Brain Metabolomics Reveal the Antipyretic Effects of Jinxin Oral Liquid in Young Rats by Using Gas Chromatography–Mass Spectrometry

Abstract
:1. Introduction
2. Results
2.1. Antipyretic and Anti-Inflammation Effects of IB and JXOL
2.2. Brain Metabolomics Profile and Multivariate Data Analysis
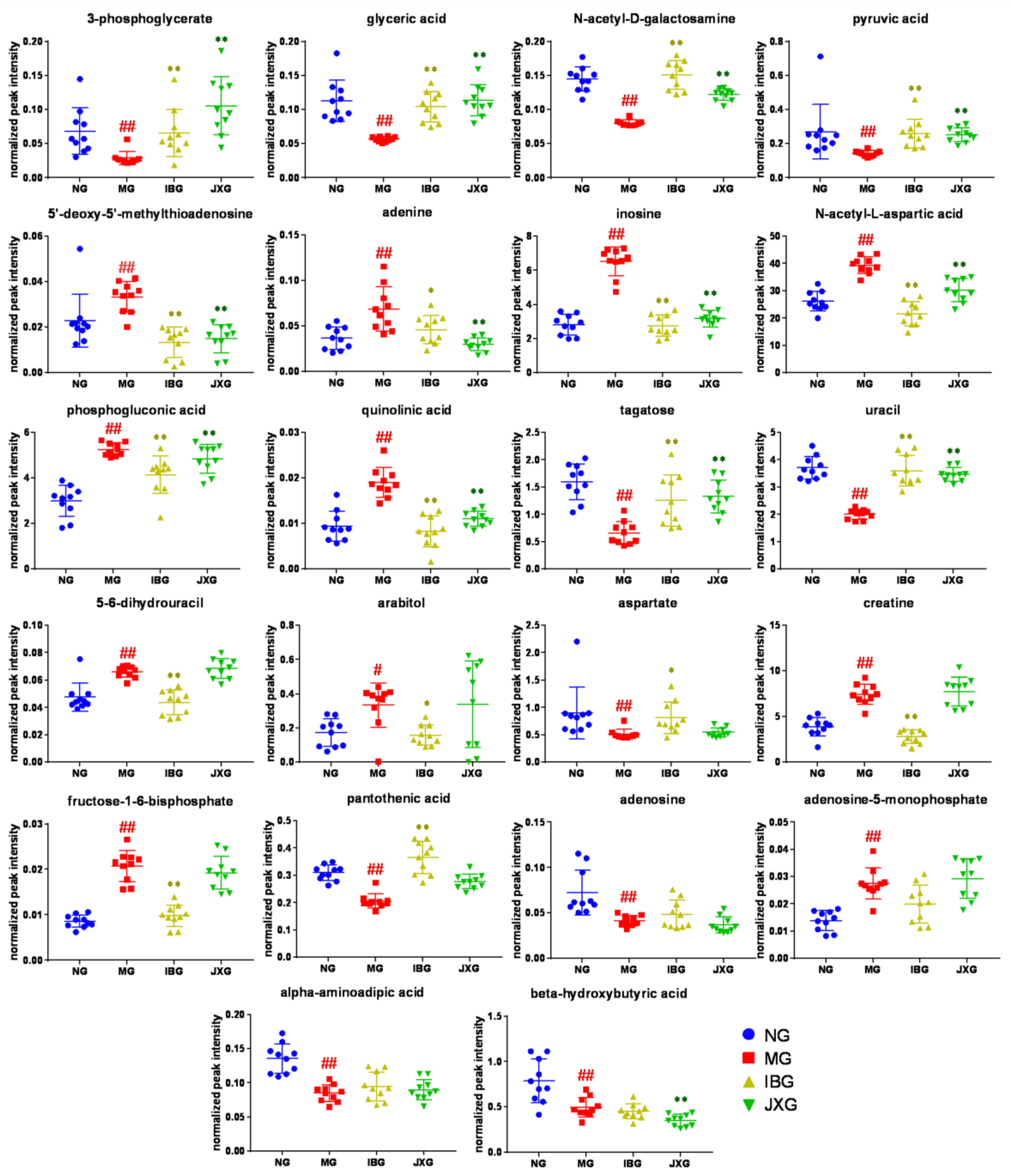
2.3. Identification of Potential Biomarkers and the Changing Trends Among NG, MG, IBG, and JXG
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals and Model Construction
4.3. Sample Collection and Cytokines
4.4. Sample Preparation and GC-MS Analysis
4.4.1. Tissue Extraction
4.4.2. GC-MS Conditions
5. Data Processing and Statistical Analysis
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Khalil, S.N.; Hahn, B.J.; Chumpitazi, C.E.; Rock, A.D.; Kaelin, B.A.; Macias, C.G. A multicenter, randomized, open-label, active-comparator trial to determine the efficacy, safety, and pharmacokinetics of intravenous ibuprofen for treatment of fever in hospitalized pediatric patients. BMC Pediatr. 2017, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Zhang, Z.; Guo, M.; Zhang, Q.; Wang, Q.; Lu, Z.; Zhao, H.; Liu, Y.; Fu, S.; Wang, M.; et al. Plasma metabolomics combined with lipidomics profiling reveals the potential antipyretic mechanisms of qingkailing injection in a rat model. Chem.-Biol. Interact. 2016, 254, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Guo, M.; Li, Q.; Peng, L.; Liu, H.; Zhang, L.; Bai, X.; Wang, Y.; Li, J.; Cai, C. Plasma metabolomic profiling to reveal antipyretic mechanism of shuang-huang-lian injection on yeast-induced pyrexia rats. PLoS ONE 2014, 9, e100017. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Huang, Q.; Shan, J.J.; Duan, J.A.; Zhu, Z.H.; Liu, P.; Bian, Y.; Shang, E.X.; Qian, D.W. Metabolomics of the antipyretic effects of bubali cornu (water buffalo horn) in rats. PLoS ONE 2016, 11, e0158478. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Han, X.M.; Li, J.Q.; Ren, X.Z.; Da, Q.G.; Guo, L.W.; Liu, T.S. Peparation of Qingfei Oral Liquid and Preparation Method Treating Viral Pneumoniain Children. China Patent ZL03131642.5, 19 November 2003. [Google Scholar]
- Shan, J.J.; Du, L.N.; Xu, J.Y.; Xu, S.; Di, L.Q.; Wang, S.C. Simultaneous determination of eight components in Jinxin Oral Liquid by UPLC. Chin. Tradit. Herb. Drugs 2013, 44, 183–185. [Google Scholar]
- Shan, J.J.; Deng, Y.T.; Yu, J.H.; Xi, X.P.; Du, L.N.; Xu, J.Y.; Xie, T.; Wang, S.C. Research on anti-inflammatory, expectorant and antipyretic effects of different polar fractions of Jinxin oral liquid. J. Nanjing Univ. Tradit. Chin. Med. 2012, 28, 464–466. [Google Scholar]
