



Energy System Contributions during Olympic Combat Sports: A Narrative Review


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Energy Systems’ Contributions in Combat Sports
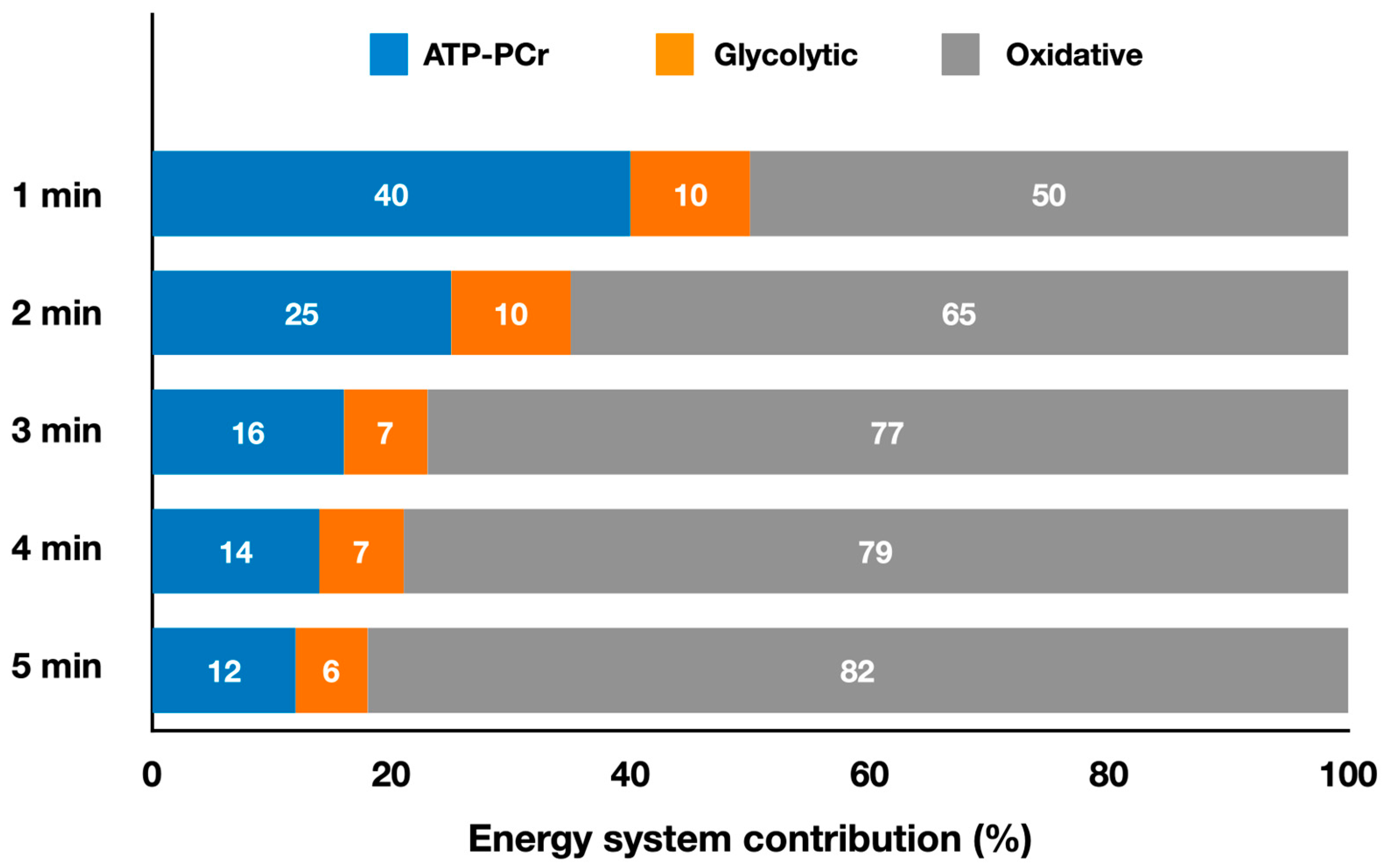
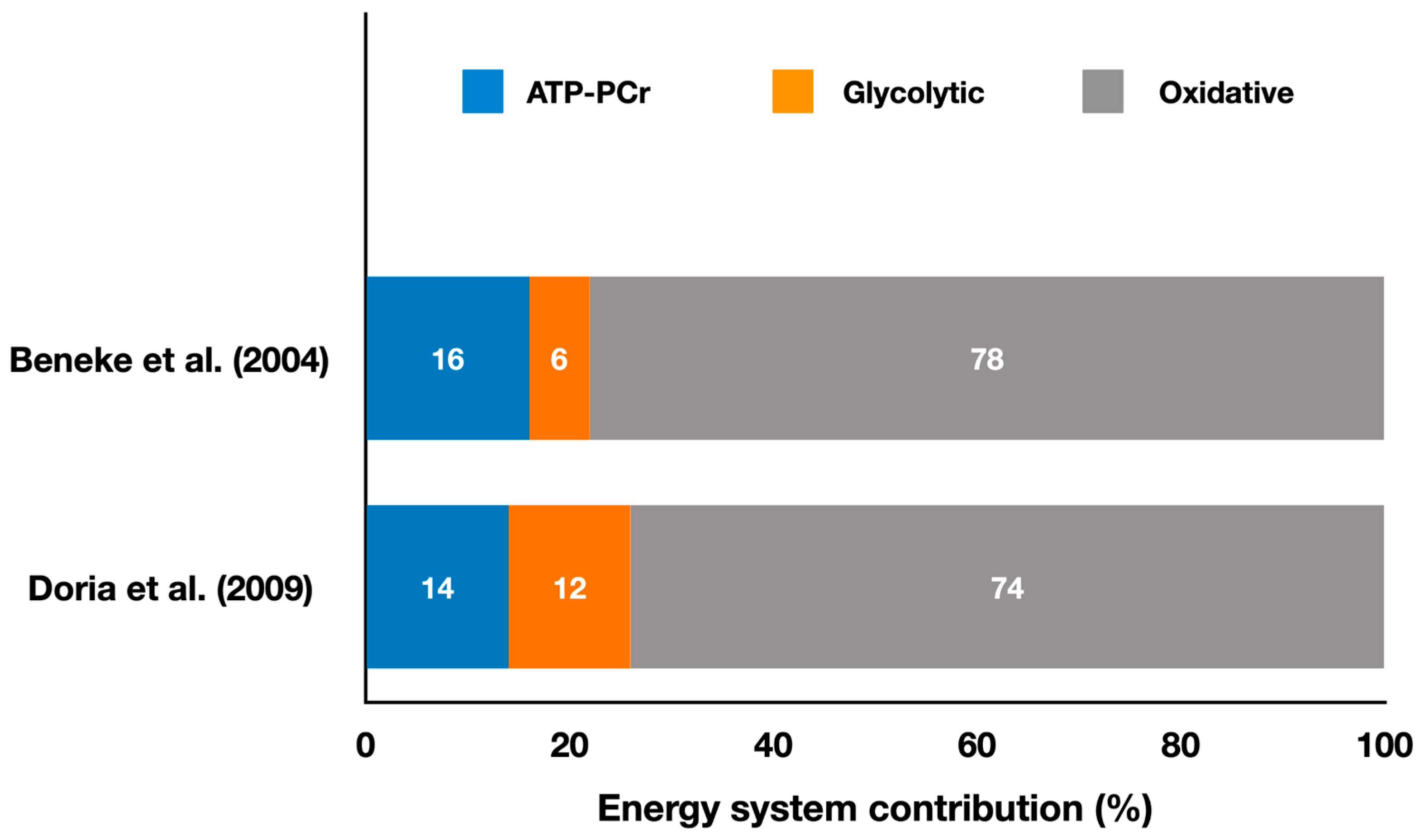
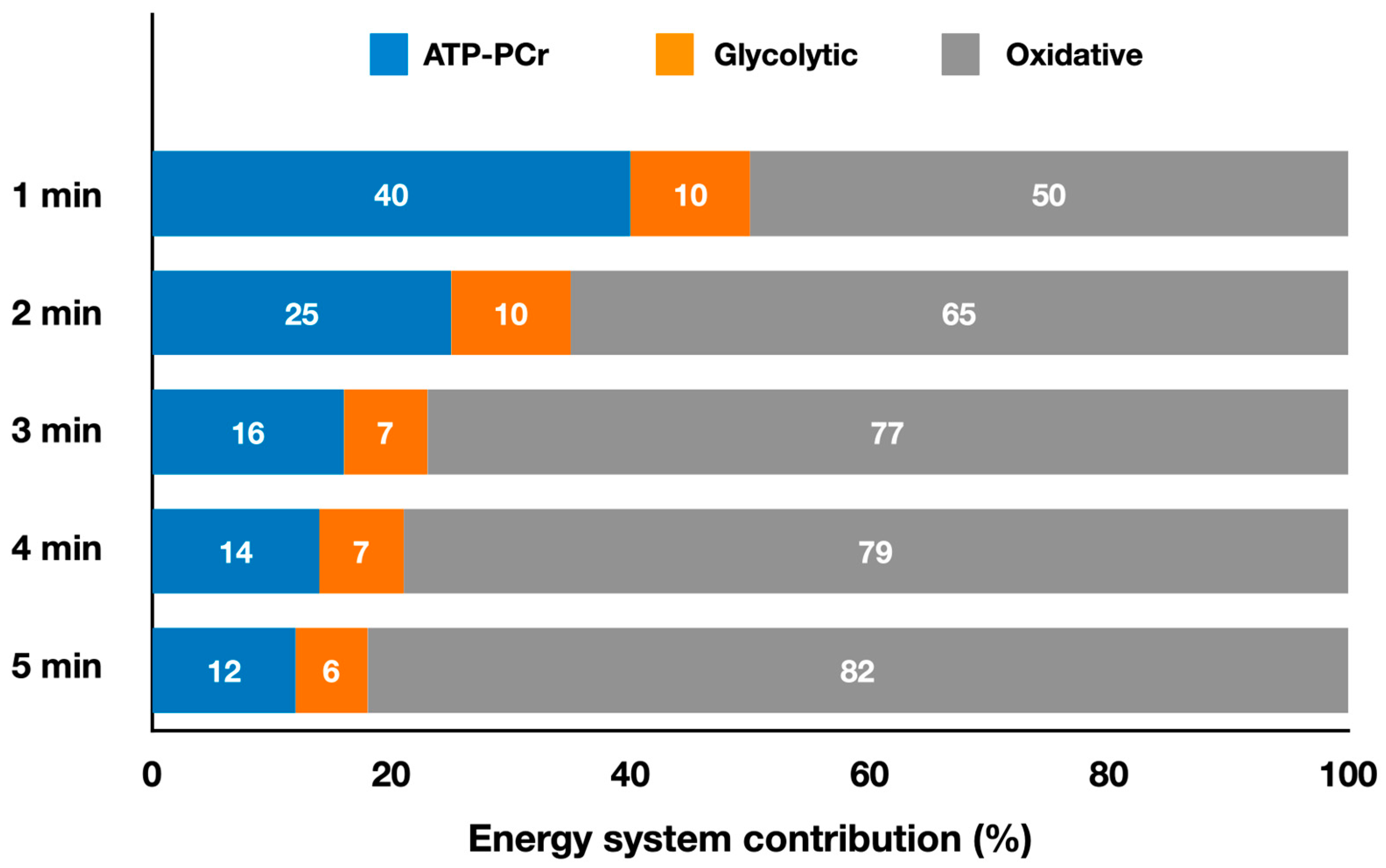
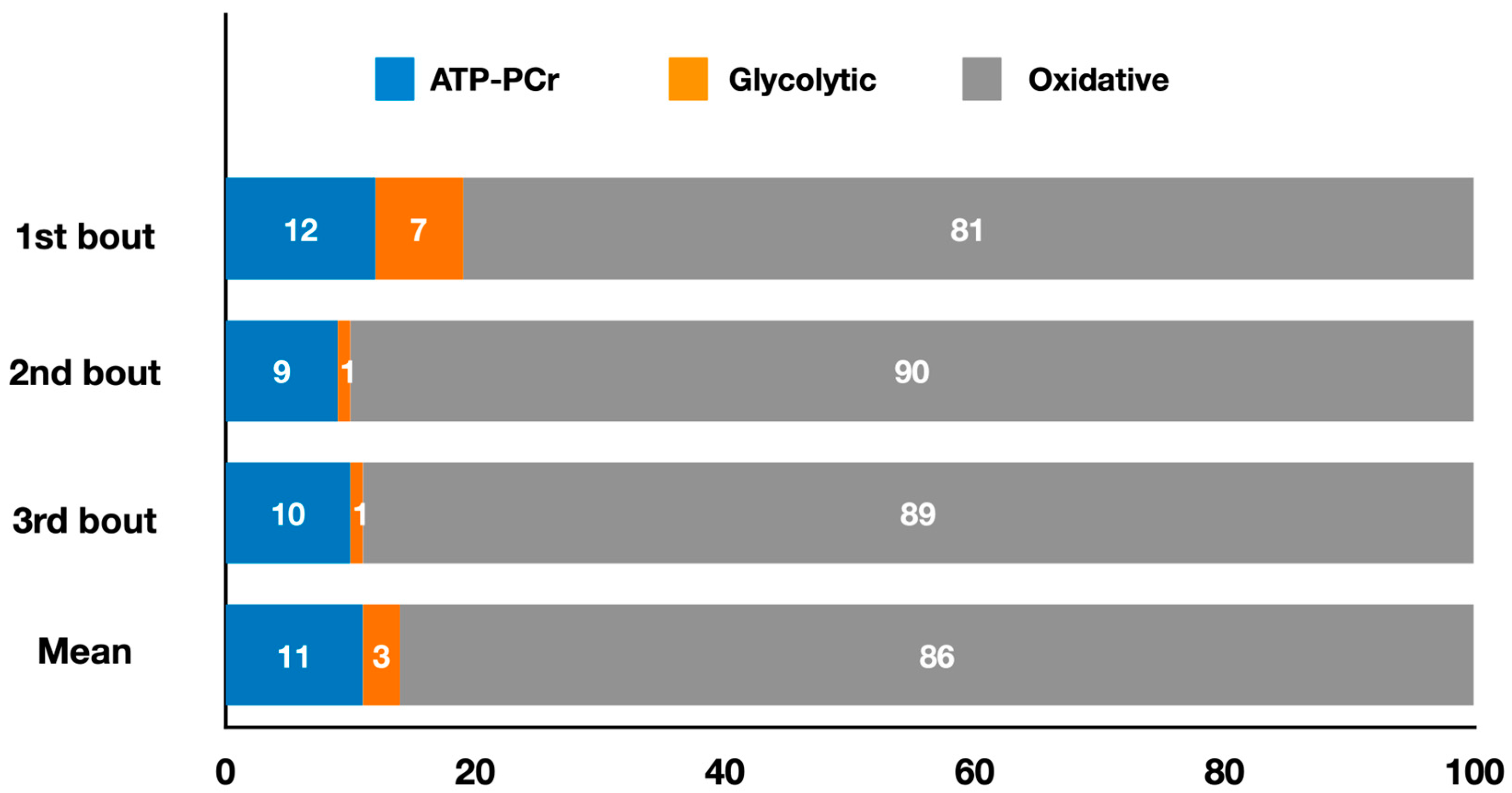
