Dopamine D3 Receptor Heteromerization: Implications for Neuroplasticity and Neuroprotection


Abstract
1. Introduction
2. Structure and Function of D3 Receptor
3. D3 Receptor Heteromerization
4. Interaction of D3R and nAChR: Relevance for Neuroplasticity and Neuroprotection
4.1. The Nicotinic Acetylcholine Receptor
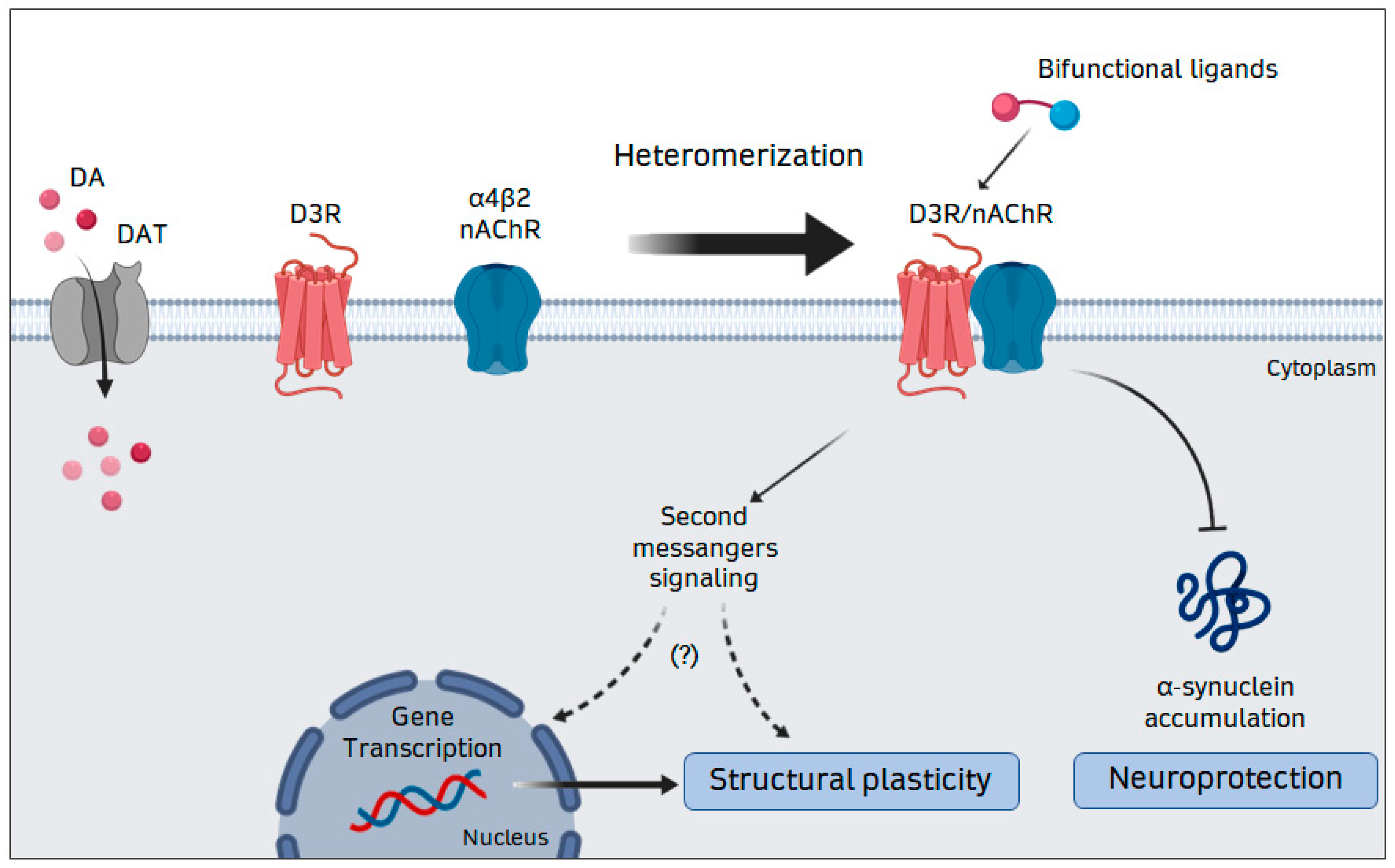
4.2. D3–nAChR Interactions
5. Therapeutic Implications of D3R-nAChR Heteromerization
6. Conclusions and Future Directions
Funding
Conflicts of Interest
References
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure to function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [CrossRef]
- Carli, M.; Kolachalam, S.; Aringhieri, S.; Rossi, M.; Giovannini, L.; Maggio, R.; Scarselli, M. Dopamine D2 Receptors Dimers: How can we Pharmacologically Target Them? Curr. Neuropharmacol. 2018, 16, 222–230. [Google Scholar] [CrossRef]
- Rangel-Barajas, C.; Coronel, I.; Florán, B. Dopamine Receptors and Neurodegeneration. Aging Dis. 2015, 6, 349–368. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, M.; Rabinowitz, A.; Singer, N.; Dagher, A. From apathy to addiction: Insights from neurology and psychiatry. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 101, 109926. [Google Scholar] [CrossRef] [PubMed]
- Diaz, J.; Lévesque, D.; Lammers, C.H.; Griffon, N.; Martres, M.P.; Schwartz, J.C.; Sokoloff, P. Phenotypical characterization of neurons expressing the dopamine D3 receptor in the rat brain. Neuroscience 1995, 65, 731–745. [Google Scholar] [CrossRef]
- Diaz, J.; Pilon, C.; Le Foll, B.; Gros, C.; Triller, A.; Schwartz, J.-C.; Sokoloff, P. Dopamine D3 receptors expressed by all mesencephalic dopamine neurons. J. Neurosci. 2000, 20, 8677–8684. [Google Scholar] [CrossRef] [PubMed]
- Lévesque, D.; Diaz, J.; Pilon, C.; Martres, M.P.; Giros, B.; Souil, E.; Schott, D.; Morgat, J.L.; Schwartz, J.C.; Sokoloff, P. Identification, characterization, and localization of the dopamine D3 receptor in rat brain using 7-[3H]hydroxy-N,N-di-n-propyl-2-aminotetralin. Proc. Natl. Acad. Sci. USA 1992, 89, 8155–8159. [Google Scholar] [CrossRef]
- Le Moine, C.; Bloch, B. Expression of the D3 dopamine receptor in peptidergic neurons of the nucleus accumbens: Comparison with the D1 and D2 dopamine receptors. Neuroscience 1996, 73, 131–143. [Google Scholar] [CrossRef]
- Sokoloff, P.; Giros, B.; Martres, M.P.; Bouthenet, M.L.; Schwartz, J.C. Molecular cloning and characterization of a novel dopamine receptor (D3) as a target for neuroleptics. Nature 1990, 347, 146–151. [Google Scholar] [CrossRef]
- Bouthenet, M.L.; Souil, E.; Martres, M.P.; Sokoloff, P.; Giros, B.; Schwartz, J.C. Localization of dopamine D3 receptor mRNA in the rat brain using in situ hybridization histochemistry: Comparison with dopamine D2 receptor mRNA. Brain Res. 1991, 564, 203–219. [Google Scholar] [CrossRef]
- Nicola, S.M.; Surmeier, J.; Malenka, R.C. Dopaminergic modulation of neuronal excitability in the striatum and nucleus accumbens. Ann. Rev. Neurosci. 2000, 23, 185–215. [Google Scholar] [CrossRef]
- Heidbreder, C. Selective antagonism at dopamine D3 receptors as a target for drug addiction pharmacotherapy: Are view of preclinical evidence. CNS Neurol. Disord. Drug Targets 2008, 7, 410–421. [Google Scholar] [CrossRef]
- Nakajima, S.; Gerretsen, P.; Takeuchi, H.; Caravaggio, F.; Chow, T.; Le Foll, B.; Mulsant, B.; Pollock, B.; Graff-Guerrero, A. The potential role of dopamine D3 receptor neurotransmission in cognition. Eur. Neuropsychopharmacol. 2013, 23, 799–813. [Google Scholar] [CrossRef]
- Marcellino, D.; Ferré, S.; Casadó, V.; Cortés, A.; Le Foll, B.; Mazzola, C.; Drago, F.; Saur, O.; Stark, H.; Soriano, A.; et al. Identification of dopamine D1–D3 receptor heteromers. Indications for a role of synergistic D1–D3 receptor interactions in the striatum. J. Biol. Chem. 2008, 283, 26016–26025. [Google Scholar] [CrossRef] [PubMed]