- Du, L.N.; Xie, T.; Xu, J.Y.; Kang, A.; Di, L.Q.; Shan, J.J.; Wang, S.C. A metabolomics approach to studying the effects of Jinxin oral liquid on rsv-infected mice using uplc/ltq-orbitrap mass spectrometry. J. Ethnopharmacol. 2015, 174, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.G.; Luo, H.; Wang, S.C.; Xu, J.Y.; Li, J.X. Antiviral effects of Jinxin oral liquid against respiratory syncytial virus infection in the balb/c mice model. J. Ethnopharmacol. 2015, 162, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; Caflisch, L.; Lodi, A.; Brenner, A.J.; Tiziani, S. Metabolomic signature of brain cancer. Mol. Carcinog. 2017, 56, 2355–2371. [Google Scholar] [CrossRef]
- Shen, C.S.; Zhang, Z.G.; Xie, T.; Xu, J.Y.; Yan, J.; Kang, A.; Dai, Q.G.; Wang, S.C.; Ji, J.J.; Shan, J.J. Jinxin oral liquid inhibits human respiratory syncytial virus-induced excessive inflammation associated with blockade of the nlrp3/asc/caspase-1 pathway. Biomed. Pharmacother. 2018, 103, 1376–1383. [Google Scholar] [CrossRef]
- Savolainen, O.I.; Sandberg, A.S.; Ross, A.B. A simultaneous metabolic profiling and quantitative multimetabolite metabolomic method for human plasma using gas-chromatography tandem mass spectrometry. J. Proteome Res. 2016, 15, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Dieme, B.; Lefevre, A.; Nadal-Desbarats, L.; Galineau, L.; Madji Hounoum, B.; Montigny, F.; Blasco, H.; Andres, C.R.; Emond, P.; Mavel, S. Workflow methodology for rat brain metabolome exploration using nmr, lc-ms and gc-ms analytical platforms. J. Pharm. Biomed. Anal. 2017, 142, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Lee, Y.H.; Kim, S.Y.; Kim, Y.G.; Moon, J.Y.; Jeong, K.H.; Lee, T.W.; Ihm, C.G.; Kim, S.; Kim, K.H.; et al. Systematic biomarker discovery and coordinative validation for different primary nephrotic syndromes using gas chromatography-mass spectrometry. J. Chromatogr. A 2016, 1453, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Li, W.W.; Yang, Y.; Dai, Q.G.; Lin, L.L.; Xie, T.; He, L.L.; Tao, J.L.; Shan, J.J.; Wang, S.C. Non-invasive urinary metabolomic profiles discriminate biliary atresia from infantile hepatitis syndrome. Metabolomics 2018, 14, 90. [Google Scholar] [CrossRef]
- Chan, E.C.; Pasikanti, K.K.; Nicholson, J.K. Global urinary metabolic profiling procedures using gas chromatography-mass spectrometry. Nat. Protoc. 2011, 6, 1483–1499. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Kang, A.; Peng, L.; Xie, T.; Ji, J.; Zhou, W.; Shan, J.; Di, L. Gas chromatography-mass spectrometry based plasma metabolomics of h1n1-induced inflammation in mice and intervention with flos lonicerae japonica-fructus forsythiae herb pair. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1092, 122–130. [Google Scholar] [CrossRef]
