
Mass Spectrometry Chromatography-Based Metabolomics: The Effect of Long-Term Aerobic Exercise on Learning Ability and the Metabolism of Intestinal Contents in Mice with Alzheimer’s Disease


Abstract
:
1. Introduction
2. Materials and Methods
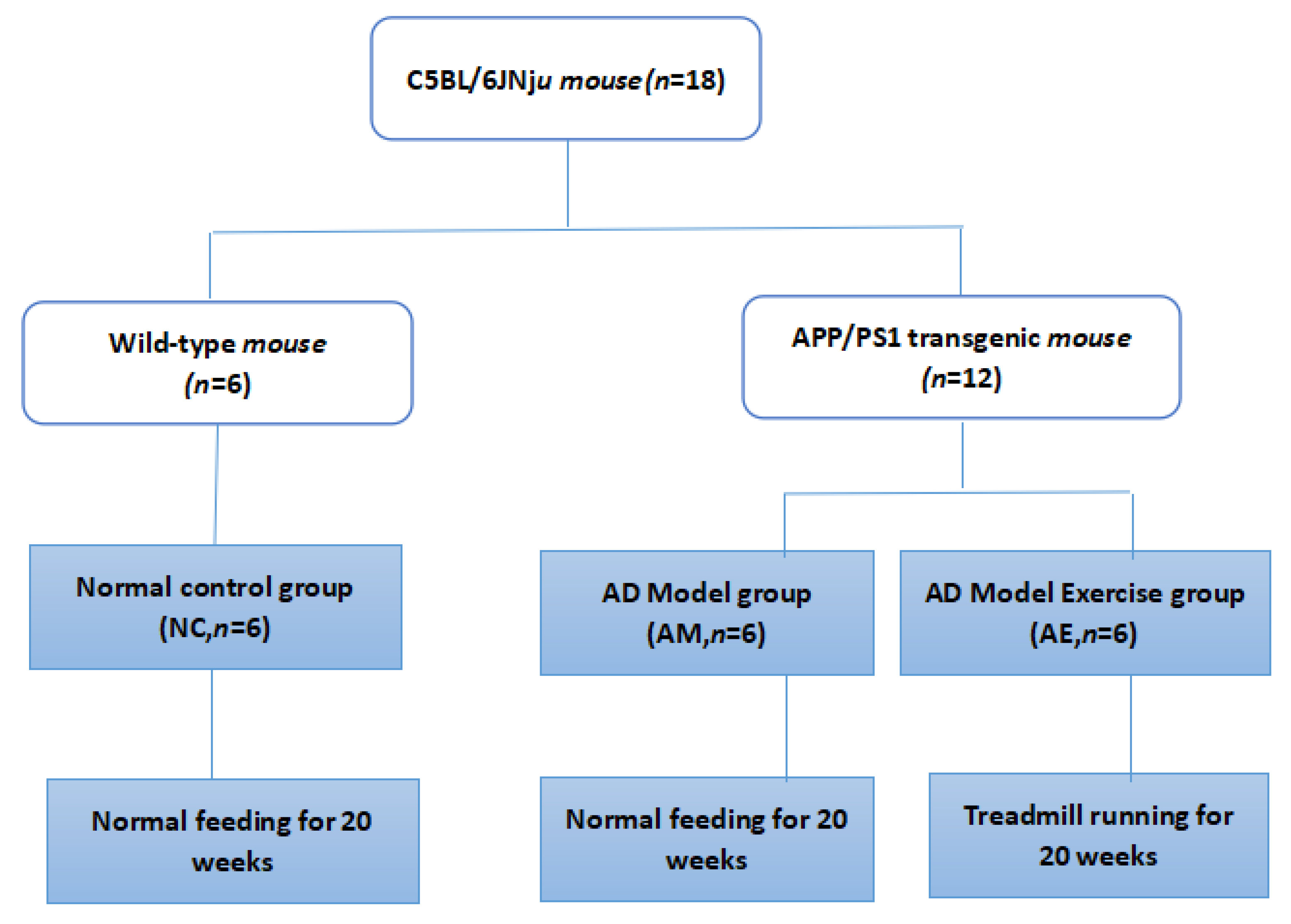
2.1. Experimental Animals
2.2. Movement Scheme
2.3. Morris Water Maze Behavioral Test
2.4. Sample Collection and Processing
2.4.1. Sample Collection
2.4.2. Pretreatment of HPLC-MS Samples
2.4.3. Chromatographic Conditions
2.4.4. Mass Spectrometry Conditions
2.5. Data Analysis
3. Results
3.1. General Health Status of Animals
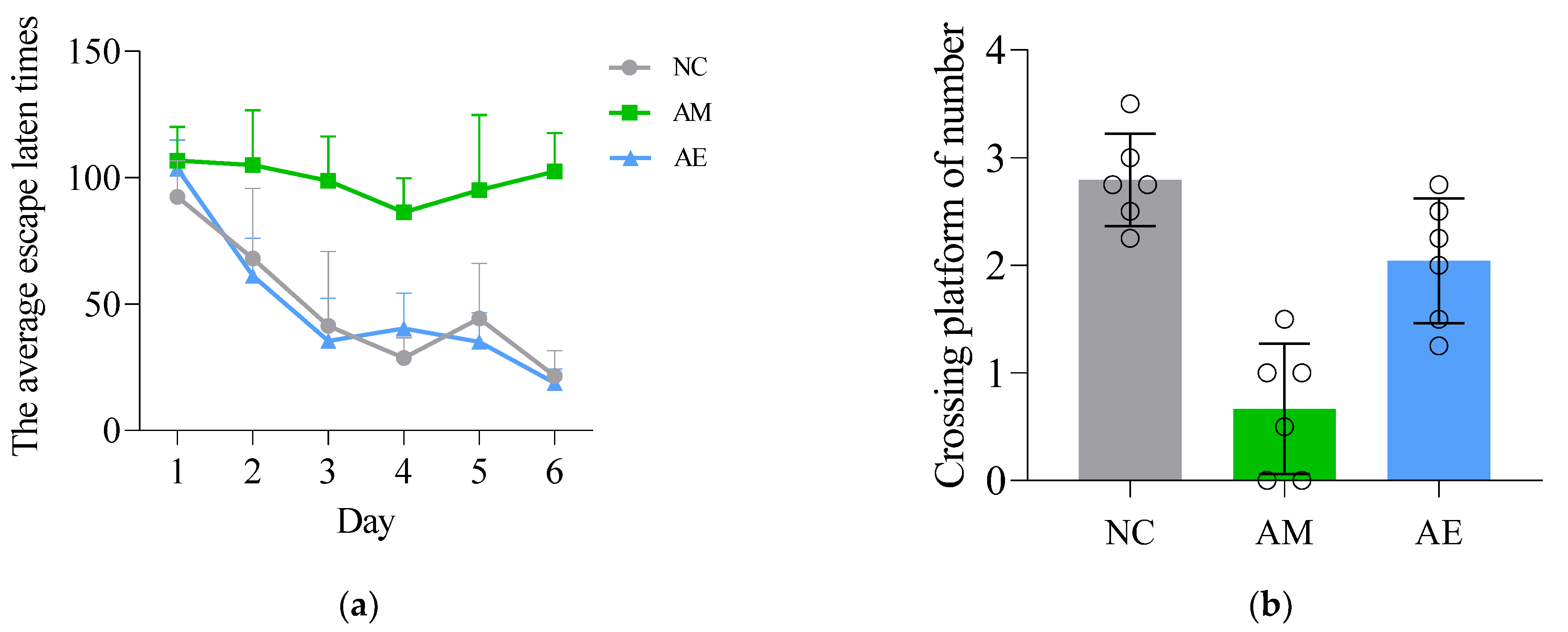
3.2. Behavioral Test—Morris Water Maze Experiment
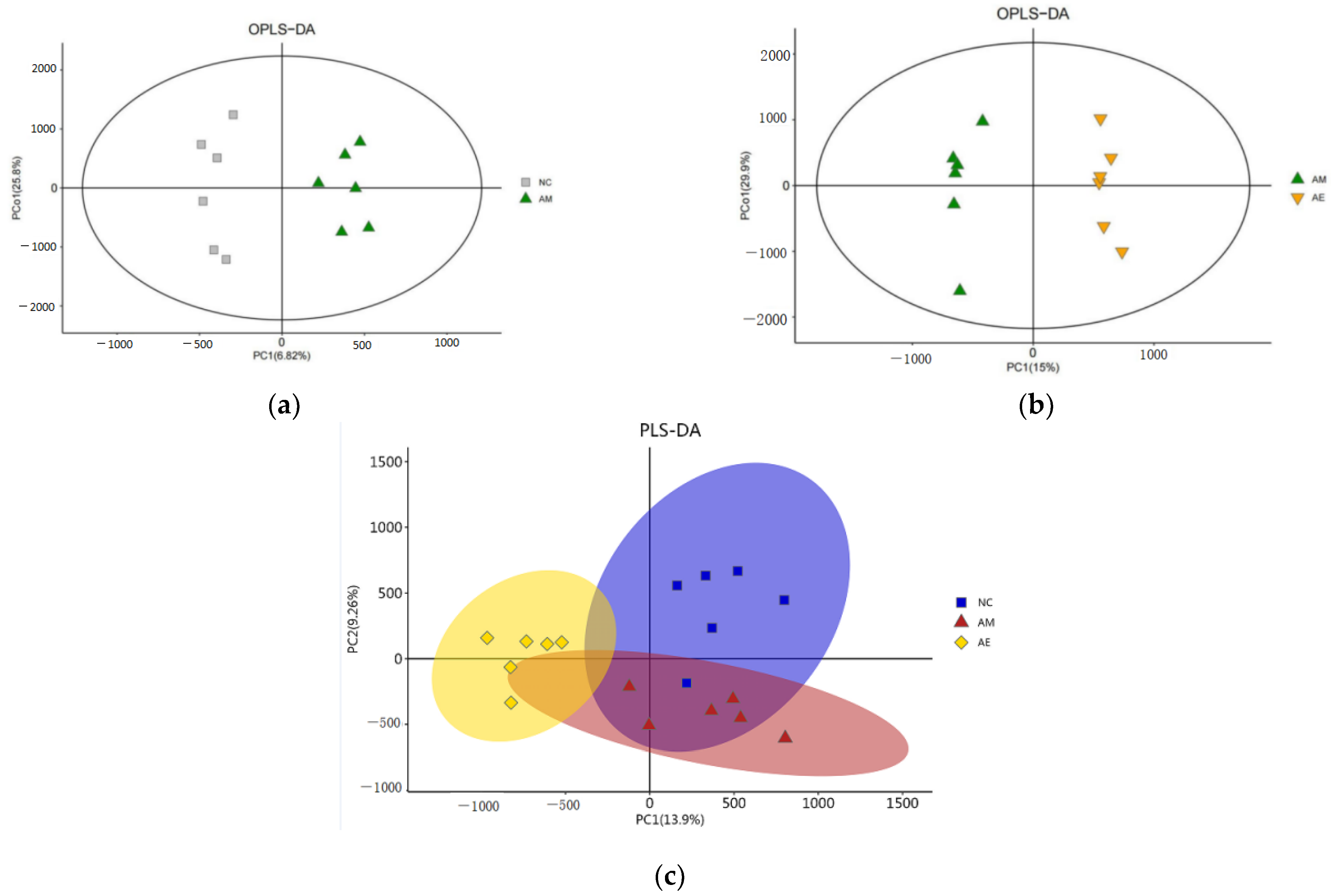
3.3. Results of (O)PLS-DA Analysis
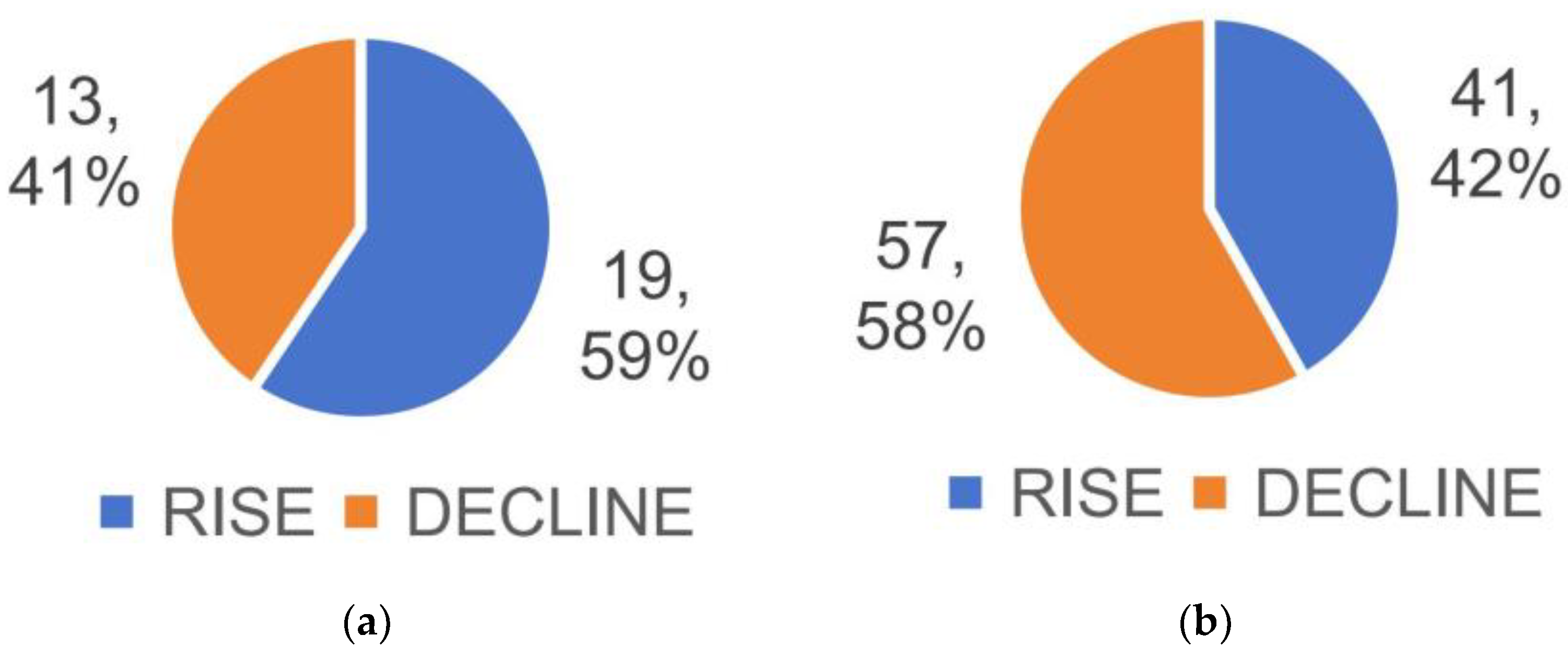
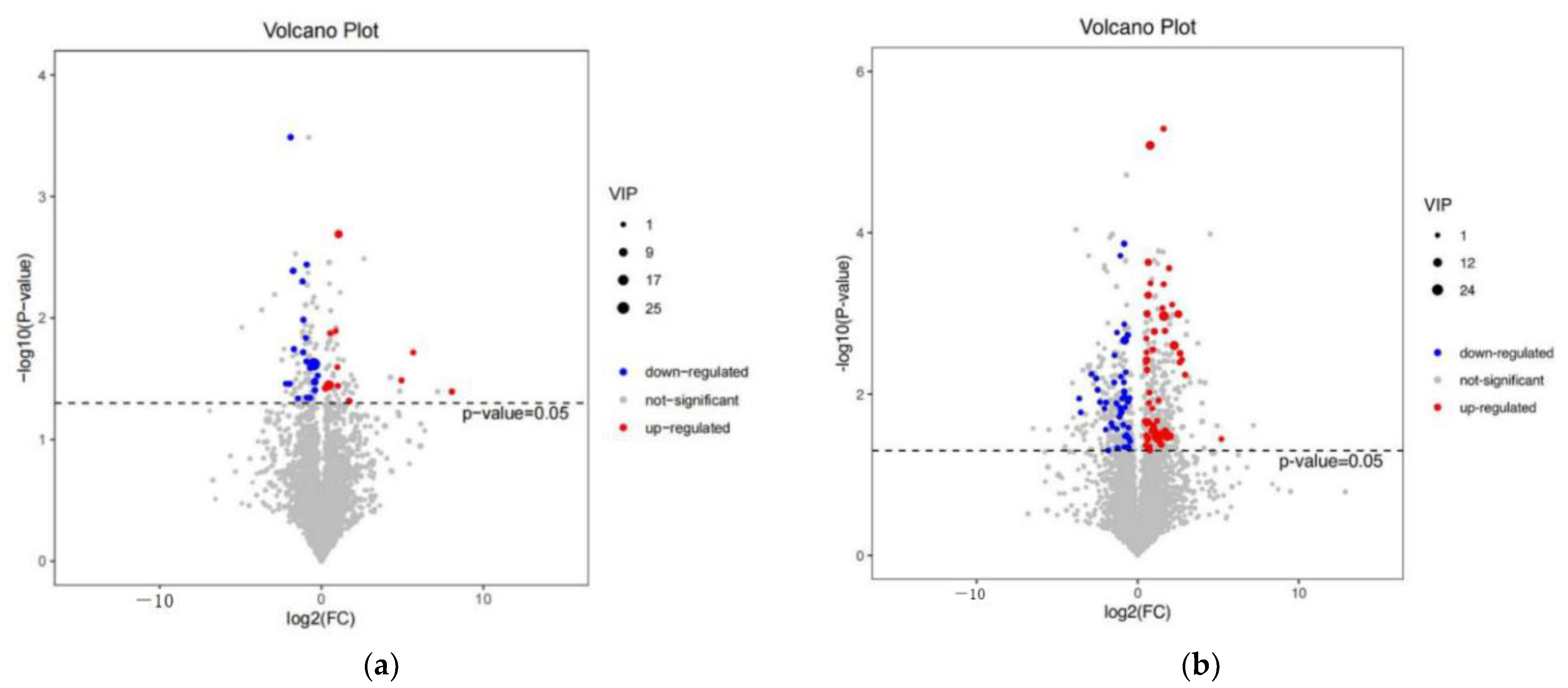
3.4. Differential Metabolite Identification
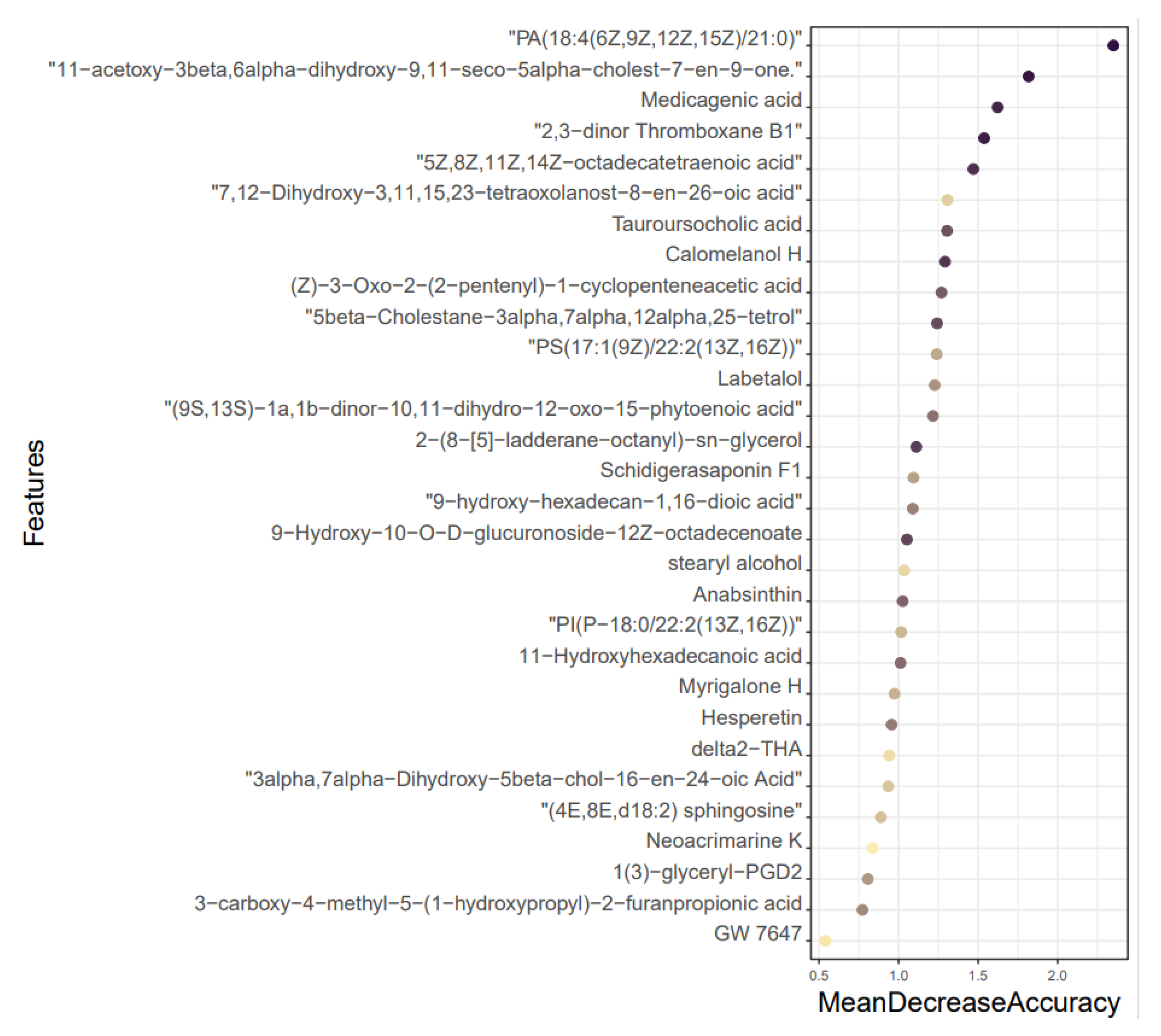
3.5. Results of Mean Decrease Accuracy
4. Discussion
4.1. Exercise Can Improve the Ability of Learning and Memory in AD Mice
4.2. Effect of Exercise on Lipid Metabolism in AD Mice
4.3. Effect of Exercise on Bile Metabolism of Intestinal Contents in AD Mice
4.4. Effect of Exercise on Phenylalanine Metabolism in AD Mice
5. Conclusions
6. Limitations of Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Patterson, C. World Alzheimer Report 2018: The State of the Art of Dementia Research: New Frontiers. An Analysis of Prevalence, Incidence, Cost and Trends; Alzheimer’s Disease International: London, UK, 2018. [Google Scholar]
- Huang, P.; Fang, R.; Li, B.Y.; Chen, S.D. Exercise-related changes of networks in aging and mild cognitive impairment brain. Front. Aging Neurosci. 2016, 8, 47. [Google Scholar] [CrossRef]
- Norton, S.; Matthews, F.E.; Barnes, D.E.; Yaffe, K.; Brayne, C. Potential for primary prevention of Alzheimer’s disease: An analysis of population-based data. Lancet Neurol. 2014, 13, 788–794. [Google Scholar] [CrossRef]
