
Maternal High-Fat Diet Consumption in Sprague Dawley Rats Compromised the Availability and Altered the Tissue Distribution of Lutein in Neonatal Offspring


Abstract
:1. Introduction
2. Materials and Methods
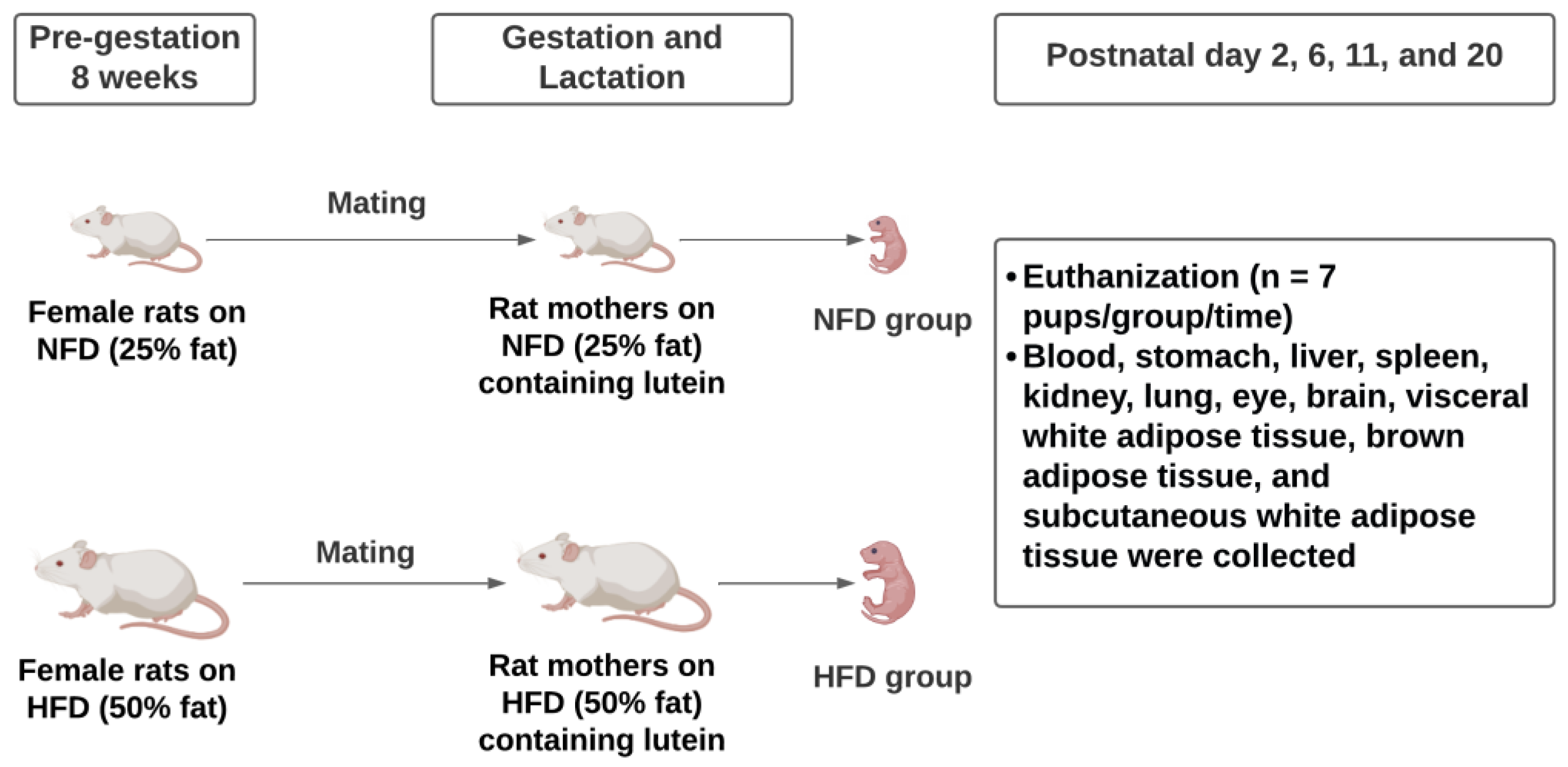
2.1. Animal Experiment
2.2. Analysis of Lutein Concentration in Serum, Milk, and Tissues
2.3. Statistical Analysis
3. Results
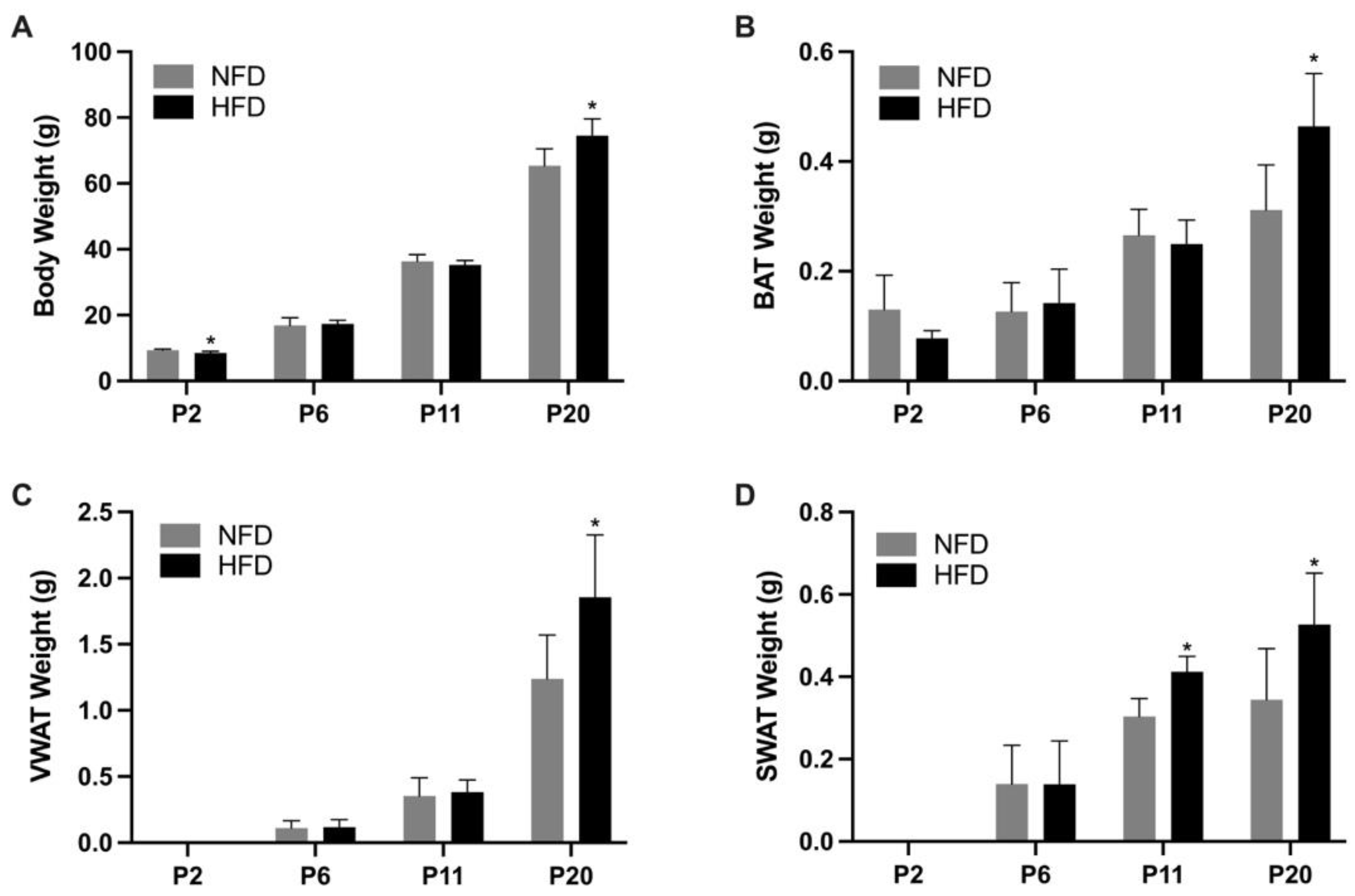
3.1. Dietary Intake, Body Weight, and Adiposity
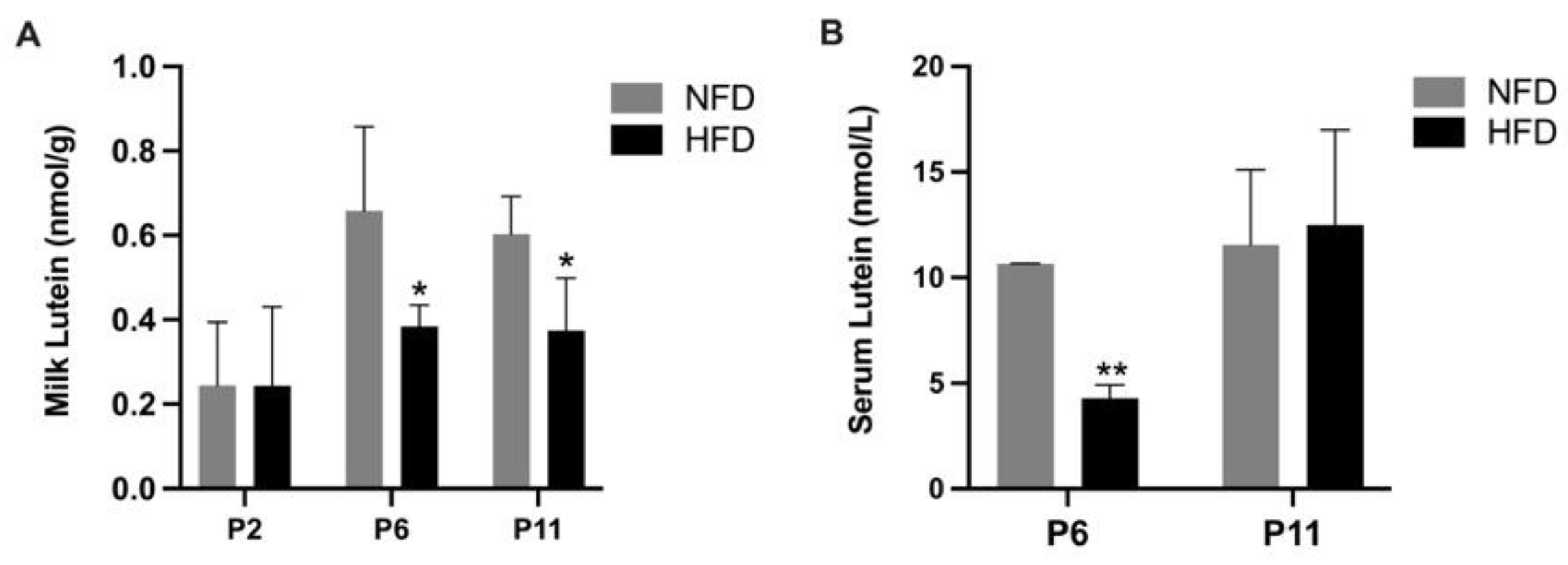
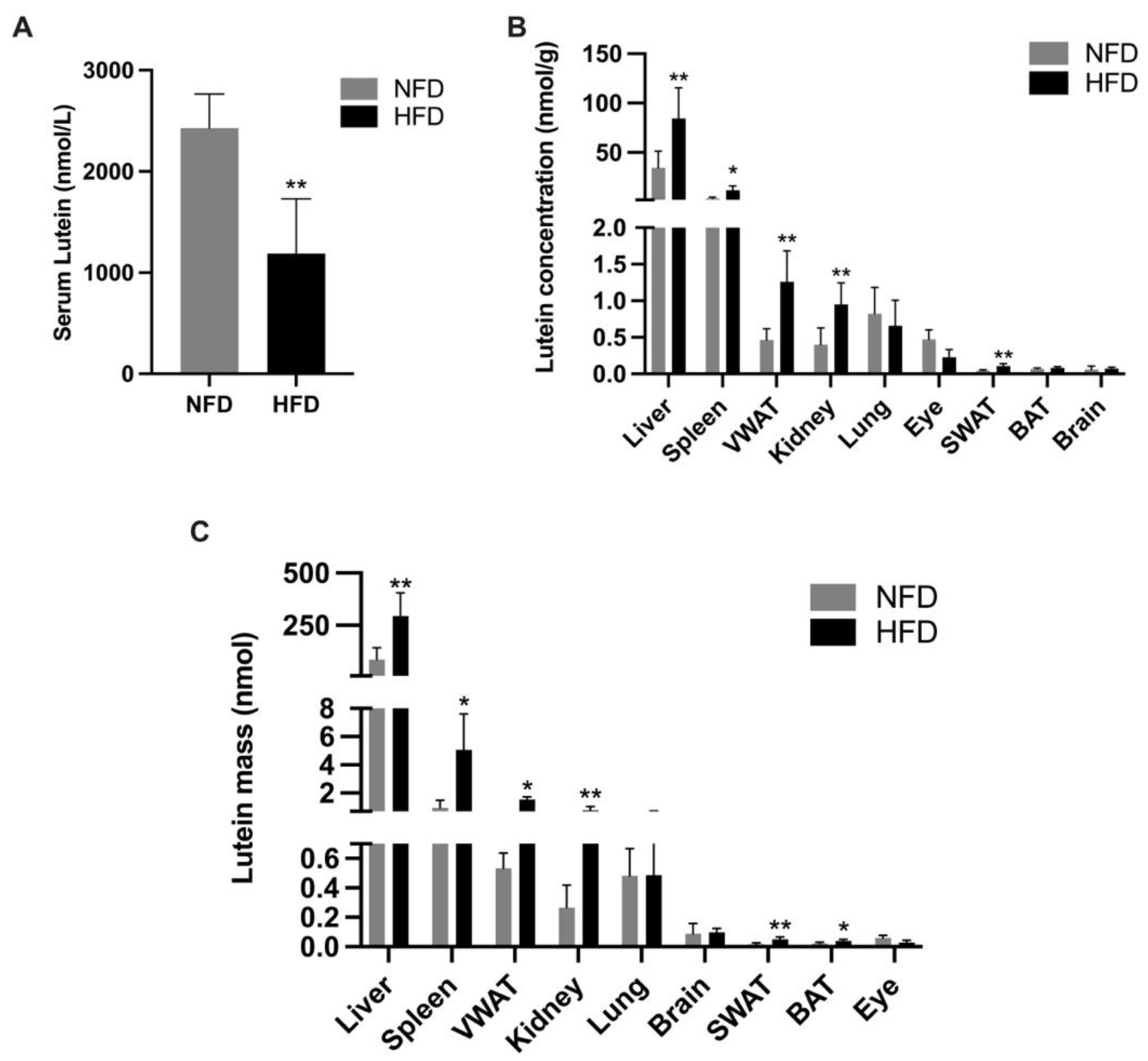
3.2. Milk and Serum Lutein Concentration in Suckling Rats (P2, P6, and P11)
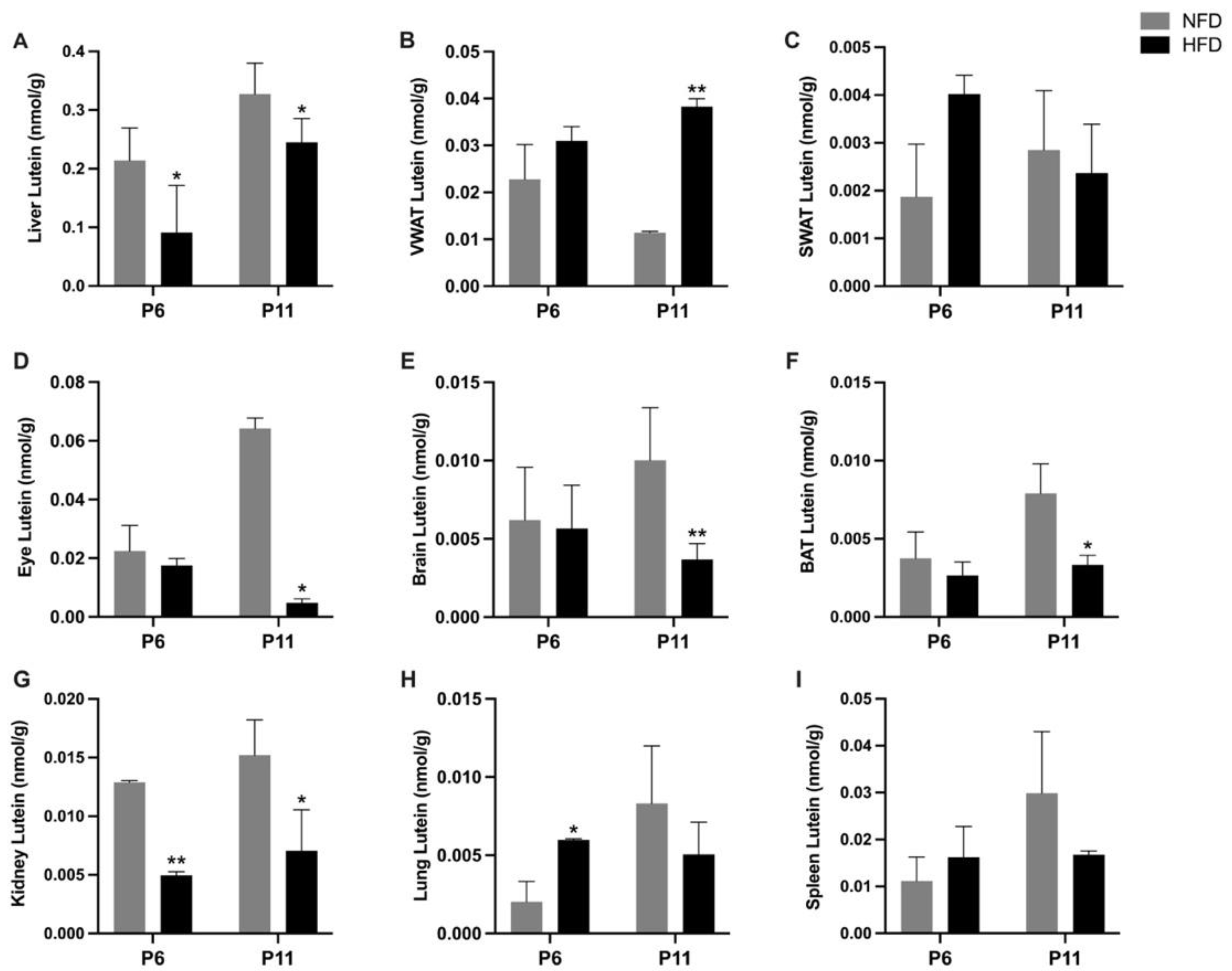
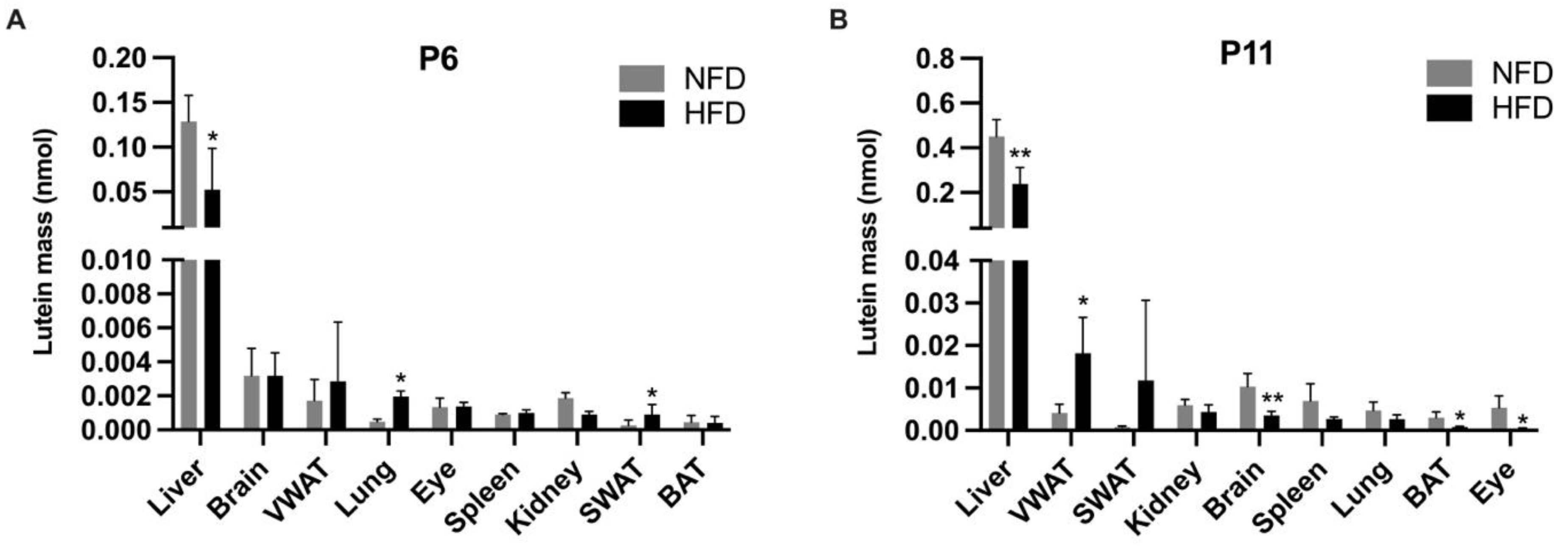
3.3. Tissue Lutein Concentration and Mass in Suckling Rats (P6 and P11)
3.4. Tissue Lutein Concentration and Mass in Weanling Rats (P20)
4. Discussion
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Becerra, M.O.; Contreras, L.M.; Lo, M.H.; Díaz, J.M.; Herrera, G.C. Lutein as a functional food ingredient: Stability and bioavailability. J. Funct. Foods 2020, 66, 10377. [Google Scholar]
- Vishwanathan, R.; Kuchan, M.J.; Sen, S.; Johnson, E.J. Lutein and preterm infants with decreased concentrations of brain carotenoids. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Haber, N.; Heuberger, R. The effect of lutein and zeaxanthin on premature infant eye development. Infant Child Adolesc. Nutr. 2013, 6, 73–85. [Google Scholar] [CrossRef]
- Landrum, J.T.; Bone, R.A. Lutein, zeaxanthin, and the macular pigment. Arch. Biochem. Biophys. 2001, 385, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, B.S.; Chan, G.; Hoffman, R.O.; Sharifzadeh, M.; Ermakov, I.V.; Gellermann, W.; Bernstein, P.S. Interrelationships between maternal carotenoid status and newborn infant macular pigment optical density and carotenoid status. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5568–5578. [Google Scholar] [CrossRef]
