
SlbHLH22-Induced Hypertrophy Development Is Related to the Salt Stress Response of the GTgamma Gene in Tomatoes

Abstract
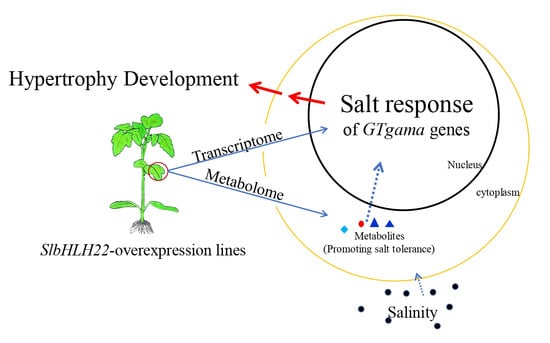
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Transcriptome and Metabolome Analysis
2.3. Hormonal and Salt Treatments
2.4. RT-PCR
2.5. Statistic Analysis
2.6. Computational Modeling
3. Results
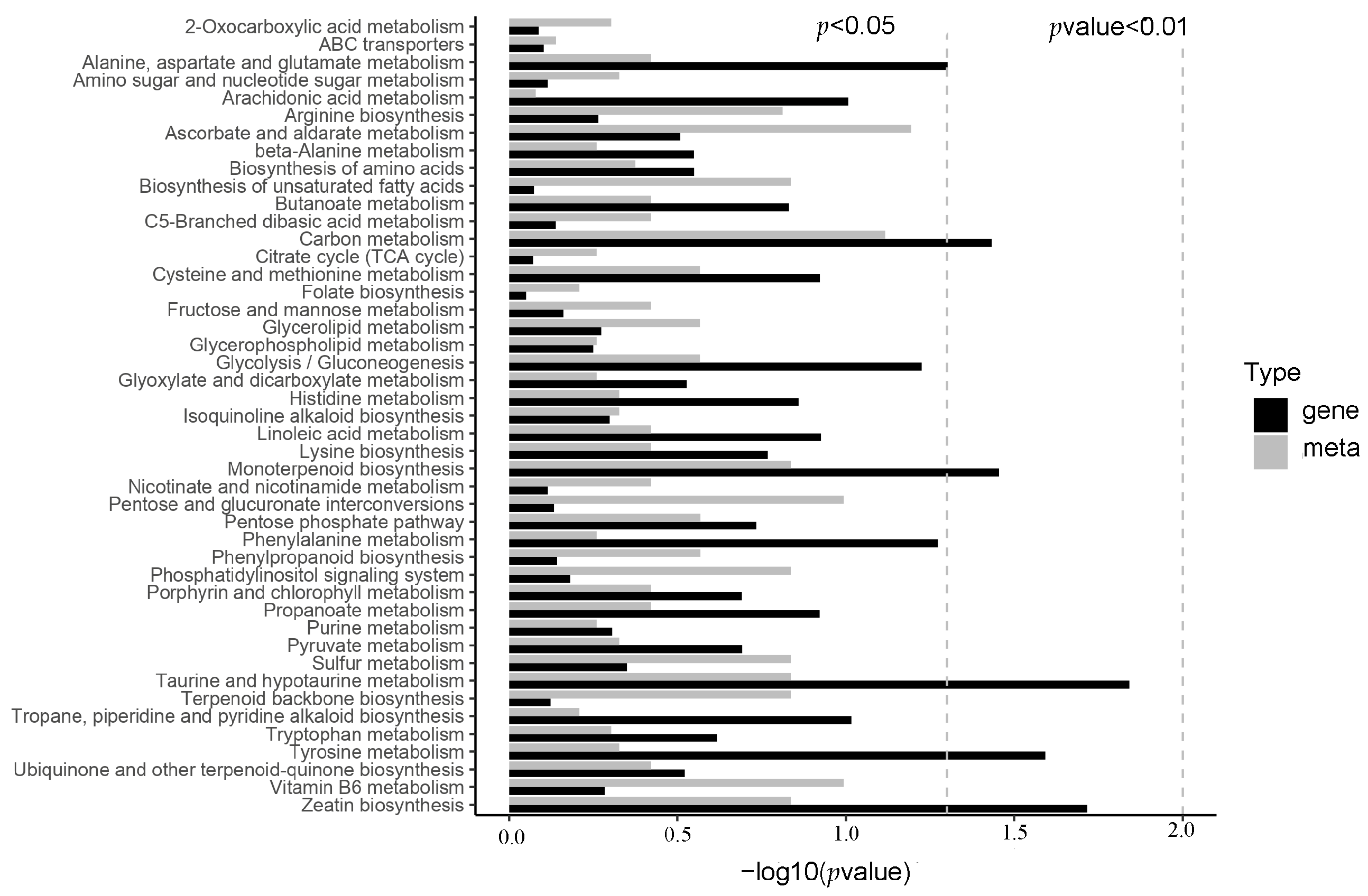
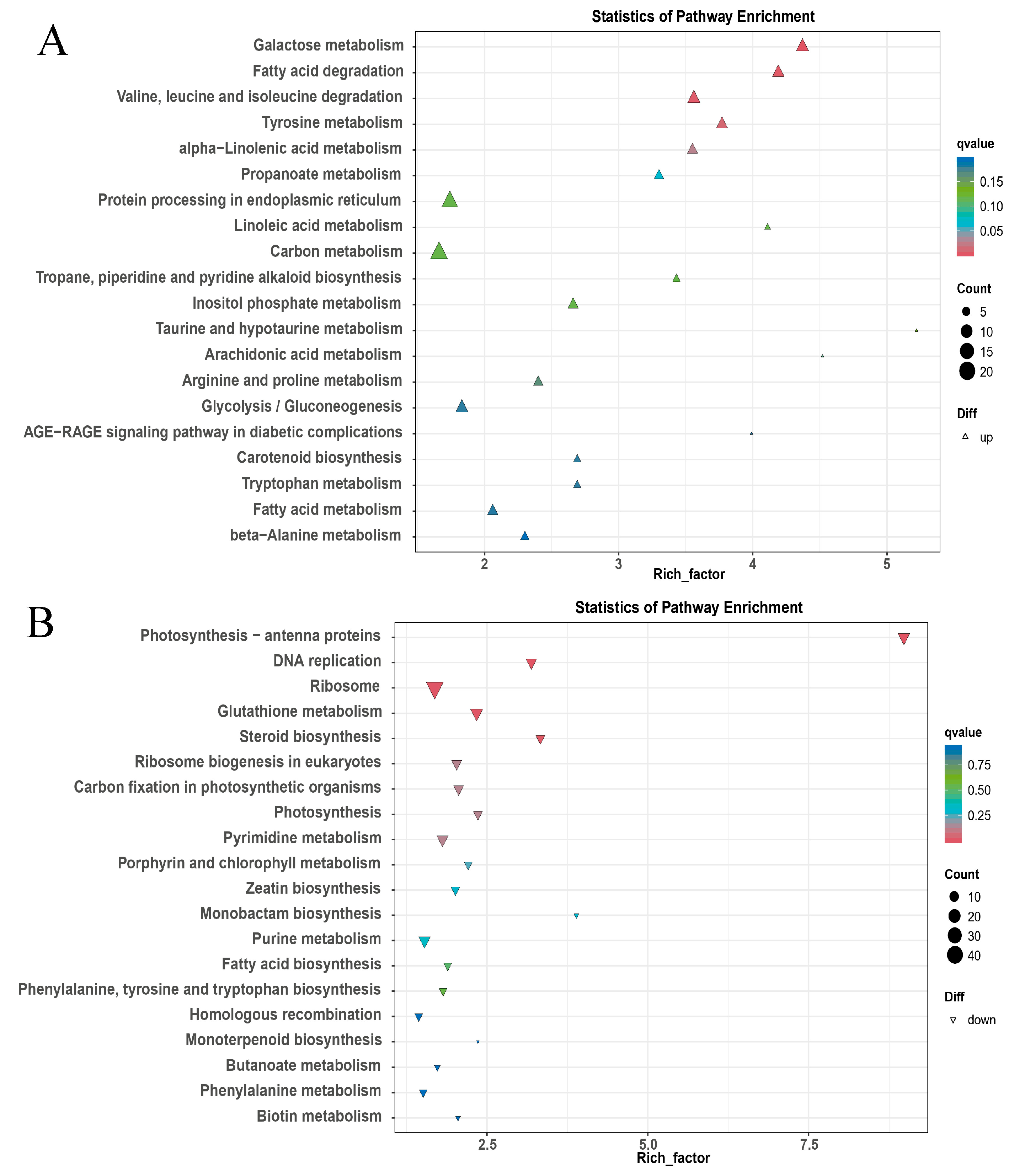
3.1. Metabolome Analysis of OE vs. WT
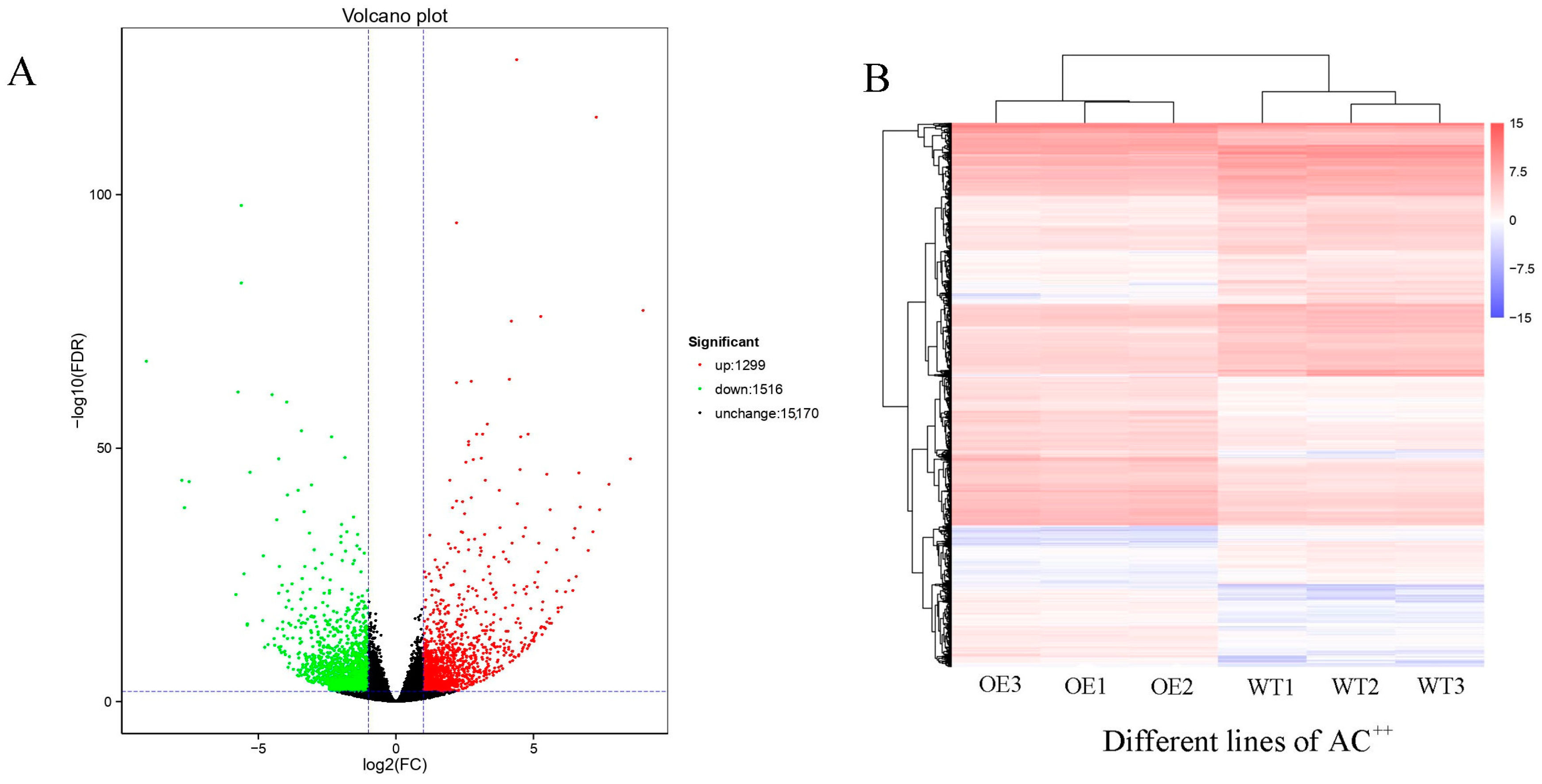
3.2. Transcriptome Analysis of OE vs. WT
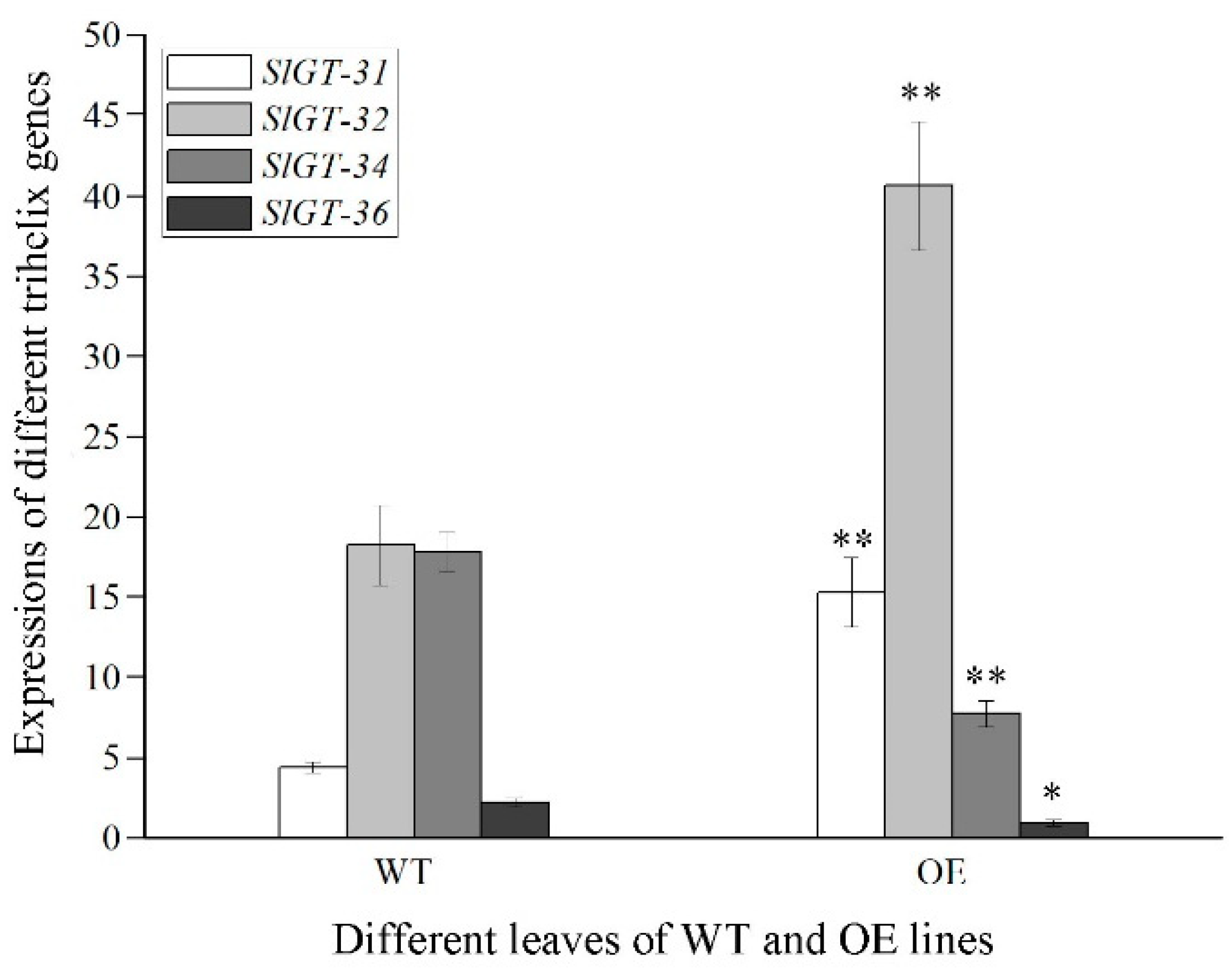
3.3. Analysis of the Transcription Factor among DEGs
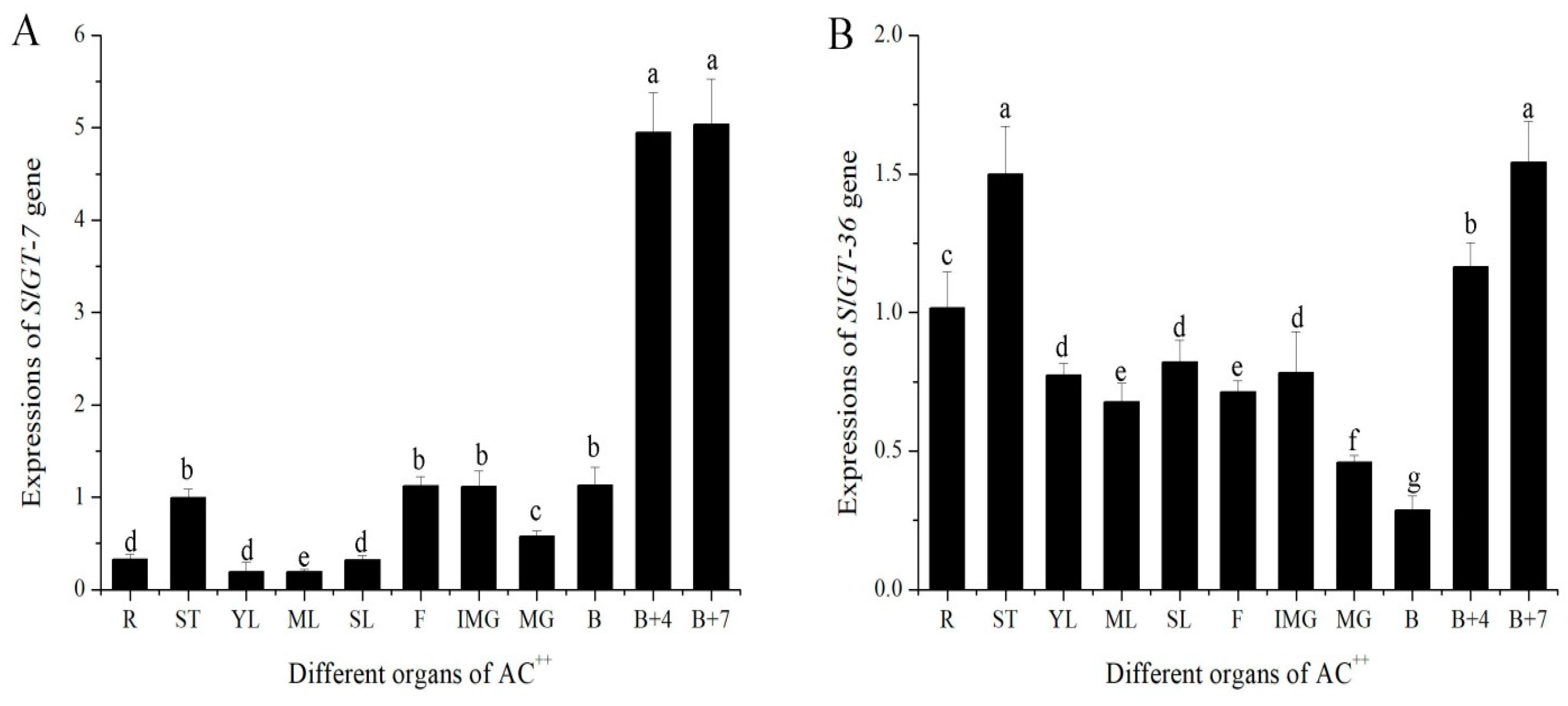
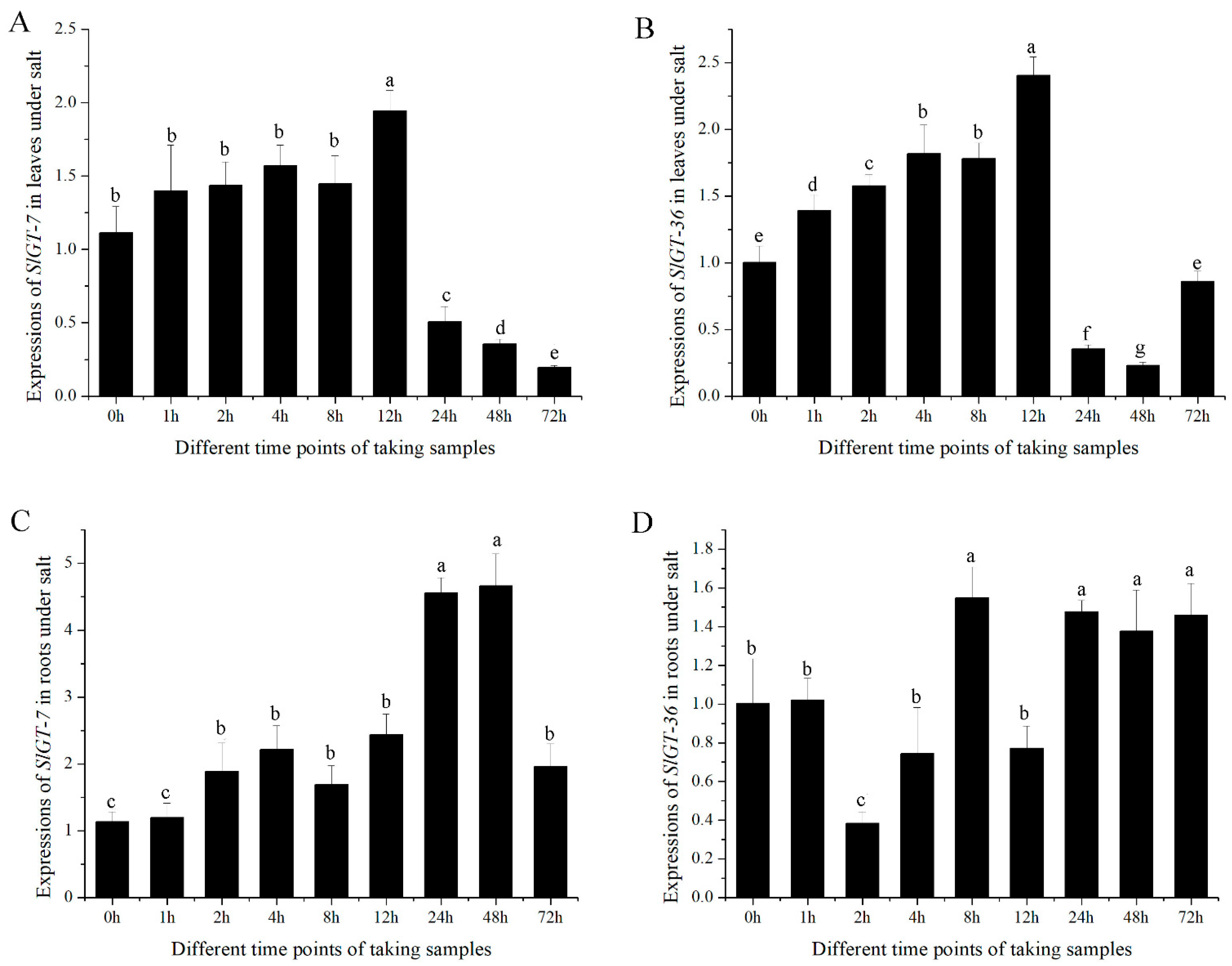
3.4. Expression Patterns of GTgamma Genes in AC++ and Their Responses to External Stimuli
3.5. Three-Dimensional Structures of SlGT-7 and Its Potential Phosphorylation Site
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Romer, P.; Recht, S.; Strauss, T.; Elsaesser, J.; Schornack, S.; Boch, J.; Wang, S.; Lahaye, T. Promoter elements of rice susceptibility genes are bound and activated by specific TAL effectors from the bacterial blight pathogen, Xanthomonas oryzae pv. oryzae. New Phytol. 2010, 187, 1048–1057. [Google Scholar] [CrossRef] [PubMed]
- Kay, S.; Hahn, S.; Marois, E.; Hause, G.; Bonas, U. A bacterial effector acts as a plant transcription factor and induces a cell size regulator. Science 2007, 318, 648–651. [Google Scholar] [CrossRef] [PubMed]
