
Comparative Metabolomics Profiling Reveals Key Metabolites and Associated Pathways Regulating Tuber Dormancy in White Yam (Dioscorea rotundata Poir.)

 , ,
, ,
Abstract
:1. Introduction
2. Experimental Design
2.1. Genetic Material
2.2. Field Study Area
2.3. Planting and Agronomic Management
2.4. Postharvest Study and Sampling
2.5. Metabolites Extraction
Sample Processing
2.6. GC-MS Analysis
Data Processing
2.7. Data Statistical Analysis and Visualization
3. Results
3.1. Metabolic Profiling of White Yam Tuber during Dormancy
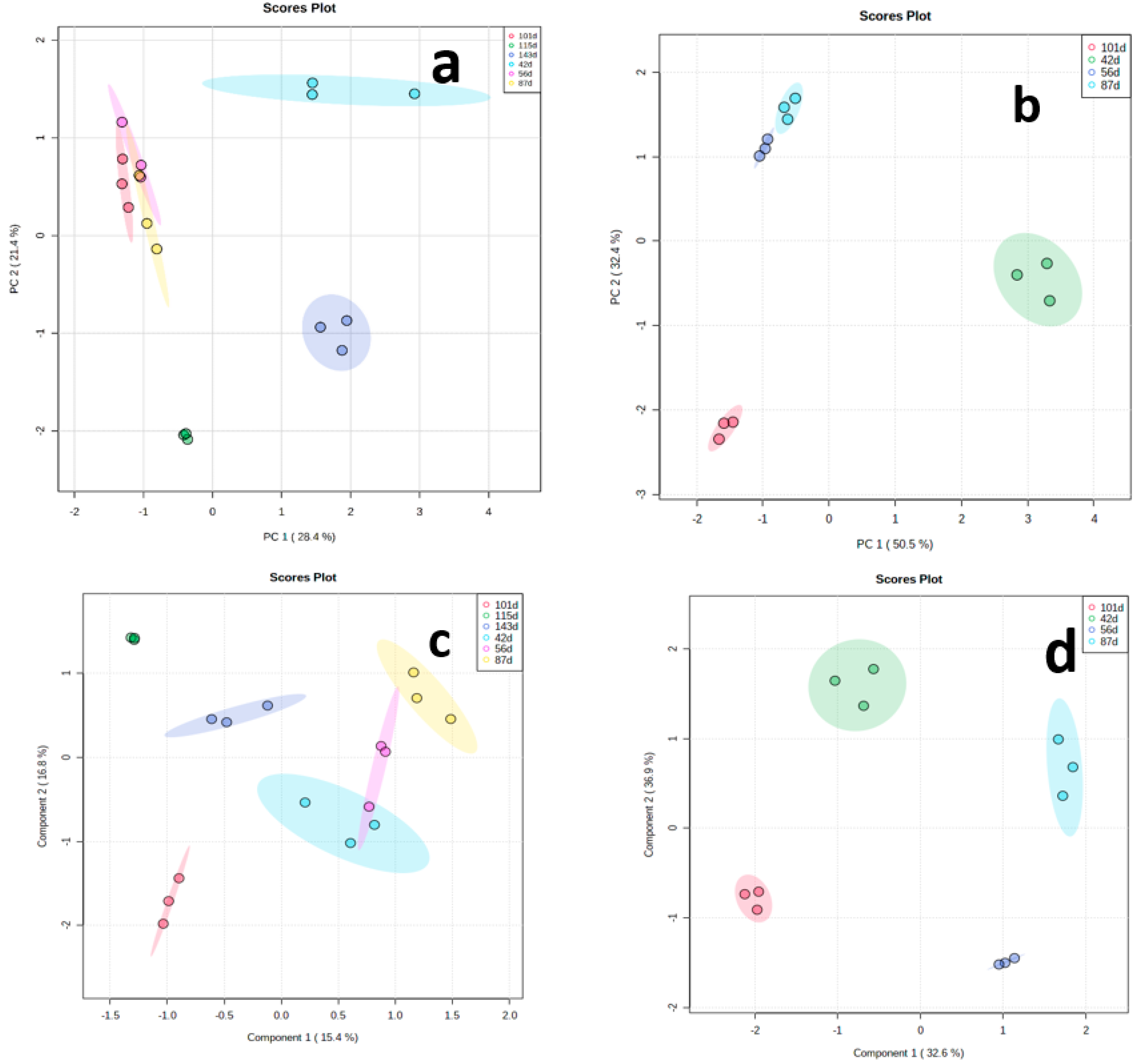
3.2. Principal Component Analysis (PCA) and Partial Least Square–Discriminant Analysis (PLS–DA) of Metabolites
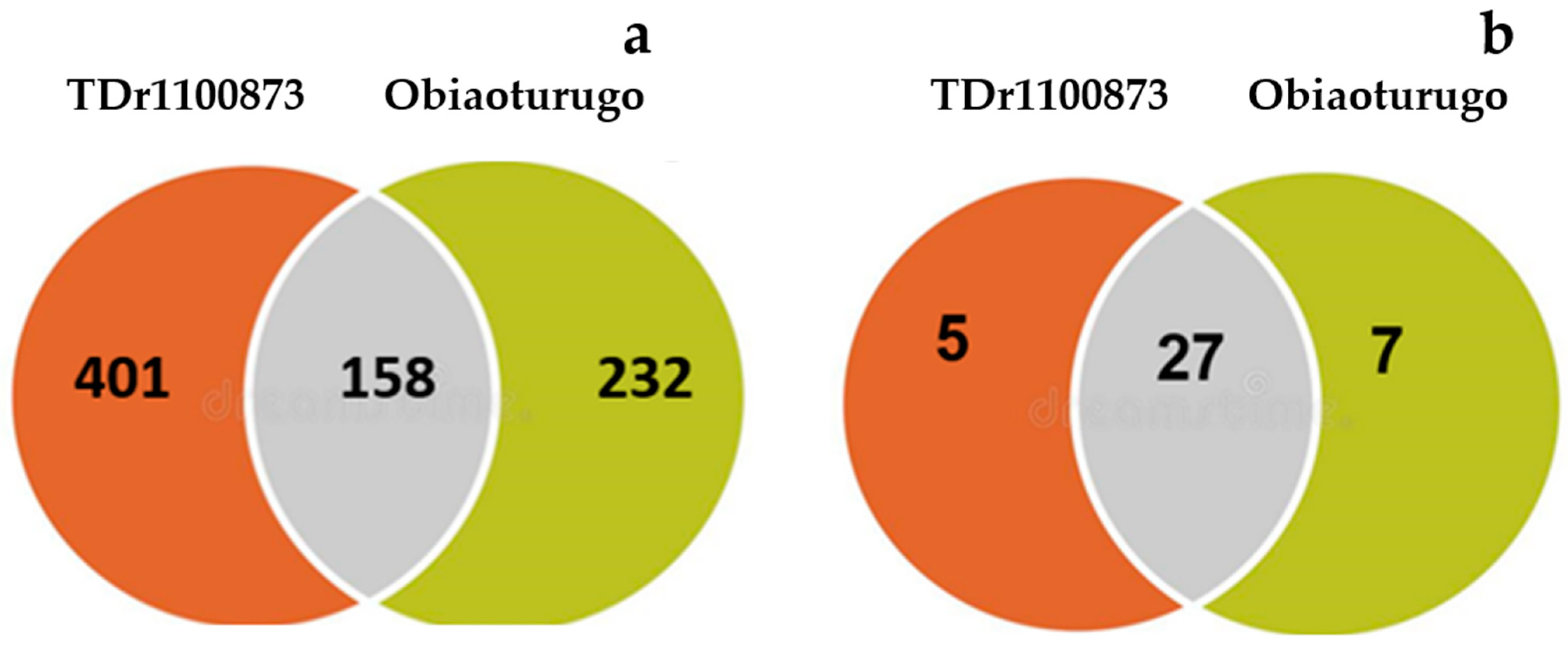
3.3. Identification of Differentially Accumulated Metabolites across the Studied Tuber Dormancy Stages
3.4. Metabolites Mapping and Chemical Functional Groups Identification
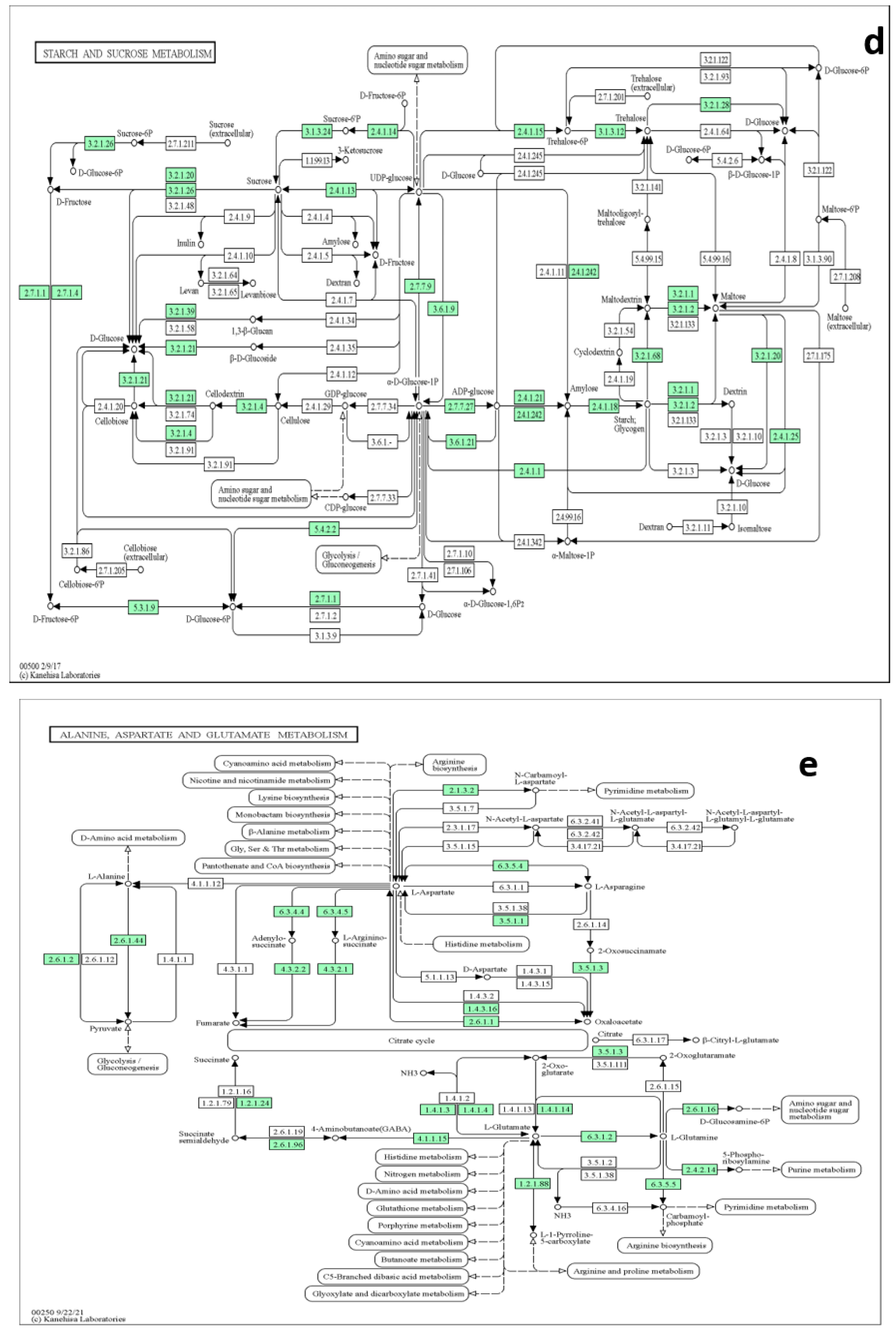
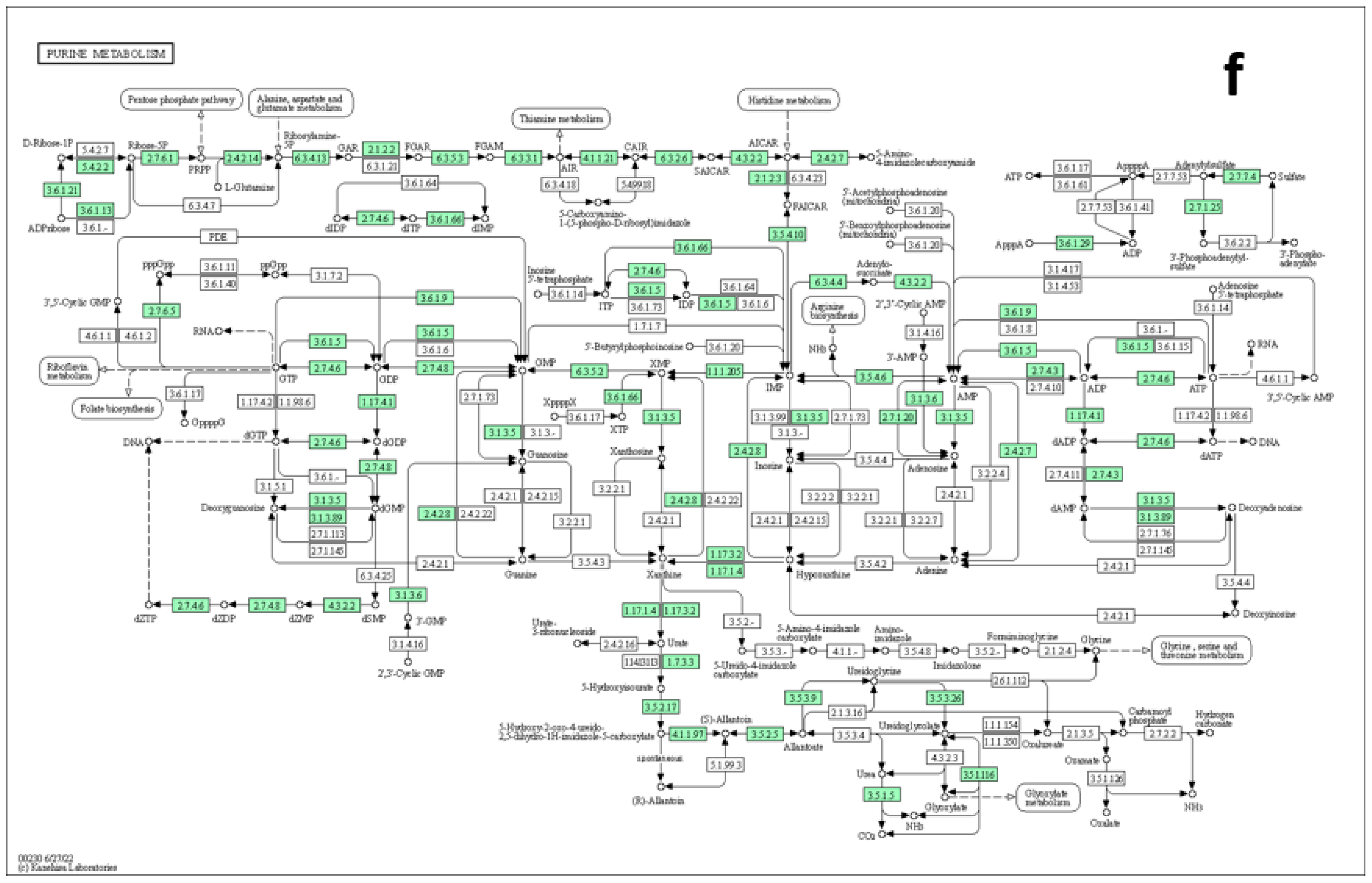