- Jeon, S.; Neuringer, M.; Johnson, E.E.; Kuchan, M.J.; Pereira, S.L.; Johnson, E.J.; Erdman, J.W. Effect of carotenoid supplemented formula on carotenoid bioaccumulation in tissues of infant Rhesus Macaques: A pilot study focused on lutein. Nutrients 2017, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Giampietri, M.; Lorenzoni, F.; Moscuzza, F.; Boldrini, A.; Ghirri, P. Lutein and neurodevelopment in preterm infants. Front. Neurosci. 2016, 10, 411. [Google Scholar] [CrossRef] [Green Version]
- Tanprasertsuk, J.; Li, B.; Bernstein, P.S.; Vishwanathan, R.; Johnson, M.A.; Poon, L.; Johnson, E.J. Relationship between concentrations of lutein and StARD3 among pediatric and geriatric human brain tissue. PLoS ONE 2016, 11, e0155488. [Google Scholar]
- Lipkie, T.E.; Morrow, A.L.; Jouni, Z.E.; McMahon, R.J.; Ferruzzi, M.G. Longitudinal survey of carotenoids in human milk from urban cohorts in China, Mexico, and the USA. PLoS ONE 2015, 10, e0127729. [Google Scholar] [CrossRef] [Green Version]
- Perrone, S.; Tei, M.; Longini, M.; Buonocore, G. The multiple facets of lutein: A call for further investigation in the perinatal period. Oxidative Med. Cell. Longev. 2016, 2016, 5381540. [Google Scholar] [CrossRef] [Green Version]
- Zielinska, M.A.; Hamulka, J.; Wesolowska, A. Carotenoid content in breastmilk in the 3rd and 6th month of lactation and its associations with maternal dietary intake and anthropometric characteristics. Nutrients 2019, 11, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Y.; Campos-Giménez, E.; Redeuil, K.M.; Lévèques, A.; Actis-Goretta, L.; Vinyes-Pares, G.; Zhang, Y.; Wang, P.; Thakkar, S.K. Concentrations of carotenoids and tocopherols in breast milk from urban Chinese mothers and their associations with maternal characteristics: A cross-sectional study. Nutrients 2017, 9, 1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albuquerque, M.N.; Diniz Ada, S.; Arruda, I.K. Elevated serum retinol and low beta-carotene but not alpha-tocopherol concentrations are associated with dyslipidemia in Brazilian adolescents. J. Nutr. Sci. Vitaminol. 2016, 62, 73–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrelli, A.; Bucovsky, M.; Horst, R.; Cremers, S.; Zhang, C.; Bessler, M.; Schrope, B.; Evanko, J.; Blanco, J.; Silverberg, S.J.; et al. Vitamin D storage in adipose tissue of obese and normal weight women. J. Bone Miner. Res. 2017, 32, 237–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, W.; Conroy, S.M.; Maskarinec, G.; Franke, A.A.; Pagano, I.S.; Cooney, R.V. Associations between obesity and serum lipid-soluble micronutrients among premenopausal women. Nutr. Res. 2010, 30, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Senkus, K.E.; Tan, L.; Crowe-White, K.M. Systemic and adipose tissue redox status in Sprague-Dawley rats fed normal- and high-fat diets supplemented with lycopene. J. Med. Food 2020, 24, 370–376. [Google Scholar] [CrossRef]
- Han, G.M.; Soliman, G.A.; Meza, J.L.; Islam, K.M.; Watanabe-Galloway, S. The influence of BMI on the association between serum lycopene and the metabolic syndrome. Br. J. Nutr. 2016, 115, 1292–1300. [Google Scholar] [CrossRef] [Green Version]