- Lukhovitskaya, N.I.; Solovieva, A.D.; Boddeti, S.K.; Thaduri, S.; Solovyev, A.G.; Savenkov, E.I. An RNA virus-encoded zinc-finger protein acts as a plant transcription factor and induces a regulator of cell size and proliferation in two tobacco species. Plant Cell 2013, 25, 960–973. [Google Scholar] [CrossRef]
- Cui, B.; Hu, Z.; Hu, J.; Zhang, Y.; Yin, W.; Zhu, Z.; Feng, Y.; Chen, G. Overexpression of SlUPA-like induces cell enlargement, aberrant development and low stress tolerance through phytohormonal pathway in tomato. Sci. Rep. 2016, 6, 23818. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Zhang, W.; Lv, J.; Yang, J.; Yang, S.; Jia, B.; Song, J.; Wu, M.; Pei, W.; Ma, J.; et al. Overexpression of cotton Trihelix transcription factor GhGT-3b_A04 enhances resistance to Verticillium dahliae and affects plant growth in Arabidopsis thaliana. J. Plant Physiol. 2023, 283, 153947. [Google Scholar] [CrossRef]
- Li, B.; Jiang, S.; Yu, X.; Cheng, C.; Chen, S.; Cheng, Y.; Yuan, J.S.; Jiang, D.; He, P.; Shan, L. Phosphorylation of trihelix transcriptional repressor ASR3 by MAP KINASE4 negatively regulates Arabidopsis immunity. Plant Cell 2015, 27, 839–856. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, M.; Zhang, Y.; Huang, M.; Wei, L.; Lin, Y.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D.; et al. Coordinated regulation of plant defense and autoimmunity by paired trihelix transcription factors ASR3/AITF1 in Arabidopsis. New Phytol. 2023, 237, 914–929. