
Description and Analysis of Glycosidic Residues in the Largest Open Natural Products Database


Abstract
1. Introduction
2. Methods
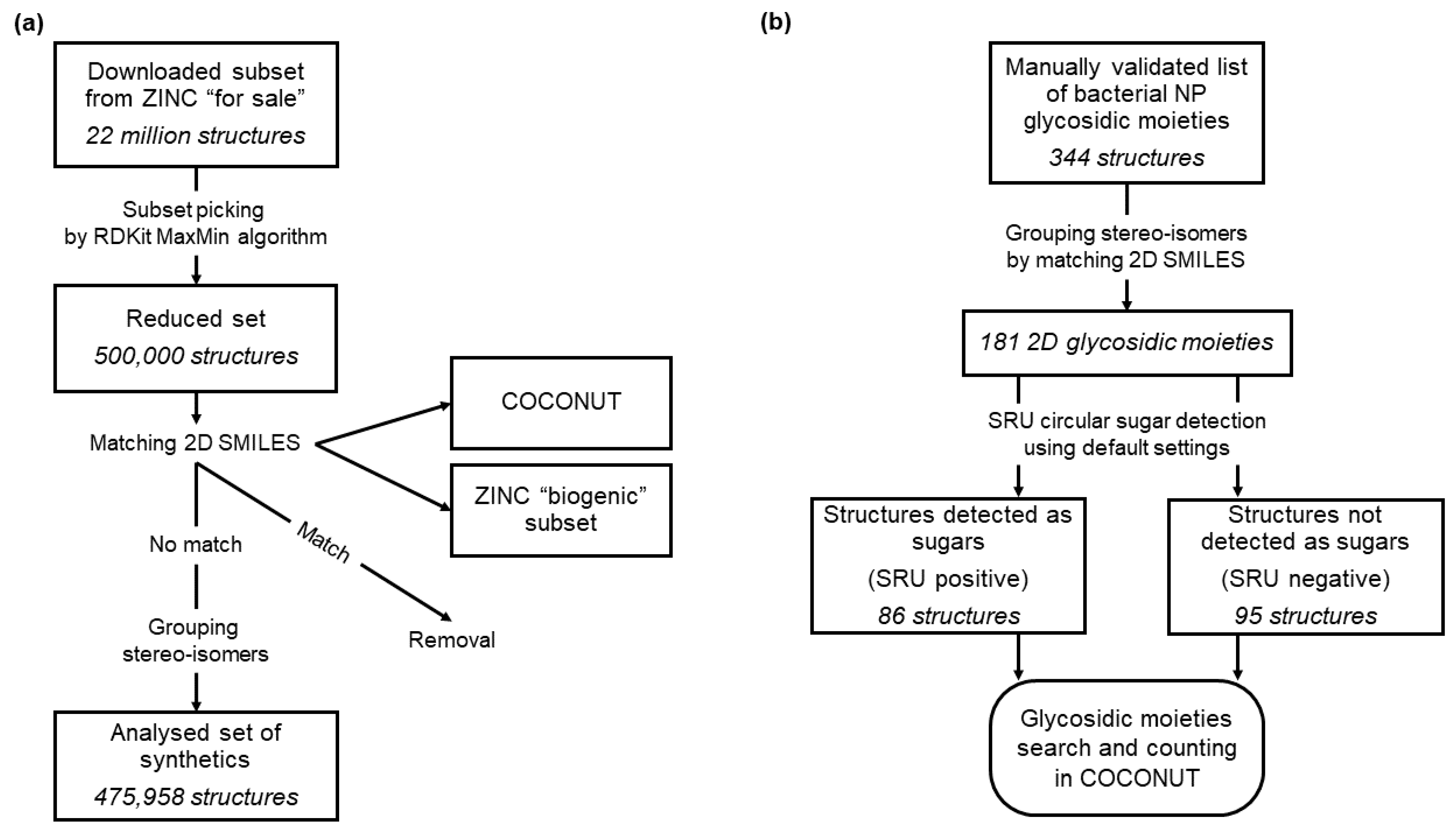
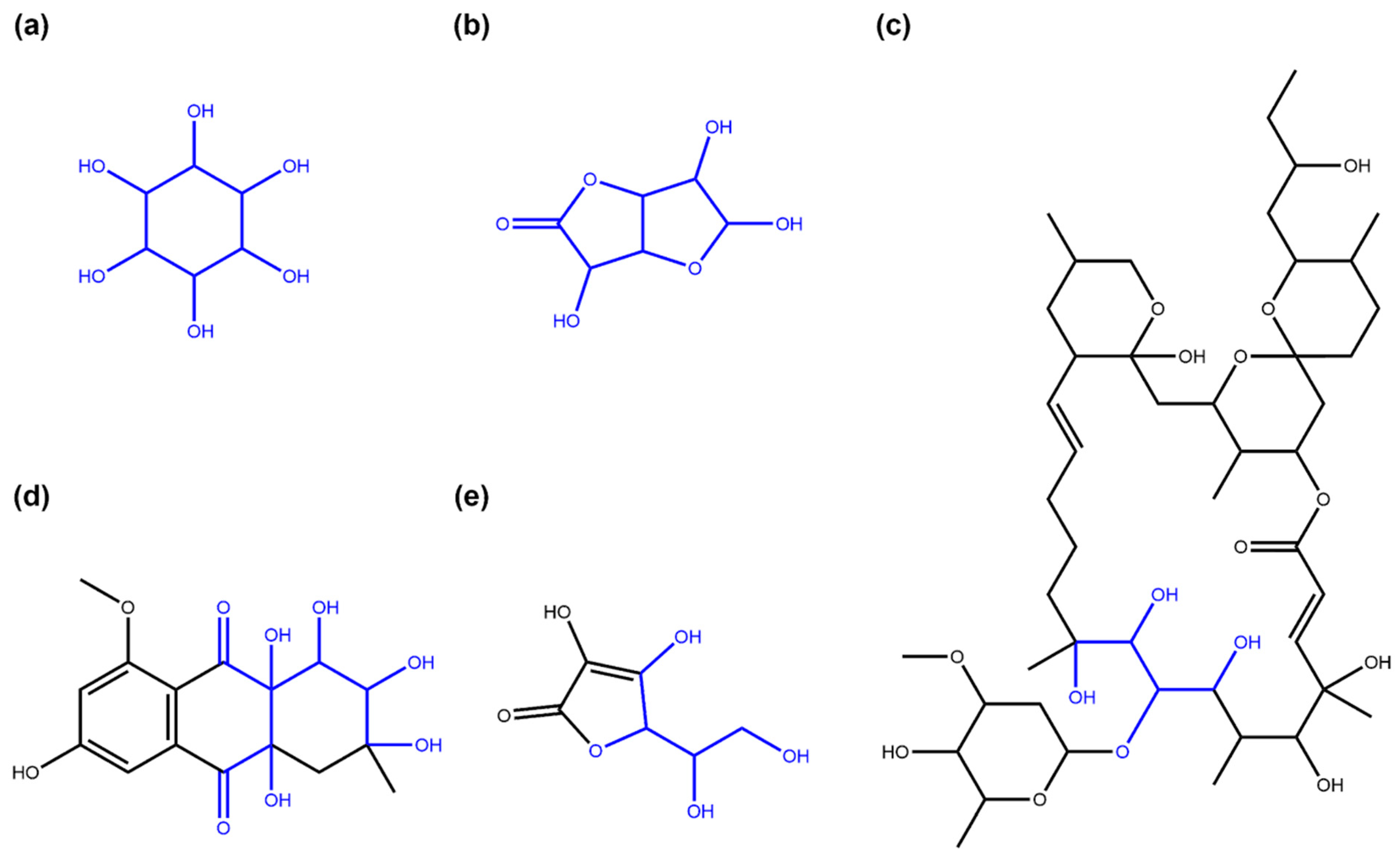
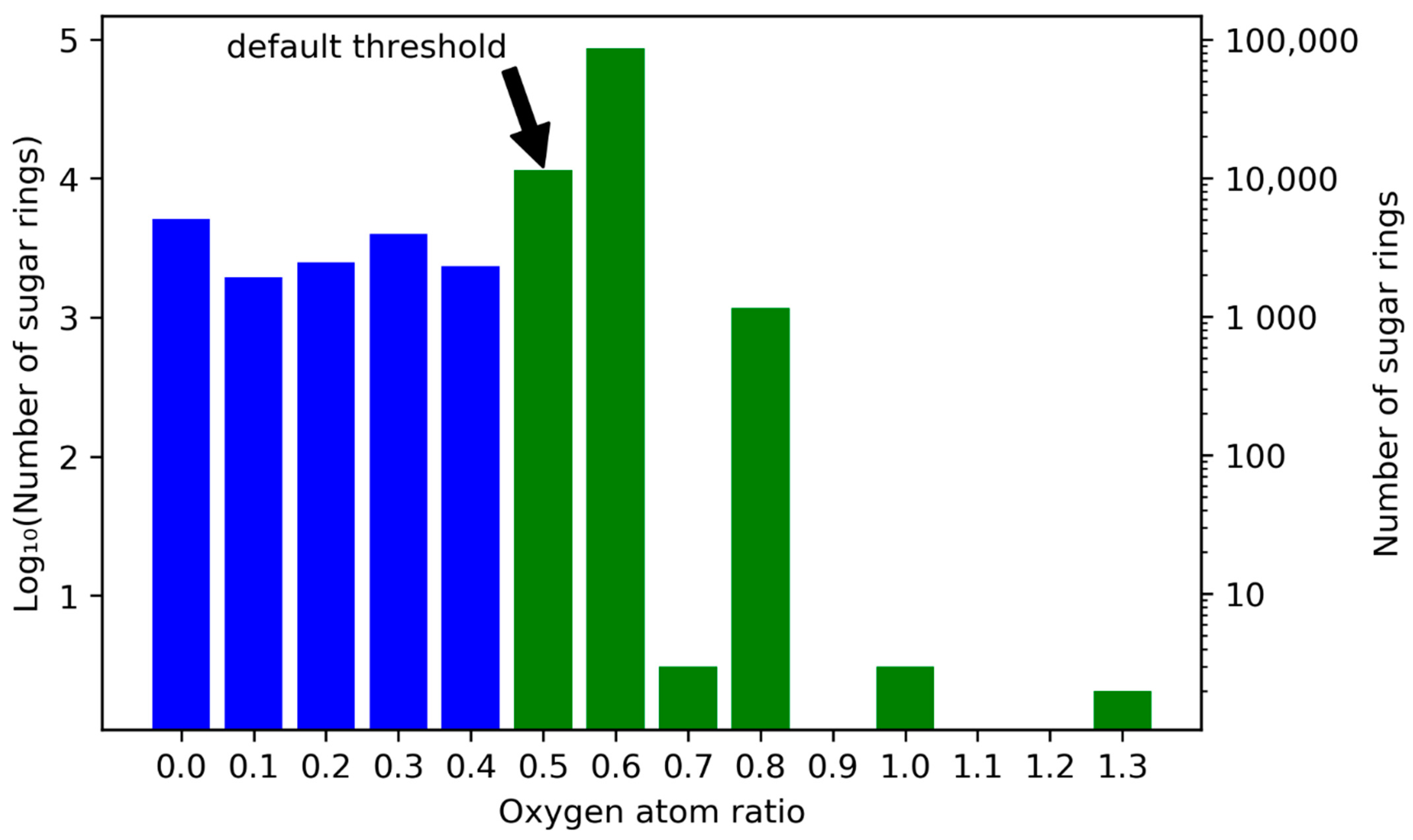
2.1. Detection of the Glycosidic Moieties
2.2. COCONUT
2.3. ZINC
2.4. Sugar Moieties in Bacterial Natural Products
3. Results
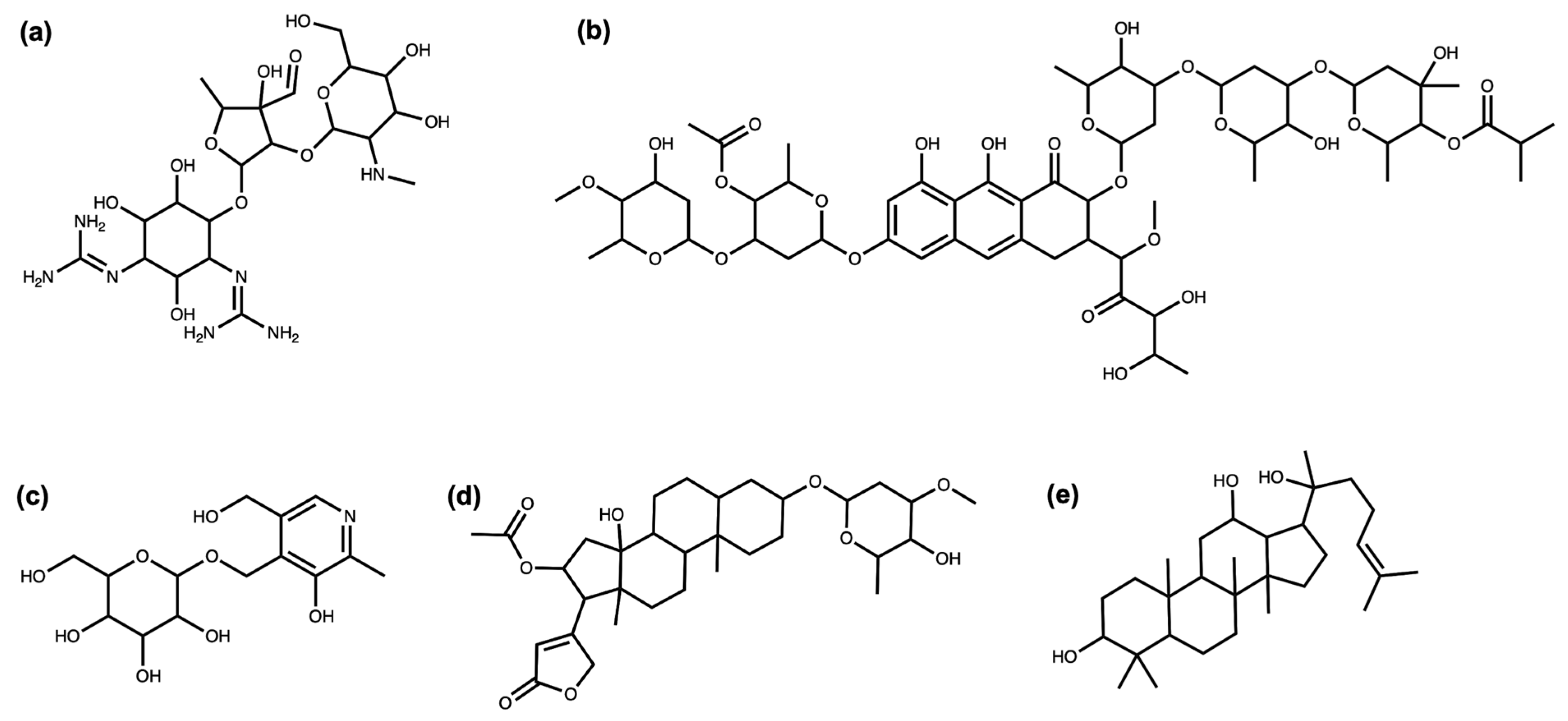
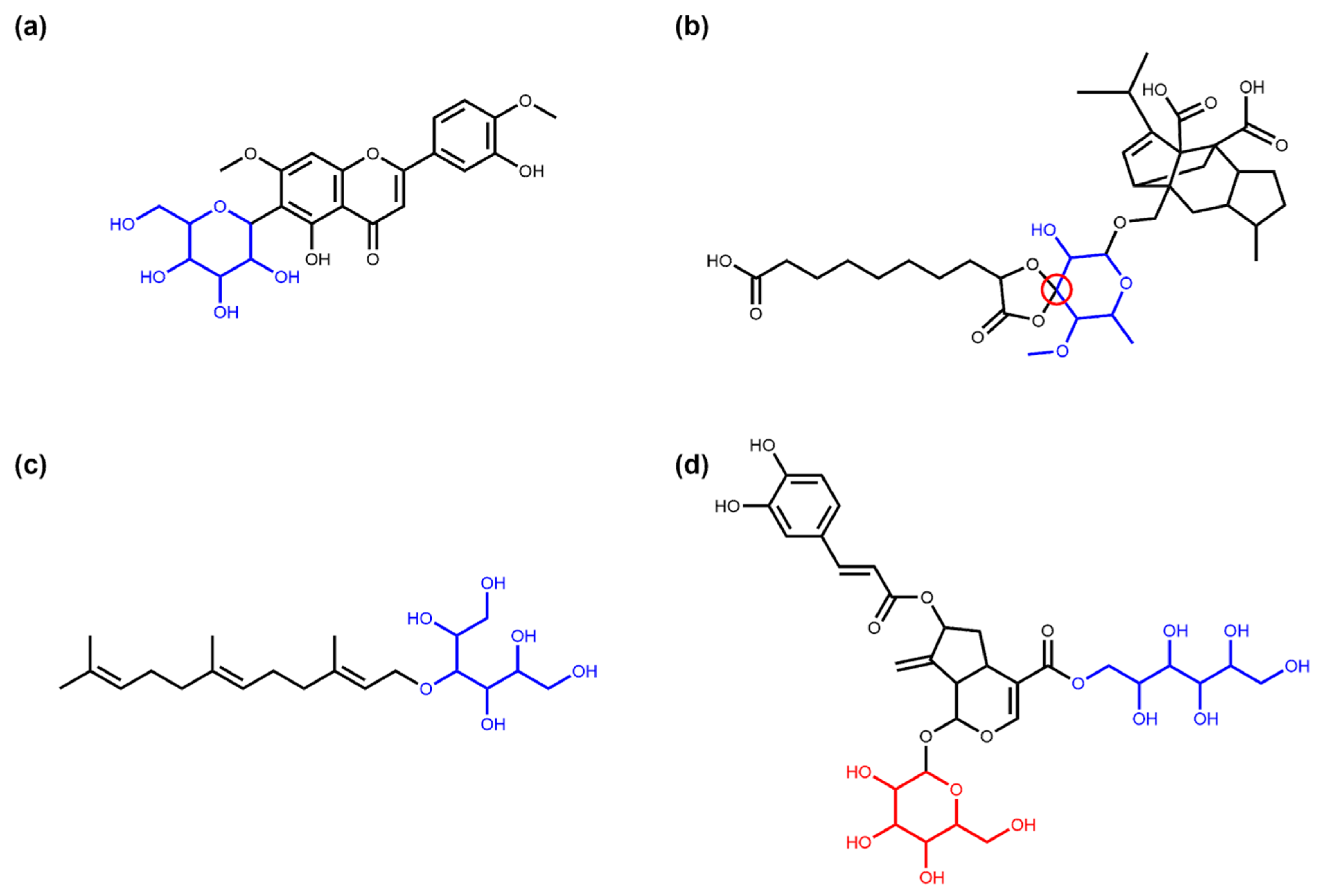
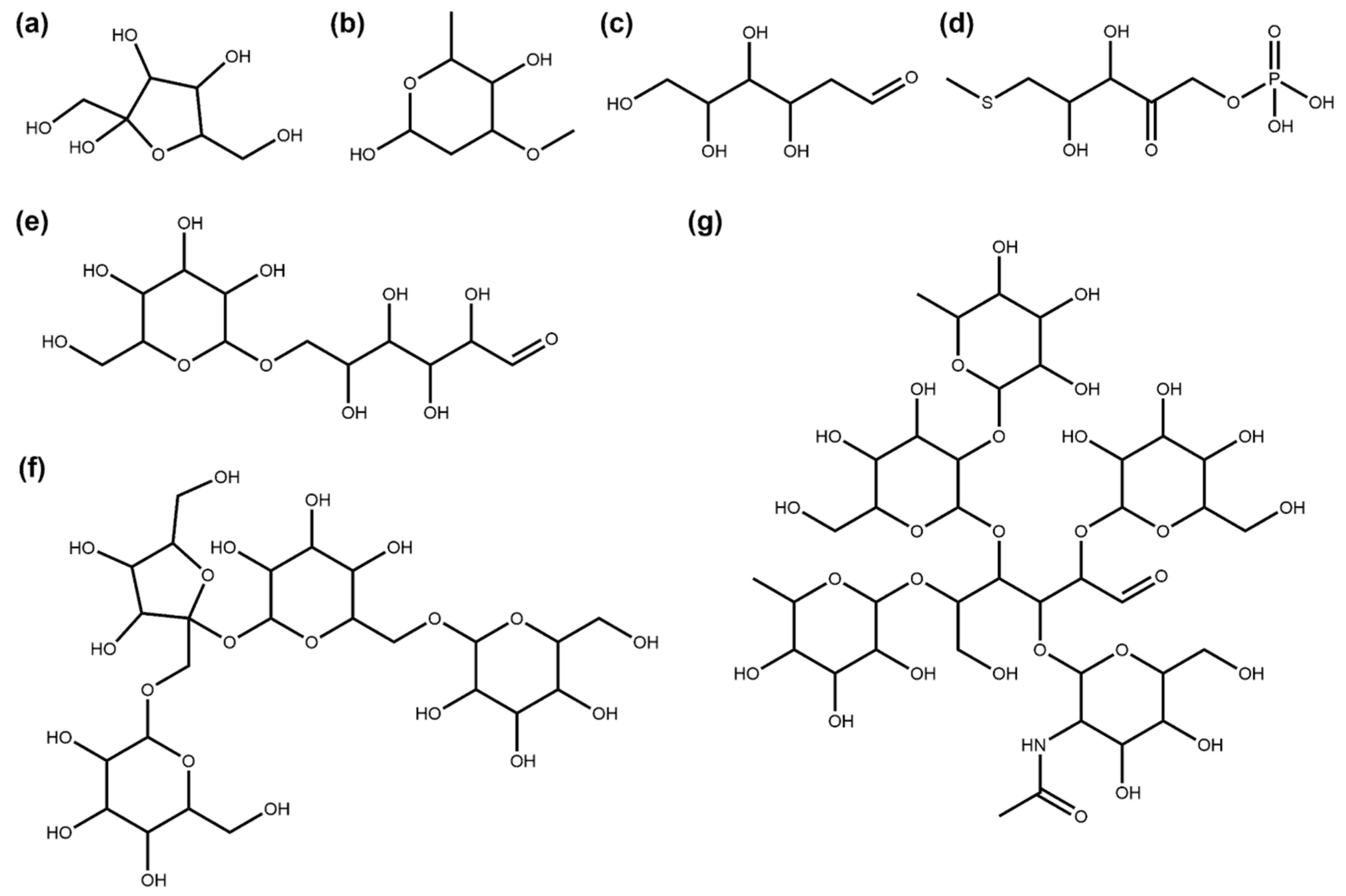
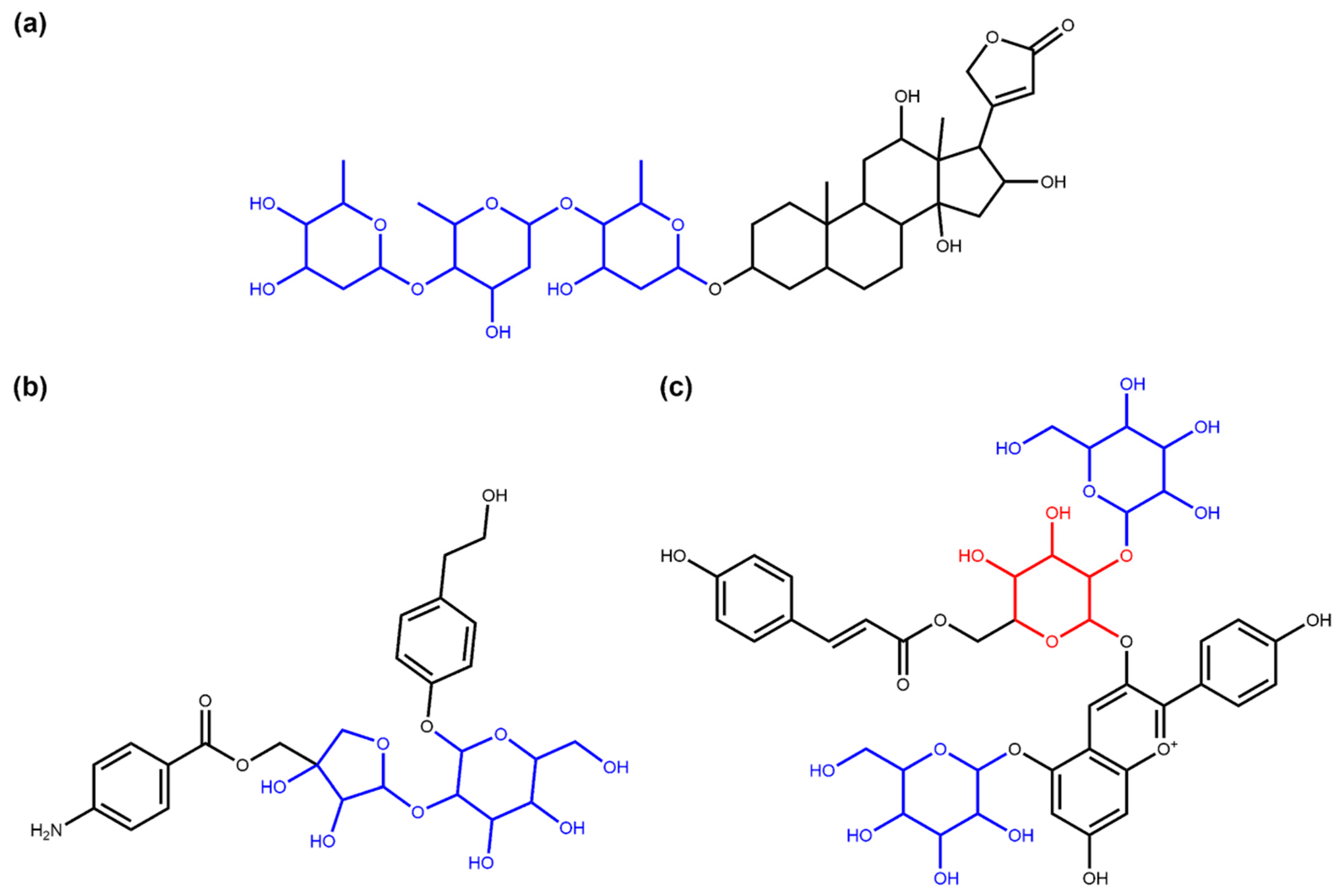
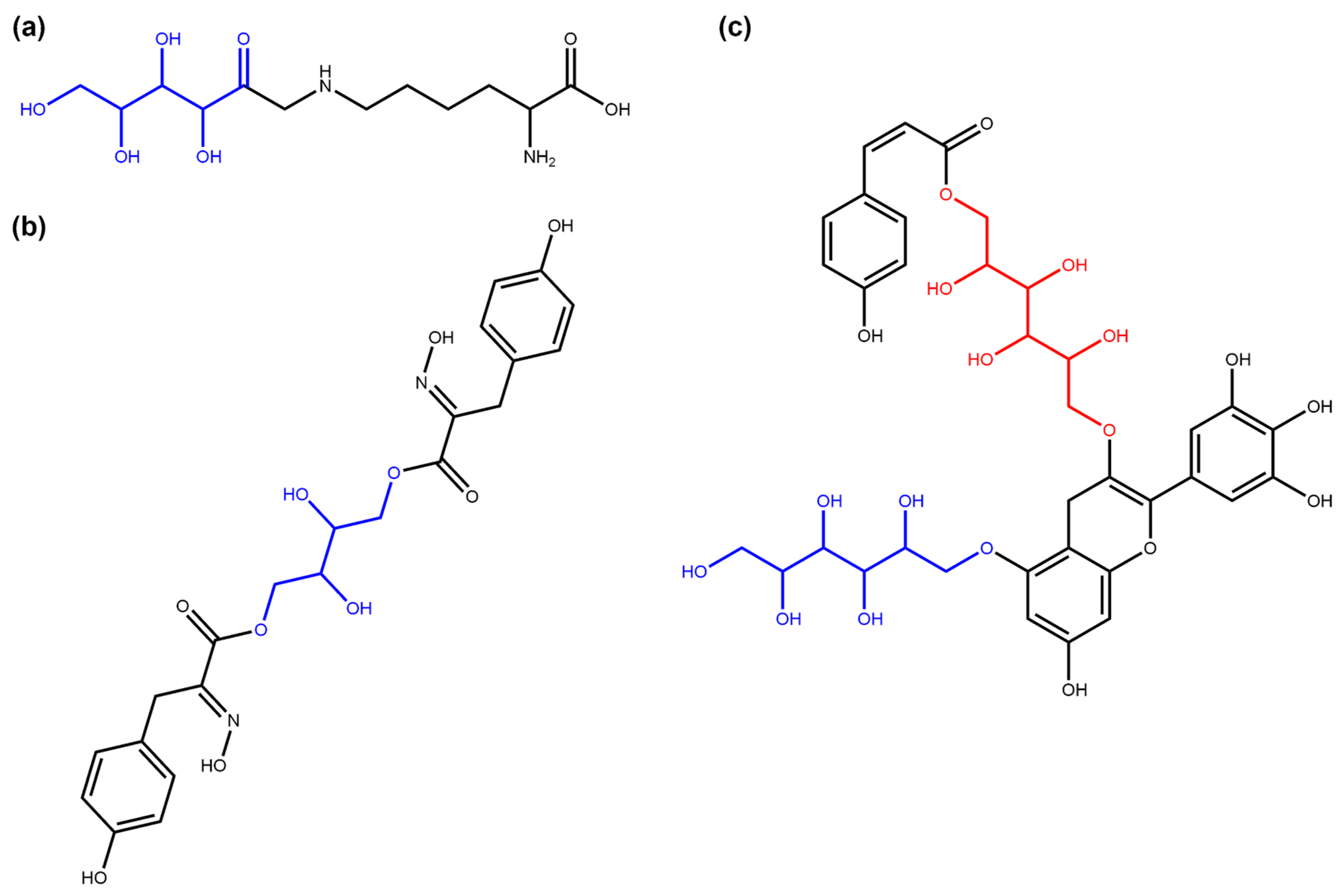
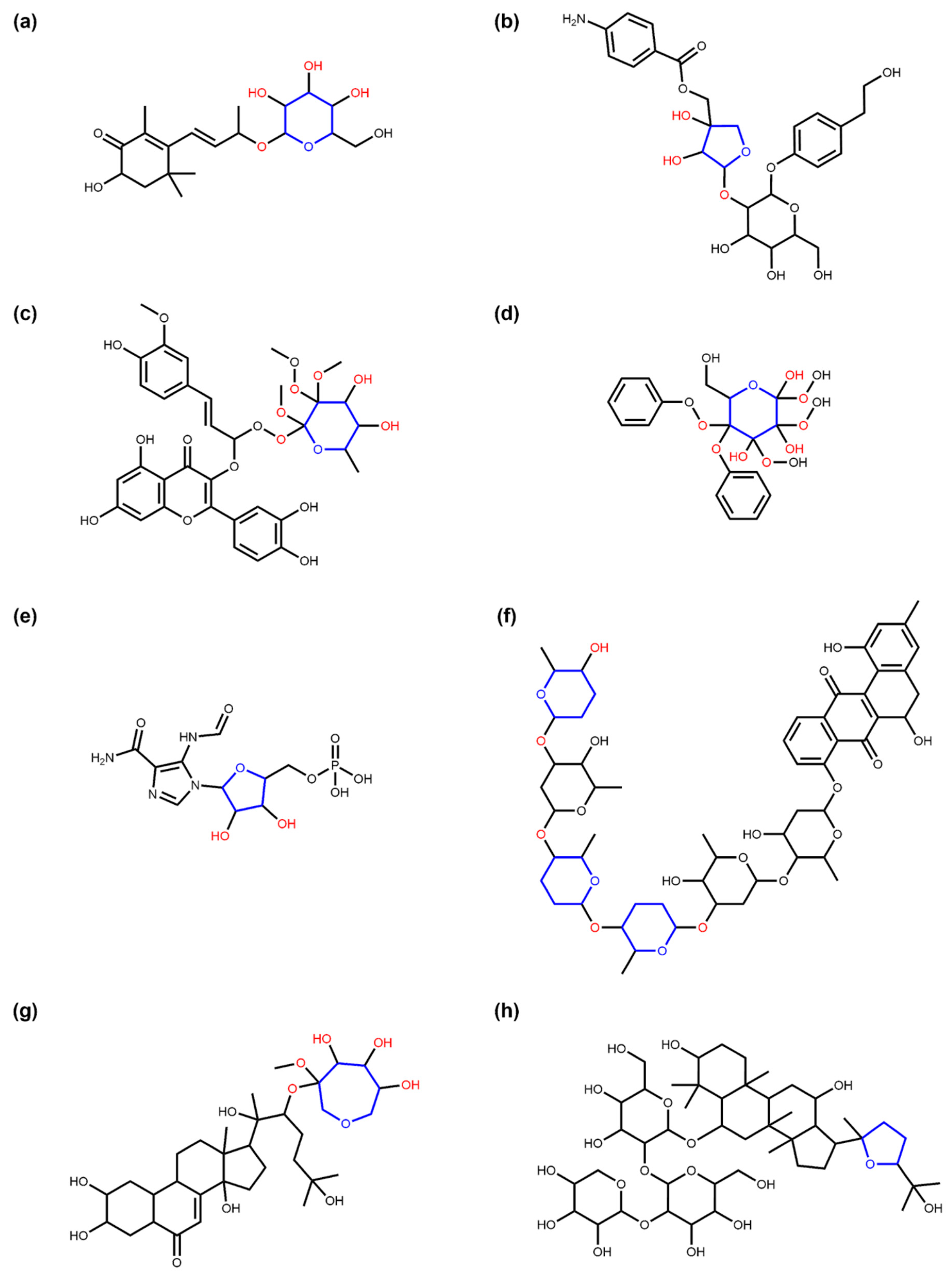
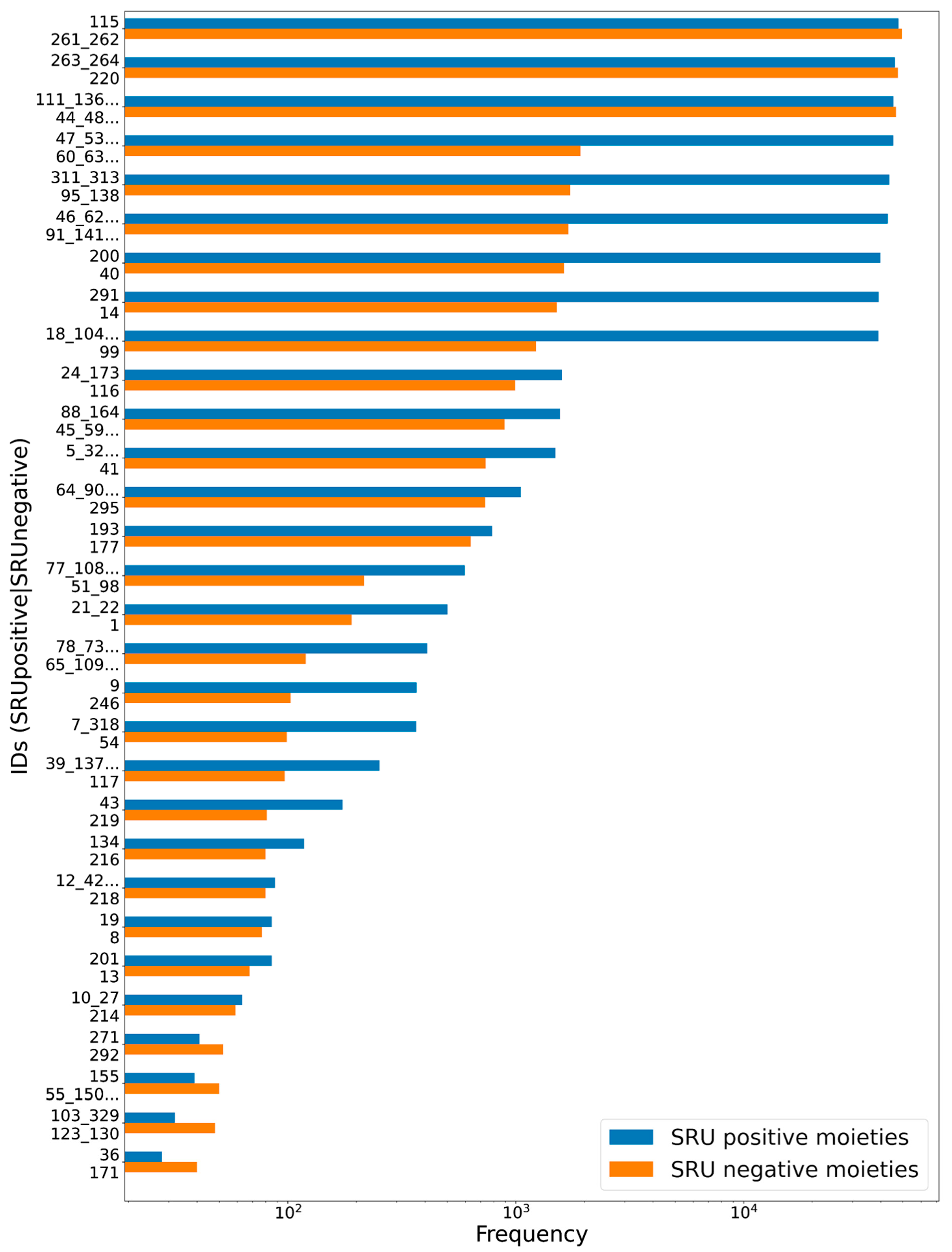
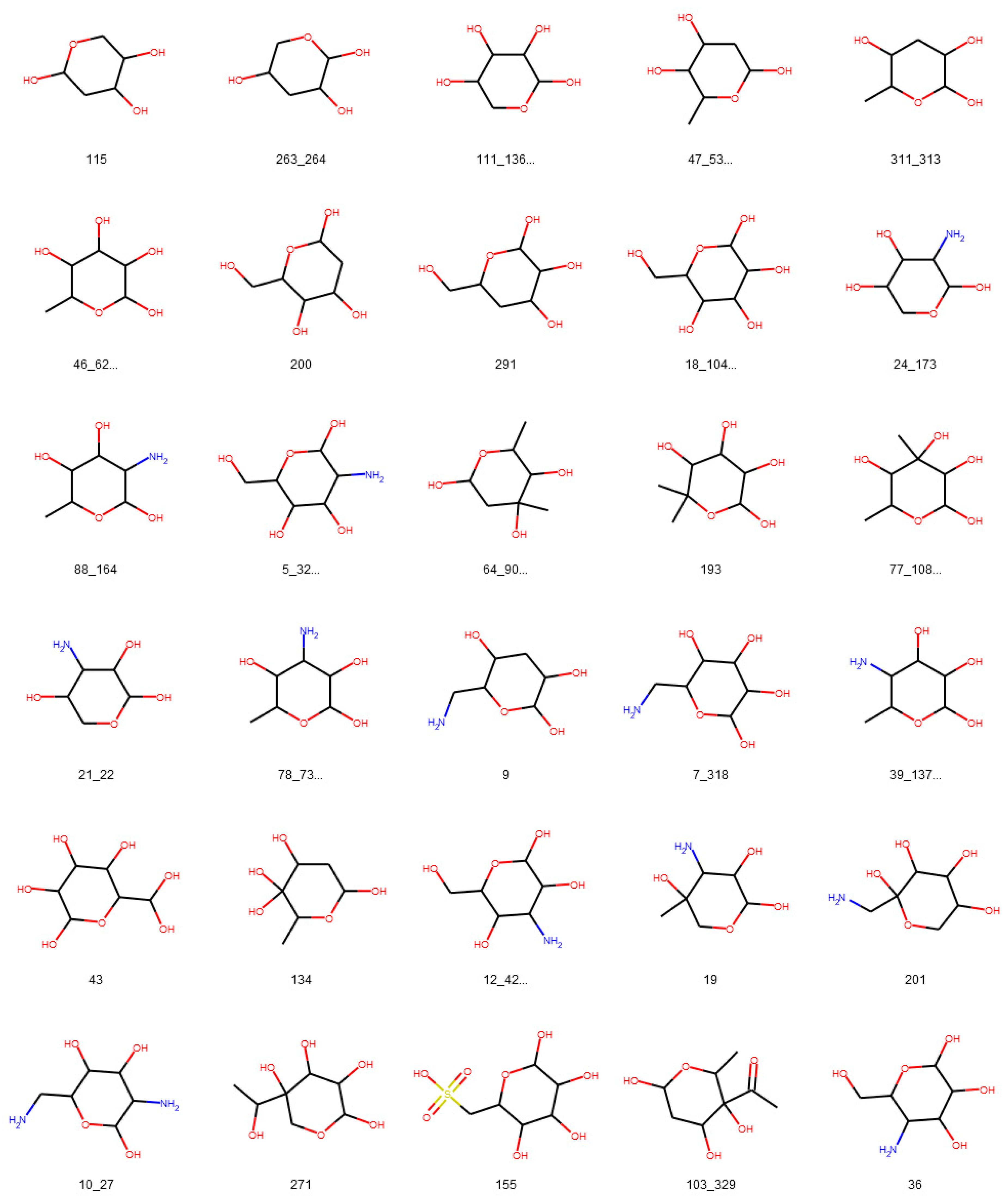
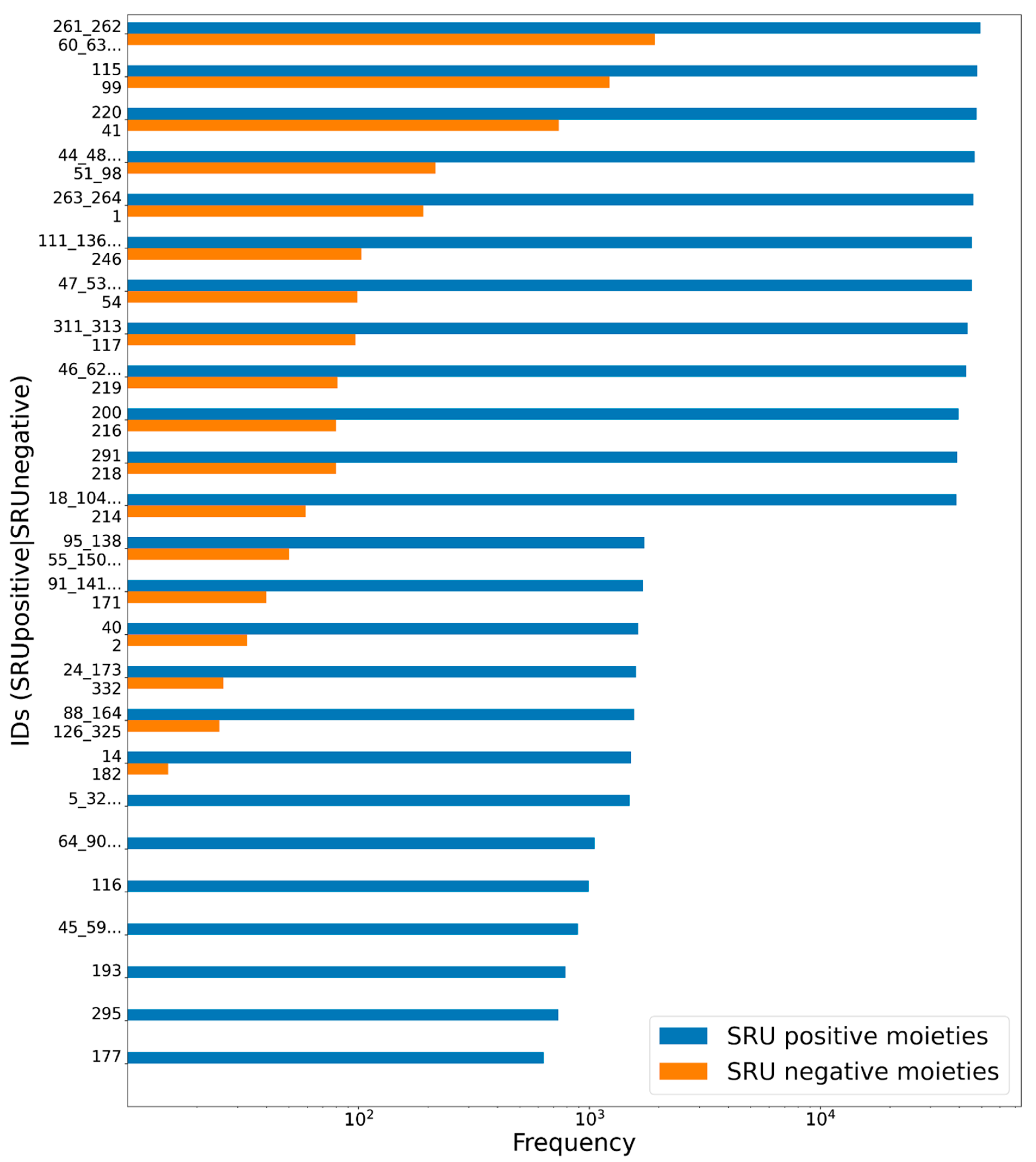
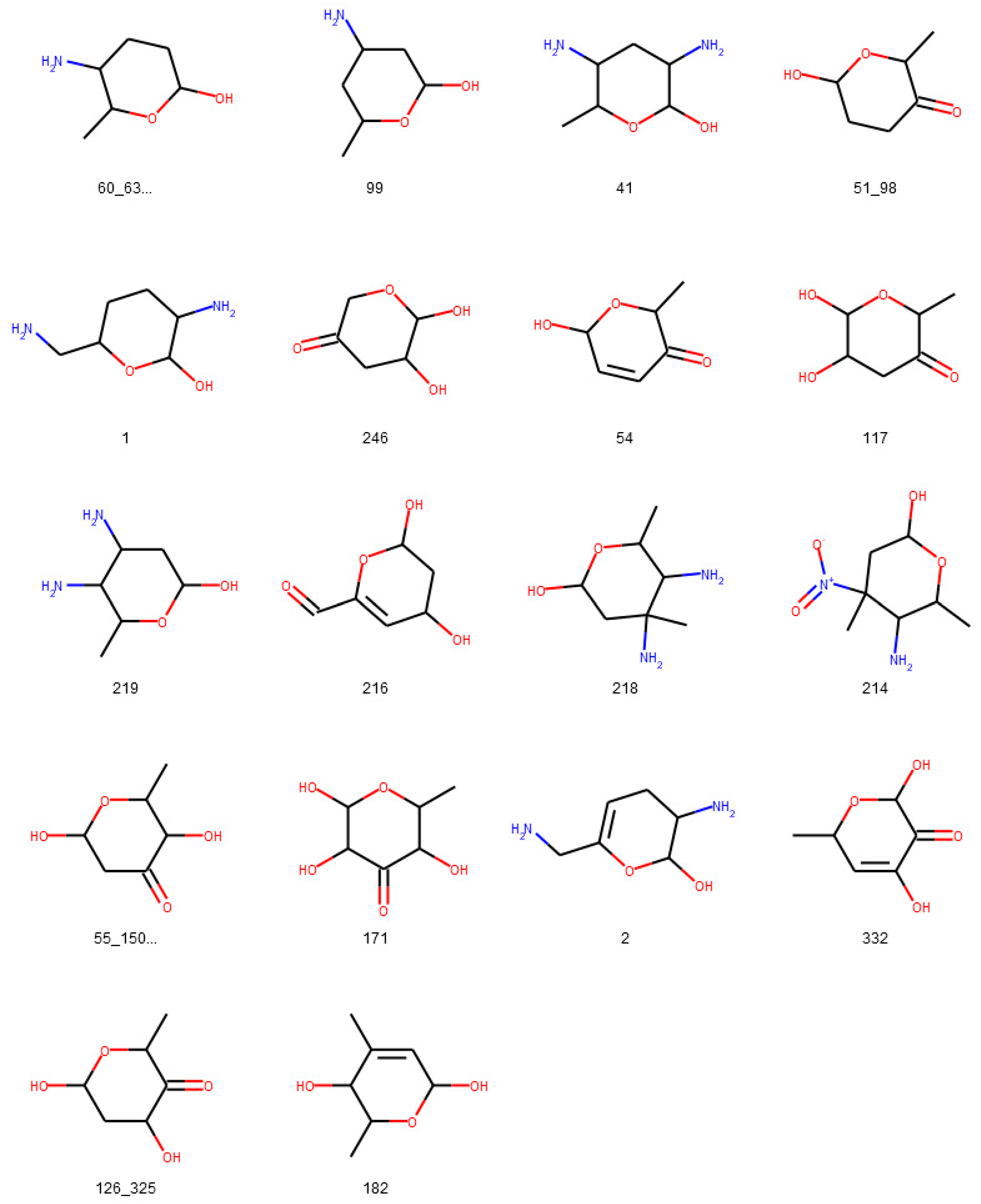
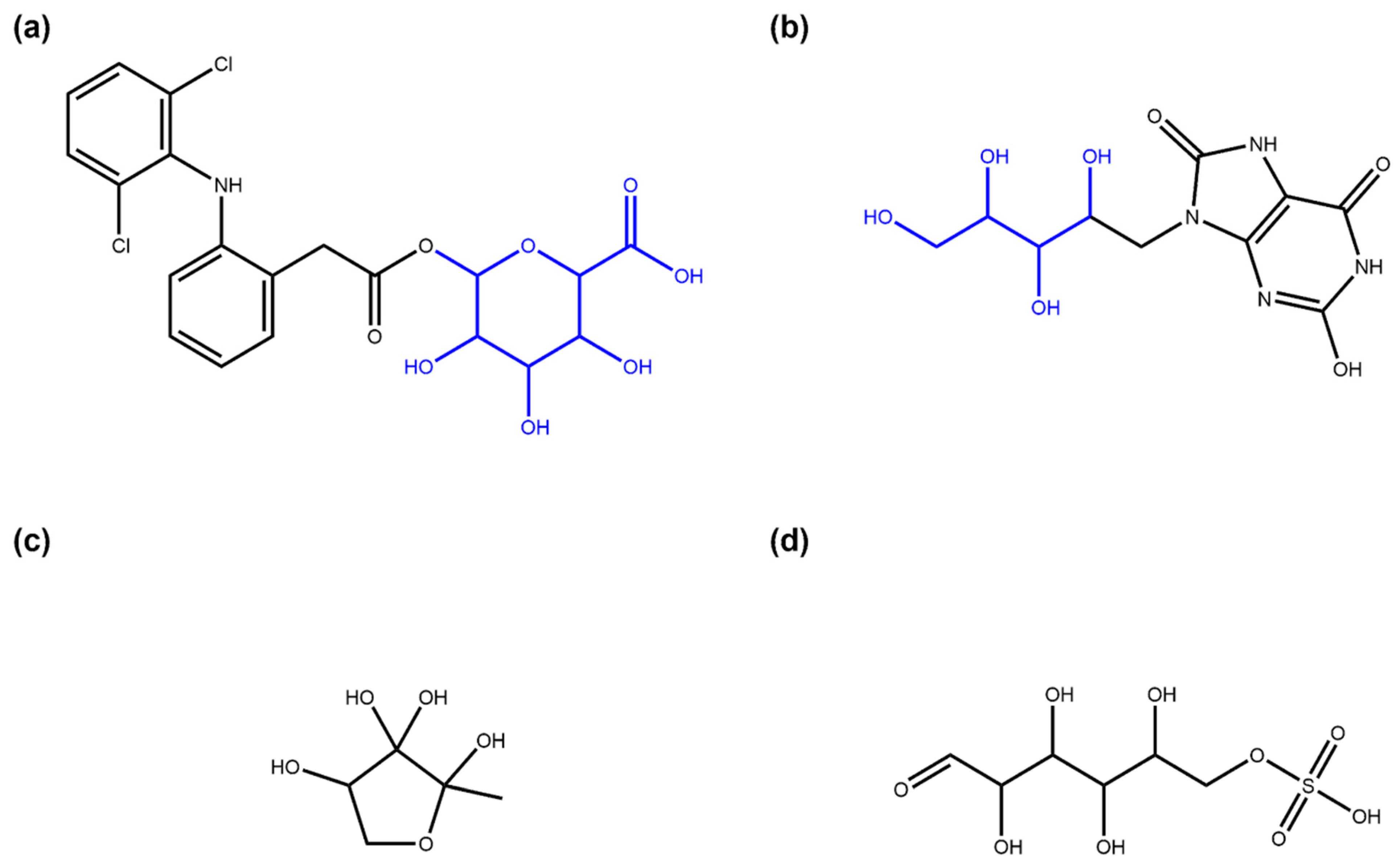
3.1. Deglycosylation and Glycoside Analysis in COCONUT