- Borges-Machado, F.; Ribeiro, Ó.; Sampaio, A.; Marques-Aleixo, I.; Meireles, J.; Carvalho, J. Feasibility and impact of a multicomponent exercise intervention in patients with Alzheimer’s disease: A pilot study. Am. J. Alzheimer’s Dis. Other Dement. 2019, 34, 95–103. [Google Scholar] [CrossRef]
- Arenaza-Urquijo, E.M.; de Flores, R.; Gonneaud, J.; Wirth, M.; Ourry, V.; Callewaert, W.; Landeay, B.; Egret, S.; Mezenge, F.; Desgranges, B.; et al. Distinct effects of late adulthood cognitive and physical activities on gray matter volume. Brain Imaging Behav. 2017, 11, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Abraham, D.; Feher, J.; Scuderi, G.L.; Szabo, D.; Dobolyi, A.; Cservenak, M.; Juhasz, J.; Ligeti, B.; Pongor, S.; Gomez-Cabrera, M.C.; et al. Exercise and probiotics attenuate the development of Alzheimer’s disease in transgenic mice: Role of microbiome. Exp. Gerontol. 2019, 115, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Rojas, C.; Aranguiz, F.; Varela-Nallar, L.; Inestrosa, N.C. Voluntary Running Attenuates Memory Loss, Decreases Neuropathological Changes and Induces Neurogenesis in a Mouse Model of Alzheimer’s Disease. Brain Pathol. 2016, 26, 62–74. [Google Scholar] [CrossRef]
- Li, X.; Wang, L.; Zhang, S.; Hu, X.; Yang, H.; Xi, L. Timing-Dependent Protection of Swimming Exercise against d-Galactose-Induced Aging-Like Impairments in Spatial Learning/Memory in Rats. Brain Sci. 2019, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gao, Q.; Gao, J.; Lv, L.; Liu, R.; Wu, Y.; Li, X.; Jin, Y.; Wang, L. Moderate-Intensity Intermittent Training Alters the DNA Methylation Pattern of PDE4D Gene in Hippocampus to Improve the Ability of Spatial Learning and Memory in Aging Rats Reduced by D-Galactose. Brain Sci. 2023, 13, 422. [Google Scholar] [CrossRef]
- Moore, K.M.; Girens, R.E.; Larson, S.K.; Jones, M.R.; Restivo, J.L.; Holtzman, D.M.; Cirrito, J.R.; Yuede, C.M.; Zimmerman, S.D.; Timson, B.F. A spectrum of exercise training reduces soluble Aβ in a dose-dependent manner in a mouse model of Alzheimer’s disease. Neurobiol. Dis. 2016, 85, 218–224. [Google Scholar] [CrossRef]