3.5. Metabolites Sets Enrichment Analysis and Pathways Annotation
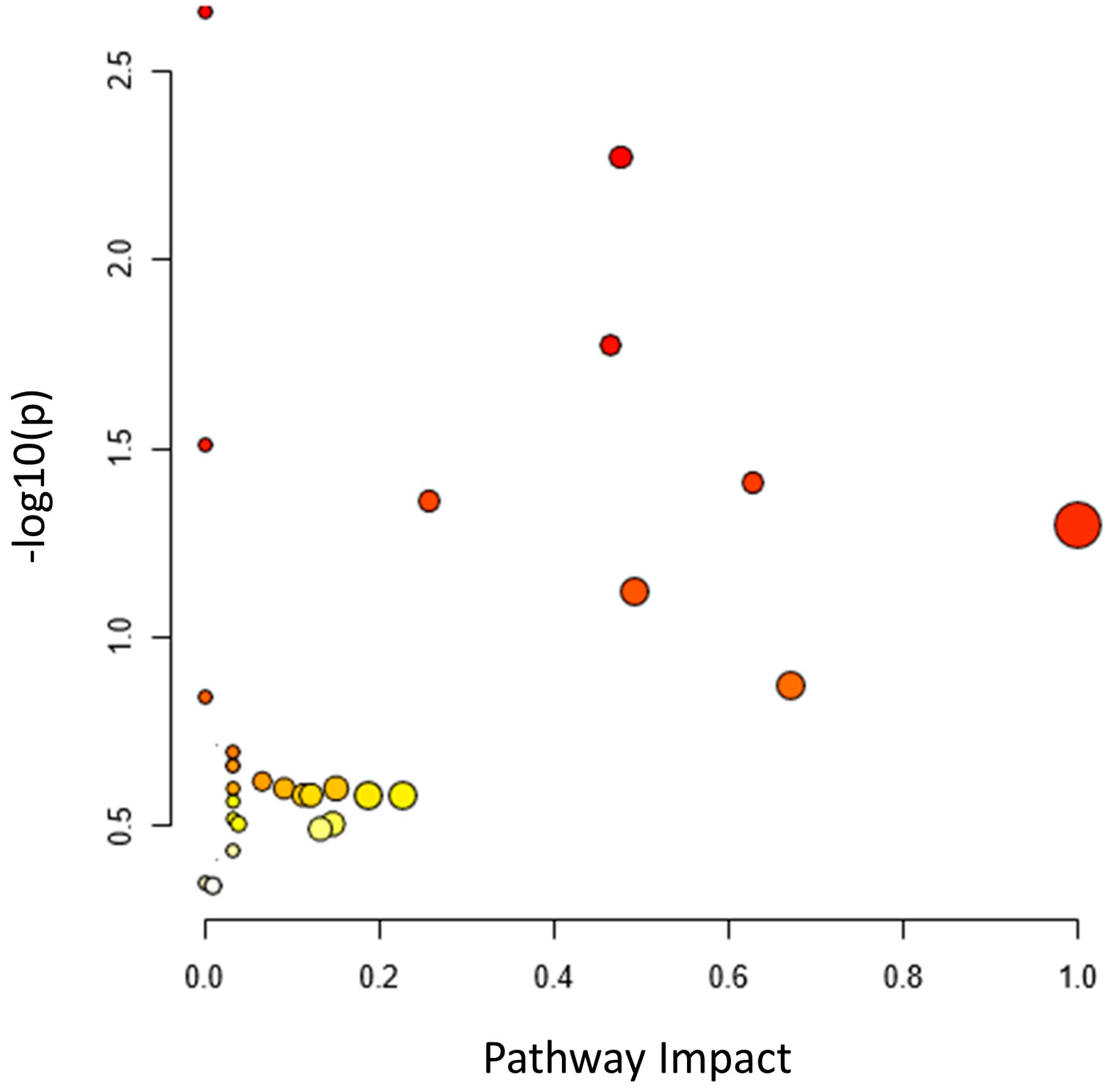
3.6. Pathway Topology Analysis
4. Discussions
4.1. Efficiency of Metabolomics in Deciphering Molecular Mechanisms in a Biological System
4.2. Differentially Accumulated Metabolites Determined the Phenotypic Variation in Dormancy Duration of the Two White Yam Genotypes
4.3. Metabolic Regulation of Yam Tuber Dormancy
4.4. Amines and Amino Acids Metabolisms
4.5. Energy Metabolism
4.6. Nucleotides Metabolism and Cellular Processes
4.7. Secondary Metabolites and Phytohormones Metabolism
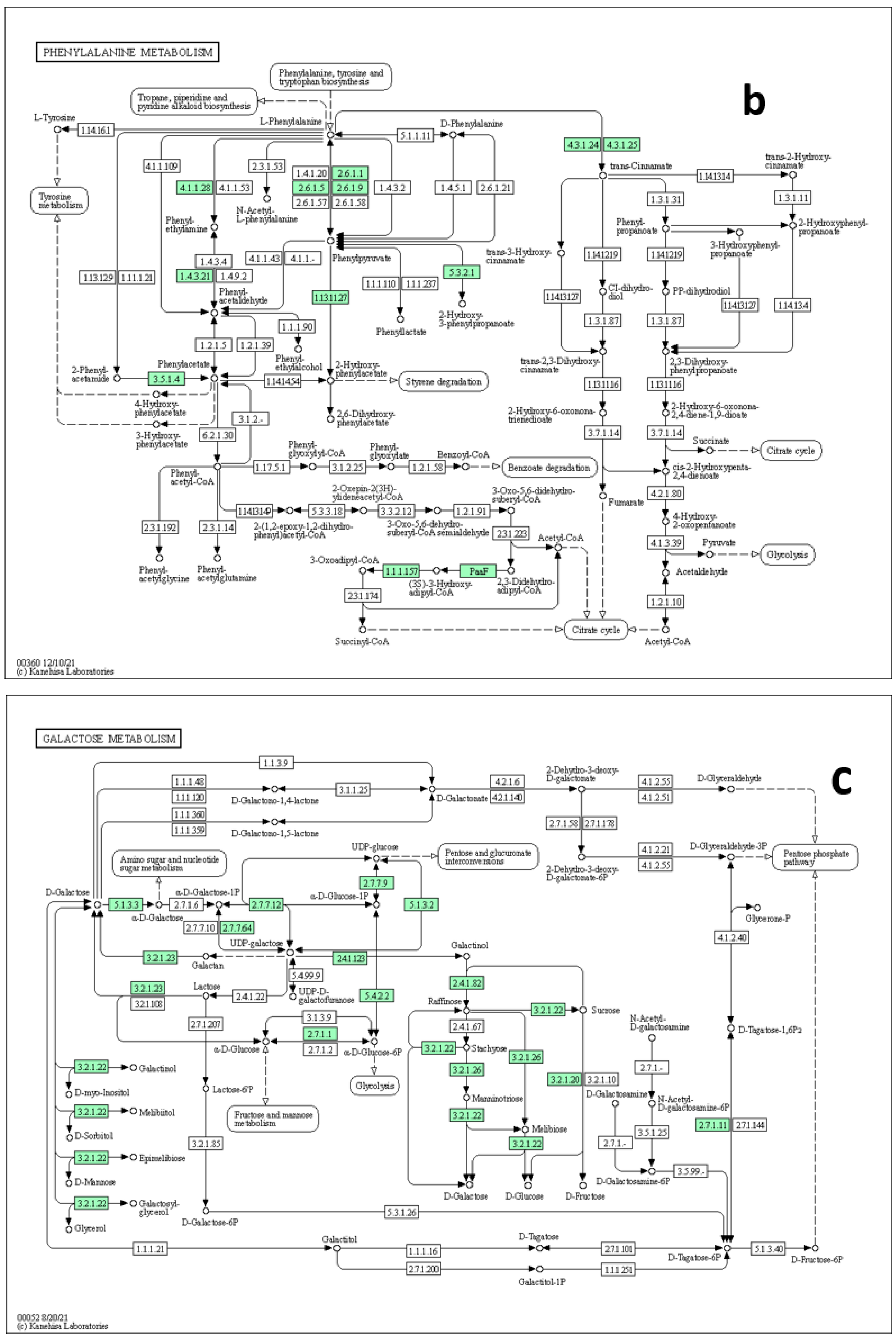
4.8. Pathways Elucidations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Price, E.J.; Wilkin, P.; Sarasan, V.; Fraser, P.D. Metabolite profiling of Dioscorea (yam) species reveals underutilised biodiversity and renewable sources for high-value compounds. Sci. Rep. 2016, 6, 29136. [Google Scholar] [CrossRef]
