1. Introduction
Agricultural intensification to meet growing global food demand has prompted investigations into sustainable management practices, including alternative fertilization and application of biostimulants to enhance both crop growth and soil health, particularly in the face of global climate change [
1,
2,
3,
4]. In conventional agriculture, fertilization is largely focused on the three macronutrients nitrogen (N), phosphorus (P) and potassium (K), which has led to soil acidification and soils that are deficient in a range of micronutrients [
5,
6]. Micronutrients are essential for plant growth and development and often function as enzyme cofactors [
7]. Micronutrient fertilizers are typically supplied in inorganic forms, such as chlorides or sulfate salts, to ameliorate deficiencies, but it is often unclear how such fertilizers impact additional processes within the soil ecosystem (e.g., microbial activity). While inorganic nutrients are rapidly available for plant uptake, they do not provide the additional benefits provided by organic fertilizers (e.g., green and livestock manure) through increased carbon (C) inputs. These valuable organic inputs from soil additions rich in a variety of C compounds have proven benefits such as increased overall soil health, soil water holding capacity, cation exchange capacity (CEC), aggregation and infiltration, pH stabilization, and enhanced microbial activities [
8,
9,
10,
11].
Organic amendments generally require microbial degradation to release nutrients into the soil solution for plant uptake, which is a relatively slow process, and the benefits are only realized long-term (a year, to several years past the current growing season). Thus, conventional chemical fertilizers are still required to obtain optimal yields [
12,
13]. A plausible alternative may be to supply micronutrients chelated to labile C compounds that can be rapidly metabolized by soil microbes and released into soil solution, thus stimulating microbial activity and providing readily available elements for plant nutrition. Furthermore, several studies have proven the ability of plant roots to directly take up small organic molecules such as low molecular weight organic acids (LMWOA), sugars, or amino acids, indicating that organic fertilizers of this nature could represent a direct source of nutrients for plant uptake [
14,
15,
16,
17,
18].
When extracellular metal concentrations are high, they are preferentially accumulated in the roots or translocated to aerial tissues via xylem; the tendency to accumulate in the roots or shoots is primarily dependent on the specific metal and plant species [
7]. For example, Cu is preferentially accumulated in the roots when external Cu concentrations are high, and thus plants under Cu toxicity usually exhibit reduced root growth [
19]. However, high external Mn concentrations may manifest stress symptoms in aboveground tissues due to preferential accumulation of Mn in the shoots and leaves [
7]. Thus, achieving the balance between micronutrient deficiency and toxicity is crucial to support optimal plant growth.
Metal lactates are an understudied class of organic micronutrient supplement that show potential to supply micronutrients and trace metals directly for plant uptake, in addition to providing a labile C source for soil and rhizosphere microbial populations. Previous work has shown lactic acid addition to soil enhances microbial activity [
20,
21], while foliar applications of lactic acid in hydroponically grown cucumber improves shoot development and yield [
22,
23]. In fact, lactic acid showed the greatest potential to enhance soil microbial activity in comparison to other LMWOA [
20,
24]. Lactic acid and lactate-conjugated metals are a common additive to vitamin supplements and widely used for food science applications [
25]. However, little is known about the capacity of crops to uptake and translocate metals bound to lactate. In
Arabidopsis thaliana, there are nine nodulin 26 intrinsic proteins (NIPs) that are highly conserved water and solute transport proteins. One of them, NIP2;1, was identified as a lactate transporter in roots induced particularly during periods of hypoxia and likely functioning in efflux [
26]. Additionally, Zn/Fe permeases (ZIPs), heavy metal ATPases (HMAs), and Yellow Stripe Like (YSL) transporters, all have shown the ability to transport a range of metals [
27,
28,
29].
The influence of lactate complexation on metal uptake and toxicity was assessed using five metals (Zn, Mn, Cu, Ni, and Co). The metals were supplied at toxic levels as either a lactate or a chloride salt form in aseptic hydroponic culture vessels, which allowed for comparison of the two forms of metals in the absence of microbial activity. Importantly, both metal chlorides and metal lactates have similar stability constants, making them suitable forms to compare on a chemical level [
30,
31,
32]. The uptake capacity and subsequent toxicity of metal lactates was compared with that of metal chlorides to investigate the importance of plant uptake of the organic ligand-bound metals. Zn, Mn, Cu, Ni, and Co concentrations were measured in root and shoot tissues using MP-AES to validate uptake of each metal. To further investigate optimal metal lactate concentrations for plant growth, we generated dose−response curves for individual metal lactates by quantifying primary root growth rates in
A. thaliana grown in a range of concentrations that more closely reflect physiologically relevant micronutrient levels according to previous reports [
33].