2.1. Striking Olympic Combat Sports
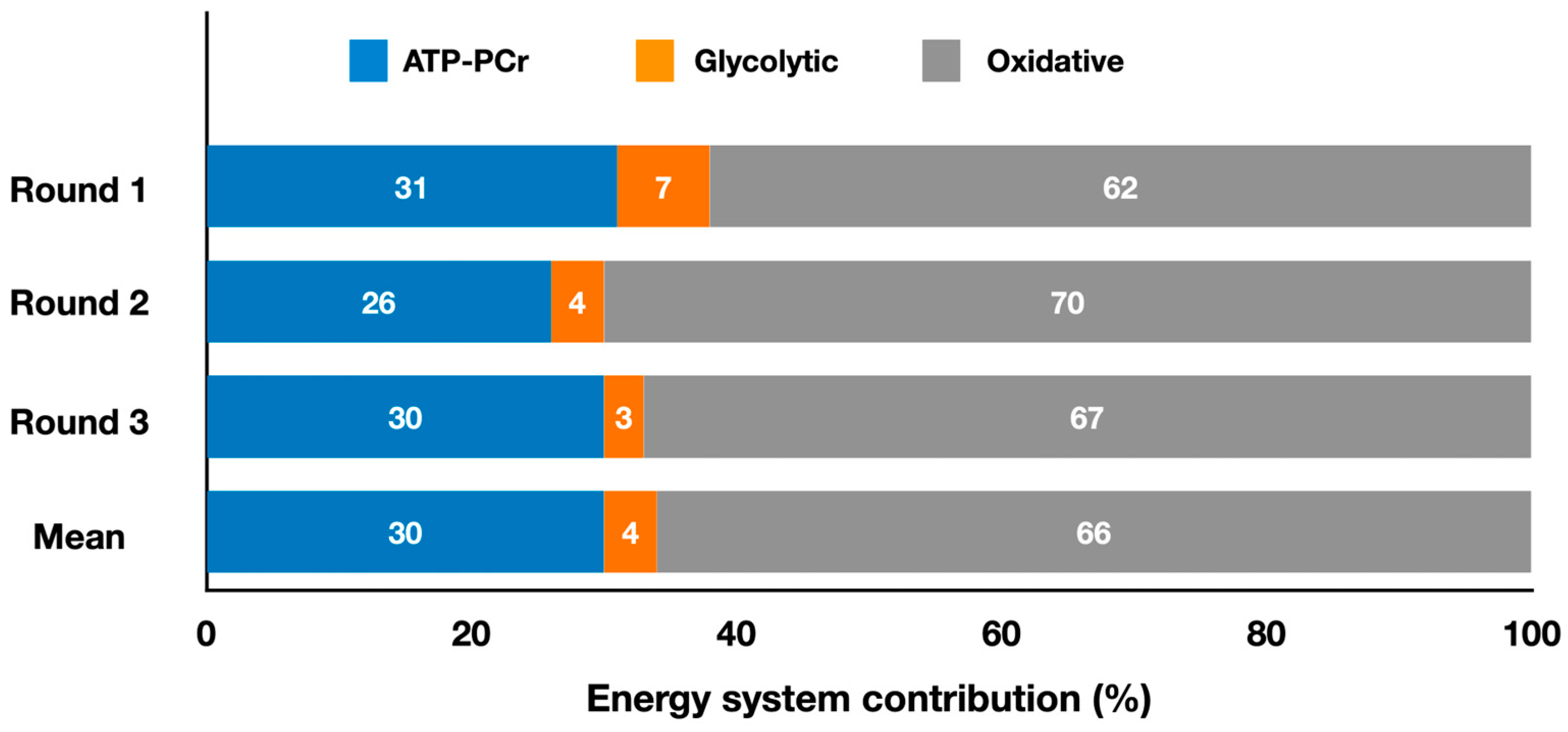
2.2. Grappling Olympic Combat Sports
2.3. Weapon-Based Combat Sport
3. Conclusions
Funding
Conflicts of Interest
References
- International Olympic Committee. 2022. Available online: https://olympics.com/es/olympic-games/tokyo-2020 (accessed on 16 November 2022).