- Sokoloff, P.; Diaz, J.; LeFoll, B.; Guillin, O.; Leriche, L.; Bezard, E.; Gross, C. The dopamine D3 receptor: A therapeutic target for the treatment of neuropsychiatric disorders. CNS Neurol. Disord. Drug Targets 2006, 5, 25–43. [Google Scholar] [PubMed]
- De Mei, C.; Ramos, M.; Iitaka, C.; Borrelli, E. Getting specialized: Presynaptic and postsynaptic dopamine D2 receptors. Curr. Opin. Pharmacol. 2009, 9, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Bellucci, A.; Collo, G.; Sarnico, I.; Battistin, L.; Missale, C.; Spano, P. Alpha-synuclein accumulation and cell death triggered by energy deprivation and dopamine overload are counteracted by D2/D3 receptor activation. J. Neurochem. 2008, 106, 560–577. [Google Scholar] [CrossRef] [PubMed]
- Bono, F.; Savoia, P.; Guglielmi, A.; Gennarelli, M.; Piovani, G.; Sigala, S.; Leo, D.; Espinoza, S.; Gainetdinov, R.R.; Devoto, P.; et al. Role of Dopamine D2/D3 Receptors in Development, Plasticity, and Neuroprotection in Human iPSC-Derived Midbrain Dopaminergic Neurons. Mol. Neurobiol. 2018, 55, 1054–1067. [Google Scholar] [CrossRef]
- Bono, F.; Mutti, V.; Savoia, P.; Barbon, A.; Bellucci, A.; Missale, C.; Fiorentini, C. Nicotine prevents alpha-synuclein accumulation in mouse and human iPSC-derived dopaminergic neurons through activation of the dopamine D3- acetylcholine nicotinic receptor heteromer. Neurobiol. Dis. 2019, 129, 1–12. [Google Scholar] [CrossRef]
- Bontempi, L.; Savoia, P.; Bono, F.; Fiorentini, C.; Missale, C. Dopamine D3 and acetylcholine nicotinic receptor heteromerization in midbrain dopamine neurons: Relevance for neuroplasticity. Eur. Neuropsychopharmacol. 2017, 27, 313–324. [Google Scholar] [CrossRef]
- Collo, G.; Zanetti, S.; Missale, C.; Spano, P.F. Dopamine D3 receptor-preferring agonists increase dendrite arborisation of mesencephalic dopaminergic neurons via extracellular signal-regulated kinase phosphorylation. Eur. J. Neurosci. 2008, 28, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Li, R.; Huang, Y.; Xuping, L.; Le, W. Dopamine D3 receptor preferring agonists induce neurotrophic effects on mesencephalic dopamine neurons. Eur. J. Neurosci. 2005, 22, 2422–2430. [Google Scholar] [CrossRef] [PubMed]
- Van Kampen, J.M.; Eckman, C.B. Dopamine D3 receptor agonist delivery to a model of Parkinson’s disease restores the nigrostriatal pathway and improves locomotor behavior. J. Neurosci. 2006, 26, 7272–7280. [Google Scholar] [CrossRef] [PubMed]
- Levant, B. The D3 dopamine receptor: Neurobiology and potential clinical relevance. Pharmacol. Rev. 1997, 49, 231–252. [Google Scholar] [PubMed]
- Chien, E.Y.; Liu, W.; Zhao, Q.; Katritch, V.; Han, G.W.; Hanson, M.A.; Shi, L.; Newman, A.H.; Javitch, J.A.; Cherezov, V.; et al. Structure of the human dopamine D3 receptor in complex with a D2/D3 selective antagonist. Science 2010, 330, 1091–1095. [Google Scholar] [CrossRef]
- De Camilli, P.; Macconi, D.; Spada, A. Dopamine inhibits adenylate cyclase in human prolactin-secreting pituitary adenomas. Nature. 1979, 278, 252–254. [Google Scholar] [CrossRef]
- Gingrich, J.A.; Caron, M.G. Recent advances in the molecular biology of dopamine receptors. Annu. Rev. Neurosci. 1993, 16, 299–321. [Google Scholar] [CrossRef]
- Robinson, S.W.; Caron, M.G. Selective inhibition of adenylyl cyclase type V by the dopamine D3 receptor. Mol. Pharmacol. 1997, 52, 508–514. [Google Scholar] [CrossRef]
- Cussac, D.; Newman-Tancredi, A.; Pasteau, V.; Millan, M.J. Human dopamine D (3) receptors mediate mitogen-activated protein kinase activation via a phosphatidyl inositol 3-kinase and an atypical protein kinase C-dependent mechanism. Mol. Pharmacol. 1999, 56, 1025–1030. [Google Scholar] [CrossRef]
- Zapata, A.; Kivell, B.; Han, Y.; Javitch, J.A.; Bolan, E.A.; Kuraguntla, D.; Jaligam, V.; Oz, M.; Jayanthi, L.D.; Samuvel, D.J.; et al. Regulation of dopamine transporter function and cell surface expression by D3 dopamine receptors. J. Biol. Chem. 2007, 282, 35842–35854. [Google Scholar] [CrossRef]
