|---|---|
| PM13 Dis Fw | ATATTCTAGACGGGAGAGGTTGAGGAAGTC |
| PM13 Dis Rv | ATATGGATCCGGCTTTCGCATCGAACTC |
| PM13 Dis1 check Up | CGAGCTGCTCAACCTGTACG |
| M13 Fw | GTAAAACGACGGCCAGT |
| PM13 Dis1 check down | GTCGTCGAACGGGTTGTTGC |
| pOJ260 Check | ACACAGGAAACAGCTATGAC |
| Compounds | Breast | Colon | Lung | Pancreas |
|---|---|---|---|---|
| MDA-MB-231 | HT29 | NSCLC A549 | PSN1 | |
| PM130391 (1) | 4.60 × 10−8 | 4.60 × 10−8 | 1.00 × 10−7 | 1.90 × 10−8 |
| PM130392 (2) | 1.60 × 10−8 | 2.00 × 10−8 | 4.80 × 10−8 | 7.00 × 10−9 |
| PM140293 (3) | 1.60 × 10−8 | 2.10 × 10−8 | 3.20 × 10−8 | 3.00 × 10−9 |
| Gefitinib | 3.48 × 10−5 | 1.74 × 10−4 | 1.33 × 10−5 | 5.18 × 10−5 |
| Erlotinib | 1.99 × 10−5 | 6.48 × 10−5 | 8.12 × 10−6 | 2.35 × 10−5 |
| 5-FU | 6.67 × 10−5 | 2.48 × 10−5 | 9.96 × 10−6 | 6.41 × 10−6 |
| Epirubicin | 1.50 × 10−7 | 5.90 × 10−7 | 2.10 × 10−7 | 1.10 × 10−7 |
| Cyclophosphamide | 1.96 × 10−4 | 6.64 × 10−4 | 3.29 × 10−4 | 1.32 × 10−4 |
| Irinotecan | 1.55 × 10−5 | 4.52 × 10−5 | 1.30 × 10−5 | 7.71 × 10−6 |
| Oxaliplatin | 3.73 × 10−5 | 1.16 × 10−5 | 4.98 × 10−6 | 2.23 × 10−5 |
| Unit Position | δC | δH (J in Hz) | Unit Position | δC | δH (J in Hz) |
|---|---|---|---|---|---|
| Ala | DHPA | ||||
| 1-C | 171.9 | 1-C | 171.1 | ||
| 2-CH | 48.2 | 4.10 (d, 6.7) | 2-CH | 50.7 | 5.28 (d, 6.6, 1.7) |
| 3-CH3 | 19.9 | 1.18 (d, 6.8) | 3-CH2 | 25.6 | 2.48 (ddt, 13.0, 6.2, 1.8) |
| NH | 6.68 (d, 6.4) | 1.98 (m) | |||
| Thr | 4-CH | 59.6 | 4.57 (dd, 12.0, 6.2) | ||
| 1-C | 171.3 | 5-CH | 149.8 | 7.01 (s) | |
| 2-CH | 49.5 | 5.95 (dd, 7.1, 3.2) | |||
| 3-CH | 69.4 | 5.19 (dd, 6.5, 3.4) | N-Me-Gly | ||
| 4-CH3 | 15.7 | 1.30 (d, 6.5) | 1-C | 168.9 | |
| NH | 6.99 (d, 7.1) | 2-CH2 | 51.3 | 4.97 (d, 17.5) | |
| PA-1 | 3.53 (d, 17.5) | ||||
| 1-C | 172.9 | N-CH3 | 36.8 | 3.19 (s) | |
| 2-CH | 49.2 | 5.64 (dd, 6.5, 1.8) | N-OH-Ala | ||
| 3-CH2 | 25.3 | 1.90 (m) | 1-C | 169.7 | |
| 2.07 (m) | 2-CH | 53.3 | 5.20 (t, 7.4) | ||
| 4-CH2 | 21.5 | 1.56 (m) | 3-CH3 | 14.3 | 1.51 (d, 7.4) |
| 5-CH2 | 47.3 | 3.05 (d, 13.5) | 2,8 DMNA | ||
| 2.67 (m) | 1-C | 175.7 | |||
| NH | 5.4 (dd, 13.2, 1.9) | 2-CH | 41.3 | 2.17 (m) | |
| PA-2 | 3-CH2 | 34.4 | 1.59 (m) | ||
| 1-C | 172.7 | 1.33 (m) | |||
| 2-CH | 43.8 | 6.09 (dd 6.6, 2.0) | 4-CH2 | 27.4 | 1.24 (m) |
| 3-CH2 | 23.5 | 1.97 (m) | 5-CH2 | 29.8 | 1.24 (m) |
| 4-CH2 | 19.6 | 2.14 (m) | 6-CH2 | 27.2 | 1.24 (m) |
| 1.54 (m) | 7-CH2 | 38.9 | 1.23 (m) | ||
| 5-CH2 | 47.4 | 3.16 (m) | 1.13 (m) | ||
| 2.75 (dt, 13.3, 3.4) | 8-CH | 27.9 | 1.50 (m) | ||
| NH | 4.80 (dd, 12.9, 1.8) | 9-CH3 | 22.6 | 0.86 (d, 6.6) | |
| 10-CH3 | 22.6 | 0.86 (d, 6.6) | |||
| 11-CH3 | 17.5 | 1.10 (d, 6.8) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceniceros, A.; Cañedo, L.; Méndez, C.; Olano, C.; Schleissner, C.; Cuevas, C.; Calle, F.d.l.; Salas, J.A. Identification of the Biosynthetic Gene Cluster of New Piperazic Acid-Containing Lipopeptides with Cytotoxic Activity in the Genome of Marine Streptomyces PHM034. Metabolites 2023, 13, 1091. https://doi.org/10.3390/metabo13101091
Ceniceros A, Cañedo L, Méndez C, Olano C, Schleissner C, Cuevas C, Calle Fdl, Salas JA. Identification of the Biosynthetic Gene Cluster of New Piperazic Acid-Containing Lipopeptides with Cytotoxic Activity in the Genome of Marine Streptomyces PHM034. Metabolites. 2023; 13(10):1091. https://doi.org/10.3390/metabo13101091
Chicago/Turabian StyleCeniceros, Ana, Librada Cañedo, Carmen Méndez, Carlos Olano, Carmen Schleissner, Carmen Cuevas, Fernando de la Calle, and José A. Salas. 2023. "Identification of the Biosynthetic Gene Cluster of New Piperazic Acid-Containing Lipopeptides with Cytotoxic Activity in the Genome of Marine Streptomyces PHM034" Metabolites 13, no. 10: 1091. https://doi.org/10.3390/metabo13101091
APA StyleCeniceros, A., Cañedo, L., Méndez, C., Olano, C., Schleissner, C., Cuevas, C., Calle, F. d. l., & Salas, J. A. (2023). Identification of the Biosynthetic Gene Cluster of New Piperazic Acid-Containing Lipopeptides with Cytotoxic Activity in the Genome of Marine Streptomyces PHM034. Metabolites, 13(10), 1091. https://doi.org/10.3390/metabo13101091