2. Materials and Methods
2.1. Wheat Seed Sterilization and Pre-Germination
Wheat seedlings were cultivated under aseptic conditions, starting with seed sterilization prior to sewing in sterile culture vessels, and using sterile nutrient solutions. ”Louise” (
Triticum aestivum L.) wheat seeds were chosen for this study due to the importance of this variety in the state of Washington [
34]. Seeds were surface sterilized through a series of rinse steps adapted from Speakman and Kruger (1983) [
35]. Briefly, 35 seeds were placed into 50 mL conical tubes and immersed in 40 mL of a 0.001% terramycin (CAS: 2058-46-0, Alfa Aesar, Haverhill, MA, USA) solution with 60 µL of 50% Tween 20 (23336-0010, Acros Organics, Fair Lawn, MA, USA) for 20 h at room temperature in the dark. Then, seeds were immersed in 40 mL of a 0.1% AgNO
3 (CAS: 7761-88-8, Alfa Aesar, Haverhill, MA, USA) solution with 0.84 mL of 50% Tween 20 for 10 min. The AgNO
3 solution was decanted and seeds were immersed in 40 mL of a 0.5% NaCl solution with 0.84 mL of 50% Tween 20 for 30 s. Seeds were rinsed five times with 40 mL of sterile Milli-Q (Milli-Q Advantage A10 System, Millipore Sigma, Burlington, MA, USA) water. After this sterilization procedure, seeds were placed on sterile Phytagel plates (0.4% Phytagel: P8169-250G, Sigma Life Science, St. Louis, MO, USA; 0.036% MgSO
4: M63-500, Fischer Scientific; 0.018% CaCl
2 dihydrate: C79-500, Alfa Aesar, Haverhill, MA, USA) and stored in the dark, at room temperature (~23 °C) for three days. After three days, pre-germinated seedlings were inspected for microbial contaminants, and only non-contaminated seedlings of approximately equivalent developmental stage were used in the study.
2.2. Wheat Cultivation Vessels and Media
To remove any possible residual and trace metals from experimental materials, all culture vessels, glass beads, and polypropylene pellets were acid-washed in a 100 mM HNO
3 and HCl solution for a minimum of two hours and subsequently rinsed with ultrapure Milli-Q water eight times. Sterilized and pre-germinated wheat seedlings were grown for 13 days in 946 mL glass culture vessels (Phyto Technology Laboratories #C607, Lenexa, KS, USA) with vented screw caps (0.5 µm pore size; Phyto Technology Laboratories #C2055, Lenexa, KS, USA) to allow for gas exchange (
Figure S1). Within each culture vessel, 250 g of 6 mm diameter glass beads were added and used as the physical plant growth support matrix. MilliQ water was then added according to volumes shown in
Table S1, and each culture vessel was sterilized under 15 PSI at 121 °C for 30 min. A 0.25× modified Hoagland’s nutrient solution (pH 5.9) described in
Table S2 was added to each culture vessel in the volumes indicated in
Table S1 [
36]. Metals and nutrients that were added as a part of the experimental design in the form of either lactate or chloride treatments that would normally be included in a complete Hoagland’s nutrient solution (i.e., Zn, Cu, and Mn), were excluded from the Hoagland’s solution for that treatment. A 0.229 M stock solution of each metal lactate (Zn, Cu, Mn, Ni, and Co lactate) and metal chloride (Zn, Cu, Mn, Ni, and Co chloride) was prepared. Metal chloride solutions were sterilized by autoclaving at 15 PSI, 121 °C for 30 min, while the metal lactate solutions were filter sterilized (Millipore Sigma #SCVPU11RE, Burlington, MA, USA; 0.1 µm pore size). Metal lactate and metal chloride solutions were each added in the volumes shown in
Table S1 to bring total solution volumes in each vessel to 50 mL to create the final treatment concentrations also shown in
Table S1. The treatment concentrations used in this study were chosen to reflect concentrations that are several orders of magnitude higher than the original concentration of Mn, Cu, and Zn in Hoagland’s nutrient solution. Since Co and Ni are not included in a complete Hoagland’s nutrient solution, the same concentrations were used across all treatments. Control culture vessels did not receive the metal added via metal lactate and metal chloride treatments.
Following the nutrient solution preparation, each sterile wheat seedling was carefully placed into the glass beads, and the surface of the glass beads in each vessel was then covered by a layer of sterile, black polypropylene pellets to prevent excessive light exposure on seedling roots. Additionally, black plastic covers (ShuBee, Macon, GA, USA) were placed on the bottom exterior portion of each culture vessel to further inhibit light exposure on roots. Wheat seedlings were grown in culture vessels at room temperature under growth lights (Ecolux w/Starcoat, F32T8 SPX35 Eco, GE, Boston, MA, USA; photon flux of 55 µmol m−2 s−1) with a photoperiod of 16 h for 13 days.
2.3. Microbial Contamination Analysis
Prior to collecting each wheat seedling for growth and tissue analysis, a 100 mL aliquot of nutrient solution from each culture vessel was inoculated onto sterile 0.8% tryptone yeast agar (tryptone: 0123-17, Difco Laboratories, Detroit, MI, USA; yeast extract: J850-500G, Amresco Inc., Solon, OH, USA; agar: J637-500G, VWR International, Radnor, PA, USA) plates to detect culturable microorganisms. The plates were stored in the dark at room temperature for a minimum of six days and visually inspected for any microbial colonies. Plates displaying microbial growth represented culture vessels that were contaminated, and those vessels were not included in the plant growth parameter results.