- Obidiegwu, J.E.; Lyons, J.B.; Chilaka, C.A. The Dioscorea Genus (Yam)—An appraisal of nutritional and therapeutic potentials. Foods 2020, 9, 1304. [Google Scholar] [CrossRef]
- Mignouna, H.D.; Abang, M.M.; Asiedu, R. Harnessing modern biotechnology for tropical tuber crop improvement: Yam (Dioscorea spp.) molecular breeding. Afr. J. Biotechnol. 2003, 2, 478–485. [Google Scholar]
- Lev, L.S.; Shriver, A.L. A trend analysis of yam production, area, yield, and trade (1961–1996). Colloq.-CIRAD 1998, 11–20. [Google Scholar]
- Dansi, A.; Dantsey-Barry, H.; Dossou-Aminon, I.; N’kpenu, E.; Agré, A.; Sunu, Y.; Kombaté, K.; Loko, Y.; Dansi, M.; Assogba, P. Varietal diversity and genetic erosion of cultivated yams (Dioscorea cayenensis Poir-D. rotundata Lam complex and D. alata L.) in Togo. Int. J. Biodivers. Conserv. 2013, 5, 223–239. [Google Scholar]
- FAOSTAT. Food and Agriculture Organization of the United Nations. 2020. Available online: http://wwwfaoorg/faostat/en/#data/QC (accessed on 23 October 2022).
