Soluble Sugar and Lipid Readjustments in the Yarrowia lipolytica Yeast at Various Temperatures and pH

Abstract
:1. Introduction
2. Results
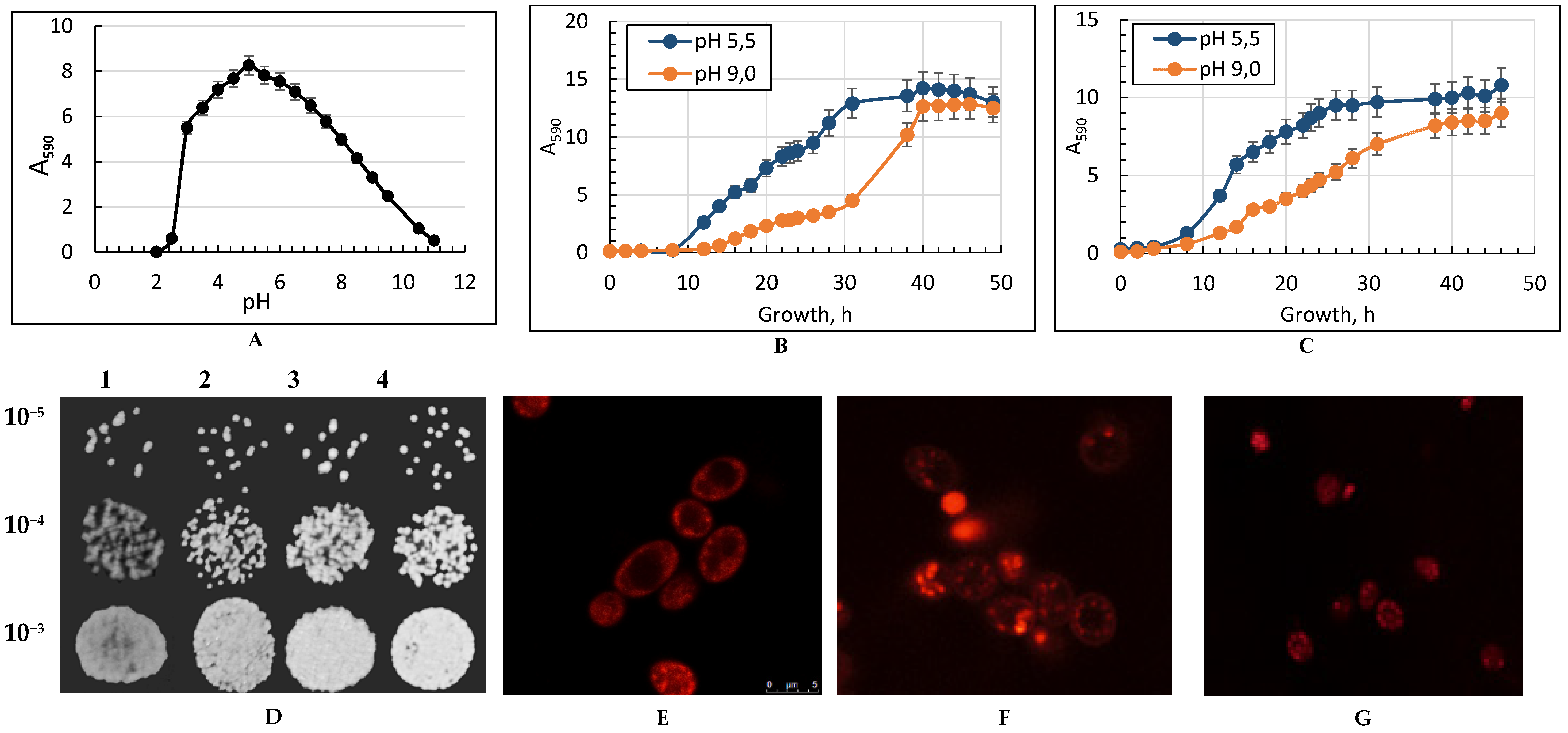
2.1. Growth of the Y. lipolytica Yeast under Different Conditions
2.2. Cell Viability and Vitality Assays
2.3. Cytosolic Soluble Carbohydrate Analysis
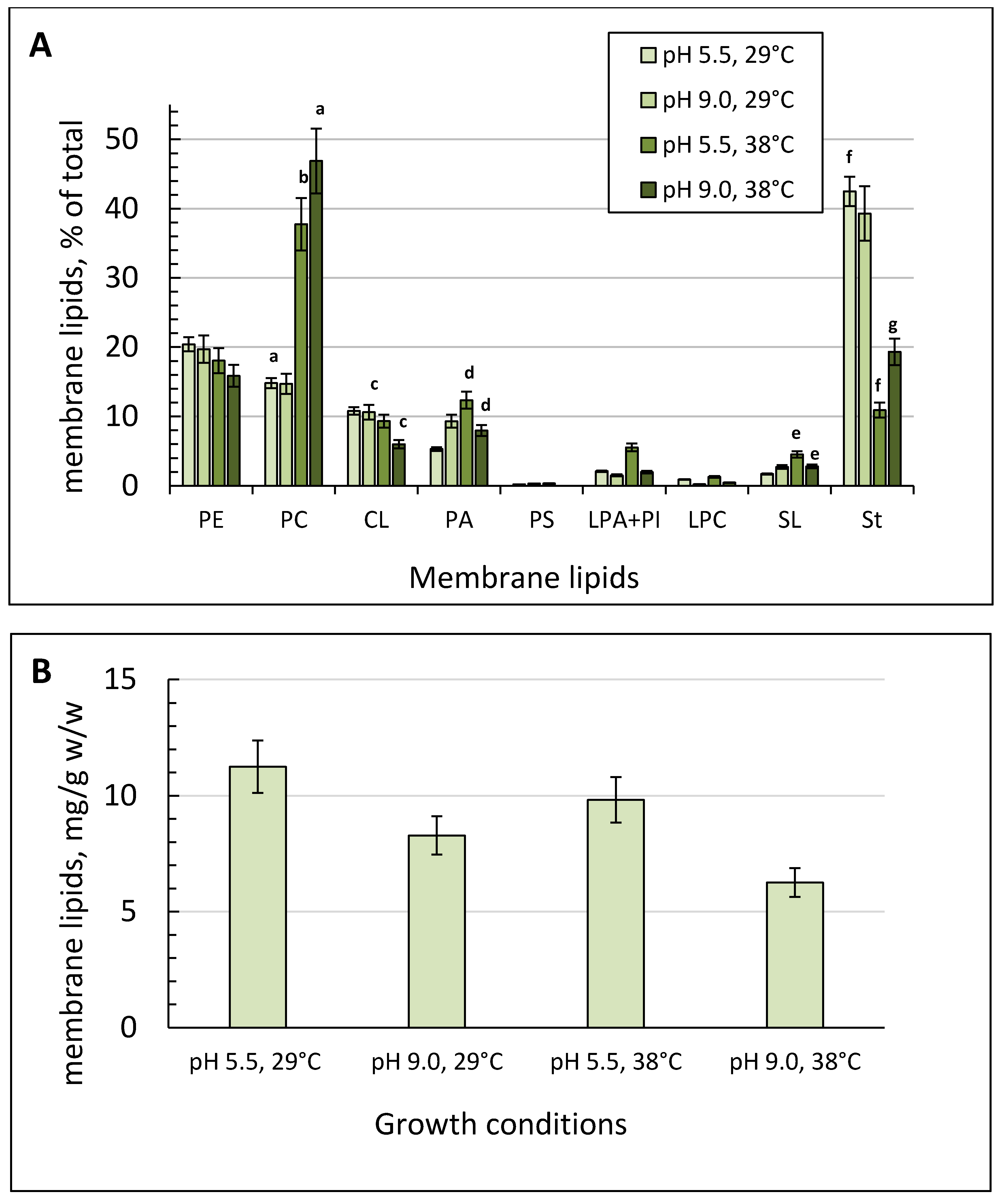
2.4. Membrane and Storage Lipids Profile at Various Ambient pH and Temperatures
2.5. Fatty Acids of the Main Phospholipids under Various Conditions
3. Discussion
3.1. Cytosol Carbohydrate Profile
3.2. Lipidome of Y. lipolytica
4. Conclusions
5. Materials and Methods
5.1. Yeast Strains and Growth Conditions
5.2. Cell Viability and Vitality Assays
5.2.1. Spotting Test
5.2.2. Staining with Methyl Blue
5.3. Potential-Dependent Staining
5.4. Transmission Electron Microscopy (TEM)
5.5. Preparation of Cellular Homogenate
5.6. Enzymes Activities Assay
5.7. Glutathione HPLC-ECD Analysis
5.8. Preparation and Analysis of Lipids
5.9. Carbohydrate Analysis
5.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zvyagilskaya, R.A.; Parkhomenko, O.A.; Gordeeva, A.V.; Deryabina, Y.I.; Persson, B.L. Bioenergetics of Yarrowia lipolytica cells grown at alkaline condition. Biosci. Rep. 2004, 24, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Egermeier, M.; Russmayer, H.; Sauer, M.; Marx, H. Metabolic flexibility of Yarrowia lipolytica growing on glycerol. Front. Microbiol. 2017, 8, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.H.; Ji, X.J.; Huang, H. Biotechnological applications of Yarrowia lipolytica: Past, present and future. Biotechnol. Adv. 2015, 33, 1522–1546. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Jackson, E.N. Metabolic engineering of Yarrowia lipolytica for industrial applications. Curr. Opin. Biotechnol. 2015, 36, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madzak, C. Engineering Yarrowia lipolytica for use in biotechnological applications: A review of major achievements and recent innovations. Mol. Biotechnol. 2018, 60, 621–635. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.P.; Cowen, L.E.; di Pietro, A.; Quinn, J. Stress Adaptation. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Iwata, H.; Yoshida, J.; Ogihara, J.; Kato, J.; Kasumi, T. Metabolic correlation between polyol and energy-storing carbohydrate under osmotic and oxidative stress condition in Moniliella megachiliensis. J. Biosci. Bioeng. 2015, 120, 405–410. [Google Scholar] [CrossRef]
- Eleutherio, E.; Panek, A.; De Mesquita, J.F.; Trevisol, E.; Magalhães, R. Revisiting yeast trehalose metabolism. Curr. Genet. 2015, 61, 263–274. [Google Scholar] [CrossRef]