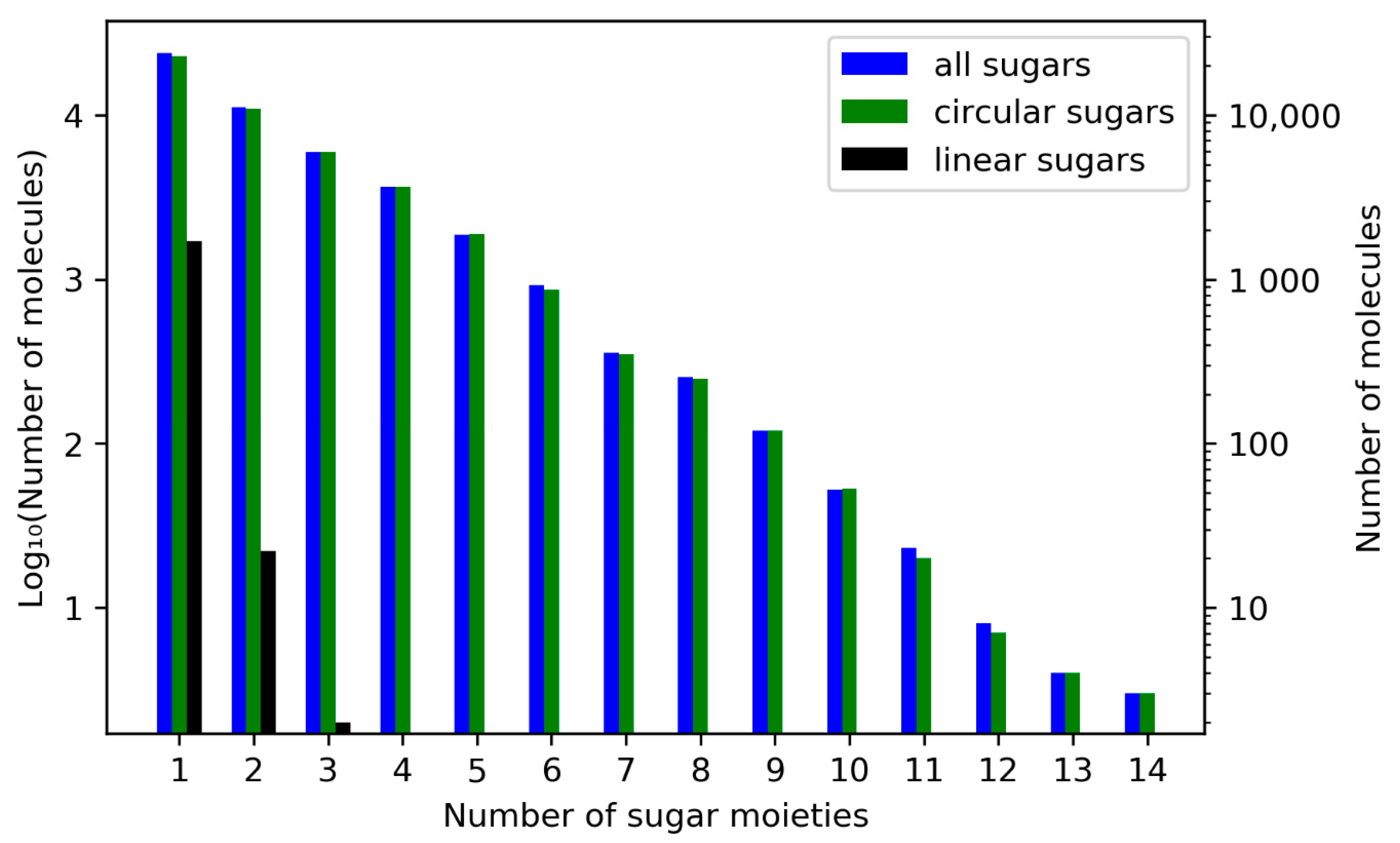
3.1.1. Description of Sugar Moieties in COCONUT
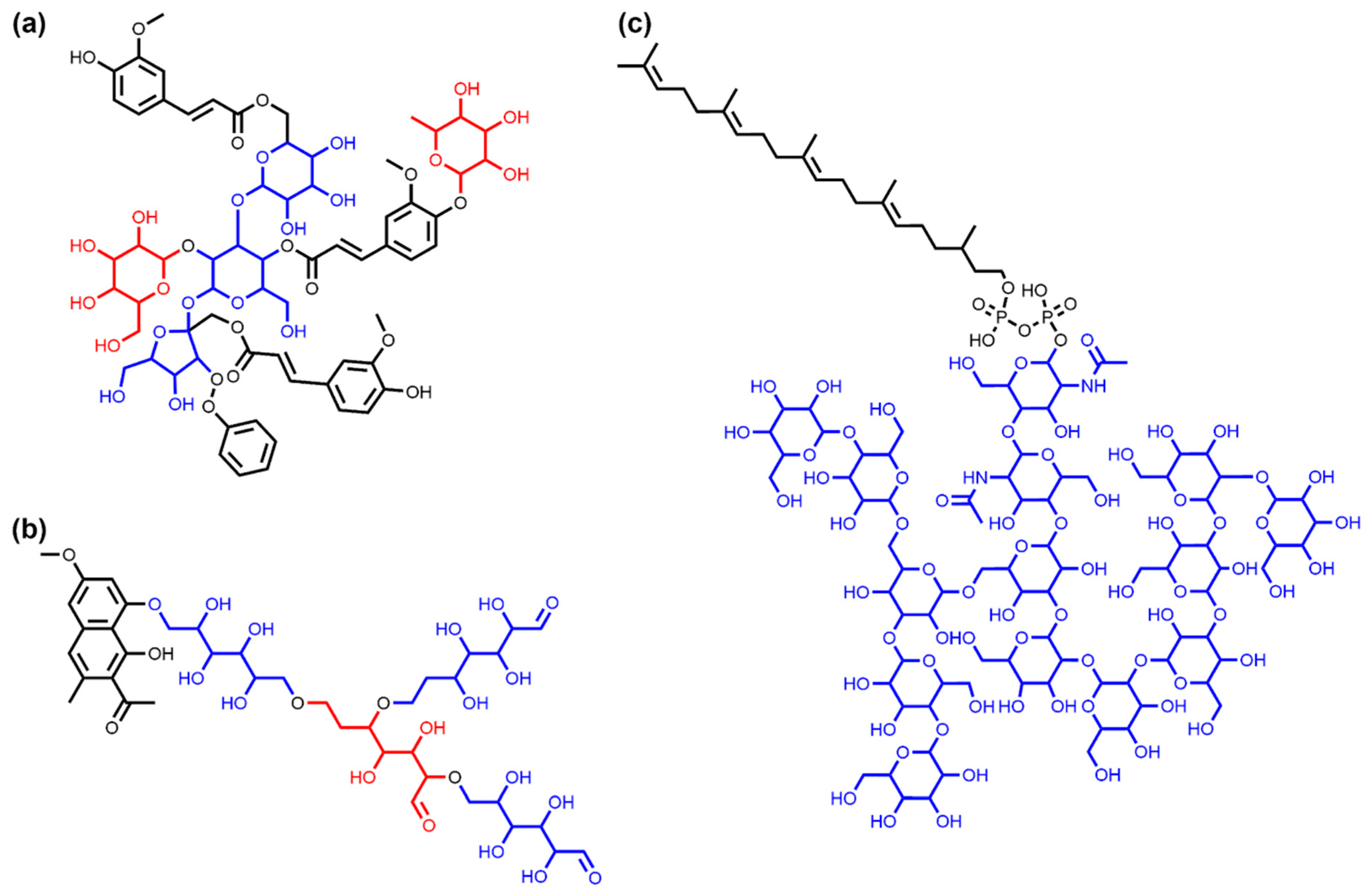
3.1.2. Bacterial NP Sugar Moieties
3.2. Glycosylation Analysis of Synthetic Molecules from ZINC
3.3. Glycosylation Analysis of “Active In Vitro” Molecules from ZINC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NP | natural product |
| SMILES | Simplified Molecular Input Line Entry System |
| SRU | Sugar Removal Utility |
| COCONUT | COlleCtion of Open Natural prodUcTs |
| ZINC | ZINC Is Not Commercial |
References
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The Re-Emergence of Natural Products for Drug Discovery in the Genomics Era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef]