- Nwogha, J.S.; Wosene, A.G.; Raveendran, M.; Oselebe, H.O.; Obidiegwu, J.E.; Amirtham, D. Physiological and Molecular basis of dormancy in yam tuber: A way forward towards genetic manipultion of dormancy in yam tubers. Glob. J. Sci. Front. Res. 2022, XXll, 47–73. [Google Scholar]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo-, para-, and ecodormancy: Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees–from what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar]
- Mounirou, Y.; Jeanne, Z.; Djakaridia, T.; Sèssi Ida, A.; Elie Idossou, A.; Hibath Audrey Agbékè Mahougnon, A.; Jean Didier, Z.; Akpovi, A. Evaluation of yam (Dioscorea cayenensis–Dioscorea rotundata) seed germination grown in Centre Benin. Int. J. 2015, 3, 277–284. [Google Scholar]
- Ile, E.; Craufurd, P.; Battey, N.; Asiedu, R. Phases of dormancy in yam tubers (Dioscorea rotundata). Ann. Bot. 2006, 97, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Passam, H. Dormancy of yams in relation to storage. In Yams. Ignames; Oxford University Press: Oxford, UK, 1982; pp. 285–293. [Google Scholar]
- Hamadina, E.I.; Craufurd, P.Q. Changes in Free Phenolics Contents during Tuber Development, Dormancy and Sprouting in White Yam (Dioscorea rotundata Poir.). Int. J. Plant Res. 2015, 5, 34–41. [Google Scholar]
- Shangguan, L.; Chen, M.; Fang, X.; Xie, Z.; Gong, P.; Huang, Y.; Wang, Z.; Fang, J. Comparative transcriptome analysis provides insight into regulation pathways and temporal and spatial expression characteristics of grapevine (Vitis vinifera) dormant buds in different nodes. BMC Plant Biol. 2020, 20, 390. [Google Scholar] [CrossRef]
- Kaczmarska, K.T.; Chandra-Hioe, M.V.; Zabaras, D.; Frank, D.; Arcot, J. Effect of germination and fermentation on carbohydrate composition of Australian sweet lupin and soybean seeds and flours. J. Agric. Food Chem. 2017, 65, 10064–10073. [Google Scholar] [CrossRef]
- González-Orenga, S.; Ferrer-Gallego, P.P.; Laguna, E.; López-Gresa, M.P.; Donat-Torres, M.P.; Verdeguer, M.; Vicente, O.; Boscaiu, M. Insights on salt tolerance of two endemic Limonium species from Spain. Metabolites 2019, 9, 294. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Liu, X.-d.; Xie, Q.; He, Z.-h. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant 2016, 9, 34–45. [Google Scholar]
- Guo, H.; Lyv, Y.; Zheng, W.; Yang, C.; Li, Y.; Wang, X.; Chen, R.; Wang, C.; Luo, J.; Qu, L. Comparative metabolomics reveals two metabolic modules affecting seed germination in rice (Oryza sativa). Metabolites 2021, 11, 880. [Google Scholar] [CrossRef] [PubMed]
- Shuai, H.; Meng, Y.; Luo, X.; Chen, F.; Zhou, W.; Dai, Y.; Qi, Y.; Du, J.; Yang, F.; Liu, J. Exogenous auxin represses soybean seed germination through decreasing the gibberellin/abscisic acid (GA/ABA) ratio. Sci. Rep. 2017, 7, 12620. [Google Scholar] [CrossRef] [PubMed]
- Kralj Cigić, I.; Rupnik, S.; Rijavec, T.; Poklar Ulrih, N.; Cigić, B. Accumulation of agmatine, spermidine, and spermine in sprouts and microgreens of alfalfa, fenugreek, lentil, and daikon radish. Foods 2020, 9, 547. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, G.; de Jiménez, E.S. Polyamine distribution among maize embryonic tissues and its relation to seed germination. Biochem. Biophys. Res. Commun. 1988, 153, 881–887. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.; Rehman, H.; Hussain, M. Seed priming with polyamines improves the germination and early seedling growth in fine rice. J. New Seeds 2008, 9, 145–155. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Y.; Zhang, X.-Q.; Ma, X.; Huang, L.-K.; Yan, Y.-H. Exogenous spermidine improves seed germination of white clover under water stress via involvement in starch metabolism, antioxidant defenses and relevant gene expression. Molecules 2014, 19, 18003–18024. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lin, C.; He, F.; Li, Z.; Guan, Y.; Hu, Q.; Hu, J. Exogenous spermidine improves seed germination of sweet corn via involvement in phytohormone interactions, H2O2 and relevant gene expression. BMC Plant Biol. 2017, 17, 1. [Google Scholar] [CrossRef] [PubMed]
- Sibian, M.S.; Saxena, D.C.; Riar, C.S. Effect of germination on chemical, functional and nutritional characteristics of wheat, brown rice and triticale: A comparative study. J. Sci. Food Agric. 2017, 97, 4643–4651. [Google Scholar] [CrossRef]
- Gipson, A.B.; Morton, K.J.; Rhee, R.J.; Simo, S.; Clayton, J.A.; Perrett, M.E.; Binkley, C.G.; Jensen, E.L.; Oakes, D.L.; Rouhier, M.F. Disruptions in valine degradation affect seed development and germination in Arabidopsis. Plant J. 2017, 90, 1029–1039. [Google Scholar] [CrossRef]
- Desmaison, A.M.; Tixier, M. Amino acids content in germinating seeds and seedlings from Castanea sativa L. Plant Physiol. 1986, 81, 692–695. [Google Scholar] [CrossRef]
- Zhang, T.; Yuan, Y.; Zhan, Y.; Cao, X.; Liu, C.; Zhang, Y.; Gai, S. Metabolomics analysis reveals Embden Meyerhof Parnas pathway activation and flavonoids accumulation during dormancy transition in tree peony. BMC Plant Biol. 2020, 20, 484. [Google Scholar] [CrossRef]
- Wang, S.Y.; Jiao, H.J.; Faust, M. Changes in metabolic enzyme activities during thidiazuron-induced lateral budbreak of apple. HortScience 1991, 26, 171–173. [Google Scholar] [CrossRef]
- Halaly, T.; Pang, X.; Batikoff, T.; Crane, O.; Keren, A.; Venkateswari, J.; Ogrodovitch, A.; Sadka, A.; Lavee, S.; Or, E. Similar mechanisms might be triggered by alternative external stimuli that induce dormancy release in grape buds. Planta 2008, 228, 79–88. [Google Scholar] [CrossRef]
- Shen, C.; Wang, H.; Wang, X.; Wang, B.; Zheng, X.; Shi, X.; Liu, W.; Liu, F. Respiratory changes during dormancy of grape buds. Sci. Agric. Sin. 2013, 46, 1201–1207. [Google Scholar]
- Deng, S.; Xiao, Q.; Xu, C.; Hong, J.; Deng, Z.; Jiang, D.; Luo, S. Metabolome profiling of stratified seeds provides insight into the regulation of dormancy in Davidia involucrata. Plant Divers. 2022, 44, 417–427. [Google Scholar] [CrossRef]
- Arc, E.; Chibani, K.; Grappin, P.; Jullien, M.; Godin, B.; Cueff, G.; Valot, B.; Balliau, T.; Job, D.; Rajjou, L. Cold stratification and exogenous nitrates entail similar functional proteome adjustments during Arabidopsis seed dormancy release. J. Proteome Res. 2012, 11, 5418–5432. [Google Scholar] [CrossRef]
- Szczotka, Z.; Pawłowski, T.; Krawiarz, K. Proteins and polyamines during dormancy breaking of European beech (Fagus sylvatica L.) seeds. Acta Physiol. Plant. 2003, 25, 423–435. [Google Scholar] [CrossRef]
- Silva, A.C.D.; Suassuna, J.F.; Melo, A.S.D.; Costa, R.R.; Andrade, W.L.D.; Silva, D.C.D. Salicylic acid as attenuator of drought stress on germination and initial development of sesame. Rev. Bras. Eng. Agrícola Ambient. 2017, 21, 156–162. [Google Scholar] [CrossRef]
- De Souza Vidigal, D.; Willems, L.; van Arkel, J.; Dekkers, B.J.; Hilhorst, H.W.; Bentsink, L. Galactinol as marker for seed longevity. Plant Sci. 2016, 246, 112–118. [Google Scholar] [CrossRef]
- Baena-González, E.; Lunn, J.E. SnRK1 and trehalose 6-phosphate–two ancient pathways converge to regulate plant metabolism and growth. Curr. Opin. Plant Biol. 2020, 55, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhu, J.-Y.; Roh, J.; Marchive, C.; Kim, S.-K.; Meyer, C.; Sun, Y.; Wang, W.; Wang, Z.-Y. TOR signaling promotes accumulation of BZR1 to balance growth with carbon availability in Arabidopsis. Curr. Biol. 2016, 26, 1854–1860. [Google Scholar] [CrossRef]
- Bledsoe, S.W.; Henry, C.; Griffiths, C.A.; Paul, M.J.; Feil, R.; Lunn, J.E.; Stitt, M.; Lagrimini, L.M. The role of Tre6P and SnRK1 in maize early kernel development and events leading to stress-induced kernel abortion. BMC Plant Biol. 2017, 17, 74. [Google Scholar] [CrossRef]
- Figueroa, C.M.; Lunn, J.E. A tale of two sugars: Trehalose 6-phosphate and sucrose. Plant Physiol. 2016, 172, 7–27. [Google Scholar] [CrossRef]
- Tiama, D.; Sawadogo, N.; Traore, R.E.; Yolou, M.; Bationo-Kando, P.; Zoundjihekpon, J.; Sawadogo MZongo, J.D. Effect of chemical fertilizers on production of yams (nyù) of passore in farmers’ environment. Agron. Afr. 2018, 30, 99–105. [Google Scholar]
- Enfissi, E.M.; Barneche, F.; Ahmed, I.; Lichtlé, C.; Gerrish, C.; McQuinn, R.P.; Giovannoni, J.J.; Lopez-Juez, E.; Bowler, C.; Bramley, P.M. Integrative transcript and metabolite analysis of nutritionally enhanced DE-ETIOLATED1 downregulated tomato fruit. Plant Cell 2010, 22, 1190–1215. [Google Scholar] [CrossRef]
- Oliveros, J.C.; VENNY. An Interactive Tool for Comparing Lists with Venn Diagrams. 2007. Available online: http://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 1 November 2022).