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhong, T.; E, L.; Xu, M.; Dai, W.; Sun, S.; Ye, J. GT Factor ZmGT-3b Is Associated with Regulation of Photosynthesis and Defense Response to Fusarium graminearum Infection in Maize Seedling. Front. Plant Sci. 2021, 12, 724133. [Google Scholar] [CrossRef]
- Yu, C.; Song, L.; Song, J.; Ouyang, B.; Guo, L.; Shang, L.; Wang, T.; Li, H.; Zhang, J.; Ye, Z. ShCIGT, a Trihelix family gene, mediates cold and drought tolerance by interacting with SnRK1 in tomato. Plant Sci. 2018, 270, 140–149. [Google Scholar] [CrossRef]
- David, S.; Levin, E.; Fallik, E.; Alkalai-Tuvia, S.; Foolad, M.R.; Lers, A. Physiological genetic variation in tomato fruit chilling tolerance during postharvest storage. Front. Plant Sci. 2022, 13, 991983. [Google Scholar] [CrossRef]
- Fang, Y.; Xie, K.; Hou, X.; Hu, H.; Xiong, L. Systematic analysis of GT factor family of rice reveals a novel subfamily involved in stress responses. Mol. Genet. Genom. MGG 2010, 283, 157–169. [Google Scholar] [CrossRef]
- Liu, X.; Wu, D.; Shan, T.; Xu, S.; Qin, R.; Li, H.; Negm, M.; Wu, D.; Li, J. The trihelix transcription factor OsGTgamma-2 is involved adaption to salt stress in rice. Plant Mol. Biol. 2020, 103, 545–560. [Google Scholar] [CrossRef] [PubMed]
- Waseem, M.; Rong, X.; Li, Z. Dissecting the Role of a Basic Helix-Loop-Helix Transcription Factor, SlbHLH22, under Salt and Drought Stresses in Transgenic Solanum lycopersicum L. Front. Plant Sci. 2019, 10, 734. [Google Scholar] [CrossRef] [PubMed]