- Collo, G.; Bono, F.; Cavalleri, L.; Plebani, L.; Merlo Pich, E.; Millan, M.J.; Spano, P.F.; Missale, C. Pre-synaptic dopamine D(3) receptor mediates cocaine-induced structural plasticity in mesencephalic dopaminergic neurons via ERK and Akt pathways. J. Neurochem. 2012, 120, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Jaworski, J.; Splanger, D.P.; Hoogenraad, C.C.; Sheng, M. Control of dendritic arborisation by the phosphoinositide-3’-kinase-Akt-mammalian target of rapamicin pathway. J. Neurosci. 2005, 25, 11300–11312. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Zhang, M.-X.; Swank, M.W.; Kunz, J.; Wu, G.-Y. Regulation of dendritic morphogenesis by Ras-PI3K-Akt-mTOR and Ras-MAPK signaling pathways. J. Neurosci. 2005, 25, 11288–11299. [Google Scholar] [CrossRef]
- Collo, G.; Bono, F.; Cavalleri, L.; Plebani, L.; Mitola, S.; Merlo Pich, E.; Millan, M.J.; Zoli, M.; Maskos, U.; Spano, P.; et al. Nicotine-induced structural plasticity in mesencephalic dopaminergic neurons is mediated by dopamine D3 receptors and Akt-mTORC1 signaling. Mol. Pharmacol. 2013, 83, 1176–1189. [Google Scholar] [CrossRef]
- Davila, V.; Yan, Z.; Craciun, L.C.; Logothetis, D.; Sulzer, D. D3 dopamine autoreceptors do not activate G-protein-gated inwardly rectifying potassium channel currents in substantia nigra dopamine neurons. J. Neurosci. 2003, 23, 5693–5697. [Google Scholar] [CrossRef]
- Van Kampen, J.M.; Robertson, H.A. A possible role for dopamine D3 receptor stimulation in the induction of neurogenesis in the adult rat substantia nigra. Neuroscience 2005, 136, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Cadoret, M.A.; Jaber, M.; Bloch, B. Prenatal D1, D1b and D3 dopamine receptor gene expression in the rat forebrain: Detection by reverse polymerase chain reaction. Neurosci. Lett. 1993, 155, 92–95. [Google Scholar] [CrossRef]
- Fishburn, C.S.; Bedford, M.; Lonai, P.; Fuchs, S. Early expression of D3 dopamine receptors in murine embryonic development. FEBS Lett. 1996, 381, 257–261. [Google Scholar] [CrossRef][Green Version]
- Diaz, J.; Ridray, S.; Mignon, V.; Griffon, N.; Schwartz, J.C.; Sokoloff, P. Selective expression of dopamine D3 receptor mRNA in proliferative zones during embryonic development of the rat brain. J. Neurosci. 1997, 17, 4282–4292. [Google Scholar] [CrossRef]
- Belinsky, G.S.; Sirois, C.L.; Rich, M.T.; Short, S.M.; Moore, A.R.; Gilbert, S.E.; Antic, S.D. Dopamine receptors in human embryonic stem cell neurodifferentiation. Stem Cells Dev. 2013, 22, 1522–1540. [Google Scholar] [CrossRef]
- Collo, G.; Cavalleri, L.; Bono, F.; Mora, C.; Fedele, S.; Invernizzi, R.W.; Gennarelli, M.; Piovani, G.; Kunath, T.; Millan, M.J.; et al. Ropinirole and Pramipexole Promote Structural Plasticity in Human iPSC-Derived Dopaminergic Neurons via BDNF and mTOR Signaling. Neural Plast. 2018, 2018, 4196961. [Google Scholar] [CrossRef] [PubMed]
- Van Kampen, J.M.; Hagg, T.; Robertson, H.A. Induction of neurogenesis in the adult rat subventricular zone and neostriatum following dopamine D3 receptor stimulation. Eur. J. Neurosci. 2004, 19, 2377–2387. [Google Scholar] [CrossRef]
- Kim, S.Y.; Choi, K.C.; Chang, M.S.; Kim, M.H.; Kim, S.Y.; Na, Y.S.; Lee, J.E.; Jin, B.K.; Lee, B.H.; Baik, J.H. The dopamine D2 receptor regulates the development of dopaminergic neurons via extracellularsignal-regulated kinase and Nurr1 activation. J. Neurosci. 2006, 26, 4567–4576. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Choi, M.H.; Chang, M.S.; Baik, J.H. Wnt5a-dopamine D2 receptor interactions regulate dopamine neuron development via extracellular signal-regulated kinase (ERK) activation. Biol. Chem. 2011, 286, 15641–15651. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Brito, I.; Romero-Fernandez, W.; Di Palma, M.; Oflijan, J.; Skieterska, K.; Duchou, J.; Van Craenenbroeck, K.; Suárez-Boomgaard, D.; Rivera, A.; et al. The G protein-coupled receptor heterodimer network (GPCR-HetNet) and its hub components. Int. J. Mol. Sci. 2014, 15, 8570–8590. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.Y.; Wess, J. Identification and molecular characterization of m3 muscarinic receptor dimers. J. Biol. Chem. 1999, 274, 19487–19497. [Google Scholar] [CrossRef]