- Andersen, L.F.; Jacobs, D.R., Jr.; Gross, M.D.; Schreiner, P.J.; Dale Williams, O.; Lee, D.H. Longitudinal associations between body mass index and serum carotenoids: The CARDIA study. Br. J. Nutr. 2006, 95, 358–365. [Google Scholar] [CrossRef] [Green Version]
- Hammond, B.R., Jr.; Ciulla, T.A.; Snodderly, D.M. Macular pigment density is reduced in obese subjects. Investig. Ophthalmol. Vis. Sci. 2002, 43, 47–50. [Google Scholar]
- Kirby, M.L.; Beatty, S.; Stack, J.; Harrison, M.; Greene, I.; McBrinn, S.; Carroll, P.; Nolan, J.M. Changes in macular pigment optical density and serum concentrations of lutein and zeaxanthin in response to weight loss. Br. J. Nutr. 2011, 105, 1036–1046. [Google Scholar] [CrossRef] [Green Version]
- Vahratian, A. Prevalence of overweight and obesity among women of childbearing age: Results from the 2002 National Survey of Family Growth. Matern. Child Health J. 2009, 13, 268. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Xu, X.; Yan, Y. Estimated global overweight and obesity burden in pregnant women based on panel data model. PLoS ONE 2018, 13, e0202183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaillard, R. Maternal obesity during pregnancy and cardiovascular development and disease in the offspring. Eur. J. Epidemiol. 2015, 30, 1141–1152. [Google Scholar] [CrossRef] [Green Version]
- Sheshappa, M.B.; Ranganathan, A.; Bhatiwada, N.; Talahalli, R.R.; Vallikannan, B. Dietary Components Affect the plasma and tissue levels of lutein in aged rats with lutein deficiency—A repeated gavage and dietary study. J. Food Sci. 2015, 80, H2322–H2330. [Google Scholar] [CrossRef]