2.4. Wheat Growth and Metal Response
At the conclusion of the 13-day growth period, each wheat seedling was carefully removed from the culture vessel, and the roots were washed in a 0.01 M CaCl2 solution to remove loosely bound metals from the root surface. The seedlings were then cut above the seed crown to separate the shoot and root tissues, and shoot tissues were measured for total length (cm), and both the shoot and root tissues were weighed to obtain fresh weights (FW). Tissues were then stored in 50 mL conical tubes, flash-frozen in liquid nitrogen, and subsequently stored at −80 °C. Frozen tissues from the control (0 mM), 0.343, and 3.43 mM treatments were lyophilized using a Labconco FreeZone 6 freeze dryer (Labconco Corporation, Kansas City, MO, USA) for at least 72 h and subsequently weighed to obtain dry weights (DW).
2.5. Wheat Tissue Analysis
Below and above ground lyophilized tissue were weighed into PTFE digestion tubes. Because these were small samples, the entire sample (1–30 mg) was added to the tubes and acid digested using 1 mL of HNO3. After digestion, 1 mL of ultrapure water was added to each vial. The solutions were filtered with a 0.45 µm PTFE filter (Thermo Fisher Scientific, Waltham, MA, USA). Filtered digests were then diluted 25× and analyzed using an Agilent 4200 microwave plasma-atomic emission spectrometer (MP-AES) (Agilent, Santa Clara, CA, USA) and run in combination with Zn, Cu, Mn, Co, and Ni ICP standards ranging from 0–500 ng g−1 for validation.