- Shan, J.; Peng, L.; Qian, W.; Xie, T.; Kang, A.; Gao, B.; Di, L. Integrated serum and fecal metabolomics study of collagen-induced arthritis rats and the therapeutic effects of the zushima tablet. Front. Pharmacol. 2018, 9, 891. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, D.; Li, X.; Li, Z.; Chao, J.; Qin, X. Metabolomic study of the fever model induced by baker’s yeast and the antipyretic effects of aspirin in rats using nuclear magnetic resonance and gas chromatography-mass spectrometry. J. Pharm. Biomed. Anal. 2013, 81–82, 168–177. [Google Scholar] [CrossRef]
- Gao, X.; Guo, M.; Peng, L.; Zhao, B.; Su, J.; Liu, H.; Zhang, L.; Bai, X.; Qiao, Y. Uplc q-tof/ms-based metabolic profiling of urine reveals the novel antipyretic mechanisms of qingkailing injection in a rat model of yeast-induced pyrexia. Evid.-Based Complement. Altern. Med. ECAM 2013, 2013, 864747. [Google Scholar] [CrossRef]
- Haley, M.J.; Lawrence, C.B. The blood-brain barrier after stroke: Structural studies and the role of transcytotic vesicles. J. Cereb. Blood Flow Metab. 2017, 37, 456–470. [Google Scholar] [CrossRef]
- Trotta, N.; Archambaud, F.; Goldman, S.; Baete, K.; Van, L.K.; Wens, V.; Van, B.P.; Chiron, C.; De, T.X. Functional integration changes in regional brain glucose metabolism from childhood to adulthood. Hum. Brain Mapp. 2016, 37, 3017–3030. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, D.C.; Brizuela, N.O.; Herrera, M.O.; Olguín, H.J.; Peraza, A.V.; Mejía, G.B. B-cyclodextrin and oleic acid increase levels of dopamine and potentiates oxidative damage in young and adult rat brain. Lipids Health Dis. 2018, 17, 172. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.P.; Pasin, J.S.; Saraiva, A.L.; Ratzlaff, V.; Rossato, M.F.; Andrighetto, R.; Rubin, M.A.; Ferreira, J.; Mello, C.F. N-acetylcysteine prevents baker’s-yeast-induced inflammation and fever. Inflamm. Res. 2012, 61, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Micheli, V.; Camici, M.; Tozzi, M.G.; Ipata, P.L.; Sestini, S.; Bertelli, M.; Pompucci, G. Neurological disorders of purine and pyrimidine metabolism. Curr. Top. Med. Chem. 2011, 11, 923–947. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, V.; Tolle, M.; Vanholder, R.; Schonfelder, G.; van der Giet, M.; Henning, L.; Schluter, H.; Paul, M.; Zidek, W.; Jankowski, J. Uridine adenosine tetraphosphate: A novel endothelium-derived vasoconstrictive factor. Nat. Med. 2005, 11, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Murtuza, F.; Farrier, A.J.; Venkatesan, M.; Smith, R.; Khan, A.; Uzoigwe, C.E.; Chami, G. Is there an association between body temperature and serum lactate levels in hip fracture patients? Ann. R. Coll. Surg. Engl. 2015, 97, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Saliba, S.W.; Vieira, E.L.; Santos, R.P.; Candelario-Jalil, E.; Fiebich, B.L.; Vieira, L.B.; Teixeira, A.L.; de Oliveira, A.C. Neuroprotective effects of intrastriatal injection of rapamycin in a mouse model of excitotoxicity induced by quinolinic acid. J. Neuroinflamm. 2017, 14, 25. [Google Scholar] [CrossRef]
- Kalonia, H.; Kumar, P.; Kumar, A. Attenuation of proinflammatory cytokines and apoptotic process by verapamil and diltiazem against quinolinic acid induced huntington like alterations in rats. Brain Res. 2011, 1372, 115–126. [Google Scholar] [CrossRef]
- Hayflick, S.J. Defective pantothenate metabolism and neurodegeneration. Biochem. Soc. Trans. 2014, 42, 1063–1068. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Zhang, X.; Yue, B.; Ge, W.; Zhang, M.; Ma, C.; Kong, M. Effects of pantothenic acid on growth performance, slaughter performance, lipid metabolism, and antioxidant function of wulong geese aged one to four weeks. Anim. Nutr. 2016, 2, 312–317. [Google Scholar] [CrossRef]
- Cajka, T.; Fiehn, O. Lc-ms-based lipidomics and automated identification of lipids using the lipidblast in-silico ms/ms library. Methods Mol. Biol. 2017, 1609, 149–170. [Google Scholar] [PubMed]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. Ms-dial: Data-independent ms/ms deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef] [PubMed]



 represents brain samples of NG,
represents brain samples of NG,  represents brain samples of MG,
represents brain samples of MG,  represents brain samples of IBG, and
represents brain samples of IBG, and  represents brain samples of JXG. The scatter plots show the mean and SD of the metabolites. # p < 0.05, ## p < 0.01, vs. NG; * p < 0.05, ** p < 0.01, vs. MG.
represents brain samples of NG, represents brain samples of MG, represents brain samples of IBG, and represents brain samples of JXG. The scatter plots show the mean and SD of the metabolites. # p < 0.05, ## p < 0.01, vs. NG; * p < 0.05, ** p < 0.01, vs. MG.
represents brain samples of JXG. The scatter plots show the mean and SD of the metabolites. # p < 0.05, ## p < 0.01, vs. NG; * p < 0.05, ** p < 0.01, vs. MG.