- Marechal, E.; Hiratsuka, K.; Delgado, J.; Nairn, A.; Qin, J.; Chait, B.T.; Chua, N.H. Modulation of GT-1 DNA-binding activity by calcium-dependent phosphorylation. Plant Mol. Biol. 1999, 40, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Nagata, T.; Niyada, E.; Fujimoto, N.; Nagasaki, Y.; Noto, K.; Miyanoiri, Y.; Murata, J.; Hiratsuka, K.; Katahira, M. Solution structures of the trihelix DNA-binding domains of the wild-type and a phosphomimetic mutant of Arabidopsis GT-1: Mechanism for an increase in DNA-binding affinity through phosphorylation. Proteins 2010, 78, 3033–3047. [Google Scholar] [CrossRef] [PubMed]
- O'Brien, M.; Kaplan-Levy, R.N.; Quon, T.; Sappl, P.G.; Smyth, D.R. PETAL LOSS, a trihelix transcription factor that represses growth in Arabidopsis thaliana, binds the energy-sensing SnRK1 kinase AKIN10. J. Exp. Bot. 2015, 66, 2475–2485. [Google Scholar] [CrossRef]
- Fu, M.; Li, F.; Zhou, S.; Guo, P.; Chen, Y.; Xie, Q.; Chen, G.; Hu, Z. Trihelix transcription factor SlGT31 regulates fruit ripening mediated by ethylene in tomato. J. Exp. Bot. 2023, 74, 5709–5721. [Google Scholar] [CrossRef]
- Fang, X.; Wang, Y.; Cui, J.; Yue, L.; Jiang, A.; Liu, J.; Wu, Y.; He, X.; Li, C.; Zhang, J.; et al. Transcriptome and metabolome analyses reveal the key genes related to grain size of big grain mutant in Tartary Buckwheat (Fagopyrum tartaricum). Front. Plant Sci. 2022, 13, 1079212. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Y.; Li, W.; Lin, Y.; Wang, C.; Xu, R.; Zhang, L. Genome-wide characterization and expression analysis of soybean trihelix gene family. PeerJ 2020, 8, e8753. [Google Scholar] [CrossRef]