- Hebert, T.E.; Moffet, S.; Morello, J.P.; Loisel, T.P.; Bichet, D.G.; Barret, C.; Bouvier, M. A peptide derived from a β2-adrenergic receptor transmembrane domain inhibits both receptor dimerization and activation. J. Biol. Chem. 1996, 271, 16384–16392. [Google Scholar] [CrossRef]
- White, J.H.; Wise, A.; Main, M.J.; Green, A.; Fraser, N.J.; Disney, G.H.; Barnes, A.A.; Emson, P.; Foord, S.M.; Marshall, F.H. Heterodimerization is required for the formation of a functional GABAB receptor. Nature 1998, 396, 679–682. [Google Scholar] [CrossRef]
- Kunishima, N.; Shimada, Y.; Tsuji, Y.; Sato, T.; Yamamoto, M. Structural basis of glutamate recognition by a dimeric metabotropic glutamate receptor. Nature 2000, 407, 971–977. [Google Scholar] [CrossRef]
- Scarselli, M.; Novi, F.; Schallmach, E.; Lin, R.; Baragli, A.; Colzi, A.; Griffon, N.; Corsini, G.U.; Sokoloff, P.; Levenson, R.; et al. D2/D3 dopamine receptor heterodimers exhibit unique functional properties. J. Biol. Chem. 2001, 276, 30308–30314. [Google Scholar] [CrossRef]
- Maggio, R.; Scarselli, M.; Capannolo, M.; Millan, M.J. Novel dimensions of D3 receptor function: Focus on heterodimerisation, transactivation and allosteric modulation. Eur. Neuropsychopharmacol. 2015, 25, 1470–1479. [Google Scholar] [CrossRef] [PubMed]
- Fiorentini, C.; Savoia, P.; Savoldi, D.; Missale, C. Receptor heteromers in Parkinson’s disease and L-DOPA-induced dyskinesia. CNS Neurol. Disord. Drug Targets 2013, 12, 1101–1113. [Google Scholar] [PubMed]
- Maggio, R.; Millan, M.J. DopamineD2–D3 receptor heteromers: Pharmacological properties and therapeutic significance. Curr. Opin. Pharmacol. 2010, 10, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Fiorentini, C.; Busi, C.; Gorruso, E.; Gotti, C.; Spano, P.F.; Missale, C. Reciprocal regulation of dopamine D1 and D3 receptor function and trafficking by heterodimerization. Mol. Pharmacol. 2008, 74, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Guitart, X.; Navarro, G.; Moreno, E.; Yano, H.; Cai, N.S.; Sánchez-Soto, M.; Kumar-Barodia, S.; Naidu, Y.T.; Mallol, J.; Cortés, A.; et al. Functional selectivity of allosteric interactions within G protein-coupled receptor oligomers: The dopamine D1–D3 receptor heterotetramer. Mol. Pharmacol. 2014, 86, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Guitart, X.; Moreno, E.; Rea, W.; Sánchez-Soto, M.; Cai, N.S.; Quiroz, C.; Kumar, V.; Bourque, L.; Cortés, A.; Canela, E.I.; et al. Biased G Protein-Independent Signaling of Dopamine D1-D3 Receptor Heteromers in the Nucleus Accumbens. Mol. Neurobiol. 2019, 56, 6756–6769. [Google Scholar] [CrossRef]
- Ferré, S.; Lluis, C.; Lanciego, J.L.; Franco, R. Prime time for G-protein-coupled receptor heteromers as therapeutic targets for CNS disorders: The dopamine D₁-D₃ receptor heteromer. CNS Neurol. Disord. Drug Targets 2010, 9, 596–600. [Google Scholar] [CrossRef]
- Bézard, E.; Ferry, S.; Mach, U.; Stark, H.; Leriche, L.; Boraud., T.; Gross, C.; Sokoloff, P. Attenuation of levodopa-induced dyskinesia by normalizing dopamine D3 receptor function. Nat. Med. 2003, 9, 762–767. [Google Scholar]
- Fiorentini, C.; Savoia, P.; Bono, F.; Tallarico, P.; Missale, C. The D3 dopamine receptor: From structural interactions to function. Eur. Neuropsychopharmacol. 2015, 25, 1462–1469. [Google Scholar] [CrossRef]
- Fiorentini, C.; Busi, C.; Spano, P.; Missale, C. Dimerization of dopamine D1 and D3 receptors in the regulation of striatal function. Curr. Opin. Pharmacol. 2010, 10, 87–92. [Google Scholar] [CrossRef]
