
Interferon Signaling in the Endometrium and in Endometriosis


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
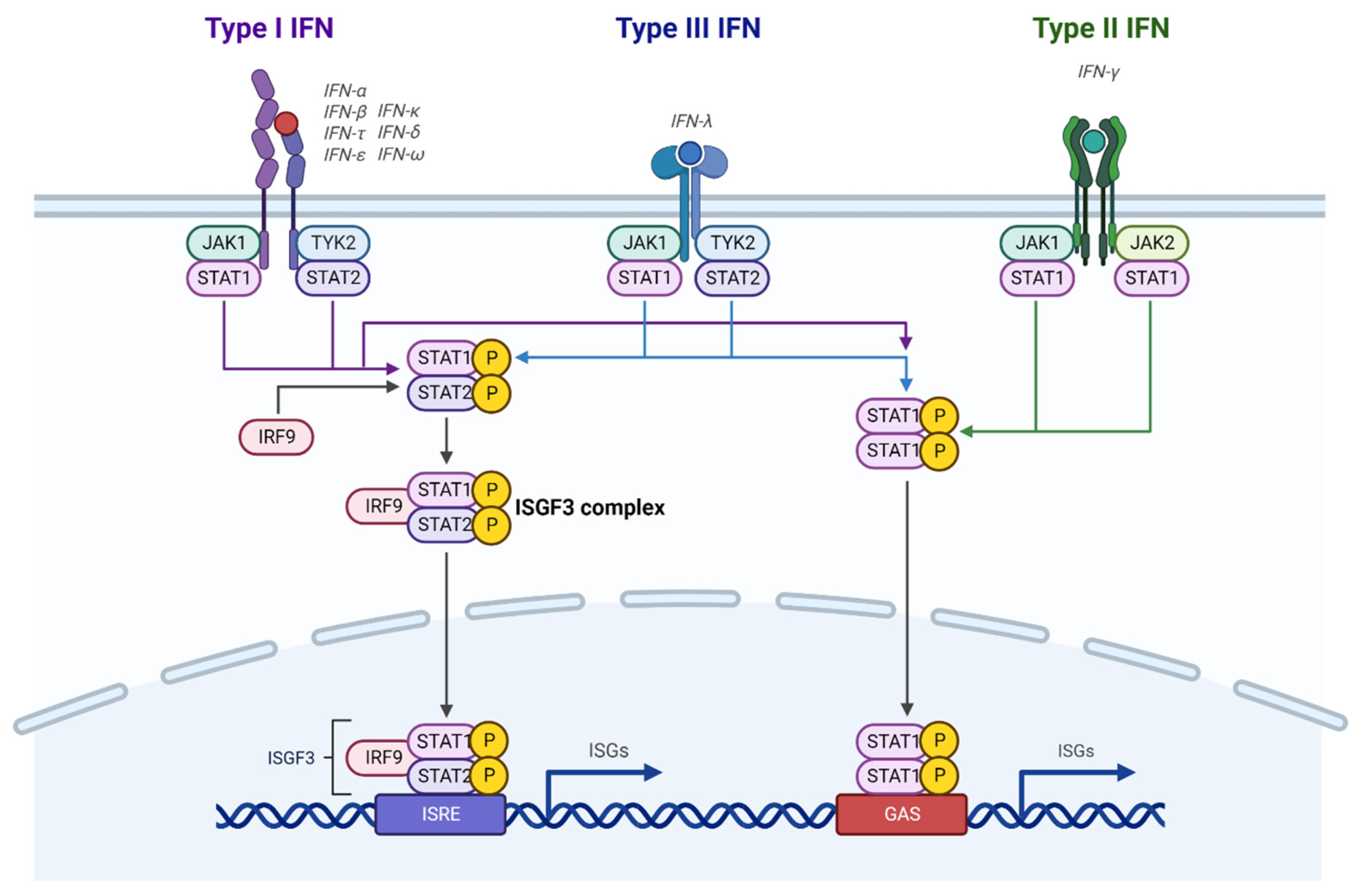
2. IFN Signaling
2.1. Types of IFNs
2.2. IFN Signaling
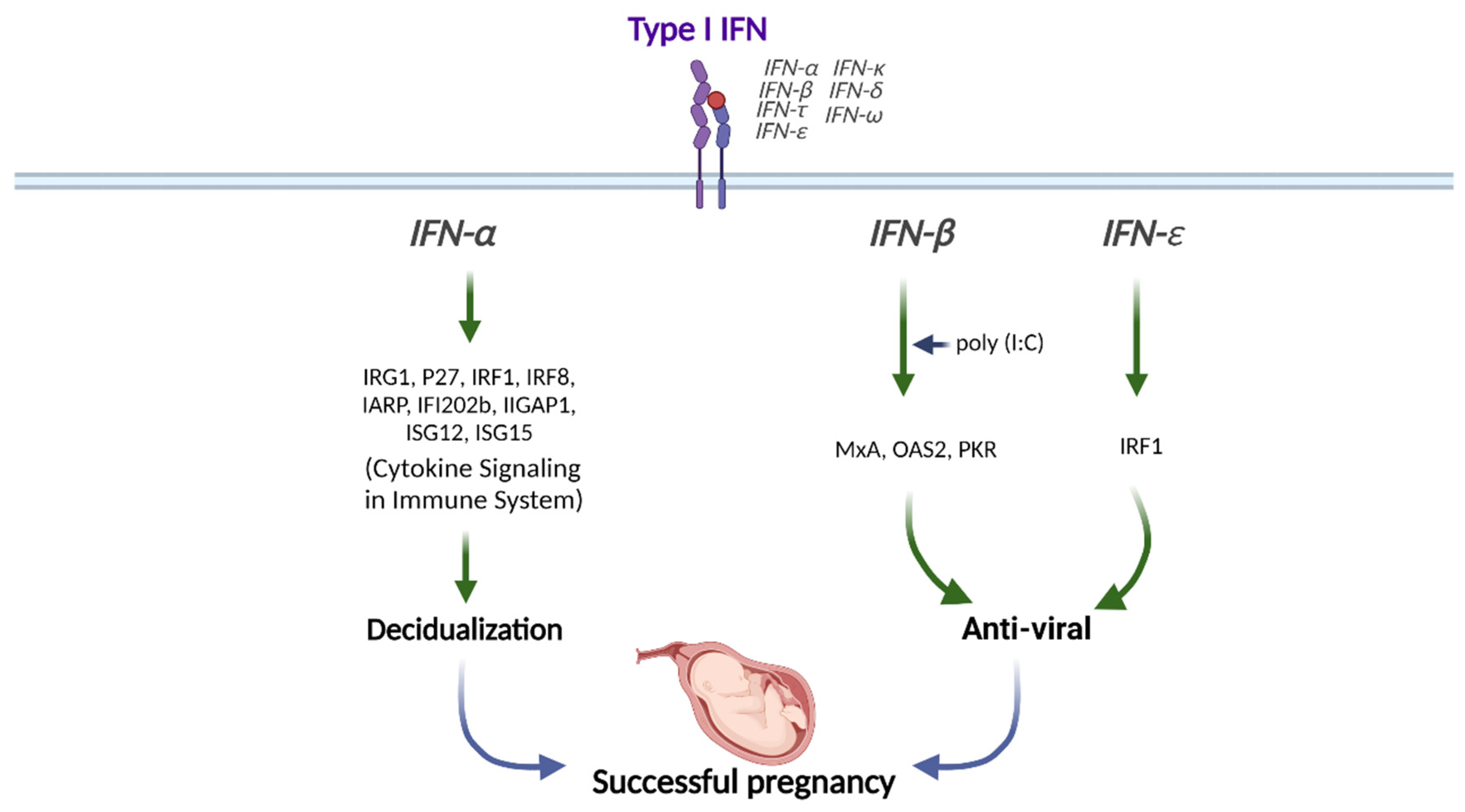
3. Roles of Interferons in Human Endometrial Function
3.1. Type I IFNs
3.2. Type II IFNs
3.3. Type III IFNs
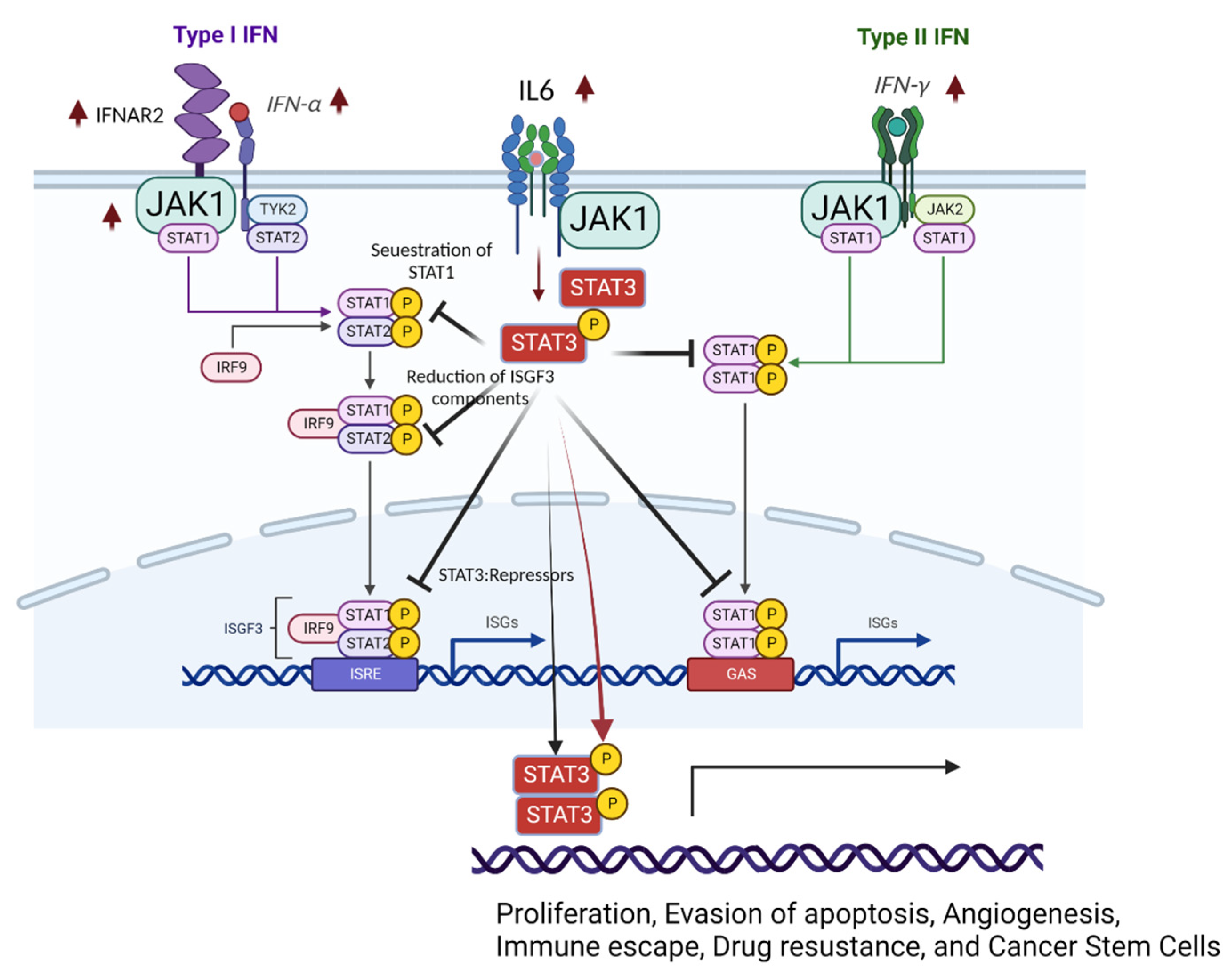
4. Dysregulation of Interferon Signaling in Endometriosis
4.1. Dysregulation of IFN Signaling in Endometriotic Tissues
4.2. Dysregulation of STAT3 in Endometriosis
4.3. Role of IFNs in Mesenchymal Stem Cells (MSCs)