- Ko, Y.J.; Kim, Y.K.; Valacich, J. Martial arts participation: Consumer motivation. Int. J. Sports Market. Sponsorship. 2010, 11, 105–123. [Google Scholar] [CrossRef]
- Franchini, E.; Cormack, S.; Takito, M.Y. Effects of high-intensity interval training on Olympic combat sports athletes’ performance and physiological adaptation: A systematic review. J. Strength Cond. Res. 2019, 33, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Franchini, E.; Herrera-Valenzuela, T. Special issue—strength and conditioning for combat sports athletes. Rev. Art. Marc. Asi. 2021, 16, 1–203. [Google Scholar] [CrossRef]
- Bridge, C.A.; Santos, J.F.S.; Chaabene, H.; Pieter, W.; Franchini, E. Physical and physiological profiles of taekwondo athletes. Sports Med. 2014, 44, 713–733. [Google Scholar] [CrossRef] [PubMed]
- Chaabene, H.; Hachana, Y.; Franchini, E.; Mkaouer, B.; Chamari, K. Physical and physiological profile of elite karate athletes. Sports Med. 2012, 42, 829–843. [Google Scholar] [CrossRef] [PubMed]
- Franchini, E.; Del Vecchio, F.B.; Matsushigue, K.A.; Artioli, G.G. Physiological profiles of elite judo athletes. Sports Med. 2011, 41, 147–166. [Google Scholar] [CrossRef] [PubMed]
- Lassalvia, C.; Julio, U.F.; Franchini, E. Physiological responses during karate kata in practitioners and athletes: A scoping review. Strength. Cond. J. 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Artioli, G.G.; Bertuzzi, R.C.; Roschel, H.; Mendes, S.H.; Lancha, A.H.; Franchini, E. Determining the contribution of the energy systems during exercise. J. Vis. Exp. 2012, 61, 3413. [Google Scholar] [CrossRef]
- Andreato, L.V.; Julio, U.F.; Panissa, V.L.; Esteves, J.V.; Hardt, F.; Moraes, S.M.; Souza, C.O.; Franchini, E. Brazilian jiu-jitsu competition Part I: Metabolic, hormonal, cellular damage, and heart rate responses. J. Strength Cond. Res. 2015, 29, 2538–2549. [Google Scholar] [CrossRef]
- Andreato, L.V.; Julio, U.F.; Panissa, V.L.; Esteves, J.V.; Hardt, F.; Moraes, S.M.; Souza, C.O.; Franchini, E. Brazilian jiu-jitsu competition Part II: Physical performance, time-motion, technical-tactical analyses, and perceptual responses. J. Strength Cond. Res. 2015, 29, 2015–2025. [Google Scholar] [CrossRef]
- Beneke, R.; Beyer, T.; Jachner, C.; Erasmus, J.; Hütler, M. Energetics of karate kumite. Eur. J. Appl. Physiol. 2004, 92, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Campos, F.A.; Bertuzzi, R.C.M.; Dourado, A.C.; Santos, V.G.; Franchini, E. Energy demands in taekwondo athletes during combat simulation. Eur. J. Appl. Physiol. 2012, 112, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.; Leithäuser, R.M.; Beneke, R. The energetics of semicontact 3 x 2-min amateur boxing. Int. J. Sports Physiol. Perform. 2014, 9, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Doria, C.; Veicsteinas, A.; Limonta, E.; Maggioni, M.A.; Aschieri, P.; Eusebi, F.; Fano, G.; Pietrangelo, T. Energetics of karate (kata and kumite techniques) in top-level athletes. Eur. J. Appl. Physiol. 2009, 107, 603–610. [Google Scholar] [CrossRef]
- Julio, U.F.; Panissa, V.L.G.; Esteves, J.V.; Cury, R.L.; Agostinho, M.F.; Franchini, E. Energy-system contributions to simulated judo matches. Int. J. Sports Physiol. Perform. 2017, 12, 676–683. [Google Scholar] [CrossRef]
- Ouergui, I.; Benyoussef, A.; Houcine, N.; Abedelmalek, S.; Franchini, E.; Gmada, N.; Bouhlel, E.; Bouassida, A. Physiological responses and time-motion analysis of kickboxing: Differences between full contact, light contact, and point fighting contests. J. Strength Cond. Res. 2021, 35, 2558–2563. [Google Scholar] [CrossRef]
- Yang, W.H.; Park, J.H.; Shin, Y.C.; Kim, J. Physiological profiling and energy system contributions during simulated epée matches in elite fencers. Int. J. Sports Physiol. Perform. 2022, 17, 943–950. [Google Scholar] [CrossRef]
- Franchini, E.; Nakamura, F.Y.; Takito, M.Y.; Kiss, M.A.P.D.; Sterkowicz, S. Specific test developed in Brazilian judoists. Biol. Sport 1998, 15, 165–170. [Google Scholar]