2.6. Plate-Growth Assays
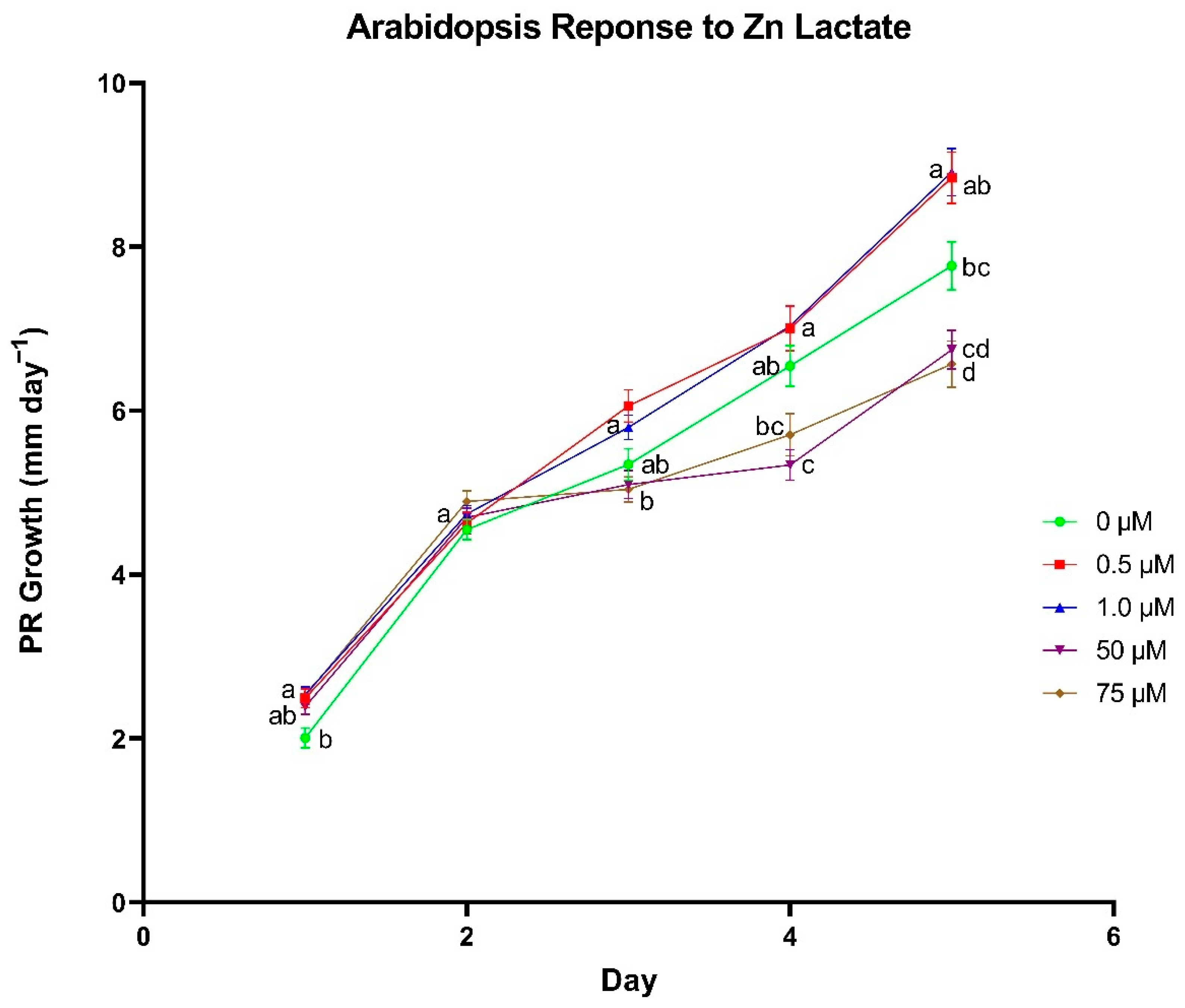
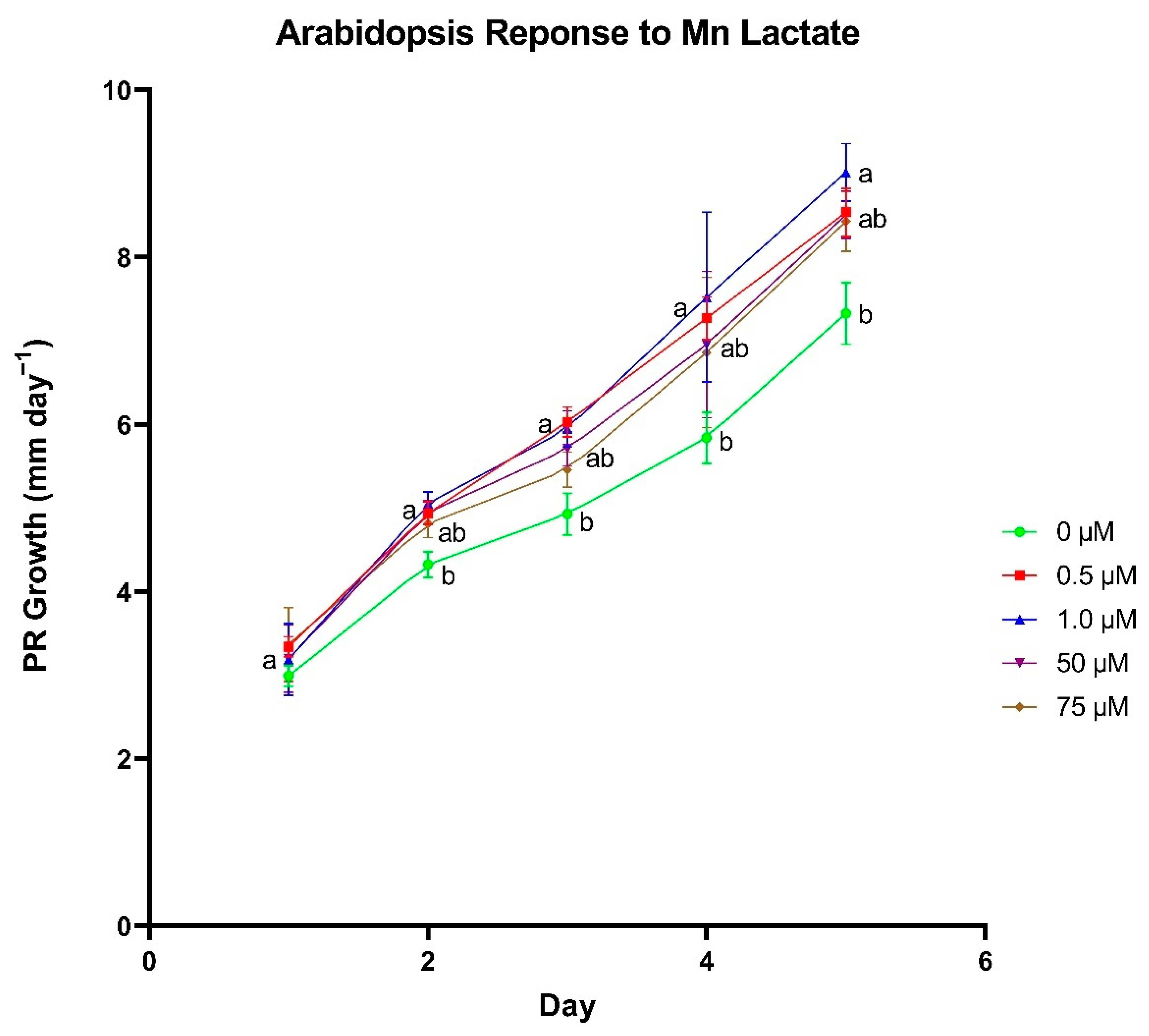
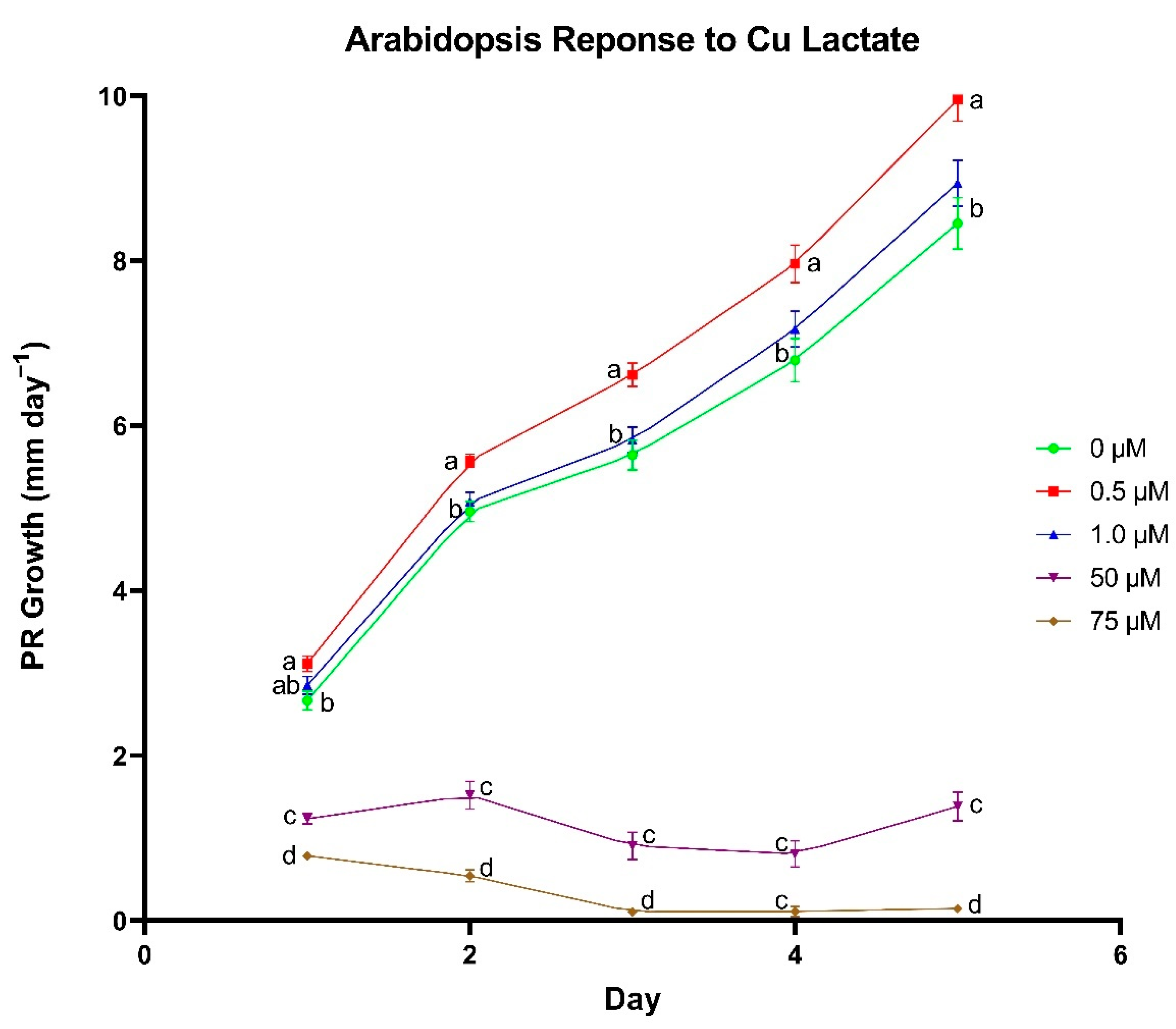
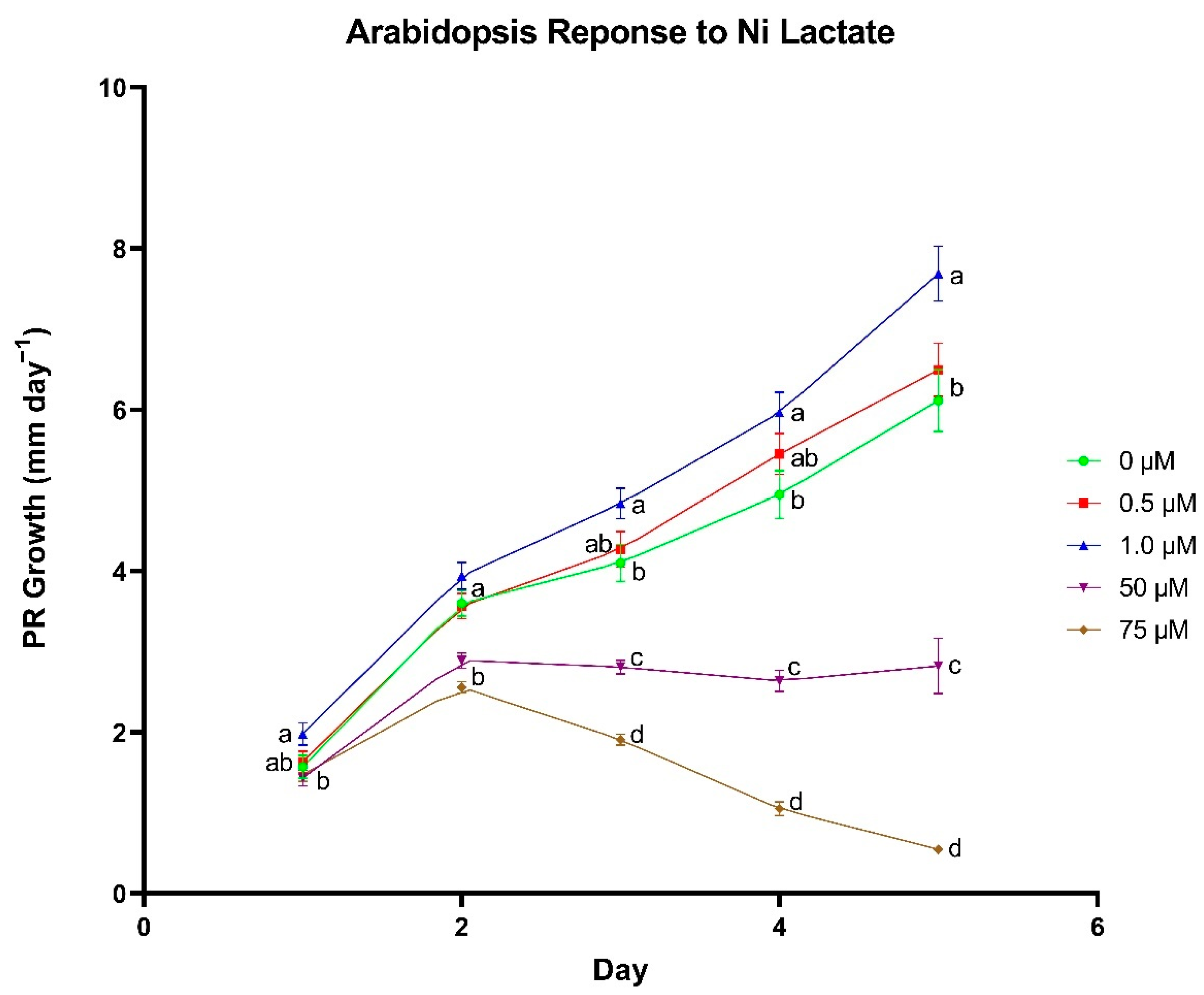
A series of plate-growth assays were used to determine optimal concentrations for each individual metal lactate. Arabidopsis thaliana (ecotype Columbia-0) seeds were surface sterilized in a 10% (v/v) bleach and 0.02% (v/v) Triton X-100 solution for 5 min followed by a 1-min wash in 70% (v/v) ethanol. Seeds were rinsed with sterile nanopure water before and after the ethanol wash. Sterilized seeds were transferred onto 1% Bacto agar plates (pH 5.8) containing 0, 0.5, 1, 50, and 75 µM of individual metal lactate and 0.25× modified Hoagland’s nutrient solution. Metals that were added as metal lactates that would normally be included in a complete Hoagland’s nutrient solution (i.e., Zn, Cu, and Mn), were excluded from the Hoagland’s solution for that treatment. Each concentration was represented by 8–10 sterilized seeds per plate on four plates. Plates were sealed with micropore tape and placed in a dark 4 °C refrigerator for 2–4 days for cold stratification. The trial was repeated in triplicate for each metal lactate.