- Sorokina, M.; Steinbeck, C. Review on Natural Products Databases: Where to Find Data in 2020. J. Cheminformatics 2020, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and Resupply of Pharmacologically Active Plant-Derived Natural Products: A Review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed]
- Kren, V.; Martinkova, L. Glycosides in Medicine: “The Role of Glycosidic Residue in Biological Activity”. Curr. Med. Chem. 2001, 8, 1303–1328. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.J.; Rather, P.N.; Hare, R.S.; Miller, G.H. Molecular Genetics of Aminoglycoside Resistance Genes and Familial Relationships of the Aminoglycoside-Modifying Enzymes. Microbiol. Mol. Biol. Rev. 1993, 57, 138–163. [Google Scholar] [CrossRef]
- Yoshizawa, S.; Fourmy, D.; Puglisi, J.D. Structural Origins of Gentamicin Antibiotic Action. EMBO J. 1998, 17, 6437–6448. [Google Scholar] [CrossRef]
- Hayasaka, T.; Inoue, Y. Chromomycin A3 Studies in Aqueous Solutions. Spectrophotometric Evidence for Aggregation and Interaction with Herring Sperm Deoxyribonucleic Acid. Biochemistry 1969, 8, 2342–2347. [Google Scholar] [CrossRef]
- Kaziro, Y.; Kamiyama, M. Mechanism of Action of Chromomycin A3: II. Inhibition of RNA Polymerase Reaction. J. Biochem. (Tokyo) 1967, 62, 424–429. [Google Scholar] [CrossRef]
- Mrudulakumari Vasudevan, U.; Lee, E.Y. Flavonoids, Terpenoids, and Polyketide Antibiotics: Role of Glycosylation and Biocatalytic Tactics in Engineering Glycosylation. Biotechnol. Adv. 2020, 41, 107550. [Google Scholar] [CrossRef]
- Nakano, H.; McMahon, L.G.; Gregory, J.F., III. Pyridoxine-5′-β-D-Glucoside Exhibits Incomplete Bioavailability as a Source of Vitamin B-6 and Partially Inhibits the Utilization of Co-Ingested Pyridoxine in Humans. J. Nutr. 1997, 127, 1508–1513. [Google Scholar] [CrossRef]