- Patel, T.K.; Williamson, J.D. Mannitol in plants, fungi, and plant-fungal interactions. Trends Plant. Sci. 2016, 21, 486–497. [Google Scholar] [CrossRef]
- Sánchez-Fresneda, R.; Guirao-Abad, J.P.; Argüelles, A.; González-Párraga, P.; Valentín, E.; Argüelles, J.C. Specific stress-induced storage of trehalose, glycerol and D-arabitol in response to oxidative and osmotic stress in Candida albicans. Biochem. Biophys. Res. Commun. 2012, 430, 1334–1339. [Google Scholar] [CrossRef]
- Kayingo, G.; Kilian, S.G.; Prior, B.A. Conservation and release of osmolytes by yeasts during hypo-osmotic stress. Arch. Microbiol. 2001, 177, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewska, L.; Rakicka, M.; Rymowicz, W.; Rywińska, A. A comparative study on glycerol metabolism to erythritol and citric acid in Yarrowia lipolytica yeast cells. FEMS Yeast Res. 2014, 14, 966–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, P.S.; Waters, O.D.; Oliver, R.P. Decoding the mannitol enigma in filamentous fungi. Trends Microbiol. 2007, 15, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.; Prasad, V.; Zehra, A.; Gupta, V.K.; Upadhyay, R.S. Mannitol metabolism during pathogenic fungal-host interactions under stressed conditions. Front. Microbiol. 2015, 6, 1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vélëz, H.; Glassbrook, N.J.; Daub, M.E. Mannitol biosynthesis is required for plant pathogenicity by Alternaria alternata. FEMS Microbiol Lett. 2008, 285, 122–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voegele, R.T.; Hahn, M.; Lohaus, G.; Link, T.; Heiser, I.; Mendgen, K. Possible roles for mannitol and mannitol dehydrogenase in the biotrophic plant pathogen Uromyces fabae. Plant Physiol. 2005, 137, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Benaroudj, N.; Lee, D.H.; Goldberg, A.L. Trehalose accumulation during cellular stress protects cells and cellular proteins from damage by oxygen radicals. J. Biol. Chem. 2001, 276, 24261–24267. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.; Wiedemann, N.; Kammerer, B. Heat Stress-Induced Metabolic Remodeling in Saccharomyces cerevisiae. Metabolites 2019, 9, 266. [Google Scholar] [CrossRef] [Green Version]
- Flores, C.L.; Gancedo, C.; Petit, T. Disruption of Yarrowia lipolytica TPS1 gene encoding trehalose-6-P synthase does not affect growth in glucose but impairs growth at high temperature. PLoS ONE 2011, 6, e23695. [Google Scholar] [CrossRef] [Green Version]
- Bondarenko, S.A.; Ianutsevich, E.A.; Danilova, O.A.; Grum-Grzhimaylo, A.A.; Kotlova, E.R.; Kamzolkina, O.V.; Bilanenko, E.N.; Tereshina, V.M. Membrane lipids and soluble sugars dynamics of the alkaliphilic fungus Sodiomyces tronii in response to ambient pH. Extremophiles 2017, 21, 743–754. [Google Scholar] [CrossRef]