Following cold stratification, plates were transferred to a Percival Model CU22L growth chamber (~100 µmol m−2 s−1) where they were maintained at 24 °C. After the first 24 h of incubation, an initial mark was made on the exterior of the plate just over the top of the tip of the primary root (PR). Plate marking was repeated every 24 h for a total of 5 days after the initial mark to monitor daily PR growth. Plates were scanned using an Epson V700 flatbed scanner, and the scanned images were imported into ImageJ software (Java 8.0) to quantify daily PR growth.
2.7. Statistical Analyses
In the wheat cultivation vessel experiment, a t-test was used to identify differences between metal chloride and metal lactate treatments within each concentration for each plant growth parameter (Excel Version 2104, Microsoft, Redmond, WA, USA). A Fischer’s Least Significant Difference test was used to discern differences in nutrient content between concentrations within the same metal delivery treatment using the R statistical computing environment (R Development Core Team, 2017). Correlations between treatment and tissue metal concentrations were analyzed using a Pearson correlation test in the R statistical computing environment. In the plate-based assay experiment, differences in daily PR growth between metal lactate concentrations were assessed using a Tukey’s Honest Significant Difference test in the R statistical computing environment.
4. Discussion
This study provides the first investigation characterizing metal lactates as a potential plant biostimulant. To the authors’ knowledge, metal lactates have only previously been studied as a feed additive for ruminants to determine their effects on nutrient digestibility, in which Co lactate has been shown to enhance bacterial fiber digestion compared to CoCO
3 and promote microbial volatile fatty acid production in cattle [
42,
43]. The proven enhancement of microbial activity driven by metal lactates in the digestive tract of ruminants suggests a similar phenomenon might be observed in the soil ecosystem; however, there is no information on plant uptake or optimal concentration of lactate-bound metals. This information is not only essential for further studies on metal lactates, but it also provides baseline evidence that metal lactates have biostimulatory effects on plants. Furthermore, when characterizing novel crop biostimulants it is apropos to first define the plant response, especially for micronutrient-containing biostimulants, since the range between deficiency and toxicity in plant micronutrient levels tends to be narrow [
7]. The current study was conducted to address the dearth of information regarding metal lactate−plant interactions.