- Zhu, M.; Chen, G.; Zhang, J.; Zhang, Y.; Xie, Q.; Zhao, Z.; Pan, Y.; Hu, Z. The abiotic stress-responsive NAC-type transcription factor SlNAC4 regulates salt and drought tolerance and stress-related genes in tomato (Solanum lycopersicum). Plant Cell Rep. 2014, 33, 1851–1863. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Mao, L.; Guan, W.; Wei, X.; Shao, Y.; Luo, Z.; Lin, X.; Li, L. Exogenous 24-epibrassinolide activates detoxification enzymes to promote degradation of boscalid in cherry tomatoes. J. Sci. Food Agric. 2021, 101, 2210–2217. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Chen, G.; Zhou, S.; Tu, Y.; Wang, Y.; Dong, T.; Hu, Z. A new tomato NAC (NAM/ATAF1/2/CUC2) transcription factor, SlNAC4, functions as a positive regulator of fruit ripening and carotenoid accumulation. Plant Cell Physiol. 2014, 55, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Abdelhedi, O.; Nasri, R.; Mora, L.; Jridi, M.; Toldra, F.; Nasri, M. In silico analysis and molecular docking study of angiotensin I-converting enzyme inhibitory peptides from smooth-hound viscera protein hydrolysates fractionated by ultrafiltration. Food Chem. 2018, 239, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Apodiakou, A.; Hoefgen, R. New insights into the regulation of plant metabolism by O-acetylserine: Sulfate and beyond. J. Exp. Bot. 2023, 74, 3361–3378. [Google Scholar] [CrossRef] [PubMed]