- Koo, J.H.; Kang, E.B.; Oh, Y.S.; Yang, D.S.; Cho, J.Y. Treadmill exercise decreases amyloid-β burden possibly via activation of SIRT-1 signaling in a mouse model of Alzheimer’s disease. Exp. Neurol. 2017, 288, 142–152. [Google Scholar] [CrossRef]
- Pimplikar, S.W.; Nixon, R.A.; Robakis, N.K.; Shen, J.; Tsai, L.H. Amyloid-independent mechanisms in Alzheimer’s disease pathogenesis. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 14946–14954. [Google Scholar] [CrossRef] [PubMed]
- Decker, H.; Lo, K.Y.; Unger, S.M.; Ferreira, S.T.; Silverman, M.A. Amyloid-beta peptide oligomers disrupt axonal transport through an Nmda receptor-dependent mechanism that is mediated by glycogen synthase kinase 3beta in primary cultured hippocampal neurons. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 9166–9171. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, H.; Ding, N.; Li, X.; Chen, X.; Chen, Z. Beneficial Effects on Brain Micro-Environment by Caloric Restriction in Alleviating Neurodegenerative Diseases and Brain Aging. Front. Physiol. 2021, 12, 715443. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhu, H.; Zhang, L.; Qin, C. The intestinal microbiome and Alzheimer’s disease: A review. Anim. Models Exp. Med. 2018, 1, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, B.O.; Bäckhed, F. Signals from the gut microbiota to distant organs in physiology and disease. Nat. Med. 2016, 22, 1079–1089. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.M.; Mailing, L.J.; Niemiro, G.M.; Moore, R.; Cook, M.D.; White, B.A.; Holschner, H.D.; Woods, J.A. Exercise alters gut microbiota composition and function in lean and obese humans. Med. Sci. Sports Exerc. 2018, 50, 747–757. [Google Scholar] [CrossRef]
- Chen, J.; Guo, Y.; Gui, Y.; Xu, D. Physical exercise, gut, gut microbiota, and atherosclerotic cardiovascular diseases. Lipids Health Dis. 2018, 17, 17. [Google Scholar] [CrossRef]