- Klose, C.; Surma, M.A.; Gerl, M.J.; Meyenhofer, F.; Shevchenko, A.; Simons, K. Flexibility of a eukaryotic lipidome-insights from yeast lipidomics. PLoS ONE 2012, 7, e35063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, C.M.; Block, D.E. Examining the role of membrane lipid composition in determining the ethanol tolerance of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2014, 80, 2966–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, B.; Schmidt, C.; Daum, G. Storage lipids of yeasts: A survey of nonpolar lipid metabolism in Saccharomyces cerevisiae, Pichia pastoris, and Yarrowia lipolytica. FEMS Microbiol. 2014, 38, 892–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodedová, M.; Sychrová, H. Changes in the Sterol Composition of the Plasma Membrane Affect Membrane Potential, Salt Tolerance and the Activity of Multidrug Resistance Pumps in Saccharomyces cerevisiae. PLoS ONE 2015, 10, e0139306. [Google Scholar] [CrossRef] [Green Version]
- Carman, G.M.; Han, G.S. Regulation of phospholipid synthesis in the yeast Saccharomyces cerevisiae. Annu. Rev. Biochem. 2011, 80, 859–883. [Google Scholar] [CrossRef]
- Breslow, D.K.; Weissman, J.S. Membranes in balance: Mechanisms of sphingolipid homeostasis. Mol. Cell 2010, 40, 267–279. [Google Scholar] [CrossRef] [Green Version]
- Henry, S.A.; Kohlwein, S.D.; Carman, G.M. Metabolism and regulation of glycerolipids in the yeast Saccharomyces cerevisiae. Genetics 2012, 190, 317–349. [Google Scholar] [CrossRef] [Green Version]
- Gálvez-López, D.; Chávez-Meléndez, B.; Vázquez-Ovando, A.; Rosas-Quijano, R. The metabolism and genetic regulation of lipids in the oleaginous yeast Yarrowia lipolytica. Braz. J. Microbiol. 2019, 50, 23–31. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S.; Bzducha-Wróbel, A.; Kot, A.M. Effect of selenium on lipid and amino acid metabolism in yeast cells. Biol. Trace Elem. Res. 2019, 187, 316–327. [Google Scholar] [CrossRef] [Green Version]
- Tezaki, S.; Iwama, R.; Kobayashi, S.; Shiwa, Y.; Yoshikawa, H.; Ohta, A.; Horiuchi, H.; Fukuda, R. Δ12-fatty acid desaturase is involved in growth at low temperature in yeast Yarrowia lipolytica. Biochem. Biophys. Res. Commun. 2017, 488, 165–170. [Google Scholar] [CrossRef]
- Cordova, L.T.; Alper, H.S. Production of α-linolenic acid in Yarrowia lipolytica using low temperature fermentation. Apple Microbiol. Biotechnol. 2018, 102, 8809–8816. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-L.; Madzak, C.; Liu, H.-H.; Song, P.; Ren, L.-J.; Huang, H.; Ji, X.-J. Engineering Yarrowia lipolytica for efficient γ-linolenic acid production. Biochem. Eng. J. 2017, 117, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Moi, I.M.; Leow, A.T.C.; Ali, M.S.M.; Rahman, R.; Salleh, A.B.; Sabri, S. Polyunsaturated fatty acids in marine bacteria and strategies to enhance their production. Appl. Microbiol. Biotechnol. 2018, 102, 5811–5826. [Google Scholar] [CrossRef] [PubMed]