- Fiorentini, C.; Savoia, P.; Savoldi, D.; Bono, F.; Busi, C.; Barbon, A.; Missale, C. Shp-2 knockdown prevents l-dopa-induced dyskinesia in a rat model of Parkinson’s disease. Mov. Disord. 2016, 31, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Fiorentini, C.; Savoia, P.; Savoldi, D.; Barbon, A.; Missale, C. Persistent activation of the D1R/Shp-2/Erk1/2 pathway in l-DOPA-induced dyskinesia in the 6-hydroxy-dopamine rat model of Parkinson’s disease. Neurobiol. Dis. 2013, 54, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Maggio, R.; Scarselli, M.; Novi, F.; Millan, M.J.; Corsini, G.U. Potent activation of dopamine D3/D2 heterodimers by the antiparkinsonian agents, S32504, pramipexole and ropinirole. J. Neurochem. 2003, 87, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Torvinen, M.; Marcellino, D.; Canals, M.; Agnati, L.F.; Lluis, C.; Franco, R.; Fuxe, K. Adenosine A2A receptor and dopamine D3 receptor interactions: Evidence of functional A2A/D3 heteromeric complexes. Mol. Pharmacol. 2005, 67, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Koschatzky, S.; Gmeiner, P. Selective agonists for dopamine/neurotensin receptor heterodimers. Chem. Med. Chem. 2012, 7, 509–514. [Google Scholar] [CrossRef]
- Zeng, C.; Asico, L.D.; Yu, C.; Villar, V.A.; Shi, W.; Luo, Y.; Wang, Z.; He, D.; Liu, Y.; Huang, L.; et al. Renal D3 dopamine receptor stimulation induces natriuresis by endothelin B receptor interactions. Kidney Int. 2008, 74, 750–759. [Google Scholar] [CrossRef]
- Reyes-Resina, I.; Alkozi, H.A.; Del Ser-Badia, A.; Sánchez-Naves, J.; Lillo, J.; Jiménez, J.; Pintor, J.; Navarro, G.; Franco, R. Expression of Melatonin and Dopamine D3 Receptor Heteromers in Eye Ciliary Body Epithelial Cells and Negative Correlation with Ocular Hypertension. Cells 2020, 9, 152. [Google Scholar] [CrossRef]
- Bouvier, M. Oligomerization of G-protein-coupled transmitter receptors. Nat. Rev. Neurosci. 2001, 2, 274–286. [Google Scholar] [CrossRef]
- Farré, D.; Muñoz, A.; Moreno, E.; Reyes-Resina, I.; Canet-Pons, J.; Dopeso-Reyes, I.G.; Rico, A.J.; Lluís, C.; Mallol, J.; Navarro, G.; et al. Stronger Dopamine D1 Receptor-Mediated Neurotransmission in Dyskinesia. Mol. Neurobiol. 2015, 52, 1408–1420. [Google Scholar] [CrossRef]
- Giros, B.; Caron, M.G. Molecular characterization of the dopamine transporter. Trends Pharmacol. Sci. 1993, 14, 43–49. [Google Scholar] [CrossRef]
- Giros, B.; Jaber, M.; Jones, S.R.; Wightman, R.M.; Caron, M.G. Hyperlocomotion and indifference to cocaine and amphetamine in mice lacking the dopamine transporter. Nature 1996, 379, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Quik, M.; Bordia, T.; Huang, L.; Perez, X. Targeting nicotinic receptors for Parkinson’s disease therapy. CNS Neurol. Disord. Drug Targets 2011, 10, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Séguéla, P.; Wadiche, J.; Dineley-Miller, K.; Dani, J.A.; Patrick, J.W. Molecular cloning, functional properties, and distribution of rat brain alpha 7: A nicotinic cation channel highly permeable to calcium. J. Neurosci. 1993, 13, 596–604. [Google Scholar] [CrossRef]
- Murray, T.A.; Bertrand, D.; Papke, R.L.; George, A.A.; Pantoja, R.; Srinivasan, R.; Liu, Q.; Wu, J.; Whiteaker, P.; Lester, H.A.; et al. α7β2 nicotinic acetylcholine receptors assemble, function, and are activated primarily via their α7-α7 interfaces. Mol. Pharmacol. 2012, 81, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Huang, Y.; Xue, F.; Simard, A.; De Chon, J.; Li, G.; Zhang, J.; Lucero, L.; Wang, M.; Sierks, M.; et al. A novel nicotinic acetylcholine receptor subtype in basal forebrain cholinergic neurons with high sensitivity to amyloid peptides. J. Neurosci. 2009, 29, 918–929. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.S.; Zwart, R.; Ursu, D.; Jensen, M.M.; Pinborg, L.H.; Gilmour, G.; Wu, J.; Sher, E.; Mikkelsen, J.D. α7 and β2 Nicotinic Acetylcholine Receptor Subunits Form Heteromeric Receptor Complexes that Are Expressed in the Human Cortex and Display Distinct Pharmacological Properties. PLoS ONE 2015, 10, e0130572. [Google Scholar] [CrossRef]
- Albuquerque, E.X.; Pereira, E.F.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef]