- Cook, M.D.; Allen, J.M.; Pence, B.D.; Wallig, M.A.; Gaskins, H.R.; White, B.A.; Woods, J.A. Exercise and gut immune function: Evidence of alterations in colon immune cell homeostasis and microbiome characteristics with exercise training. Immunol. Cell Biol. 2016, 94, 58–63. [Google Scholar] [CrossRef]
- National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Baker, E.J.; Gleeson, T.T. The effects of intensity on the energetics of brief locomotor activity. J. Exp. Biol. 1999, 202 Pt 22, 3081. [Google Scholar] [CrossRef]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Anderson, E.M.; Moenk, M.D.; Barbaro, L.; Clarke, D.A.; Matuszewich, L. Effects of pretraining and water temperature on female rats’ performance in the Morris water maze. Physiol. Behav. 2013, 122, 216–221. [Google Scholar] [CrossRef]
- Lombroso, P.J.; Ogren, M.P. Learning and memory, part I: Brain regions involved in two types of learning and memory. J. Am. Acad. Child Adolesc. Psychiatry 2008, 47, 1228–1232. [Google Scholar] [CrossRef]
- Alkadhi, K.A. Exercise as a Positive Modulator of Brain Function. Mol. Neurobiol. 2018, 55, 3112–3130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yi, P.; Yang, J.; Huang, J.; Xu, P.; Hu, M.; Znahg, C.; Wang, B.; Peng, W. Integrated network pharmacology analysis and serum metabolomics to reveal the cognitive improvement effect of Bushen Tiansui formula on Alzheimer’s disease. J. Ethnopharmacol. 2020, 249, 112371. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.A.; Scheff, S.W. Oxidative stress in the progression of Alzheimer disease in the frontal cortex. J. Neuropathol. Exp. Neurol. 2010, 69, 155–167. [Google Scholar] [CrossRef]
- Sweeney, G.; Song, J. The association between PGC-1α and Alzheimer’s disease. Anat. Cell Biol. 2016, 49, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Castro-Gómez, P.; Garcia-Serrano, A.; Visioli, F.; Fontecha, J. Relevance of dietary glycerophospholipids and sphingolipids to human health. Prostaglandins Leukot. Essent. Fatty Acids 2015, 101, 41–51. [Google Scholar] [CrossRef]