- Kusakari, K.; Fukuhara, T.; Motoyama, A.; Ochiai, N.; Watanabe, T.; Sugimoto, Y. The corrected structure of depressoside, an antioxidative iridoid glucoside extracted from the flowers of Gentiana urnula Harry Sm. Nat. Prod. Res. 2016, 30, 954–959. [Google Scholar] [CrossRef]
- Araujo, W.L.; Martins, A.O.; Fernie, A.R.; Tohge, T. 2-Oxoglutarate: Linking TCA cycle function with amino acid, glucosinolate, flavonoid, alkaloid, and gibberellin biosynthesis. Front. Plant Sci. 2014, 5, 552. [Google Scholar] [CrossRef]
- Hubberten, H.M.; Klie, S.; Caldana, C.; Degenkolbe, T.; Willmitzer, L.; Hoefgen, R. Additional role of O-acetylserine as a sulfur status-independent regulator during plant growth. Plant J. 2012, 70, 666–677. [Google Scholar] [CrossRef]
- Ding, F.; Wang, R.; Chen, B. Effect of exogenous ammonium gluconate on growth, ion flux and antioxidant enzymes of maize (Zea Mays L.) seedlings under NaCl stress. Plant Biol. 2019, 21, 643–651. [Google Scholar] [CrossRef]
- Green, P.J.; Kay, S.A.; Chua, N.H. Sequence-specific interactions of a pea nuclear factor with light-responsive elements upstream of the rbcS-3A gene. EMBO J. 1987, 6, 2543–2549. [Google Scholar] [CrossRef]
- Kaplan-Levy, R.N.; Brewer, P.B.; Quon, T.; Smyth, D.R. The trihelix family of transcription factors—Light, stress and development. Trends Plant Sci. 2012, 17, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Alamri, S.; Alsubaie, Q.D.; Al-Amri, A.A.; Al-Munqedi, B.; Ali, H.M.; Kushwaha, B.K.; Singh, V.P.; Siddiqui, M.H. Priming