- Millar, N.S.; Gotti, C. Diversity of vertebrate nicotinic acetylcholine receptors. Neuropharmacology 2009, 56, 237–246. [Google Scholar] [CrossRef]
- Zoli, M.; Moretti, M.; Zanardi, A.; McIntosh, J.M.; Clementi, F.; Gotti, C. Identification of the nicotinic receptor subtypes expressed on dopaminergic terminals in the rat striatum. J. Neurosci. 2002, 22, 8785–8789. [Google Scholar] [CrossRef]
- Gotti, C.; Clementi, F.; Fornari, A.; Gaimarri, A.; Guiducci, S.; Manfredi, I.; Moretti, M.; Pedrazzi, P.; Pucci, L.; Zoli, M. Structural and functional diversity of native brain neuronal nicotinic receptors. Biochem. Pharmacol. 2009, 78, 703–711. [Google Scholar] [CrossRef]
- Benowitz, N.L. Pharmacology of nicotine: Addiction, smoking-induced disease, and therapeutics. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Gotti, C.; Clementi, F. Neuronal nicotinic receptors: From structure to pathology. Prog. Neurobiol. 2004, 74, 363–396. [Google Scholar] [CrossRef] [PubMed]
- Champtiaux, N.; Gotti, C.; Cordero-Erausquin, M.; David, D.J.; Przybylski, C.; Léna, C.; Clementi, F.; Moretti, M.; Rossi, F.M.; Le Novère, N.; et al. Subunit composition of functional nicotinic receptors in dopaminergic neurons investigated with knock-out mice. J. Neurosci. 2003, 23, 7820–7829. [Google Scholar] [CrossRef] [PubMed]
- Pidoplichko, V.I.; DeBiasi, M.; Williams, J.T.; Dani, J.A. Nicotine activates and desensitizes midbrain dopamine neurons. Nature 1997, 390, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Picciotto, M.R.; Zoli, M. Neuroprotection via nAChRs: The role of nAChRs in neurodegenerative disorders such as Alzheimer’s and Parkinson’s disease. Front. Biosci. 2008, 13, 492–504. [Google Scholar] [CrossRef]
- Mudo, G.; Belluardo, N.; Fuxe, K. Nicotinic receptor agonists as neuroprotective/neurotrophic drugs. Progress in molecular mechanisms. J. Neural Transm. 2007, 114, 135–147. [Google Scholar] [CrossRef]
- Grilli, M.; Zappettini, S.; Zoli, M.; Marchi, M. Pre-synaptic nicotinic and D receptors functionally interact on dopaminergic nerve endings of rat and mouse nucleus accumbens. J. Neurochem. 2009, 108, 1507–1514. [Google Scholar] [CrossRef]
- Quarta, D.; Ciruela, F.; Patkar, K.; Borycz, J.; Solinas, M.; Lluis, C.; Franco, R.; Wise, R.A.; Goldberg, S.R.; Hope, B.T.; et al. Heteromeric nicotinic acetylcholine- dopamine autoreceptor complexes modulate striatal dopamine release. Neuropsychopharmacology 2007, 32, 35–42. [Google Scholar] [CrossRef]
- Quik, M.; Parameswaran, N.; McCallum, S.E.; Bordia, T.; Bao, S.; Cormack, A.M.; Kim, A.; Tyndale, R.F. Chronic oral nicotine treatment protects against striatal degeneration in MPTP-treated primates. J. Neurochem. 2006, 98, 1866–1875. [Google Scholar] [CrossRef]
- Doura, M.B.; Luu, T.V.; Lee, N.H.; Perry, D.C. Persistent gene expression changes in ventral tegmental area of adolescent but not adult rats in response to chronic nicotine. Neuroscience 2010, 170, 503–513. [Google Scholar] [CrossRef]
- Bono, F.; Fiorentini, C. Exploring pre-degenerative alterations in humans using induced pluripotent stem cell-derived dopaminergic neurons. Neural Regen. Res. 2017, 12, 1068–1070. [Google Scholar] [CrossRef]
- Portoghese, P.S. Bivalent ligands and the message-address concept in the design of selective opioid receptor antagonists. Trends Pharmacol. Sci. 1989, 10, 230–235. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, Z.; Kan, Y. Receptor dimerization—Rationale for the design of bivalent ligands. Curr. Top. Med. Chem. 2007, 7, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Bonger, K.M.; van den Berg, R.J.; Knijnenburg, A.D.; Heitman, L.H.; Ijzerman, A.P.; Oosterom, J.; Timmers, C.M.; Overkleeft, H.S.; van der Marel, G.A. Synthesis and evaluation of homodimeric GnRHR antagonists having a rigid bis-propargylated benzene core. Bioorg. Med. Chem. 2008, 16, 3744–3758. [Google Scholar] [CrossRef]