- Llano, D.A.; Devanarayan, V.; Alzheimer’s Disease Neuroimaging Initiative. Serum Phosphatidylethanolamine and Lysophosphatidylethanolamine Levels Differentiate Alzheimer’s Disease from Controls and Predict Progression from Mild Cognitive Impairment. J. Alzheimer’s Dis. 2021, 80, 311–319. [Google Scholar] [CrossRef]
- Bravo, F.V.; Da Silva, J.; Chan, R.B.; Di Paolo, G.; Teixeira-Castro, A.; Oliveira, T.G. Phospholipase D functional ablation has a protective effect in an Alzheimer’s disease Caenorhabditis elegans model. Sci. Rep. 2018, 8, 3540. [Google Scholar] [CrossRef]
- Sokolova, D.; Childs, T.; Hong, S. Insight into the role of phosphatidylserine in complement-mediated synapse loss in Alzheimer’s disease. Fac. Rev. 2021, 10, 19. [Google Scholar] [CrossRef]
- Wollen, K. Alzheimer’s disease: The pros and cons of pharmaceutical, nutritional, botanical, and stimulatory therapies, with a discussion of treatment strategies from the perspective of patients and practitioners. Altern. Med. Rev. 2010, 15, 223–244. [Google Scholar]
- Rockenfeller, P.; Koska, M.; Pietrocola, F.; Minois, N.; Knittelfelder, O.; Sica, V.; Franz, J.; Carmona-Gutierrez, D.; Kroemer, G.; Madeo, F. Phosphatidylethanolamine positively regulates autophagy and longevity. Cell Death Differ. 2015, 22, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, W.; Zan, J.; Wu, C.; Tan, W. Untargeted lipidomics reveals progression of early Alzheimer’s disease in APP/PS1 transgenic mice. Sci. Rep. 2020, 10, 14509. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, A.A.; Ong, W.-Y.; Horrocks, L.A. Biochemical aspects of neurodegeneration in human brain: Involvement of neural membrane phospholipids and phospholipases A 2. Neurochem. Res. 2004, 29, 1961–1977. [Google Scholar] [CrossRef]
- Sun, G.Y.; Xu, J.; Jensen, M.D.; Simonyi, A. Phospholipase A2 in the central nervous system: Implications for neurodegenerative diseases. J. Lipid Res. 2004, 45, 205–213. [Google Scholar] [CrossRef]
- Farooqui, A.A.; Horrocks, L.A. Phospholipase A₂-generated lipid mediators in the brain: The good, the bad, and the ugly. Neuroscientist 2006, 12, 245–260. [Google Scholar] [CrossRef]