- Nwogha, J.S.; Abtew, W.G.; Raveendran, M.; Oselebe, H.O.; Obidiegwu, J.E.; Chilaka, C.A.; Amirtham, D.D. Role of Non-Structural Sugar Metabolism in Regulating Tuber Dormancy in White Yam (Dioscorea rotundata). Agriculture 2023, 13, 343. [Google Scholar] [CrossRef]
- Han, C.; Zhen, S.; Zhu, G.; Bian, Y.; Yan, Y. Comparative metabolome analysis of wheat embryo and endosperm reveals the dynamic changes of metabolites during seed germination. Plant Physiol. Biochem. 2017, 115, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, M.; Dong, S.; Liu, Y.-L.; Li, Z.-H. The Opposite Roles of White Light in Regulating Germination of Fresh and Aged Seed in Tobacco. Plants 2021, 10, 2457. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yang, J.; Su, L.; Sun, K.; Li, D.; Liu, Y.; Wang, H.; Chen, Z.; Guo, T. Metabolic profile analysis and identification of key metabolites during rice seed germination under low-temperature stress. Plant Sci. 2019, 289, 110282. [Google Scholar] [CrossRef]
- Mazlan, O.; Aizat, W.M.; Baharum, S.N.; Azizan, K.A.; Noor, N.M. Metabolomics analysis of developing Garcinia mangostana seed reveals modulated levels of sugars, organic acids and phenylpropanoid compounds. Sci. Hortic. 2018, 233, 323–330. [Google Scholar] [CrossRef]
- Yan, S.; Huang, W.; Gao, J.; Fu, H.; Liu, J. Comparative metabolomic analysis of seed metabolites associated with seed storability in rice (Oryza sativa L.) during natural aging. Plant Physiol. Biochem. 2018, 127, 590–598. [Google Scholar] [CrossRef]
- Feenstra, A.D.; Alexander, L.E.; Song, Z.; Korte, A.R.; Yandeau-Nelson, M.D.; Nikolau, B.J.; Lee, Y.J. Spatial mapping and profiling of metabolite distributions during germination. Plant Physiol. 2017, 174, 2532–2548. [Google Scholar] [CrossRef]
- Yang, L.; Liu, S.; Lin, R. The role of light in regulating seed dormancy and germination. J. Integr. Plant Biol. 2020, 62, 1310–1326. [Google Scholar] [CrossRef]
- Horikoshi, H.M.; Sekozawa, Y.; Kobayashi, M.; Saito, K.; Kusano, M.; Sugaya, S. Metabolomics analysis of ‘Housui’ Japanese pear flower buds during endodormancy reveals metabolic suppression by thermal fluctuation. Plant Physiol. Biochem. 2018, 126, 134–141. [Google Scholar] [CrossRef]
- Amarowicz, R.; Weidner, S. Biological activity of grapevine phenolic compounds. In Grapevine Molecular Physiology & Biotechnology; Springer: Dordrecht, The Netherlands, 2009; pp. 389–405. [Google Scholar]
- Wang, L.; Ruan, Y.-L. Regulation of cell division and expansion by sugar and auxin signaling. Front. Plant Sci. 2013, 4, 163. [Google Scholar] [CrossRef]
- Hartig, K.; Beck, E. Crosstalk between auxin, cytokinins, and sugars in the plant cell cycle. Plant Biol. 2006, 8, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.G.; Lea, P.J. Glutamate in plants: Metabolism, regulation, and signalling. J. Exp. Bot. 2007, 58, 2339–2358. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-Y.; Chen, B.-X.; Chen, Z.-J.; Gao, Y.-T.; Chen, Z.; Liu, J. Reactive oxygen species generated by NADPH oxidases promote radicle protrusion and root elongation during rice seed germination. Int. J. Mol. Sci. 2017, 18, 110. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Cai, S.; Ye, L.; Hu, H.; Li, C.; Zhang, G. The effects of GA and ABA treatments on metabolite profile of germinating barley. Food Chem. 2016, 192, 928–933. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zeng, L.; Zhao, H.; Ye, Q. Proteomic analysis of the early development of the Phalaenopsis amabilis flower bud under low temperature induction using the iTRAQ/MRM approach. Molecules 2020, 25, 1244. [Google Scholar] [CrossRef] [PubMed]
- Tiburcio, A.F.; Alcazar, R. Potential applications of polyamines in agriculture and plant biotechnology. In Polyamines: Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; pp. 489–508. [Google Scholar]
- Ma, Z.; Bykova, N.V.; Igamberdiev, A.U. Cell signaling mechanisms and metabolic regulation of germination and dormancy in barley seeds. Crop J. 2017, 5, 459–477. [Google Scholar]
- Hildebrandt, T.M.; Nesi, A.N.; Araújo, W.L.; Braun, H.-P. Amino acid catabolism in plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar]
- Yang, Q.; Zhao, D.; Liu, Q. Connections between amino acid metabolisms in plants: Lysine as an example. Front. Plant Sci. 2020, 11, 928. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of exogenous proline to abiotic stresses tolerance in plants: A review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Fujita, Y.; Fujita, M.; Satoh, R.; Maruyama, K.; Parvez, M.M.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis. Plant Cell 2005, 17, 3470–3488. [Google Scholar] [CrossRef] [PubMed]
- Yadav, U.P.; Ivakov, A.; Feil, R.; Duan, G.Y.; Walther, D.; Giavalisco, P.; Piques, M.; Carillo, P.; Hubberten, H.-M.; Stitt, M. The sucrose–trehalose 6-phosphate (Tre6P) nexus: Specificity and mechanisms of sucrose signalling by Tre6P. J. Exp. Bot. 2014, 65, 1051–1068. [Google Scholar] [CrossRef] [PubMed]
- Muller, K.; Linkies, A.; Vreeburg, R.A.; Fry, S.C.; Krieger-Liszkay, A.; Leubner-Metzger, G. In vivo cell wall loosening by hydroxyl radicals during cress seed germination and elongation growth. Plant Physiol. 2009, 150, 1855–1865. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Calle, V.; Barrero-Sicilia, C.; Carbonero, P.; Iglesias-Fernandez, R.R. Mannans and endo-β-mannanases (MAN) in Brachypodium distachyon: Expression profiling and possible role of the BdMAN genes during coleorhiza-limited seed germination. J. Exp. Bot. 2015, 66, 3753–3764. [Google Scholar] [CrossRef]
- Falco, S.; Guida, T.; Locke, M.; Mauvais, J.; Sanders, C.; Ward, R.; Webber, P. Transgenic canola and soybean seeds with increased lysine. Bio/technology 1995, 13, 577–582. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, C.; Wang, X.; Liu, Q.; Yuan, D.; Pan, G.; Sun, S.S.; Tu, J. Development of high-lysine rice via endosperm-specific expression of a foreign LYSINE RICH PROTEIN gene. BMC Plant Biol. 2016, 16, 147. [Google Scholar] [CrossRef]
- Jia, M.; Wu, H.; Clay, K.L.; Jung, R.; Larkins, B.A.; Gibbon, B.C. Identification and characterization of lysine-rich proteins and starch biosynthesis genes in the opaque2mutant by transcriptional and proteomic analysis. BMC Plant Biol. 2013, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Li, X.; Wang, X.; Chen, H.; Chen, F.; Shen, S. Proteomic analysis of rice (Oryza sativa) seeds during germination. Proteomics 2007, 7, 3358–3368. [Google Scholar] [PubMed]
- Nakabayashi, K.; Okamoto, M.; Koshiba, T.; Kamiya, Y.; Nambara, E. Genome-wide profiling of stored mRNA in Arabidopsis thaliana seed germination: Epigenetic and genetic regulation of transcription in seed. Plant J. 2005, 41, 697–709. [Google Scholar] [CrossRef]
- Gianinetti, A.; Finocchiaro, F.; Bagnaresi, P.; Zechini, A.; Faccioli, P.; Cattivelli, L.; Valè, G.; Biselli, C. Seed dormancy involves a transcriptional program that supports early plastid functionality during imbibition. Plants 2018, 7, 35. [Google Scholar] [CrossRef]
- Guo, G.; Liu, X.; Sun, F.; Cao, J.; Huo, N.; Wuda, B.; Xin, M.; Hu, Z.; Du, J.; Xia, R. Wheat miR9678 affects seed germination by generating phased siRNAs and modulating abscisic acid/gibberellin signaling. Plant Cell 2018, 30, 796–814. [Google Scholar] [CrossRef]