- Grillitsch, K.; Connerth, M.; Köfeler, H.; Arrey, T.N.; Rietschel, B.; Wagner, B.; Karas, M.; Daum, G. Lipid particles/droplets of the yeast Saccharomyces cerevisiae revisited: Lipidome meets proteome. Biochim. Biophys. Acta 2011, 1811, 1165–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicaud, J.M. Yarrowia lipolytica. Yeast 2012, 29, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.A.G.; Colen, G.; Takahashi, J.A. Yarrowia lipolytica and its multiple applications in the biotechnological industry. Sci. World J. 2014. [Google Scholar] [CrossRef] [Green Version]
- Grum-Grzhimaylo, O.A.; Debets, A.J.; Bilanenko, E.N. The diversity of microfungi in peatlands originated from the White Sea. Mycologia 2016, 108, 233–254. [Google Scholar] [CrossRef]
- Augstein, A.; Barth, K.; Gentsch, M.; Kohlwein, S.D.; Barth, G. Characterization, localization and functional analysis of Gpr1 p, a protein affecting sensitivity to acetic acid in the yeast Yarrowia lipolytica. Microbiology 2003, 149, 589–600. [Google Scholar] [CrossRef] [Green Version]
- Fickers, P.; Fudalej, F.; Nicaud, J.M.; Destain, J.; Thonart, P. Selection of new over-producing derivatives for the improvement of extracellular lipase production by the non-conventional yeast Yarrowia lipolytica. J. Biotechnol. 2005, 115, 379–386. [Google Scholar] [CrossRef]
- Sekova, V.Y.; Dergacheva, D.I.; Isakova, E.P.; Deryabina, Y.I.; Tereshina, V.M. Carbohydrate spectrum of extremophilic yeasts Yarrowia lipolytica under pH-stress. Mikrobiologiya 2018, 87, 173–180. [Google Scholar] [CrossRef]
- Yancey, P.H.; Siebenaller, J.F. Co-evolution of proteins and solutions: Protein adaptation versus cytoprotective micromolecules and their roles in marine organisms. J. Exp. Biol. 2015, 218, 1880–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walther, T.; Mtimet, N.; Alkim, C.; Vax, A.; Loret, M.O.; Ullah, A.; Gancedo, C.; Smits, G.J.; François, J.M. Metabolic phenotypes of Saccharomyces cerevisiae mutants with altered trehalose 6-phosphate dynamics. Biochem. J. 2013, 454, 227–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gancedo, C.; Flores, C.L. The importance of a functional trehalose biosynthetic pathway for the life of yeasts and fungi. FEMS Yeast Res. 2004, 4, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Conlin, L.K.; Nelson, H.C. The natural osmolyte trehalose is a positive regulator of the heat-induced activity of yeast heat shock transcription factor. Mol. Cell. Biol. 2007, 27, 1505–1515. [Google Scholar] [CrossRef] [Green Version]