represents brain samples of NG, represents brain samples of MG, represents brain samples of IBG, and represents brain samples of JXG. The scatter plots show the mean and SD of the metabolites. # p < 0.05, ## p < 0.01, vs. NG; * p < 0.05, ** p < 0.01, vs. MG.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | Metabolites | HMDB | KEGG | FC NG vs. MG | p Value: | ||
|---|---|---|---|---|---|---|---|
| MG vs. NG | IBG vs. MG | JXG vs. MG | |||||
| 1 | 3-phosphoglycerate | HMDB0000807 | C00597 | 0.455 | <0.0001 ## | 0.0040 ** | <0.0001 ** |
| 2 | 5-6-dihydrouracil | HMDB0000076 | C00429 | 1.506 | 0.0010 ## | <0.0001 ** | 0.8870 |
| 3 | 5′-deoxy-5′-methylthioadenosine | HMDB0001173 | C00170 | 1.705 | 0.0020 ## | <0.0001 ** | <0.0001 ** |
| 4 | adenine | HMDB0000034 | C00147 | 1.814 | 0.0080 ## | 0.0400 * | <0.0001 ** |
| 5 | adenosine | HMDB0000050 | C00212 | 0.658 | <0.0001 ## | 1.0000 | 0.1070 |
| 6 | adenosine-5-monophosphate | HMDB0000045 | C00020 | 1.734 | <0.0001 ## | 0.1380 | 0.6430 |
| 7 | alpha-aminoadipic acid | HMDB0000510 | C00956 | 0.623 | <0.0001 ## | 0.8180 | 0.8340 |
| 8 | arabitol | HMDB0000568 | C01904 | 1.939 | 0.0470 # | 0.0470 * | 0.4550 |
| 9 | aspartate | HMDB0000191 | C00049 | 0.589 | <0.0001 ## | 0.0400 * | 0.1390 |
| 10 | beta-hydroxybutyric acid | HMDB0000357 | C01089 | 0.631 | 0.0080 ## | 0.5000 | 0.0010 ** |
| 11 | creatine | HMDB0000064 | C00300 | 1.925 | <0.0001 ## | <0.0001 ** | 0.8870 |
| 12 | fructose-1-6-bisphosphate | HMDB0001058 | C00354 | 2.419 | <0.0001 ## | 0.0020 ** | 0.5190 |
| 13 | glyceric acid | HMDB0000139 | C00258 | 0.582 | <0.0001 ## | <0.0001 ** | <0.0001 ** |
| 14 | inosine | HMDB0000195 | C00294 | 2.273 | <0.0001 ## | <0.0001 ** | <0.0001 ** |
| 15 | N-acetyl-d-galactosamine | HMDB0000212 | C01074 | 0.566 | <0.0001 ## | <0.0001 ** | <0.0001 ** |
| 16 | N-acetyl-l-aspartic acid | HMDB0000812 | C01042 | 1.541 | <0.0001 ## | <0.0001 ** | <0.0001 ** |
| 17 | pantothenic acid | HMDB0000210 | C00864 | 0.673 | <0.0001 ## | 0.0020 ** | <0.0001 ** |
| 18 | phosphogluconic acid | HMDB0001316 | C00345 | 1.624 | <0.0001 ## | 0.0010 ** | 0.4300 |
| 19 | pyruvic acid | HMDB0000243 | C00022 | 0.576 | <0.0001 ## | <0.0001 ** | <0.0001 ** |
| 20 | quinolinic acid | HMDB0000232 | C03722 | 2.116 | <0.0001 ## | 0.0010 ** | <0.0001 ** |
| 21 | tagatose | HMDB0003418 | C00795 | 0.391 | <0.0001 ## | 0.0030 ** | <0.0001 ** |
| 22 | uracil | HMDB0000300 | C00106 | 0.588 | <0.0001 ## | 0.0020 ** | <0.0001 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, W.; Shan, J.; Shen, C.; Yang, R.; Xie, T.; Di, L. Brain Metabolomics Reveal the Antipyretic Effects of Jinxin Oral Liquid in Young Rats by Using Gas Chromatography–Mass Spectrometry. Metabolites 2019, 9, 6. https://doi.org/10.3390/metabo9010006
Qian W, Shan J, Shen C, Yang R, Xie T, Di L. Brain Metabolomics Reveal the Antipyretic Effects of Jinxin Oral Liquid in Young Rats by Using Gas Chromatography–Mass Spectrometry. Metabolites. 2019; 9(1):6. https://doi.org/10.3390/metabo9010006
Chicago/Turabian StyleQian, Wenjuan, Jinjun Shan, Cunsi Shen, Rui Yang, Tong Xie, and Liuqing Di. 2019. "Brain Metabolomics Reveal the Antipyretic Effects of Jinxin Oral Liquid in Young Rats by Using Gas Chromatography–Mass Spectrometry" Metabolites 9, no. 1: 6. https://doi.org/10.3390/metabo9010006
APA StyleQian, W., Shan, J., Shen, C., Yang, R., Xie, T., & Di, L. (2019). Brain Metabolomics Reveal the Antipyretic Effects of Jinxin Oral Liquid in Young Rats by Using Gas Chromatography–Mass Spectrometry. Metabolites, 9(1), 6. https://doi.org/10.3390/metabo9010006