- Liu, J.-H.; Honda, C.; Moriguchi, T. Involvement of polyamine in floral and fruit development. Jpn. Agric. Res. Q. JARQ 2006, 40, 51–58. [Google Scholar] [CrossRef]
- Nambeesan, S.; Handa, A.K.; Mattoo, A.K. Polyamines and Regulation of Ripening and Senescence. In Postharvest Biology and Technology of Fruit, Vegetables and Flowers; Paliyath, G., Murr, D.P., Handa, A.K., Lurie, S., Eds.; Wiley-Blackwell: Oxford, UK, 2016. [Google Scholar]
- Feduraev, P.; Skrypnik, L.; Riabova, A.; Pungin, A.; Tokupova, E.; Maslennikov, P.; Chupakhina, G. Phenylalanine and tyrosine as exogenous precursors of wheat (Triticum aestivum L.) secondary metabolism through PAL-associated pathways. Plants 2020, 9, 476. [Google Scholar] [CrossRef]
- Jiao, Y.; Chen, Y.; Ma, C.; Qin, J.; Nguyen, T.H.N.; Liu, D.; Gan, H.; Ding, S.; Luo, Z.-B. Phenylalanine as a nitrogen source induces root growth and nitrogen-use efficiency in Populus× canescens. Tree Physiol. 2018, 38, 66–82. [Google Scholar] [CrossRef]
- Sato, K.; Yamane, M.; Yamaji, N.; Kanamori, H.; Tagiri, A.; Schwerdt, J.G.; Fincher, G.B.; Matsumoto, T.; Takeda, K.; Komatsuda, T. Alanine aminotransferase controls seed dormancy in barley. Nat. Commun. 2016, 7, 11625. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Kim, D.-W.; Khadka, P.; Rakwal, R.; Rohila, J.S. Unraveling key metabolomic alterations in wheat embryos derived from freshly harvested and water-imbibed seeds of two wheat cultivars with contrasting dormancy status. Front. Plant Sci. 2017, 8, 1203. [Google Scholar] [CrossRef]
- Verslues, P.E.; Sharma, S. Proline Metabolism and Its Implications for Plant-Environment Interaction. Arab. Book/Am. Soc. Plant Biol. 2010, 8, e0140. [Google Scholar] [CrossRef] [PubMed]
- Lechowska, K.; Wojtyla, Ł.; Quinet, M.; Kubala, S.; Lutts, S.; Garnczarska, M. Endogenous Polyamines and Ethylene Biosynthesis in Relation to Germination of Osmoprimed Brassica napus Seeds under Salt Stress Katarzyna. Int. J. Mol. Sci. 2022, 23, 349. [Google Scholar] [CrossRef] [PubMed]
- Llebrés, M.-T.; Pascual, M.-B.; Debille, S.; Trontin, J.-F.; Harvengt, L.P.; Avila, C.; Cánovas, F.M. The role of arginine metabolic pathway during embryogenesis and germination in maritime pine (Pinus pinaster Ait.). Tree Physiol. 2018, 38, 471–484. [Google Scholar] [PubMed]
- Dilworth, M.F.; Dure, L., III. Developmental biochemistry of cotton seed embryogenesis and germination: X. Nitrogen flow from arginine to asparagine in germination. Plant Physiol. 1978, 61, 698–702. [Google Scholar] [CrossRef]
- Shalaby, A. Changes in biogenic amines in mature and germinating legume seeds and their behavior during cooking. Food/Nahrung 2000, 44, 23–27. [Google Scholar] [CrossRef]
- MacGregor, D.R.; Kendall, S.L.; Florance, H.; Fedi, F.; Moore, K.; Paszkiewicz, K.; Smirnoff, N.; Penfield, S. Seed production temperature regulation of primary dormancy occurs through control of seed coat phenylpropanoid metabolism. New Phytol. 2014, 205, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, J.E. Seed coat and water absorption properties of seed of near-isogenic snap bean lines differing in seed coat color. J. Am. Soc. Hortic. Sci. 1977, 102, 478–480. [Google Scholar] [CrossRef]
- Corbineau, F.; Picard, M.; Côme, D. Germinability of leek seeds and its improvement by osmopriming. Acta Hortic. 1993, 371, 45–52. [Google Scholar] [CrossRef]
- Edwards, M. MDormancy in seeds of Charlock: III. Occurrence and mode of action of an inhibitor associated with dormancy. J. Exp. Bot. 1968, 19, 601–610. [Google Scholar] [CrossRef]
- Tobe, K.; Zhang, L.; Qiu, G.Y.; Shimizu, H.; Omasa, K. Characteristics of seed germination in five non-halophytic Chinese desert shrub species. J. Arid. Environ. 2001, 47, 191–201. [Google Scholar] [CrossRef]
- Wingler, A.; Henriques, R. Sugars and the speed of life—Metabolic signals that determine plant growth, development and death. Physiol. Plant. 2022, 174, e13656. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.H.; Veit, B.; Hanson, M.R. The Arabidopsis AtRaptor genes are essential for post-embryonic plant growth. BMC Biol. 2005, 3, 12. [Google Scholar] [CrossRef]
- Tixier, A.; Gambetta, G.A.; Godfrey, J.; Orozco, J.; Zwieniecki, M.A. Non-structural carbohydrates in dormant woody perennials; the tale of winter survival and spring arrival. Front. For. Glob. Chang. 2019, 2. [Google Scholar] [CrossRef]
- Tarancon, C.; González-Grando, E.; Oliveros, J.C.; Nicolas, M.; Cubas, P. A conserved carbon starvation response underlies bud dormancy in woody and herbaceous species. Front. Plant Sci. 2017, 8, 788. [Google Scholar]
- Park, J.Y.; Canam, T.; Kang, K.Y.; Unda, F.; Mansfield, S.D. Sucrose phosphate synthase expression influences poplar phenology. Tree Physiol. 2009, 29, 937–946. [Google Scholar] [CrossRef]
- Vandecasteele, C.; Teulat-Merah, B.; MorÈRe-LE Paven, M.C.; Leprince, O.; Ly Vu, B.; Viau, L.; Ledroit, L.; Pelletier, S.; Payet, N.; Satour, P.; et al. Quantitative trait loci analysis reveals a correlation between the ratio of sucrose/raffinose family oligosaccharides and seed vigour in Medicago truncatula. Plant Cell Environ. 2011, 34, 1473–1487. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fernie, A.R. On the role of the tricarboxylic cid cycle in plant productivity. J. Integr. Plant Biol. 2018, 60, 1199–1216. [Google Scholar] [CrossRef]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef]
- Fernie, A.R.; Malate, E. Jack of all trades or master of a few? Phytochemistry 2009, 70, 828–883. [Google Scholar] [CrossRef]
- Kim, H.U. Lipid metabolism in plants. Plants 2020, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.M.; Sun, Y.Y.; Ye, X.Y.; Li, Z.G. Signaling Role of Glutamate in Plants. Front. Plant Sci. 2020, 10, 1743. [Google Scholar] [CrossRef] [PubMed]
- Greco, M.; Chiappetta, A.; Bruno, L.; Bitonti, M.B. In Posidonia oceanica cadmium induces changes in DNA methylation and chromatin patterning. J. Exp. Bot. 2012, 63, 695–709. [Google Scholar] [CrossRef] [PubMed]
- Nieuwland, J.; Menges, M.; Murray, J.A. The plant cyclins. In Cell Cycle Control and Plant Development; Blackwell Publishing: Oxford, UK, 2007; Volume 41. [Google Scholar]
- Zsigmond, L.; Tomasskovics, B.; Deák, V.; Rigó, G.; Szabados, L.; Bánhegyi, G.; Szarka, A. Plant Physiology and Biochemistry Enhanced activity of galactono-1, 4-lactone dehydrogenase and ascorbate e glutathione cycle in mitochondria from complex III deficient Arabidopsis. Plant Physiol. Biochem. 2011, 49, 809–815. [Google Scholar] [CrossRef]
- Blokhina, O.; Fagerstedt, K.V. Reactive oxygen species and nitric oxide in plant mitochondria: Origin and redundant regulatory systems. Physiol. Plant. 2010, 138, 447–462. [Google Scholar]
- Paradiso, A.; Pinto, M.C.D.; Locato, V.; Gara, L.D. Galactone-γ-lactone-dependent ascorbate biosynthesis alters wheat kernel maturation. Plant Biol. 2012, 14, 1435–8603. [Google Scholar] [CrossRef]
- Xool-Tamayo, J.; Tamayo-Ordoñez, Y.; Monforte-González, M.; Muñoz-Sánchez, J.A.; Vázquez-Flota, F. Alkaloid Biosynthesis in the Early Stages of the Germination of Argemone mexicana L. (Papaveraceae). Plants 2021, 10, 2226. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Smith, D.L. Flavonoids in agriculture: Chemistry and roles in, biotic and abiotic stress responses, and microbial associations. Agronomy 2020, 10, 1209. [Google Scholar] [CrossRef]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds? New Phytol. 2018, 220, 692–702. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as Important Molecules of Plant Interactions with the Environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef]
