
AP-64, Encoded by C5orf46, Exhibits Antimicrobial Activity against Gram-Negative Bacteria

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Expression and Purification of Recombinant Proteins
2.3. Antibacterial Activity Detection of Purification Proteins
2.4. Antibacterial Activity of AP-64 and Gm94 against DH5α
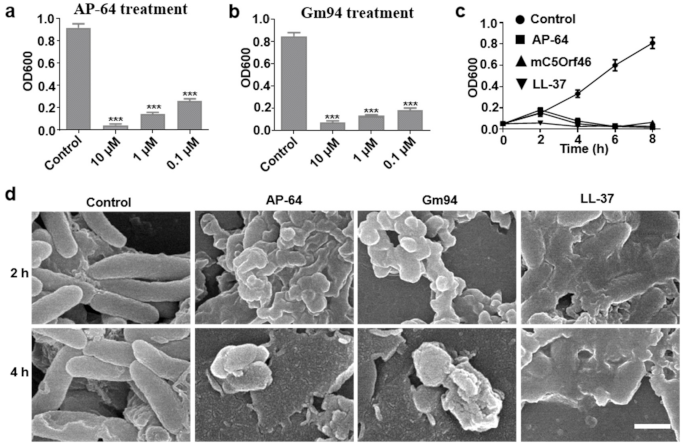
2.5. SEM Detection for DH5α Cells
2.6. Antibacterial Activity against Pathogenic Bacteria
2.7. Antibacterial Activity against Escherichia coli O157:H7 in Mouse
2.8. Antitumor Capacity
2.9. Statistical Analysis
3. Results
3.1. Amino Acid Sequence Features of AP-64/C5orf46
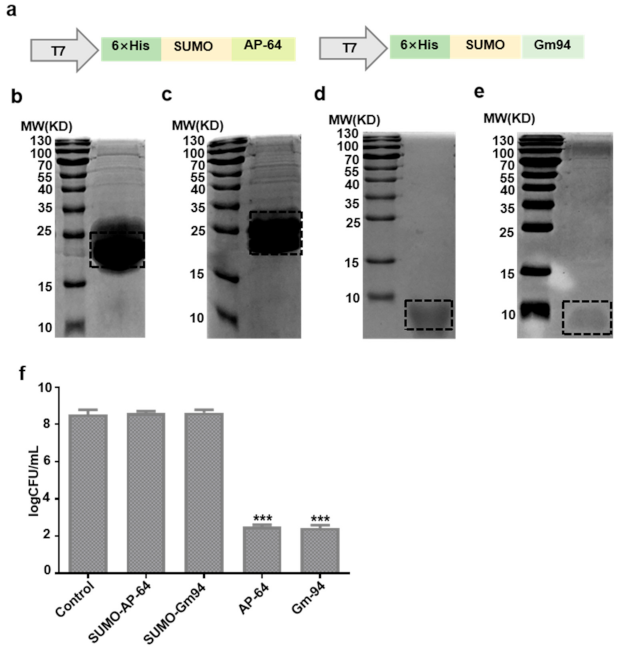
3.2. Expression and Purification of Recombinant Proteins
3.3. Antibacterial Activity against DH5α Cells
3.4. Antibacterial Activity against Pathogenic Bacteria
3.5. Antitumor Capacity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marston, H.D.; Dixon, D.M.; Knisely, J.M.; Palmore, T.N.; Fauci, A.S. Antimicrobial Resistance. JAMA 2016, 316, 1193–1220. [Google Scholar] [CrossRef] [PubMed]
- Gruenheid, S.; Le Moual, H. Resistance to antimicrobial peptides in Gram-negative bacteria. FEMS Microbiol. Lett. 2012, 330, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Taubes, G. The Bacteria Fight Back. Science 2008, 321, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L.; Maisetta, G.; Di Luca, M.; Gaddi, L.M.H.; Esin, S.; Florio, W.; Brancatisano, F.L.; Barra, D.; Campa, M.; Batoni, G. Comparative Analysis of the Bactericidal Activities of Amphibian Peptide Analogues against Multidrug-Resistant Nosocomial Bacterial Strains. Antimicrob. Agents Chemother. 2007, 52, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Radek, K.; Gallo, R. Antimicrobial peptides: Natural effectors of the innate immune system. Semin. Immunopathol. 2007, 29, 27–43. [Google Scholar] [CrossRef]
- Kavanagh, K.; Dowd, S. Histatins: Antimicrobial peptides with therapeutic potential. J. Pharm. Pharmacol. 2004, 56, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, L.; Maisetta, G.; Batoni, G.; Tavanti, A. Insights into the Antimicrobial Properties of Hepcidins: Advantages and Drawbacks as Potential Therapeutic Agents. Molecules 2015, 20, 6319–6341. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Dürr, U.H.; Sudheendra, U.; Ramamoorthy, A. LL-37, the only human member of the cathelicidin family of antimicrobial peptides. Biochim. Biophys. Acta BBA Biomembr. 2006, 1758, 1408–1425. [Google Scholar] [CrossRef] [PubMed]
- Iacob, D.G.; Iacob, S.A.J.P. Antibacterial Function of the Human Cathelicidin-18 Peptide (LL-37) between Theory and Practice. Protein Pept. Lett. 2014, 21, 1247–1256. [Google Scholar] [PubMed]