- Nakano, H.; Gregory, J.F., III. Pyridoxine-5′-β-D-Glucoside Influences the Short-Term Metabolic Utilization of Pyridoxine in Rats. J. Nutr. 1995, 125, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Sa, D.; Pn, L. Digoxin in Clinical Practice: Sorting out the Facts. Br. J. Clin. Pract. 1995, 49, 257–261. [Google Scholar]
- El-Seedi, H.R.; Khalifa, S.A.M.; Taher, E.A.; Farag, M.A.; Saeed, A.; Gamal, M.; Hegazy, M.-E.F.; Youssef, D.; Musharraf, S.G.; Alajlani, M.M.; et al. Cardenolides: Insights from Chemical Structure and Pharmacological Utility. Pharmacol. Res. 2019, 141, 123–175. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Bae, B.-S.; Park, H.-W.; Ahn, N.-G.; Cho, B.-G.; Cho, Y.-L.; Kwak, Y.-S. Characterization of Korean Red Ginseng (Panax Ginseng Meyer): History, Preparation Method, and Chemical Composition. J. Ginseng Res. 2015, 39, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Shibata, S. Saponins with Biological and Pharmacological Activity. In Proceedings of the New Natural Products and Plant Drugs with Pharmacological, Biological or Therapeutical Activity; Wagner, H., Wolff, P., Eds.; Springer: Berlin/Heidelberg, Germany, 1977; pp. 177–196. [Google Scholar]
- Cho, I.-H. Effects of Panax Ginseng in Neurodegenerative Diseases. J. Ginseng Res. 2012, 36, 342–353. [Google Scholar] [CrossRef]
- Sorokina, M.; Merseburger, P.; Rajan, K.; Yirik, M.A.; Steinbeck, C. COCONUT Online: Collection of Open Natural Products Database. J. Cheminformatics 2021, 13, 2. [Google Scholar] [CrossRef]
- Schaub, J.; Zielesny, A.; Steinbeck, C.; Sorokina, M. Too Sweet: Cheminformatics for Deglycosylation in Natural Products. J. Cheminformatics 2020, 12, 67. [Google Scholar] [CrossRef]
- Willighagen, E.L.; Mayfield, J.W.; Alvarsson, J.; Berg, A.; Carlsson, L.; Jeliazkova, N.; Kuhn, S.; Pluskal, T.; Rojas-Chertó, M.; Spjuth, O.; et al. The Chemistry Development Kit (CDK) v2.0: Atom Typing, Depiction, Molecular Formulas, and Substructure Searching. J. Cheminformatics 2017, 9, 33. [Google Scholar] [CrossRef]
- Sterling, T.; Irwin, J.J. ZINC 15 – Ligand Discovery for Everyone. J. Chem. Inf. Model. 2015, 55, 2324–2337. [Google Scholar] [CrossRef]
- Ashton, M.; Barnard, J.; Casset, F.; Charlton, M.; Downs, G.; Gorse, D.; Holliday, J.; Lahana, R.; Willett, P. Identification of Diverse Database Subsets Using Property-Based and Fragment-Based Molecular Descriptions. Quant. Struct. Act. Relatsh. 2002, 21, 598–604. [Google Scholar] [CrossRef]
- RDKit: Open-Source Cheminformatics Software. Available online: http://rdkit.org/ (accessed on 17 February 2021).