The first objective was to compare the uptake and toxicity range of five metals (Zn, Mn, Cu, Ni, and Co) supplied as either a lactate or a chloride salt form, to determine uptake in the absence of microbial breakdown processes using wheat grown in sterile culture vessels. Based on plant growth metrics and tissue metal content, metals supplied in lactate form appear to have a similar affinity for plant uptake compared to metals in chloride form. A previous study by Neocleous et al. (2020) found similar results when comparing the impact of chelated and inorganic forms of Mn and Zn on plant performance metrics and nutrient uptake in hydroponically grown bean crops [
44]. In their study, bean plants receiving Mn and Zn chelated with ethylenediaminetetra acetic acid (EDTA) showed negligible differences in tissue micronutrient content compared to those receiving sulfate mineral forms. Chelating agents such as EDTA are often used to enhance plant micronutrient uptake in both soil and soilless growing systems by protecting them from redox changes and/or precipitation, thereby making them more available in solution [
45]. Similarly, LMWOA (e.g., acetic, malic, and oxalic acid) also serve as small, organic complexing molecules and have been shown to increase heavy metal uptake in hydroponically grown wheat [
46,
47,
48]. Although the precise mechanism by which complexed metals are taken up by plant roots is still under investigation, the data indicate they do play a beneficial role in facilitating metal uptake in plants. Based on the work from Neocleous et al. (2020), as well as the findings from the present study, it appears that organometallic complexes may be as effective in delivering metal nutrients compared to mineral forms, at least in hydroponic systems [
44]. Further investigation is required to determine the efficacy of metal lactates to supply plants with metal nutrients in more complex soil systems.
A negative trend was observed between Mn and Zn where Mn tissue concentrations decreased with increasing ZnCl
2 and Zn lactate concentrations. Lewis et al. (2012) observed Zn treatment increased Mn translocation in nodulated
Medicago truncatula plants [
49], and others have observed that increased Zn treatment is associated with decreased Mn tissue concentrations in various plants including sunflower, wheat, and lemon balm (
Melissa officinalis) [
50,
51,
52]. In peanut (
Arachis hypogaea L.), Zn and Mn uptake are facilitated by the transporter AhNRAMP1, which could indicate that excessive Zn supply may suppress Mn uptake as a result of competition for transporters in root cells, thus providing a potential explanation for the findings presented above [
53]. Interestingly, our findings suggest there is a difference in translocation of Zn depending on the form in which it is supplied. This could indicate that Zn supplied in lactate form may be regulated by different transporters than Zn ions from chloride forms once inside the plant; however, further investigation is needed to elucidate potential differences and mechanisms involved in the translocation and partitioning of metals in plants when supplied these two forms.
To further investigate the physiological responses to metal lactates beyond uptake capacity and toxicity thresholds, we identified concentrations for each individual metal lactate that yielded optimal root growth in plate-grown
A. thaliana seedlings. This allowed us to gauge nutrient status and concentration as a result of plant uptake as has been described previously [
33]. Root growth measurements have frequently been used as an indicator for responsiveness to fertilizer treatments in a variety of plant species [
54,
55]. While root growth can be quantified in different ways (e.g., growth rates, density, and biomass), root growth promotion in any aspect generally correlates to increased plant vigor through numerous mechanisms including enhanced nutrient and water acquisition and heightened tolerance to abiotic and biotic stressors [
56,
57,
58]. Our finding that metal lactates delivered at specific concentrations can enhance primary root growth in
A. thaliana seedlings shows promise in their ability to promote overall plant health.
The ability of metal lactates to improve primary root growth in
A. thaliana seedlings may be attributed, at least in part, to enhanced activity in the enzymes and cellular processes that require micronutrients. Likewise, microbial growth and activity may also be stimulated by metal lactates by providing metal nutrients required for metabolic processes. For example, Zn in plants and microbes is primarily involved in the functionality and structural integrity of enzymes and proteins. Some of the more well-characterized plant enzymes and proteins requiring Zn include carbonic anhydrase, the Zn-containing superoxide dismutase (SOD), and ribosomal proteins, which have roles in C fixation, detoxification of reactive oxygen species, and protein synthesis, respectively [
59,
60,
61,
62,
63]. Zinc supplementation has been shown to enhance the activity of carbonic anhydrase [
61] and the Zn-containing SOD [
64], as well as increase protein synthesis by mitigating ribosomal disintegration [
65], and in each case plants receiving Zn had greater overall growth compared to Zn-deficient plants. Microbial alkaline phosphatase is particularly important in crop production settings due to its catalytic function in cleaving assimilable phosphate from organic P sources [
66]. Zinc is a cofactor required for microbial alkaline phosphatase activity; however, a report by Łukowski and Dec (2018) found acid-soluble and exchangeable fractions of Zn to be negatively correlated with alkaline phosphatase activity [
67]. Interestingly, the same study found a significant positive correlation between Zn and dehydrogenase activity, which is generally used as an indicator of overall biological activity [
68].
Manganese mainly serves as a cofactor, activating about 35 different enzymes primarily involved in oxidation-reduction, decarboxylation, and hydrolytic reactions [
7,
69]. For example, Mn is a cofactor for phenylalanine ammonia-lyase and peroxidase which are enzymes involved in lignin biosynthesis [
70]. Manganese deficiency is shown to reduce lignin content in plant roots [
71], which has been linked to decreased resistance against root-infecting pathogens due to the role of lignin in pathogen defense [
72]. Microbial Mn peroxidase also has an absolute requirement for Mn and is an ecologically important enzyme involved in the degradation of lignin, and thereby contributes to the recycling of more recalcitrant plant-derived C in soils [
73,
74]. The activity of Mn peroxidase appears to be enhanced by high concentrations of Mn which subsequently leads to greater lignin breakdown [
75].