- Vangaveti, V.; Baune, B.T.; Kennedy, R.L. Hydroxyoctadecadienoic acids: Novel regulators of macrophage differentiation and atherogenesis. Ther. Adv. Endocrinol. Metab. 2010, 1, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, E.; Owada, Y.; Ikawa, S.; Adachi, Y.; Egawa, T.; Nemoto, K.; Suzuki, K.; Hishinuma, T.; Kawashima, H.; Kondo, H.; et al. Epidermal FABP (FABP5) regulates keratinocyte differentiation by 13 (S)-HODE-mediated activation of the NF-κB signaling pathway. J. Investig. Dermatol. 2011, 131, 604–612. [Google Scholar] [CrossRef]
- Nieman, D.C.; Shanely, R.A.; Luo, B.; Meaney, M.P.; Dew, D.A.; Pappan, K.L. Metabolomics approach to assessing plasma 13-and 9-hydroxy-octadecadienoic acid and linoleic acid metabolite responses to 75-km cycling. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R68–R74. [Google Scholar] [CrossRef]
- MahmoudianDehkordi, S.; Arnold, M.; Nho, K.; Ahmad, S.; Jia, W.; Xie, G.; Louie, G.; Kueider-Paisley, A.; Srthur Moseley, M.; Thompson, J.W.; et al. Altered bile acid profile associates with cognitive impairment in Alzheimer’s disease—An emerging role for gut microbiome. Alzheimer’s Dement. 2019, 15, 76–92. [Google Scholar] [CrossRef]
- Morville, T.; Sahl, R.E.; Trammell, S.A.; Svenningsen, J.S.; Gillum, M.P.; Helge, J.W.; Clemmensen, C. Divergent effects of resistance and endurance exercise on plasma bile acids, FGF19, and FGF21 in humans. JCI Insight 2018, 3, e122737. [Google Scholar] [CrossRef] [PubMed]
- Squier, T.C. Oxidative stress and protein aggregation during biological aging. Exp. Gerontol. 2001, 36, 1539–1550. [Google Scholar] [CrossRef] [PubMed]
- Gregersen, N.; Bross, P. Protein misfolding and cellular stress: An overview. In Protein Misfolding and Cellular Stress in Disease and Aging; Humana Press: Totowa, NJ, USA, 2010; pp. 3–23. [Google Scholar] [CrossRef]