- Sekova, V.Y.; Gessler, N.N.; Isakova, E.P.; Antipov, A.N.; Dergacheva, D.I.; Deryabina, Y.I.; Trubnikova, E.V. Redox status of extremophilic yeast Yarrowia lipolytica during adaptation to pH-stress. Prikl. Biokhim. Mikrobiol. 2015, 51, 570–577. [Google Scholar] [CrossRef]
- Koganti, S.; Kuo, T.M.; Kurtzman, C.P.; Smith, N.; Ju, L.K. Production of arabitol from glycerol: Strain screening and study of factors affecting production yield. Appl. Microbiol. Biotechnol. 2011, 90, 257–267. [Google Scholar] [CrossRef]
- Toyoda, T.; Ohtaguchi, K. Effect of temperature on D-arabitol production from lactose by Kluyveromyces lactis. J. Ind. Microbiol. Biotechnol. 2011, 38, 1179–1185. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S.; Bzducha-Wróbel, A.; Kot, A.M. Effect of selenium on growth and antioxidative system of yeast cells. Mol. Biol. Rep. 2019, 46, 1797–1808. [Google Scholar] [CrossRef] [Green Version]
- Gientka, I.; Gadaszewska, M.; Błażejak, S.; Kieliszek, M.; Bzducha-Wróbel, A.; Stasiak-Różańska, L.; Kot, A.M. Evaluation of lipid biosynthesis ability by Rhodotorula and Sporobolomyces strains in medium with glycerol. Eur. Food Res. Technol. 2017, 243, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Kot, A.M.; Błażejak, S.; Kieliszek, M.; Gientka, I.; Bryś, J. Simultaneous Production of Lipids and Carotenoids by the Red Yeast Rhodotorula from Waste Glycerol Fraction and Potato Wastewater. Appl. Biochem. Biotechnol. 2019, 189, 589–607. [Google Scholar] [CrossRef]
- Mattanna, P.; Dallé da Rosa, P.; Gusso, A.P.; Richards, N.S.; Valente, P. Enhancement of microbial oil production by alpha-linolenic acid producing Yarrowia lipolytica strains QU22 and QU137. Food Sci. Biotechnol. 2014, 23, 1929–1934. [Google Scholar] [CrossRef]
- Pascual-Ahuir, A.; Manzanares-Estreder, S.; Timón-Gómez, A.; Proft, M. Ask yeast how to burn your fats: Lessons learned from the metabolic adaptation to salt stress. Curr. Genet. 2017, 64, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Haines, T.H.; Dencher, N.A. Cardiolipin: A proton trap for oxidative phosphorylation. FEBS Lett. 2002, 528, 35–39. [Google Scholar] [CrossRef] [Green Version]
- Joshi, A.S.; Zhou, J.; Gohil, V.M.; Chen, S.; Greenberg, M.L. Cellular functions of cardiolipin in yeast. Biochim. Biophys. Acta 2009, 1793, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Rostovtseva, T.K.; Gurnev, P.A.; Chen, M.; Bezrukov, S.M. Membrane lipid composition regulates tubulin interaction with mitochondrial voltage-dependent anion channel. J. Biol. Chem. 2012, 287, 29589–29598. [Google Scholar] [CrossRef] [Green Version]