- Corso, M.; Perreau, F.; Mouille, G.; Lepiniec, L. Specialized phenolic compounds in seeds: Structures, functions, and regulations. Plant Sci. 2020, 296, 110471. [Google Scholar]
- Higdon, J.V.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef]
- Arora, A.; Byrem, T.M.; Nair, M.G.; Strasburg, G.M. Modulation of liposomal membrane fluidity by flavonoids and isoflavonoids. Arch. Biochem. Biophys. 2000, 373, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Cotelle, N.; Bernier, J.L.; Catteau, J.P.; Pommery, J.; Wallet, J.C.; Gaydou, E.M. Antioxidant properties of hydroxy-flavones. Free Radic. Biol. Med. 1996, 20, 35–43. [Google Scholar] [CrossRef]
- Araniti, F.; Prinsi, B. Seed Germination Induced by Coumarin Is Mediated by a Lower Ability to Sustain the Energetic Metabolism. Plants 2022, 11, 843. [Google Scholar] [CrossRef] [PubMed]
- Chattha, F.A.; Munawar, M.A.; Nisa, M.; Kousar, S. Plant growth regulating activity of coumarins. In Coumarin-Based Heteroaromatics as Plant Growth Regulators; University of the Punjab: Lahore, Pakistan, 2016; pp. 91–104. [Google Scholar]
- Chen, Z.; Ma, Y.; Yang, R.; Gu, Z.; Wang, P. Effects of exogenous Ca2+ on phenolic accumulation and physiological changes in germinated wheat (Triticum aestivum L.) under UV-B radiation. Food Chem. 2019, 288, 368–376. [Google Scholar] [CrossRef]
- Pergo, É.M.; Abrahim, D.; Soares da Silva, P.C.; Kern, K.A.; Da Silva, L.J.; Voll, E.; Ishii-Iwamoto, E.L. Bidens pilosa L. exhibits high sensitivity to coumarin in comparison with three other weed species. J. Chem. Ecol. 2008, 34, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Berrie, A.M.M.; Parker, W.; Knights, B.A.; Hendrie, M.R. Studies on lettuce seed germination-I. Coumarin induced dormancy. Phytochemistry 1968, 7, 567–573. [Google Scholar] [CrossRef]
- Goren, R.; Tomer, E. Effect of seselin and coumarin on growth, indoleacetic acid oxidase and peroxidase with special reference to cucumber radicles. Plant Physiol. 1971, 47, 312–316. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed]
- Ninkuu, V.; Zhang, L.; Yan, J.; Fu, Z.; Yang, T.; Zeng, H. Biochemistry of Terpenes and Recent Advances in Plant Protection. Int. J. Mol. Sci. 2021, 22, 5710. [Google Scholar] [CrossRef]
- Das, A.K.; Anik, T.R.; Rahman, M.M.; Keya, S.S.; Islam, M.R.; Rahman, M.A.; Sultana, S.; Ghosh, P.K.; Khan, S.; Ahamed, T.; et al. Ethanol Treatment Enhances Physiological and Biochemical Responses to Mitigate Saline Toxicity in Soybean. Plants 2022, 11, 272. [Google Scholar] [CrossRef] [PubMed]
- Ventura, I.; Brunello, L.; Iacopino, S.; Valeri, M.C.; Novi, G.; Dornbusch, T.; Perata, P.; Loreti, E. Arabidopsis phenotyping reveals the importance of alcohol dehydrogenase and pyruvate decarboxylase for aerobic plant growth. Sci. Rep. 2020, 10, 16669. [Google Scholar] [CrossRef]
- Su, W.; Ren, Y.; Wang, D.; Su, Y.; Feng, J.; Zhang, C.; Tang, H.; Xu, L.; Muhammad, K.; Que, Y. The alcohol dehydrogenase gene family in sugarcane and its involvement in cold stress regulation. BMC Genom. 2020, 21, 521. [Google Scholar]
- Skalák, J.; Vercruyssen, L.; Claeys, H.; Hradilová, J.; Černý, M.; Novák, O.; Plačková, L.; Saiz-Fernández, I.; Skaláková, P.; Coppens, F.; et al. Multifaceted activity of cytokinin in leaf development shapes its size and structure in Arabidopsis. Plant J. 2019, 97, 805–824. [Google Scholar] [CrossRef]
- Mao, J.; Li, W.; Mi, B.; Mujitaba, M.; Ma, Z.; Zhang, Y.; Chen, B.; Caldero, A. Different exogenous sugars affect the hormone signal pathway and sugar metabolism in “Red Globe” (Vitis vinifera L.) plantlets grown in vitro as shown by transcriptomic analysis. Planta 2017, 246, 537–552. [Google Scholar] [CrossRef]
- Magome, H.; Nomura, T.; Hanada, A.; Takeda-Kamiya, N.; Ohnishi, T.; Shinma, Y.; Katsumata, T.; Kawaide, H.; Kamiya, Y.; Yamaguchi, S. CYP714B1 and CYP714B2 encode gibberellin 13-oxidases that reduce gibberellin activity in rice. Proc. Natl. Acad. Sci. USA 2013, 110, 1947–1952. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, A.; Savka, M.A.; Hudson, A.O. The synthesis and role of β-alanine in plants. Front. Plant Sci. 2019, 10, 921. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.; Sodek, L.; Licausi, F.; Hameed, M.W.; Dornelas, M.C.; Van Dongen, J.T. Analysis of alanine aminotransferase in various organs of soybean (Glycine max) and in dependence of different nitrogen fertilisers during hypoxic stress. Amino Acids 2010, 39, 1043–1053. [Google Scholar] [CrossRef]
- Zhong, M.; Liu, X.; Liu, F.; Ren, Y.; Wang, Y.; Zhu, J.; Teng, X.; Duan, E.; Wang, F.; Zhang, H. FLOURY ENDOSPERM12 encoding alanine aminotransferase 1 regulates carbon and nitrogen metabolism in rice. J. Plant Biol. 2019, 62, 61–73. [Google Scholar] [CrossRef]
- Tzin, V.; Malitsky, S.; Aharoni, A.; Galili, G. Expression of a bacterial bi-functional chorismate mutase/prephenate dehydratase modulates primary and secondary metabolism associated with aromatic amino acids in Arabidopsis. Plant J. 2009, 60, 156–167. [Google Scholar] [CrossRef]
- Vanholme, R.; De Meester, B.; Ralph, J.; Boerjan, W. Lignin biosynthesis and its integration into metabolism. Curr. Opin. Biotechnol. 2019, 56, 230–239. [Google Scholar] [CrossRef]
- Schenck, C.A.; Maeda, H.A. Tyrosine biosynthesis, metabolism, and catabolism in plants. Phytochemistry 2018, 149, 82–102. [Google Scholar] [PubMed]
- Dennis, R. The Role of Primary Carbohydrate Metabolism in Wheat Grain Dormancy and Germination. Ph.D. Thesis, The Australian National University, Canberra, Australia, 2019. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Query | Match | Chem-Fun Group | HMDB | Pub Chem | KEGG |
|---|---|---|---|---|---|
| 9,12,15-Octadecatrienoic acid, (Z,Z,Z)- | Alpha-Linolenic acid | Fatty acids | HMDB0001388 | 5280934 | C06427 |
| Ethylenediamine-N,N′-dipropionic acid | Edetic Acid | Flavonoids | HMDB0015109 | 6049 | C00284 |
| 1-Butanol, 2-methyl-, acetate | Methyl methacrylate | Esters | HMDB0032385 | 6658 | C19504 |
| Tetraacetyl-d-xylonic nitrile | Sarcosine | Amino Acids and Derivatives | HMDB0000271 | 1088 | C00213 |
| (+)-N-Acetylmuramic acid | UDP-N-acetylmuraminate | nucleotide-sugar | HMDB0011720 | 24755495 | C01050 |
| 1-Butanol, 3-methyl-, formate | 3-Methylbutyl formate | Fatty esters | HMDB0034163 | 8052 | C12293 |
| 1,3-Propanediol, 2-ethyl-2-(hydroxymethyl)- | 1,3-Butanediol | Alcohols and Polyols | HMDB0031320 | 7896 | C20335 |
| 1,8-Di(4-nitrophenylmethyl)-3,6-diazahomoadamantan-9-one | Tyramine | Amino Acids and Derivatives | HMDB0000306 | 5610 | C00483 |
| Benzeneethanamine, 2-fluoro-á,3,4-trihydroxy-N-isopropyl- | Phenylethylamine | Amines, Aromatic | HMDB0012275 | 1001 | C05332 |
| Desulphosinigrin | Delphinidin 3-O-sophoroside | Flavonoids | 47205615 | C16307 | |
| Glycerin | Glycerol | Alcohols and Polyols | HMDB0000131 | 753 | C00116 |
| 9,12-Octadecadienoic acid (Z,Z)- | Linoleic acid | Lipids | HMDB0000673 | 5280450 | C01595 |
| Dasycarpidan-1-methanol, acetate (ester) | Acetaminophen | Flavonoids | HMDB0001859 | 1983 | C06804 |
| 2,2,4-Trimethyl-1,3-pentanediol diisobutyrate | 3-Phenylpropyl 2-methylpropanoate | Alcohols | HMDB0034472 | 7662 | C02008 |