of tomato seedlings with 2-oxoglutarate induces arsenic toxicity alleviatory responses by involving endogenous nitric oxide. Physiol. Plant 2021, 173, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Waseem, M.; Li, N.; Su, D.; Chen, J.; Li, Z. Overexpression of a basic helix-loop-helix transcription factor gene, SlbHLH22, promotes early flowering and accelerates fruit ripening in tomato (Solanum lycopersicum L.). Planta 2019, 250, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Le Gourrierec, J.; Li, Y.F.; Zhou, D.X. Transcriptional activation by Arabidopsis GT-1 may be through interaction with TFIIA-TBP-TATA complex. Plant J. 1999, 18, 663–668. [Google Scholar] [CrossRef]
- Yu, C.; Cai, X.; Ye, Z.; Li, H. Genome-wide identification and expression profiling analysis of trihelix gene family in tomato. Biochem. Biophys. Res. Commun. 2015, 468, 653–659. [Google Scholar] [CrossRef]
- Minguez, P.; Parca, L.; Diella, F.; Mende, D.R.; Kumar, R.; Helmer-Citterich, M.; Gavin, A.C.; van Noort, V.; Bork, P. Deciphering a global network of functionally associated post-translational modifications. Mol. Syst. Biol. 2012, 8, 599. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial Number | TF Family | DEGs Numbers | Serial Number | TF Family | DEGs Numbers | Serial Number | TF Family | DEGs Numbers |
|---|---|---|---|---|---|---|---|---|
| 1 | AP2/ERF-AP2 | 2 | 17 | E2F-DP | 1 | 33 | MYB-related | 4 |