- Glatz, A.; Pilbat, A.M.; Németh, G.L.; Vince-Kontár, K.; Jósvay, K.; Hunya, Á.; Udvardy, A.; Gombos, I.; Péter, M.; Balogh, G.; et al. Involvement of small heat shock proteins, trehalose, and lipids in the thermal stress management in Schizosaccharomyces pombe. Cell Stress Chaperones 2016, 21, 327–333. [Google Scholar] [CrossRef] [Green Version]
- McMaster, C.R. From yeast to humans—Roles of the Kennedy pathway for phosphatidylcholine synthesis. FEBS Lett. 2018, 592, 1256–1272. [Google Scholar] [CrossRef] [Green Version]
- Niu, L.; Nomura, K.; Iwahashi, H.; Matsuoka, H.; Kawachi, S.; Suzuki, Y.; Tamura, K. Petit-high pressure carbon dioxide stress increases synthesis of S-adenosylmethionine and phosphatidylcholine in yeast Saccharomyces cerevisiae. Biophys. Chem. 2017, 231, 79–86. [Google Scholar] [CrossRef]
- Boumann, H.A.; Gubbens, J.; Koorengevel, M.C.; Oh, C.S.; Martin, C.E.; Heck, A.J.; Patton-Vogt, J.; Henry, S.A.; de Kruijff, B.; de Kroon, A.I. Depletion of phosphatidylcholine in yeast induces shortening and increased saturation of the lipid acyl chains: Evidence for regulation of intrinsic membrane curvature in a eukaryote. Mol. Biol. Cell 2006, 17, 1006–1017. [Google Scholar] [CrossRef]
- Ianutsevich, E.A.; Danilova, O.A.; Groza, N.V.; Kotlova, E.R.; Tereshina, V.M. Heat shock response of thermophilic fungi: Membrane lipids and soluble carbohydrates under elevated temperatures. Microbiology 2016, 162, 989–999. [Google Scholar] [CrossRef]
- Klose, C.; Tarasov, K. Profiling of Yeast Lipids by Shotgun Lipidomics. Methods Mol. Biol. 2016, 1361, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Arinbasarova, A.Y.; Biryukova, E.N.; Medentsev, A.G. Antistress systems of the yeast Yarrowia lipolitica. Prikl. Biokhim. Mikrobiol. 2015, 51, 122–131. [Google Scholar] [CrossRef]
- Epova, E.Y.; Balovneva, M.V.; Isakova, E.P.; Kudykina, Y.K.; Zylkova, M.V.; Deryabina, Y.I.; Shevelev, A.B. Expression System for Yarrowia lipolytica Based on a Promoter of the Mitochondrial Potential-dependent Porin VDAC Gene. Biotechnol. Bioprocess Eng. 2016, 21, 408–413. [Google Scholar] [CrossRef]
- Kwolek-Mirek, M.; Zadrag-Tecza, R. Comparison of methods used for assessing the viability and vitality of yeast cells. FEMS Yeast Res. 2014, 14, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- Deryabina, Y.; Isakova, E.; Antipov, A.; Saris, N.-E.L. The inhibitors of antioxidant cell enzymes induce permeability transition in yeast mitochondria. J. Bioenerg. Biomembr. 2013, 45, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Deryabina, Y.; Isakova, E.; Sekova, V.; Antipov, A.; Saris, N.-E.L. Inhibition of free radical scavenging enzymes affects mitochondrial membrane permeability transition during growth and aging of yeast cells. J. Bioenerg. Biomembr. 2014, 46, 479–492. [Google Scholar] [CrossRef]