- Krstulovic, S.; Kuvacic, G.; Erceg, M.; Franchini, E. Reliability and validity of the new judo physical fitness test. Ido Mov. Cult. 2019, 19, 41–45. [Google Scholar] [CrossRef]
- Ouergui, I.; Houcine, N.; Marzouki, H.; Davis, P.; Zaouali, M.; Franchini, E.; Gmada, N.; Bouhlel, E. Development of a noncontact kickboxing circuit training protocol that simulates elite male kickboxing competition. J. Strength Cond. Res. 2015, 29, 3405–3411. [Google Scholar] [CrossRef]
- Santos, J.F.D.S.; Franchini, E. Frequency speed of kick test performance comparison between female taekwondo athletes of different competitive levels. J. Strength Cond. Res. 2018, 32, 2934–2938. [Google Scholar] [CrossRef]
- Santos, L.; Fernández-Rio, J.; Iglesias-Soler, E.; Blanco-Traba, M.; Jakobsen, M.D.; González-Díez, V.; Franchini, E.; Gutiérrez, C.; Dopico-Calvo, X.; Carballeira-Fernández, E.; et al. Postural control and physiological responses to simulated match in U-20 judo competitors. Sports Biomech. 2020, 19, 281–294. [Google Scholar] [CrossRef]
- Herrera-Valenzuela, T.; Cuadra, D.; Valdés-Badilla, P.; Cofre-Bolados, C.; Pardo-Tamayo, C.; Ojeda-Aravena, A.; Franchini, E. Relación del Special Wrestling Fitness Test con el rendimiento aeróbico. Rev. And. Med. Dep. 2021, 14, 98–102. [Google Scholar] [CrossRef]
- Davis, P.; Benson, P.R.; Pitty, J.D.; Connorton, A.J.; Waldock, R. The activity profile of elite male amateur boxing. Int. J. Sports Physiol. Perform. 2015, 10, 53–57. [Google Scholar] [CrossRef]
- Franchini, E.; Artioli, G.G.; Brito, C.J. Judo combat: Time-motion analysis and physiology. Int. J. Perf. Anal. Sport 2013, 13, 624–641. [Google Scholar] [CrossRef]
- Miarka, B.; Vecchio, F.B.; Carney, S.; Amatmann, J.A. Comparisons: Technical-tactical and time-motion analysis of mixed martial arts by outcomes. J. Strength Cond. Res. 2016, 30, 1975–1984. [Google Scholar] [CrossRef]
- Nilsson, J.; Csergö, S.; Gullstrand, L.; Tveit, P.; Refsnes, P.E. Work-time profile, blood lactate concentration and rating of perceived exertion in the 1998 Greco-Roman Wrestling World Championship. J. Sports Sci. 2002, 20, 939–945. [Google Scholar] [CrossRef]
- Ouergui, I.; Hssin, N.; Haddad, M.; Franchini, E.; Behm, D.G.; Wong, P.; Gamma, N.; Boule, E. Time-motion analysis of elite male kickboxing competition. J. Strength Cond. Res. 2014, 28, 3537–3543. [Google Scholar] [CrossRef]
- Santos, V.G.; Franchini, E.; Lima-Silva, A.E. Relationship between attack and skipping in taekwondo contests. J. Strength Cond. Res. 2011, 25, 1743–1751. [Google Scholar] [CrossRef]
- Tabben, M.; Coquart, J.; Chaabene, H.; Franchini, E.; Ghoul, N.; Tourny, C. Time-motion, tactical and technical analysis in top-level karatekas according to gender, match outcome and weight categories. J. Sports Sci. 2015, 33, 841–849. [Google Scholar] [CrossRef]
- Wylde, M.J.; Tan, F.H.Y.; O’Donoghue, P.G. A time-motion analysis of elite women’s foil fencing. Int. J. Perf. Anal. Sport 2013, 13, 365–376. [Google Scholar] [CrossRef]
- Franchini, E.; Takito, M.Y.; Kiss, M.A.P.D.M.; Sterkowicz, S. Physical fitness and anthropometrical differences between elite and non-elite judo players. Biol. Sport. 2005, 22, 315–328. [Google Scholar]
- Hoschel, H.; Batista, M.; Monteiro, R.; Bertuzzi, R.C.; Barroso, R.; Loturco, I.; Ugrinowitsch, C.; Tricoli, V.; Franchinim, E. Association between neuromuscular tests and kumite performance on the Brazilian karate national team. J. Sports Sci. Med. 2009, 8, 20–24. [Google Scholar]
- James, L.P.; Beckman, E.M.; Kelly, V.G.; Haff, G.G. The neuromuscular qualities of higher- and lower-level mixed martial arts competitors. Int. J. Sports Physiol. Perform. 2017, 12, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Marinho, B.F.; Andreato, L.V.; Follmer, B.; Franchini, E. Comparison of body composition and physical fitness in elite and non-elite Brazilian jiu-jitsu athletes. Sci. Sports 2016, 31, 129–134. [Google Scholar] [CrossRef]