| 2 | AP2/ERF-ERF | 19 | 18 | EIL | 1 | 34 | NAC | 19 |
| 3 | B3 | 5 | 19 | GARP-ARR-B | 1 | 35 | NF-YA | 4 |
| 4 | B3-ARF | 2 | 20 | GARP-G2-like | 2 | 36 | NF-YB | 1 |
| 5 | BBR-BPC | 1 | 21 | GeBP | 1 | 37 | NF-YC | 1 |
| 6 | bHLH | 15 | 22 | GRAS | 4 | 38 | OFP | 1 |
| 7 | bZIP | 10 | 23 | HB-BELL | 2 | 39 | PLATZ | 2 |
| 8 | C2C2-CO-like | 2 | 24 | HB-HD-ZIP | 15 | 40 | RWP-RK | 1 |
| 9 | C2C2-Dof | 5 | 25 | HB-KNOX | 2 | 41 | SRS | 1 |
| 10 | C2C2-GATA | 3 | 26 | HB-other | 4 | 42 | TCP | 6 |
| 11 | C2C2-YABBY | 2 | 27 | HMG | 2 | 43 | Tify | 2 |
| 12 | C2H2 | 11 | 28 | HSF | 6 | 44 | Trihelix | 4 |
| 13 | C3H | 2 | 29 | LOB | 1 | 45 | WRKY | 8 |
| 14 | CPP | 1 | 30 | MADS-MIKC | 8 | 46 | zf-HD | 1 |
| 15 | DBB | 1 | 31 | MADS-M-type | 3 | |||
| 16 | DBP | 1 | 32 | MYB | 16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, B.; Yu, M.; Bai, J.; Zhu, Z. SlbHLH22-Induced Hypertrophy Development Is Related to the Salt Stress Response of the GTgamma Gene in Tomatoes. Metabolites 2023, 13, 1195. https://doi.org/10.3390/metabo13121195
Cui B, Yu M, Bai J, Zhu Z. SlbHLH22-Induced Hypertrophy Development Is Related to the Salt Stress Response of the GTgamma Gene in Tomatoes. Metabolites. 2023; 13(12):1195. https://doi.org/10.3390/metabo13121195
Chicago/Turabian StyleCui, Baolu, Min Yu, Jiaojiao Bai, and Zhiguo Zhu. 2023. "SlbHLH22-Induced Hypertrophy Development Is Related to the Salt Stress Response of the GTgamma Gene in Tomatoes" Metabolites 13, no. 12: 1195. https://doi.org/10.3390/metabo13121195