- Nichols, B.W. Separation of the lipids of photosynthetic tissues: Improvements in analysis by thin-layer chromatography. Biochim. Biophys. Acta 1963, 70, 417–422. [Google Scholar] [CrossRef]
- Kates, M. Techniques of Lipidology: Isolation, Analysis, and Identification of Lipids, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 1986; p. 464. [Google Scholar]
- Benning, C.; Huang, Z.H.; Gage, D.A. Accumulation of a novel glycolipid and a betaine lipid in cells of Rhodobacter sphaeroides grown under phosphate limitation. Arch. Biochem. Biophys. 1995, 317, 103–111. [Google Scholar] [CrossRef]
- Somogyi, M. Determination of blood sugar. J. Biol. Chem. 1945, 160, 69–73. [Google Scholar]
- Brobst, K.M.; Tai, H. Determination of chlorohydrins in hydroxypropyl starch ethers. J. Assoc. Anal. Chem. 1971, 54, 1093–1094. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antioxidant Compound | Growth Conditions | |||
|---|---|---|---|---|
| pH 5.5; 29 °C | pH 9.0; 29 °C | pH 5.5; 38 °C | pH 9.0; 38 °C | |
| SODs * | 141 ± 11 a | 831 ± 48 a | 1725.48 ± 63 a | 1407.13 ± 55 a |
| CATs ** | 26.58 ± 5.67 b | 21.7 ± 1.78 | 2.33 × 107 ± 1.12 × 106 b | 4.59 × 105 ± 2.59 × 104 b |
| [GSH] † | 15.22 ± 1.31 c,d | 8.68 ± 0.75 c | 54.90 ± 4.22 d | 23.30 ± 3.15 |
| [GSSG] ‡ | 3.62 ± 0.25 e,f | 4.15 ± 0.56 | 36.70 ± 4.01 e | 11.39 ± 1.32 f |
| [GSSG]/[GSH] | ~0.24 | ~0.48 | ~0.67 | ~0.48 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sekova, V.Y.; Dergacheva, D.I.; Isakova, E.P.; Gessler, N.N.; Tereshina, V.M.; Deryabina, Y.I. Soluble Sugar and Lipid Readjustments in the Yarrowia lipolytica Yeast at Various Temperatures and pH. Metabolites 2019, 9, 307. https://doi.org/10.3390/metabo9120307
Sekova VY, Dergacheva DI, Isakova EP, Gessler NN, Tereshina VM, Deryabina YI. Soluble Sugar and Lipid Readjustments in the Yarrowia lipolytica Yeast at Various Temperatures and pH. Metabolites. 2019; 9(12):307. https://doi.org/10.3390/metabo9120307
Chicago/Turabian StyleSekova, Varvara Yu, Daria I. Dergacheva, Elena P. Isakova, Natalya N. Gessler, Vera M. Tereshina, and Yulia I. Deryabina. 2019. "Soluble Sugar and Lipid Readjustments in the Yarrowia lipolytica Yeast at Various Temperatures and pH" Metabolites 9, no. 12: 307. https://doi.org/10.3390/metabo9120307
APA StyleSekova, V. Y., Dergacheva, D. I., Isakova, E. P., Gessler, N. N., Tereshina, V. M., & Deryabina, Y. I. (2019). Soluble Sugar and Lipid Readjustments in the Yarrowia lipolytica Yeast at Various Temperatures and pH. Metabolites, 9(12), 307. https://doi.org/10.3390/metabo9120307