- Chaabene, H.; Negra, Y.; Bouguezzi, R.; Capranica, L.; Franchini, E.; Prieske, O.; Hbacha, H.; Granacher, U. Tests for the assessment of sport-specific performance in Olympic combat sports: A systematic review with practical recommendations. Front. Physiol. 2018, 9, 386. [Google Scholar] [CrossRef] [PubMed]
- Franchini, E. High-intensity interval training prescription for combat-sport athletes. Int. J. Sports Physiol. Perform. 2020, 15, 767–776. [Google Scholar] [CrossRef]
- Slimani, M.; Chaabene, H.; Miarka, B.; Franchini, E.; Chamari, K.; Cheour, F. Kickboxing review: Anthropometric, psychophysiological and activity profiles and injury epidemiology. Biol. Sport 2017, 34, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Slimani, M.; Chaabene, H.; Davis, P.; Franchini, E.; Cheour, F.; Chamari, K. Performance aspects and physiological responses in male amateur boxing competitions: A brief review. J. Strength Cond. Res. 2017, 31, 1132–1141. [Google Scholar] [CrossRef]
- Chaabene, H.; Tabben, M.; Mkaouer, B.; Franchini, E.; Negra, Y.; Hammami, M.; Amara, S.; Chaabene, R.B.; Hachana, Y. Amateur boxing: Physical and physiological attributes. Sports Med. 2015, 45, 337–352. [Google Scholar] [CrossRef]
- Chaabene, H.; Negra, Y.; Bouguezzi, R.; Mkaouer, B.; Franchini, E.; Julio, U.; Hachana, Y. Physical and physiological attributes of wrestlers: An update. J. Strength Cond. Res. 2017, 31, 1411–1442. [Google Scholar] [CrossRef] [PubMed]
- Finlay, M.J.; Greig, M.; Page, R.M.; Bridge, C.A. Acute physiological, endocrine, biochemical and performance responses associated with amateurs boxing: A systematic review with meta-analysis. Eur. J. Sport Sci. 2022. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Kons, R.; Orsatto, L.B.R.; Detanico, D. Acute performance responses during repeated matches in combat sports: A systematic review. J. Sci. Med. Sport 2020, 23, 512–518. [Google Scholar] [CrossRef]
- Smith, D.J. A framework for understanding the training process leading to elite performance. Sports Med. 2003, 33, 1103–1126. [Google Scholar] [CrossRef] [PubMed]
- Gastin, P. Energy system interaction and relative contribution during maximal exercise. Sports Med. 2001, 31, 725–741. [Google Scholar] [CrossRef] [PubMed]
- Gaitanos, G.C.; Williams, C.; Boobis, L.H.; Brooks, S. Human muscle metabolism during intermittent maximal exercise. J. Appl. Physiol. 1993, 75, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Medbo, J.; Mohn, A.C.; Tanita, I.; Bahr, R.; Vaage, O.; Sejersted, O.M. Anaerobic capacity determined by maximal accumulated O2 deficit. J. Appl. Physiol. 1988, 64, 50–60. [Google Scholar] [CrossRef] [Green Version]
- di Prampero, P.E.; Ferretti, G. The energetics of anaerobic muscle metabolism: A reappraisal of older and recent concepts. Respir. Physiol. 1999, 118, 103–115. [Google Scholar] [CrossRef]
- Péronnet, F.; Thibault, G. Mathematical analysis of running performance and world running records. J. Appl. Physiol. 1989, 67, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Lopes-Silva, J.P.; Silva Santos, J.F.; Branco, B.H.; Abad, C.C.; Oliveira, L.F.; Loturco, I.; Franchini, E. Caffeine ingestion increases estimated glycolytic metabolism during taekwondo combat simulation but does not improve performance or parasympathetic reactivation. PLoS ONE 2015, 10, e0142078. [Google Scholar] [CrossRef]
- Lopes-Silva, J.P.; Da Silva Santos, J.F.; Artioli, G.G.; Loturco, I.; Abbiss, C.; Franchini, E. Sodium bicarbonate ingestion increases glycolytic contribution and improves performance during simulated taekwondo combat. Eur. J. Sport Sci. 2018, 18, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Loturco, I.; Nakamura, F.Y.; Lopes-Silva, J.P.; Silva-Santos, J.F.; Pereira, L.A.; Franchini, E. Physical and physiological traits of a double world karate champion and responses to a simulated kumite bout: A case study. Int. J. Sports Sci. Coach. 2017, 2, 138–147. [Google Scholar] [CrossRef]