- Saveanu, A.; Lavaque, E.; Gunz, G.; Barlier, A.; Kim, S.; Taylor, J.E.; Culler, M.D.; Enjalbert, A.; Jaquet, P. Demonstration of enhanced potency of a chimeric somatostatin-dopamine molecule, BIM-23A387, in suppressing growth hormone and prolactin secretion from human pituitary somatotroph adenoma cells. J. Clin. Endocrinol. Metab. 2002, 87, 5545–5552. [Google Scholar] [CrossRef] [PubMed]
- Waldhoer, M.; Fong, J.; Jones, R.M.; Lunzer, M.; Sharma, S.K.; Kostenis, E.; Portoghese, P.S.; Whistler, J.L. A heterodimer-selective agonist shows in vivo relevance of G protein-coupled receptor dimers. Proc. Natl. Acad. Sci. USA 2005, 102, 9050–9055. [Google Scholar] [CrossRef]
- Ansonoff, M.A.; Portoghese, P.S.; Pintar, J.E. Consequences of opioid receptor mutation on actions of univalent and bivalent kappa and delta ligands. Psychopharmacology 2010, 210, 161–168. [Google Scholar] [CrossRef]
- Bonuccelli, U.; Del Dotto, P.; Rascol, O. Role of dopamine receptor agonists in the treatment of early Parkinson’s disease. Parkinsonism Relat. Disord. 2009, 15 (Suppl. 4), S44–S53. [Google Scholar] [CrossRef]
- Fawcett, J.; Rush, A.J.; Vukelich, J.; Diaz, S.H.; Dunklee, L.; Romo, P.; Yarns, B.C.; Escalona, R. Clinical experience with high dose pramipexole in treatment resistant mood disorder patients. Am. J. Psychiatry 2016, 173, 107–111. [Google Scholar] [CrossRef]
- Fox, S.H.; Katzenschlager, R.; Lim, S.Y.; Ravina, B.; Seppi, K.; Coelho, M.; Poewe, W.; Rascol, O.; Goetz, C.G.; Sampaio, C. The Movement Disorder Society Evidence-Based Medicine Review Update: Treatments for the motor symptoms of Parkinson’s disease. Mov. Disord. 2011, 26 (Suppl. 3), S2–S41. [Google Scholar] [CrossRef]
- Schapira, A.H.; McDermott, M.P.; Barone, P.; Comella, C.L.; Albrecht, S.; Hsu, H.H.; Massey, D.H.; Mizuno, Y.; Poewe, W.; Rascol, O.; et al. Pramipexole in patients with early Parkinson’s disease (PROUD): A randomised delayed-start trial. Lancet Neurol. 2013, 12, 747–755. [Google Scholar] [CrossRef]
- Kalia, L.V.; Kalia, S.K.; Lang, A.E. Disease-modifying strategies for Parkinson’s disease. Mov. Disord. 2015, 30, 1442–1450. [Google Scholar] [CrossRef]
- Ma, C.; Liu, Y.; Neumann, S.; Gao, X. Nicotine from cigarette smoking and diet and Parkinson disease: A review. Transl. Neurodegener. 2017, 6, 18. [Google Scholar] [CrossRef]
- Matera, C.; Bono, F.; Pelucchi, S.; Collo, G.; Bontempi, L.; Gotti, C.; Zoli, M.; De Amici, M.; Missale, C.; Fiorentini, C.; et al. The novel hybrid agonist HyNDA-1 targets the D3R-nAChR heteromeric complex in dopaminergic neurons. Biochem. Pharmacol. 2019, 163, 154–168. [Google Scholar] [CrossRef]
- Joyce, J.N.; Millan, M.J. Dopamine D3 receptor agonists for protection and repair in Parkinson’s disease. Curr. Opin. Pharmacol. 2007, 7, 100–105. [Google Scholar] [CrossRef]
- Rosini, M. Polypharmacology: The rise of multitarget drugs over combination therapies. Future Med. Chem. 2014, 6, 485–487. [Google Scholar] [CrossRef]
- Corson, T.W.; Aberle, N.; Crews, C.M. Design and Applications of Bifunctional Small Molecules: Why Two Heads Are Better Than One. ACS Chem. Biol. 2008, 3, 677–692. [Google Scholar] [CrossRef]
- Costantino, L.; Barlocco, D. Designed multiple ligands: Basic research vs clinical outcomes. Curr. Med. Chem. 2012, 19, 3353–3387. [Google Scholar] [CrossRef]
- Achat-Mendes, C.; Grundt, P.; Cao, J.; Platt, D.M.; Newman, A.H.; Spealman, R.D. Dopamine D3 and D2 receptor mechanisms in the abuse-related behavioral effects of cocaine: Studies with preferential antagonists in squirrel monkeys. J. Pharmacol. Exp. Ther. 2010, 334, 556–565. [Google Scholar] [CrossRef]
- Heidbreder, C.A.; Andreoli, M.; Marcon, C.; Hutcheson, D.M.; Gardner, E.L.; Ashby, C.R., Jr. Evidence for the role of dopamine D3 receptors in oral operant alcohol self-administration and reinstatement of alcohol-seeking behavior in mice. Addict. Biol. 2007, 12, 35–50. [Google Scholar] [CrossRef]
- Higley, A.E.; Kiefer, S.W.; Li, X.; Gaál, J.; Xi, Z.X.; Gardner, E.L. Dopamine D(3) receptor antagonist SB-277011A inhibits methamphetamine self-administration and methamphetamine-induced reinstatement of drug-seeking in rats. Eur. J. Pharmacol. 2011, 659, 187–192. [Google Scholar] [CrossRef]