- Elshahawi, S.I.; Shaaban, K.A.; Kharel, M.K.; Thorson, J.S. A Comprehensive Review of Glycosylated Bacterial Natural Products. Chem. Soc. Rev. 2015, 44, 7591–7697. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, N.H.; Ahmed, A.A.; Ishak, M.S.; Kandil, F.E. Luteolin 7,4′-Dimethyl Ether 6-C-Glucoside from Parkinsonia Aculeata. Phytochemistry 1991, 30, 2442. [Google Scholar] [CrossRef]
- Zhang, M.-Q.; Xu, K.-X.; Xue, Y.; Cao, F.; Yang, L.-J.; Hou, X.-M.; Wang, C.-Y.; Shao, C.-L. Sordarin Diterpene Glycosides with an Unusual 1,3-Dioxolan-4-One Ring from the Zoanthid-Derived Fungus Curvularia Hawaiiensis TA26-15. J. Nat. Prod. 2019, 82, 2477–2482. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-H.; Lu, C.-K.; Wang, G.-J.; Chang, Y.-C.; Yang, W.-B.; Ju, Y.-M. Sesquiterpene Glycosides from Cosmospora Joca. J. Nat. Prod. 2011, 74, 1561–1567. [Google Scholar] [CrossRef] [PubMed]
- Taskova, R.M.; Kokubun, T.; Ryan, K.G.; Garnock-Jones, P.J.; Jensen, S.R. Iridoid and Phenylethanoid Glucosides from Veronica Lavaudiana. J. Nat. Prod. 2011, 74, 1477–1483. [Google Scholar] [CrossRef]
- Hopf, H.; Gruber, G.; Zinn, A.; Kandler, O. Physiology and Biosynthesis of Lychnose in Cerastium Arvense. Planta 1984, 162, 283–288. [Google Scholar] [CrossRef]
- Warren, C.D.; Chaturvedi, P.; Newburg, A.R.; Oftedal, O.T.; Tilde, C.D.; Newburg, D.S. Comparison of Oligosaccharides in Milk Specimens from Humans and Twelve Other Species. In Bioactive Components of Human Milk; Newburg, D.S., Ed.; Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 2001; Volume 501, pp. 325–332. ISBN 978-1-4613-5521-2. [Google Scholar]
- Otsuki, T.; Matsufuji, H.; Takeda, M.; Toyoda, M.; Goda, Y. Acylated Anthocyanins from Red Radish (Raphanus Sativus L.). Phytochemistry 2002, 60, 79–87. [Google Scholar] [CrossRef]
- Ingavat, N.; Dobereiner, J.; Wiyakrutta, S.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Aspergillusol A, an α-Glucosidase Inhibitor from the Marine-Derived Fungus Aspergillus Aculeatus. J. Nat. Prod. 2009, 72, 2049–2052. [Google Scholar] [CrossRef] [PubMed]
- Scialis, R.J.; Manautou, J.E. Elucidation of the Mechanisms through Which the Reactive Metabolite Diclofenac Acyl Glucuronide Can Mediate Toxicity. J. Pharmacol. Exp. Ther. 2016, 357, 167–176. [Google Scholar] [CrossRef]
- Schaefer, K.; Albers, J.; Sindhuwinata, N.; Peters, T.; Meyer, B. A New Concept for Glycosyltransferase Inhibitors: Nonionic Mimics of the Nucleotide Donor of the Human Blood Group B Galactosyltransferase. ChemBioChem 2012, 13, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.T.; Xavier, K.B.; Campagna, S.R.; Taga, M.E.; Semmelhack, M.F.; Bassler, B.L.; Hughson, F.M. Salmonella Typhimurium Recognizes a Chemically Distinct Form of the Bacterial Quorum-Sensing Signal AI-2. Mol. Cell 2004, 15, 677–687. [Google Scholar] [CrossRef]
- Huang, G.; Lv, M.; Hu, J.; Huang, K.; Xu, H. Glycosylation and Activities of Natural Products. Mini Rev. Med. Chem. 2016, 16, 1013–1016. [Google Scholar] [CrossRef]
- Goel, B.; Tripathi, N.; Mukherjee, D.; Jain, S.K. Glycorandomization: A Promising Diversification Strategy for the Drug Development. Eur. J. Med. Chem. 2021, 113156. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Molecules | % | |
|---|---|---|
| NPs in COCONUT | 401,624 | 100.00% |
| Has sugars | 48,118 | 11.98% |
| Has circular sugars | 46,979 | 11.70% |
| Has only circular sugars | 46,393 | 11.55% |
| Has linear sugars | 1725 | 0.43% |
| Has only linear sugars | 1139 | 0.28% |
| Has circular and linear sugars | 586 | 0.15% |
| Consists only of sugars | 1404 | 0.35% |
| Sugar monomer | 425 | 0.11% |
| Circular sugar monomer | 216 | 0.05% |
| Linear sugar monomer | 209 | 0.05% |
| Sugar polymer | 979 | 0.24% |
| Has terminal circular sugars | 39,217 | 9.76% |
| Has only terminal circular sugars | 35,167 | 8.76% |
| Has non-terminal circular sugars | 11,182 | 2.78% |
| Has only non-terminal circular sugars | 7762 | 1.93% |
| Has terminal and non-terminal circular sugars | 4050 | 1.01% |
| Has terminal linear sugars | 1119 | 0.28% |
| Has only terminal linear sugars | 1117 | 0.28% |
| Has non-terminal linear sugars | 608 | 0.15% |
| Has only non-terminal linear sugars | 606 | 0.15% |
| Has terminal and non-terminal linear sugars | 2 | 0.00% |
| Type of Glycosidic Moieties | No. of Glycosidic Moieties | % |
|---|---|---|
| In COCONUT in total | 99,943 | 100.00% |
| Circular | 98,189 | 98.24% |
| Circular, terminal | 80,927 | 80.97% |
| Circular, terminal, O-glycosidic bond | 78,755 | 78.80% |
| Linear | 1754 | 1.76% |
| Linear, terminal | 1139 | 1.14% |
| Terminal | 82,066 | 82.11% |
| Non-terminal | 17,877 | 17.89% |
| Circular, furanose | 4036 | 4.04% |
| Circular, pyranose | 94,149 | 94.20% |
| Circular, heptose | 4 | 0.00% |
| Linear, tetrose | 686 | 0.69% |
| Linear, pentose | 322 | 0.32% |
| Linear, hexose | 719 | 0.72% |
| Linear, heptose | 27 | 0.03% |
| Linear, ring substructure | 11,950 | 11.96% |
| Number of Molecules in | ||||||||
|---|---|---|---|---|---|---|---|---|
| COCONUT | % | ZINC Synthetics “For Sale” | % | ZINC Synthetics “In Vitro” | % | ZINC “In Vitro” | % | |
| Total molecules | 401,624 | 100.00% | 475,958 | 100.00% | 65,620 | 100.00% | 182,514 | 100.00% |
| Has sugars | 48,118 | 11.98% | 851 | 0.18% | 2974 | 4.53% | 12,732 | 6.98% |
| Has circular sugars | 46,979 | 11.70% | 523 | 0.11% | 2700 | 4.11% | 12,043 | 6.60% |
| Has linear sugars | 1725 | 0.43% | 334 | 0.07% | 320 | 0.49% | 840 | 0.46% |
| Has circular and linear sugars | 586 | 0.15% | 6 | 0.00% | 46 | 0.07% | 151 | 0.08% |
| Consists only of sugars | 1404 | 0.35% | 213 | 0.04% | 572 | 0.87% | 1087 | 0.60% |
| Has terminal circular sugars | 39,217 | 9.76% | 423 | 0.09% | 2258 | 3.44% | 9796 | 5.37% |
| Has non-terminal circular sugars | 11,812 | 2.94% | 100 | 0.02% | 505 | 0.77% | 2561 | 1.40% |
| Has terminal and non-terminal circular sugars | 4050 | 1.01% | 0 | 0.00% | 63 | 0.10% | 314 | 0.17% |
| Has terminal linear sugars | 1119 | 0.28% | 300 | 0.06% | 225 | 0.34% | 608 | 0.33% |
| Has non-terminal linear sugars | 608 | 0.15% | 34 | 0.01% | 95 | 0.14% | 232 | 0.13% |
| Has terminal and non-terminal linear sugars | 2 | 0.00% | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schaub, J.; Zielesny, A.; Steinbeck, C.; Sorokina, M. Description and Analysis of Glycosidic Residues in the Largest Open Natural Products Database. Biomolecules 2021, 11, 486. https://doi.org/10.3390/biom11040486
Schaub J, Zielesny A, Steinbeck C, Sorokina M. Description and Analysis of Glycosidic Residues in the Largest Open Natural Products Database. Biomolecules. 2021; 11(4):486. https://doi.org/10.3390/biom11040486
Chicago/Turabian StyleSchaub, Jonas, Achim Zielesny, Christoph Steinbeck, and Maria Sorokina. 2021. "Description and Analysis of Glycosidic Residues in the Largest Open Natural Products Database" Biomolecules 11, no. 4: 486. https://doi.org/10.3390/biom11040486
APA StyleSchaub, J., Zielesny, A., Steinbeck, C., & Sorokina, M. (2021). Description and Analysis of Glycosidic Residues in the Largest Open Natural Products Database. Biomolecules, 11(4), 486. https://doi.org/10.3390/biom11040486