- Yang, W.H.; Heine, O.; Grau, M. Rapid weight reduction does not impair athletic performance of taekwondo athletes—A pilot study. PLoS ONE 2018, 13, e0196568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossiter, H.B. Exercise: Kinetic considerations for gas exchange. Compr. Physiol. 2011, 1, 203–244. [Google Scholar] [CrossRef] [PubMed]
- Bertuzzi, R.C.M.; Franchini, E.; Ugrinowitsch, C.; Kokubun, E.; Lima-Silva, A.E.; Pires, F.O.; Nakamura, F.Y.; Kiss, M.A.P.D.M. Predicting MAOD using only a supra maximal exhaustive test. Int. J. Sports Med. 2010, 31, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Zagatto, A.M.; Bertuzzi, R.; Miyagi, W.E.; Padulo, J.; Papoti, M. MAOD determined in a single supramaximal test: A study on the relia- bility and effects of supramaximal intensities. Int. J. Sports Med. 2016, 37, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Hill, D.W.; Riojas, A.E.; McFarlin, B.K.; Vingren, J.L. An alternative to oxygen deficit as a way to quantify anaerobic contribution in running. J. Hum. Sport Exerc. 2020, 15, 837–848. [Google Scholar] [CrossRef]
- Hill, D.W. Calculation of anaerobic capacity in running and cycling using post-exercise measures. Sci. Sport 2023. ahead of print. [Google Scholar] [CrossRef]
- Valenzuela, J.R.; Riojas, A.E.; McFarlin, B.K.; Vingren, J.L.; Hill, D.W. Determining MAOD using a single exhaustive severe intensity test. Int. J. Exerc. Sci. 2020, 13, 702–713. [Google Scholar]
- Miyagi, W.E.; de Poli, R.A.B.; Papoti, M.; Bertuzzi, R.; Zagatto, A.M. Anaerobic capacity estimated in a single supramaximal test in cycling: Validity and reliability analysis. Sci. Rep. 2017, 7, 42485. [Google Scholar] [CrossRef] [Green Version]
- Panissa, V.L.G.; Fukuda, D.H.; Caldeira, R.S.; Gerosa-Neto, J.; Lira, F.S.; Zagatto, A.M.; Franchini, E. Is oxygen uptake measurement enough to estimate the energy expenditure during high-intensity intermittent exercise? Quantification of anaerobic contribution by different methods. Front. Physiol. 2018, 9, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franchini, E.; Brito, C.J.; Artioli, G.G. Weight loss in combat sports: Physiological, psychological and performance effects. J. Int. Soc. Sports Nut. 2012, 9, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, J.F.S.; Artioli, G.G.; Takito, M.Y.; Franchini, E. Weight loss practices in taekwondo athletes of different competitive levels. J. Exerc. Rehab. 2016, 12, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Brechney, G.C.; Cannon, J.; Goodman, S.P. Effects of weight cutting on exercise performance in combat athletes: A meta-analysis. Int. J. Sports Physiol. Perform. 2022, 17, 995–1010. [Google Scholar] [CrossRef]
- Reale, R.; Slater, G.; Burke, L.M. Acute weight loss strategies for combat sports and applications to Olympic success. Int. J. Sports Physiol. Perform. 2017, 12, 142–151. [Google Scholar] [CrossRef]
- Kashiwagura, D.; Courel-Ibáñez, J.; Kashiwagura, F.B.; Agostinho, M.F.; Franchini, E. Judo technical-tactical dynamics: Analysis of attack system effectiveness in high-level athletes. Int. J. Perf. Anal. Sport 2022, 21, 922–933. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franchini, E. Energy System Contributions during Olympic Combat Sports: A Narrative Review. Metabolites 2023, 13, 297. https://doi.org/10.3390/metabo13020297
Franchini E. Energy System Contributions during Olympic Combat Sports: A Narrative Review. Metabolites. 2023; 13(2):297. https://doi.org/10.3390/metabo13020297
Chicago/Turabian StyleFranchini, Emerson. 2023. "Energy System Contributions during Olympic Combat Sports: A Narrative Review" Metabolites 13, no. 2: 297. https://doi.org/10.3390/metabo13020297
APA StyleFranchini, E. (2023). Energy System Contributions during Olympic Combat Sports: A Narrative Review. Metabolites, 13(2), 297. https://doi.org/10.3390/metabo13020297