- Khaled, M.A.; Araki, K.F.; Li, B.; Coen, K.M.; Marinelli, P.W.; Varga, J.; Gaal, J.; Foll, B.L. The selective dopamine D3 receptor antagonist SB 277011-A, but not the partial agonist BP 897, blocks cue-induced reinstatement of nicotine-seeking. Int. J. Neuropsychopharmacol. 2010, 13, 181–190. [Google Scholar] [CrossRef]
- Spiller, K.; Xi, Z.X.; Peng, X.Q.; Newman, A.H.; Ashby, C.R., Jr.; Heidbreder, C.; Gaal, J.; Gardner, E.L. The selective dopamine D3 receptor antagonists SB-277011A and NGB 2904 and the putative partial D3 receptor agonist BP-897 attenuate methamphetamine-enhanced brain stimulation reward in rats. Psychopharmacology 2008, 196, 533–542. [Google Scholar] [CrossRef]
- Coe, J.W.; Brooks, P.R.; Vetelino, M.G.; Wirtz, M.C.; Arnold, E.P.; Huang, J.; Sands, S.B.; Davis, T.I.; Lebel, L.A.; Fox, C.B.; et al. Varenicline: An alpha4beta2 nicotinic receptor partial agonist for smoking cessation. J. Med. Chem. 2005, 48, 3474–3477. [Google Scholar] [CrossRef]
- Rollema, H.; Chambers, L.K.; Coe, J.W.; Glowa, J.; Hurst, R.S.; Lebel, L.A.; Lu, Y.; Mansbach, R.S.; Mather, R.J.; Rovetti, C.C.; et al. Pharmacological profile of the alpha4beta2 nicotinic acetylcholine receptor partial agonist varenicline, an effective smoking cessation aid. Neuropharmacology 2007, 52, 985–994. [Google Scholar] [CrossRef]
- Giulietti, F.; Filipponi, A.; Rosettani, G.; Giordano, P.; Iacoacci, C.; Spannella, F.; Sarzani, R. Pharmacological Approach to Smoking Cessation: An Updated Review for Daily Clinical Practice. High Blood Press. Cardiovasc. Prev. 2020, 23, 1–14. [Google Scholar] [CrossRef]
- Matera, C.; Pucci, L.; Fiorentini, C.; Fucile, S.; Missale, C.; Grazioso, G.; Clementi, F.; Zoli, M.; De Amici, M.; Gotti, C.; et al. Bifunctional compounds targeting both D2 and non-α7 nACh receptors: Design, synthesis and pharmacological characterization. Eur. J. Med. Chem. 2015, 101, 367–383. [Google Scholar] [CrossRef]


{kind=link}
| Receptor Complex | Detection Method | Suggested Relevance |
|---|---|---|
| DA receptors | ||
| D1R–D3R | FRET and BRET in transfected cell lines [14,54] | Parkinson’s disease [14,54,62] |
| CO-IP in transfected cell lines and rat striatum [54] | Addiction [56,62] | |
| PLA in rat and monkey striatum [69] | LID [54,56,62,69] | |
| D2R–D3R | CO-IP in transfected cell lines [50] | Parkinson’s disease [51,63] |
| Schizophrenia [51] | ||
| Other GPCRs | ||
| D3R–A2AR | FRET in transfected cell lines [64] | Parkinson’s disease [64] |
| Schizophrenia [64] | ||
| D3R–MT1R/MT2R | BRET in transfected cell lines [67] | Intraocular pressure [67] |
| PLA in human non-pigmented ciliary body epithelial cells [67] | ||
| Ion Channels | ||
| D3R-nAChR | BRET in transfected cell lines [20] | Parkinson’s disease [19,20] |
| PLA in mouse primary mesencephalic DA neurons and midbrain sections [20], and in hiPSC-derived DA neurons [19] | Addiction [20] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bono, F.; Mutti, V.; Fiorentini, C.; Missale, C. Dopamine D3 Receptor Heteromerization: Implications for Neuroplasticity and Neuroprotection. Biomolecules 2020, 10, 1016. https://doi.org/10.3390/biom10071016
Bono F, Mutti V, Fiorentini C, Missale C. Dopamine D3 Receptor Heteromerization: Implications for Neuroplasticity and Neuroprotection. Biomolecules. 2020; 10(7):1016. https://doi.org/10.3390/biom10071016
Chicago/Turabian StyleBono, Federica, Veronica Mutti, Chiara Fiorentini, and Cristina Missale. 2020. "Dopamine D3 Receptor Heteromerization: Implications for Neuroplasticity and Neuroprotection" Biomolecules 10, no. 7: 1016. https://doi.org/10.3390/biom10071016
APA StyleBono, F., Mutti, V., Fiorentini, C., & Missale, C. (2020). Dopamine D3 Receptor Heteromerization: Implications for Neuroplasticity and Neuroprotection. Biomolecules, 10(7), 1016. https://doi.org/10.3390/biom10071016