- Yang, C.; Zhao, A.; Ren, Z.; Zhang, J.; Wang, P.; Zhang, Y. Vitamin A nutritional status of urban lactating Chinese women and its associated factors. Nutrients 2022, 14, 3184. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.S. Absorption, metabolism, and transport of carotenoids. FASEB J. 1996, 10, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Kern, P.A.; Ong, J.M.; Saffari, B.; Carty, J. The effects of weight loss on the activity and expression of adipose-tissue lipoprotein lipase in very obese humans. N. Engl. J. Med. 1990, 322, 1053–1059. [Google Scholar] [CrossRef]
- Wang, H.; Eckel, R.H. Lipoprotein lipase: From gene to obesity. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E271–E288. [Google Scholar] [CrossRef] [Green Version]
- Walker, R.E.; Harvatine, K.J.; Ross, A.C.; Wagner, E.A.; Riddle, S.W.; Gernand, A.D.; Nommsen-Rivers, L.A. Fatty acid transfer from blood to milk is disrupted in mothers with low milk production, obesity, and inflammation. J. Nutr. 2022, 152, 2716–2726. [Google Scholar] [CrossRef] [PubMed]
- Keaney, J.F., Jr.; Larson, M.G.; Vasan, R.S.; Wilson, P.W.; Lipinska, I.; Corey, D.; Massaro, J.M.; Sutherland, P.; Vita, J.A.; Benjamin, E.J. Obesity and systemic oxidative stress: Clinical correlates of oxidative stress in the Framingham Study. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 434–439. [Google Scholar] [CrossRef] [Green Version]
- Moussa, M.; Gouranton, E.; Gleize, B.; Yazidi, C.E.; Niot, I.; Besnard, P.; Borel, P.; Landrier, J.F. CD36 is involved in lycopene and lutein uptake by adipocytes and adipose tissue cultures. Mol. Nutr. Food Res. 2011, 55, 578–584. [Google Scholar] [CrossRef]
- Shyam, R.; Vachali, P.; Gorusupudi, A.; Nelson, K.; Bernstein, P.S. All three human scavenger receptor class B proteins can bind and transport all three macular xanthophyll carotenoids. Arch. Biochem. Biophys. 2017, 634, 21–28. [Google Scholar] [CrossRef]