| Sucrose | Sucrose | Disaccharides | HMDB0000258 | 5988 | C00089 |
| 5-O-Methyl-d-gluconic acid dimethylamide | 3-Cresotinic acid | Phytohormone | HMDB0002390 | 6738 | C14088 |
| 2,6-Dioxa-tricyclo [3.3.2.0(3,7)] decan-9-one | Ibuprofen | Amines | HMDB0001925 | 3672 | C01588 |
| 1,3-Cyclohexanedione, 2,5,5-trimethyl- | 1,4-Cyclohexanedione | Alcohols | 10263 | C08063 | |
| 5-Methoxypyrrolidin-2-one | Galactosylglycerol | Alkaloids | HMDB0006790 | 656504 | C05401 |
| 2(3H)-Furanone, 5-heptyldihydro | Galactonolactone | Sugar Acids | HMDB0002541 | 5640 | C03383 |
| 2-Propanol, 1,1′-oxybis- | Propranolol | Amino Alcohols | HMDB0001849 | 4946 | C07407 |
| 10-Hydroxydecanoic acid | 12-Hydroxydodecanoic acid | Fatty Acids | HMDB0002059 | 79034 | C08317 |
| Succinamide | Putrescine | Biogenic Polyamines | HMDB0001414 | 1045 | C00134 |
| 2-Octenoic acid, 4,5,7-trhydroxy | Methyl acrylate | Carboxylic Acids | HMDB0033977 | 7294 | C19443 |
| 2-Hydroxy-3-methoxy-succinic acid, dimethyl ester | L-Malic acid | carboxylic acid | HMDB0000156 | 222656 | C00149 |
| Octanoic acid, 2-phenylethyl ester | Styrene | Benzene Derivatives | HMDB0034240 | 7501 | C19506 |
| 2-Cyclohexen-1-one, 4-(3-hydroxy-1-butenyl)-3,5,5-trimethyl- | Amoxycillin | Terpenoids | HMDB0030500 | 2171 | C06827 |
| 8-Hydroxy-7-methoxycoumarin | Oxybenzone | Coumarins | HMDB0015497 | 4632 | C14285 |
| Pyrimidine-4,6-diol, 5-methyl- | Uracil | Nucleic Acids and Derivatives | HMDB0000300 | 1174 | C00106 |
| Oxime-, methoxy-phenyl-_ | 3,4-Dihydroxyphenylacetaldehyde | phenols | HMDB0003791 | 119219 | C04043 |
| Adenine | Adenine | Nucleotides | HMDB0000034 | 190 | C00147 |
| 2,5-Cyclohexadiene-1,4-dione, dioxime | Quinone | Benzoquinones | HMDB0003364 | 4650 | C00472 |
| 2,5-Pyrrolidinedione, 3-(1-aminoethylidene)-4-methyl- | Isopyridoxal | Alkaloids | HMDB0004290 | 440899 | C06051 |
| Phthalic acid, hept-4-yl isobutyl ester | Thalidomide | Carboxylic Acids | HMDB0015175 | 92142 | C07910 |
| L-Glutamine | L-Glutamine | Amino Acids and Derivatives | HMDB0000641 | 5961 | C00064 |
| Methyl cis-13,16-Docosadienate | Methylitaconate | carboxylic acid | METPA0268 | C02295 | |
| 2-Hydroxyhippuric acid-3TMS | Salicyluric acid | Phytohormone | HMDB0000840 | 10253 | C07588 |
| 2,4-Di-tert-butylphenoxytrimethylsilane | Butenafine | Amines | HMDB0015223 | 2484 | C08067 |
| Pentasiloxane, dodecamethyl- | Dodecamethylpentasiloxane | organosiloxane | HMDB0062731 | 8853 |
| Total | Expected | Hits | Raw p | −log10 (p) | Holm Adjust | FDR | Impact | |
|---|---|---|---|---|---|---|---|---|
| Isoquinoline alkaloid biosynthesis | 6 | 0.08 | 2 | 2.26 × 10−3 | 2.65 × 10 | 2.15 × 10−1 | 2.03 × 10−1 | 0.00 |
| Galactose metabolism | 27 | 0.35 | 3 | 4.28 × 10−3 | 2.37 × 10 | 4.02 × 10−1 | 2.03 × 10−1 | 0.76 |
| Tyrosine metabolism | 18 | 0.23 | 2 | 2.11 × 10−2 | 1.68 × 10 | 1.00 × 10 | 6.67 × 10−1 | 0.04 |
| Biosynthesis of unsaturated fatty acids | 22 | 0.28 | 2 | 3.09 × 10−2 | 1.51 × 10 | 1.00 × 10 | 7.33 × 10−1 | 0.00 |
| Linoleic acid metabolism | 4 | 0.05 | 1 | 5.04 × 10−2 | 1.30 × 10 | 1.00 × 10 | 8.15 × 10−1 | 1.00 |
| Glyoxylate and dicarboxylate | 29 | 0.37 | 2 | 5.14 × 10−2 | 1.29 × 10 | 1.00 × 10 | 8.15 × 10−1 | 0.06 |
| metabolism | ||||||||
| Pyrimidine metabolism | 38 | 0.49 | 2 | 8.32 × 10−2 | 1.08 × 10 | 1.00 × 10 | 1.00 × 10 | 0.06 |
| Lysine biosynthesis | 9 | 0.12 | 1 | 1.10 × 10−1 | 9.59 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.47 |
| Nitrogen metabolism | 12 | 0.15 | 1 | 1.44 × 10−1 | 8.41 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.54 |
| Phenylalanine metabolism | 12 | 0.15 | 1 | 1.44 × 10−1 | 8.41 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.79 |
| Purine metabolism | 63 | 0.81 | 2 | 1.92 × 10−1 | 7.17 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.26 |
| Arginine biosynthesis | 18 | 0.23 | 1 | 2.09 × 10−1 | 6.81 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.00 |
| beta-Alanine metabolism | 18 | 0.23 | 1 | 2.09 × 10−1 | 6.81 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.00 |
| Citrate cycle (TCA cycle) | 20 | 0.26 | 1 | 2.29 × 10−1 | 6.40 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.33 |
| Zeatin biosynthesis | 21 | 0.27 | 1 | 2.39 × 10−1 | 6.22 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.00 |
| Carbon fixation in photosynthetic organ- | 21 | 0.27 | 1 | 2.39 × 10−1 | 6.22 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.06 |
| isms | ||||||||
| Glycerolipid metabolism | 21 | 0.27 | 1 | 2.39 × 10−1 | 6.22 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.26 |
| Phenylalanine, tyrosine, and tryptophan | 22 | 0.28 | 1 | 2.49 × 10−1 | 6.04 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.08 |
| biosynthesis | ||||||||
| Starch and sucrose metabolism | 22 | 0.28 | 1 | 2.49 × 10−1 | 6.04 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.59 |
| Pyruvate metabolism | 22 | 0.28 | 1 | 2.49 × 10−1 | 6.04 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.24 |
| Alanine, aspartate and glutamate | 22 | 0.28 | 1 | 2.49 × 10−1 | 6.04 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.57 |
| metabolism | ||||||||
| Pantothenate and CoA biosynthesis | 23 | 0.30 | 1 | 2.59 × 10−1 | 5.87 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.00 |
| Cyanoamino acid metabolism | 26 | 0.33 | 1 | 2.87 × 10−1 | 5.42 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.00 |
| Glutathione metabolism | 27 | 0.35 | 1 | 2.97 × 10−1 | 5.28 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.01 |
| alpha-Linolenic acid metabolism | 27 | 0.35 | 1 | 2.97 × 10−1 | 5.28 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.11 |
| Arginine and proline metabolism | 28 | 0.36 | 1 | 3.06 × 10−1 | 5.14 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.16 |
| Glycine, serine, and threonine metabolism | 33 | 0.42 | 1 | 3.50 × 10−1 | 4.56 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.00 |
| Aminoacyl-tRNA biosynthesis | 46 | 0.59 | 1 | 4.53 × 10−1 | 3.44 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.00 |
| Porphyrin and chlorophyll metabolism | 47 | 0.60 | 1 | 4.61 × 10−1 | 3.37 × 10−1 | 1.00 × 10 | 1.00 × 10 | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nwogha, J.S.; Wosene, A.G.; Raveendran, M.; Obidiegwu, J.E.; Oselebe, H.O.; Kambale, R.; Chilaka, C.A.; Rajagopalan, V.R. Comparative Metabolomics Profiling Reveals Key Metabolites and Associated Pathways Regulating Tuber Dormancy in White Yam (Dioscorea rotundata Poir.). Metabolites 2023, 13, 610. https://doi.org/10.3390/metabo13050610
Nwogha JS, Wosene AG, Raveendran M, Obidiegwu JE, Oselebe HO, Kambale R, Chilaka CA, Rajagopalan VR. Comparative Metabolomics Profiling Reveals Key Metabolites and Associated Pathways Regulating Tuber Dormancy in White Yam (Dioscorea rotundata Poir.). Metabolites. 2023; 13(5):610. https://doi.org/10.3390/metabo13050610
Chicago/Turabian StyleNwogha, Jeremiah S., Abtew G. Wosene, Muthurajan Raveendran, Jude E. Obidiegwu, Happiness O. Oselebe, Rohit Kambale, Cynthia A. Chilaka, and Veera Ranjani Rajagopalan. 2023. "Comparative Metabolomics Profiling Reveals Key Metabolites and Associated Pathways Regulating Tuber Dormancy in White Yam (Dioscorea rotundata Poir.)" Metabolites 13, no. 5: 610. https://doi.org/10.3390/metabo13050610