- Yang, M.; Tang, M.; Ma, X.; Yang, L.; He, J.; Peng, X.; Guo, G.; Zhou, L.; Luo, N.; Yuan, Z.; et al. AP-57/C10orf99 is a new type of mutifunctional antimicrobial peptide. Biochem. Biophys. Res. Commun. 2015, 457, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Harney, D.J.; Hutchison, A.T.; Su, Z.; Hatchwell, L.; Heilbronn, L.K.; Hocking, S.; James, D.E.; Larance, M. Small-protein Enrichment Assay Enables the Rapid, Unbiased Analysis of Over 100 Low Abundance Factors from Human Plasma. Mol. Cell. Proteom. 2019, 18, 1899–1915. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Xiang, Q.; Liu, H.; Qu, H.; Tang, X.; Xiao, X.; Zhang, Q.; Su, Z.; Huang, Y. Expression, purification, and biological activity of the recombinant pramlintide precursor. Appl. Microbiol. Biotechnol. 2014, 98, 7837–7844. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, Z.; Wu, H.; Lv, F. Study on the antibiotic activity of microcapsule curcumin against foodborne pathogens. Int. J. Food Microbiol. 2009, 136, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Sondi, I.; Salopek-Sondi, B. Silver nanoparticles as antimicrobial agent: A case study on E. coli as a model for Gram-negative bacteria. J. Colloid Interface Sci. 2004, 275, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Clinical Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; Approved Standard M07-A8; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Kim, D.S.; Kim, S.W.; Song, J.M.; Kim, S.Y.; Kwon, K.-C. A new prokaryotic expression vector for the expression of antimicrobial peptide abaecin using SUMO fusion tag. BMC Biotechnol. 2019, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, X.; Li, L.; Zhang, T.; Wu, F.; Wang, D.; Hu, H.; Tian, C.; Liao, D.; Zhao, L.; et al. Coagulation factors VII, IX and X are effective antibacterial proteins against drug-resistant Gram-negative bacteria. Cell Res. 2019, 29, 711–724. [Google Scholar] [CrossRef] [PubMed]
- GTEx Consortium. The Genotype-Tissue Expression (GTEx) pilot analysis: Multitissue gene regulation in humans. Science 2015, 348, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Skarnes, W.C.; Rosen, B.P.; West, A.P.; Koutsourakis, M.; Bushell, W.; Iyer, V.; Mujica, A.O.; Thomas, M.; Harrow, J.; Cox, T.; et al. A conditional knockout resource for the genome-wide study of mouse gene function. Nat. Cell Biol. 2011, 474, 337–342. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, K.; Wang, Y.; Wang, Z.; Zhang, Z.; Zhao, S.; Li, H.; Huang, J.; Guo, W.; Zheng, X.; Guo, G.; et al. AP-64, Encoded by C5orf46, Exhibits Antimicrobial Activity against Gram-Negative Bacteria. Biomolecules 2021, 11, 485. https://doi.org/10.3390/biom11040485
Zhong K, Wang Y, Wang Z, Zhang Z, Zhao S, Li H, Huang J, Guo W, Zheng X, Guo G, et al. AP-64, Encoded by C5orf46, Exhibits Antimicrobial Activity against Gram-Negative Bacteria. Biomolecules. 2021; 11(4):485. https://doi.org/10.3390/biom11040485
Chicago/Turabian StyleZhong, Kunhong, Yuelong Wang, Zeng Wang, Zongliang Zhang, Shasha Zhao, Hexian Li, Jianhan Huang, Wenhao Guo, Xi Zheng, Gang Guo, and et al. 2021. "AP-64, Encoded by C5orf46, Exhibits Antimicrobial Activity against Gram-Negative Bacteria" Biomolecules 11, no. 4: 485. https://doi.org/10.3390/biom11040485
APA StyleZhong, K., Wang, Y., Wang, Z., Zhang, Z., Zhao, S., Li, H., Huang, J., Guo, W., Zheng, X., Guo, G., Zhou, L., Yang, H., & Tong, A. (2021). AP-64, Encoded by C5orf46, Exhibits Antimicrobial Activity against Gram-Negative Bacteria. Biomolecules, 11(4), 485. https://doi.org/10.3390/biom11040485