- Luo, X.; Li, Y.; Yang, P.; Chen, Y.; Wei, L.; Yu, T.; Xia, J.; Ruan, X.Z.; Zhao, L.; Chen, Y. Obesity induces preadipocyte CD36 expression promoting inflammation via the disruption of lysosomal calcium homeostasis and lysosome function. eBioMed 2020, 56, 102797. [Google Scholar] [CrossRef]
- Bohn, T.; Desmarchelier, C.; Dragsted, L.O.; Nielsen, C.S.; Stahl, W.; Rühl, R.; Keijer, J.; Borel, P. Host-related factors explaining interindividual variability of carotenoid bioavailability and tissue concentrations in humans. Mol. Nutr. Food Res. 2017, 61, 1600685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravikrishnan, R.; Rusia, S.; Ilamurugan, G.; Salunkhe, U.; Deshpande, J.; Shankaranarayanan, J.; Shankaranarayana, M.L.; Soni, M.G. Safety assessment of lutein and zeaxanthin (Lutemax 2020): Subchronic toxicity and mutagenicity studies. Food Chem. Toxicol. 2011, 49, 2841–2848. [Google Scholar] [CrossRef]
- Jeon, S.; Ranard, K.M.; Neuringer, M.; Johnson, E.E.; Renner, L.; Kuchan, M.J.; Pereira, S.L.; Johnson, E.J.; Erdman Jr, J.W. Lutein is differentially deposited across brain regions following formula or breast feeding of infant rhesus macaques. J. Nutr. 2018, 148, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhao, X.; Berde, Y.; Low, Y.L.; Kuchan, M.J. Milk and plasma lutein and zeaxanthin concentrations in Chinese breast-feeding mother-infant dyads with healthy maternal fruit and vegetable intake. J. Am. Coll. Nutr. 2019, 38, 179–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Vachali, P.P.; Gorusupudi, A.; Shen, Z.; Sharifzadeh, H.; Besch, B.M.; Nelson, K.; Horvath, M.M.; Frederick, J.M.; Baehr, W. Inactivity