Currently, more than 100 different Cu-containing proteins have been identified in plants including polyphenol oxidase and diamine oxidase [
76]. Polyphenol oxidases catalyze the oxygenation reactions converting plant phenols to orthoquinones, which are important precursors in the biosynthesis of lignin and in the production of melanotic substances that are formed from tissue damage in crops such as in potatoes and apples [
77]. In addition to the role of lignin in pathogen defense, melanotic compounds act as inhibitors of spore germination and fungal growth [
7]. Diamine oxidase is involved in the degradation of the nitrogen-containing compounds putrescine and spermidine, producing hydrogen peroxide and ammonia as byproducts of catalysis [
78]. Hydrogen peroxide produced during these reactions is thought to be involved in cell signaling, pathogen defense, and increasing the activity of peroxidases associated with lignification [
79,
80]. The activity of both polyphenol oxidase and diamine oxidase have shown clear responses to Cu supplementation, wherein Cu-sufficient plants had higher activities of the enzymes and greater plant biomass compared to plants grown in Cu-deficient conditions [
81]. In microbes, Cu is required by several enzymes including laccase and polysaccharide oxygenase which function in the breakdown of lignin and cellulose, respectively [
82,
83]. The activity of both enzymes can be enhanced by Cu supplementation in culture-based setups [
82,
84]; however, further investigation is needed to determine how Cu amendments may improve lignin and cellulose degradation in soils.
To date, only two plant proteins are known to require Ni for proper functioning: urease and Ni-urease accessory protein (Eu3). Urease catalyzes the hydrolytic reaction converting urea into ammonia and carbon dioxide, and Ni-urease accessory protein is essential for urease activity [
85,
86]. In studies using urea as the major or sole source of nitrogen, Ni-sufficient plants showed a significant increase in growth and urease activity compared to Ni-deficient plants [
87,
88,
89]. Likewise, Ni is required for urease function in microbes and the activity of the enzyme can be inhibited in Ni-deficient soils as well as stimulated upon Ni supplementation [
90].
Cobalt, although not required for plant growth, is considered a beneficial element that can enhance growth, particularly in leguminous plants, due to its requirement by nitrogen-fixing symbionts [
39]. For example, in
Rhizobium and
Bradyrhizobium, Co-dependent cobalamin is required for the synthesis of methyl malonyl-coA mutase which is involved in leghemoglobin synthesis in root nodules, wherein leghemoglobin is crucial to the protection of nitrogenase from oxygen [
91]. The absence of Co in nodule-forming plants can lead to reduced bacteroid numbers in root nodules and reduced nodule formation, ultimately decreasing nitrogen fixation [
7,
92]. This is particularly important in plants relying on microbial nitrogen fixation as a source of N when mineral or other forms are in low abundance or unavailable.
The importance of micronutrients in crop production is gaining awareness as micronutrient-deficient soils are becoming increasingly prominent across the globe [
5]. Generally, conventionally managed soils tend to have lower micronutrient availability compared to soils under organic management which can disrupt or inhibit plant metabolic processes. Organic cropping systems tend to procure sufficient levels of soil micronutrients which is likely due, in part, to the use of organic fertilizers and amendments that supply C to soil microbial communities. Organic C availability in soils is directly linked to soil microbial activity, wherein soils with high amounts of available C generally favor microbial growth and plant growth promoting functions such as P solubilization, siderophore production, phytohormone production, and biocontrol activities [
93,
94]. A variety of C amendments have been applied to soils with the goal of enhancing microbial functions including C supplied in the form of rhizospheric organic acids. In their report, Macias-Benitez et al. (2020) determined that organic acids, particularly lactic and citric acid, are effective in stimulating soil microbial activity and enhancing bacterial taxa described as plant growth promoting bacteria [
24]. Along with our work, the findings by Macias-Benitez et al. (2020) provide substantial evidence supporting the use of metal lactates as potential biostimulants.
In addition to serving as a C source of microbial nutrition, lactate is a key primary metabolite and may serve a beneficial role in early seedling growth and development. In our study, we identified several differences in plant growth between metal lactate and metal chloride treatments at similar concentrations, and in each case, metal lactates had less of an inhibitory effect (Tables 1, 3, 5, 7 and 9). For example, Ni lactate supplied at a rate of 0.343 mM led to significantly higher shoot FW compared to NiCl
2 treatments at the same concentration (
Table 7). Carbon sources are often used in in vitro plant cultures and serve as an energy and C source for highly demanding developmental processes such as embryogenesis, organogenesis, shoot proliferation, and root induction since developing seedlings grown in culture are not entirely autotrophic [
95]. The most widely used C source for growing in vitro plants is the disaccharide sucrose; however, a variety of other carbohydrates including other disaccharides, monosaccharides, trisaccharides, and sugar alcohols are also applied depending on the plant species and growth stage. Organic acids are typically not used for this purpose, but rather they have been incorporated in in vitro culture to facilitate metal nutrient uptake due to their role as a complexing agent. For example, lactic acid salts in hydroponic solution have been shown to significantly improve cucumber development and yield in nutrient-deficient conditions compared to non-deficient conditions without lactic acid salts [
22]. In addition to its role in enhancing metallonutrient uptake, the finding that Co lactate enhanced
A. thaliana root growth in our study suggests that lactate may fulfill the same role as carbohydrates to supply
A. thaliana seedlings with the C and energy, considering they were grown in nutrient sufficient conditions and the unlikelihood that Co supplementation benefitted growth and development.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}