- Stocker, R.; Yamamoto, Y.; McDonagh, A.F.; Glazer, A.N.; Ames, B.N. Bilirubin is an antioxidant of possible physiological importance. Science 1987, 235, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Yang, Q.; Yu, N.; Cao, Y.; Wang, X.; Wang, Z.; Wen-Ying, Q.; Ma, C. Phenylalanine Metabolism is Dysregulated in Human Hippocampus with Alzheimer’s Disease Related Pathological Changes. J. Alzheimer’s Dis. 2021, 83, 609–622. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, H.H.; Luo, L.; Qin, L.Q.; Yin, J.; Yu, Z.; Zhang, L.; Wan, Z. Voluntary wheel running is capable of improving cognitive function only in the young but not the middle-aged male APPSwe/PS1De9 mice. Neurochem. Int. 2021, 145, 105010. [Google Scholar] [CrossRef] [PubMed]
- Hagena, H.; Manahan-Vaughan, D. Dopamine D1/D5, but not D2/D3, receptor dependency of synaptic plasticity at hippocampal mossy fiber synapses that is enabled by patterned afferent stimulation, or spatial learning. Front. Synaptic Neurosci. 2016, 8, 31. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Day 1 | Day 2 | Formal Campaign Interventions (20 Week) | ||
|---|---|---|---|---|---|
| Speed (m/min) | 5 | 8 | 10 | 15 | 10 |
| Time (min) | 10 | 10 | 5 | 20 | 5 |
| Group | Average Weight (Mean ± SD) | Hair | Appetite | Excreta |
|---|---|---|---|---|
| AE | 21 ± 2.45 | Normal | Normal | Normal |
| AM | 20 ± 1.65 | Sparse | Low | Low |
| NC | 24 ± 2.3 | Normal | Normal | Normal |
| NC | AM | AD | |
|---|---|---|---|
| Day1 | 92.44 ± 14.16 | 106.75 ± 13.39 | 103.64 ± 11.24 |
| Day2 | 68.84 ± 27.75 | 104.92 ± 21.79 | 61.11 ± 14.92 ▲ |
| Day3 | 41.38 ± 29.39 | 98.73 ± 17.57 ■ | 35.33 ± 16.88 ▲▲ |
| Day4 | 28.60 ± 8.03 | 86.34 ± 13.49 ■■ | 40.29 ± 13.97 ▲▲ |
| Day5 | 44.31 ± 21.86 | 95.04 ± 29.79 ■ | 35.04 ± 11.57 ▲▲ |
| Day6 | 21.49 ± 10.13 | 102.39 ± 15.26 ■■ | 18.79 ± 5.58 ▲▲ |
| Groups | Times of Crossing the Platform |
|---|---|
| NC | 2.96 ± 0.60 |
| AM | 0.67 ± 0.55 ■■ |
| AE | 2.01 ± 0.48 ▲ |
| No. | Metabolites | Amount of Expression | AM/NC | AE/AM | ||||
|---|---|---|---|---|---|---|---|---|
| NC | AM | AE | VIP | p-Value | VIP | p-Value | ||