of human β, β-carotene-9′, 10′-dioxygenase (BCO2) underlies retinal accumulation of the human macular carotenoid pigment. Proc. Natl. Acad. Sci. USA 2014, 111, 10173–10178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamil, A.; Smith, D.E.; Blumberg, J.B.; Astete, C.; Sabliov, C.; Oliver Chen, C.Y. Bioavailability and biodistribution of nanodelivered lutein. Food Chem. 2016, 192, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Park, S.U. Current results on the potential health benefits of lutein. EXCLI J. 2016, 15, 308–314. [Google Scholar]
- Koushan, K.; Rusovici, R.; Li, W.; Ferguson, L.R.; Chalam, K.V. The role of lutein in eye-related disease. Nutrients 2013, 5, 1823–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Fat Diet a | Normal Fat Diet + Lutein b | High-Fat Diet c | High-Fat Diet + Lutein d | |||||
|---|---|---|---|---|---|---|---|---|
| g | kcal | g | kcal | g | kcal | g | kcal | |
| Ingredient | ||||||||
| Lutein (Xangold, 30%) | 0 | 0 | 9.8 | 88 | 0 | 0 | 9.8 | 88 |
| Casein | 200 | 800 | 200 | 800 | 200 | 800 | 200 | 800 |
| L-Cystine | 3 | 12 | 3 | 12 | 3 | 12 | 3 | 12 |
| Corn Starch | 353.8 | 1415 | 353.8 | 1415 | 101.2 | 405 | 101.2 | 405 |
| Maltodextrin 10 | 125 | 500 | 125 | 500 | 125 | 500 | 125 | 500 |
| Sucrose | 68.8 | 275 | 68.8 | 275 | 68.8 | 275 | 68.8 | 275 |
| Cellulose, BW200 | 50 | 0 | 50 | 0 | 50 | 0 | 50 | 0 |
| Soybean Oil | 25 | 225 | 25 | 225 | 25 | 225 | 25 | 225 |
| Lard | 87.7 | 789 | 87.7 | 789 | 200 | 1800 | 200 | 1800 |
| Mineral Mix S10026 | 10 | 0 | 10 | 0 | 10 | 0 | 10 | 0 |
| DiCalcium Phosphate | 13 | 0 | 13 | 0 | 13 | 0 | 13 | 0 |
| Calcium Carbonate | 5.5 | 0 | 5.5 | 0 | 5.5 | 0 | 5.5 | 0 |
| Potassium Citrate, 1 H2O | 16.5 | 0 | 16.5 | 0 | 16.5 | 0 | 16.5 | 0 |
| Vitamin Mix V10001 | 10 | 40 | 10 | 40 | 10 | 40 | 10 | 40 |
| Choline Bitartrate | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 |
| Food color | 0.05 | 0 | 0.05 | 0 | 0.05 | 0 | 0.05 | 0 |
| Total | 970.35 | 4057 | 980.15 | 4145 | 830.05 | 4057 | 839.85 | 4145 |
| g/kg | Kcal% | g/kg | Kcal% | g/kg | Kcal% | g/kg | Kcal% | |
| Fat | 120 | 25 | 110 | 27 | 270 | 50 | 270 | 51 |
| Carbohydrate | 570 | 30 | 570 | 30 | 370 | 29 | 360 | 29 |
| Protein | 210 | 20 | 210 | 20 | 240 | 20 | 240 | 20 |
| Total | - | 100 | - | 100 | - | 100 | - | 100 |
| kcal/g | 4.2 | - | 4.2 | - | 4.9 | - | 4.9 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Tan, L. Maternal High-Fat Diet Consumption in Sprague Dawley Rats Compromised the Availability and Altered the Tissue Distribution of Lutein in Neonatal Offspring. Metabolites 2023, 13, 544. https://doi.org/10.3390/metabo13040544
Zhang Y, Tan L. Maternal High-Fat Diet Consumption in Sprague Dawley Rats Compromised the Availability and Altered the Tissue Distribution of Lutein in Neonatal Offspring. Metabolites. 2023; 13(4):544. https://doi.org/10.3390/metabo13040544
Chicago/Turabian StyleZhang, Yanqi, and Libo Tan. 2023. "Maternal High-Fat Diet Consumption in Sprague Dawley Rats Compromised the Availability and Altered the Tissue Distribution of Lutein in Neonatal Offspring" Metabolites 13, no. 4: 544. https://doi.org/10.3390/metabo13040544
APA StyleZhang, Y., & Tan, L. (2023). Maternal High-Fat Diet Consumption in Sprague Dawley Rats Compromised the Availability and Altered the Tissue Distribution of Lutein in Neonatal Offspring. Metabolites, 13(4), 544. https://doi.org/10.3390/metabo13040544