| 1 | PA(18:4(6Z,9Z,12Z,15Z)/21:0) | 96.45 | 363.70 | 118.30 | 2.81 | 0.000 | 1.82 | <0.001 |
| 2 | 4,8-dimethylnonanoyl | 359.19 | 177.77 | 481.43 | 1.91 | 0.036 | 1.90 | 0.003 |
| 3 | PS(17:1(9Z)/22:2(13Z,16Z)) | 318.16 | 173.47 | 365.02 | 1.63 | 0.013 | 1.59 | <0.001 |
| 4 | 13(S)-HODE | 2320.98 | 3202.33 | 1940.018 | - | - | 3.26 | 0.04 |
| 5 | 5beta-Cyprinolsulfate | 4356.15 | 1570.77 | 2802.11 | - | - | 3.54 | 0.02 |
| 6 | Bilirubin | 4845.45 | 618.18 | 920.51 | - | - | 7.49 | <0.001 |
| 7 | D-Urobilinogen | 1157.84 | 491.56 | 709.93 | - | - | 3.72 | 0.04 |
| 8 | PE(14:0/22:2(13Z,16Z)) | 2616.74 | 953.76 | 1119.14 | - | - | 1.95 | 0.01 |
| 9 | Phenylacetylglycine | 1298.78 | 245.91 | 815.51 | - | - | 2.09 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhang, Y.; Ding, X.; Jin, Y.; Wei, C.; Xu, J. Mass Spectrometry Chromatography-Based Metabolomics: The Effect of Long-Term Aerobic Exercise on Learning Ability and the Metabolism of Intestinal Contents in Mice with Alzheimer’s Disease. Metabolites 2023, 13, 1150. https://doi.org/10.3390/metabo13111150
Li X, Zhang Y, Ding X, Jin Y, Wei C, Xu J. Mass Spectrometry Chromatography-Based Metabolomics: The Effect of Long-Term Aerobic Exercise on Learning Ability and the Metabolism of Intestinal Contents in Mice with Alzheimer’s Disease. Metabolites. 2023; 13(11):1150. https://doi.org/10.3390/metabo13111150
Chicago/Turabian StyleLi, Xue, Yuanting Zhang, Xianyi Ding, Yu Jin, Changling Wei, and Jisheng Xu. 2023. "Mass Spectrometry Chromatography-Based Metabolomics: The Effect of Long-Term Aerobic Exercise on Learning Ability and the Metabolism of Intestinal Contents in Mice with Alzheimer’s Disease" Metabolites 13, no. 11: 1150. https://doi.org/10.3390/metabo13111150
APA StyleLi, X., Zhang, Y., Ding, X., Jin, Y., Wei, C., & Xu, J. (2023). Mass Spectrometry Chromatography-Based Metabolomics: The Effect of Long-Term Aerobic Exercise on Learning Ability and the Metabolism of Intestinal Contents in Mice with Alzheimer’s Disease. Metabolites, 13(11), 1150. https://doi.org/10.3390/metabo13111150