5. The Interplay between Estrogen and IFN Signaling in Endometriosis Progression
6. Is IFN a Potential Target for Nonhormonal Endometriosis Treatment?
7. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zondervan, K.T.; Becker, C.M.; Missmer, S.A. Endometriosis. N. Engl. J. Med. 2020, 382, 1244–1256. [Google Scholar] [CrossRef] [PubMed]
- Nisolle, M.; Donnez, J. Peritoneal endometriosis, ovarian endometriosis, and adenomyotic nodules of the rectovaginal septum are three different entities. Fertil. Steril. 1997, 68, 585–596. [Google Scholar] [CrossRef]
- Cornillie, F.J.; Oosterlynck, D.; Lauweryns, J.M.; Koninckx, P.R. Deeply infiltrating pelvic endometriosis: Histology and clinical significance. Fertil. Steril. 1990, 53, 978–983. [Google Scholar] [CrossRef]
- Saunders, P.T.K.; Horne, A.W. Endometriosis: Etiology, pathobiology, and therapeutic prospects. Cell 2021, 184, 2807–2824. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.W. Recurrence of endometriosis and its control. Hum. Reprod. Update 2009, 15, 441–461. [Google Scholar] [CrossRef]
- Nirgianakis, K.; Ma, L.; McKinnon, B.; Mueller, M.D. Recurrence Patterns after Surgery in Patients with Different Endometriosis Subtypes: A Long-Term Hospital-Based Cohort Study. J. Clin. Med. 2020, 9, 496. [Google Scholar] [CrossRef] [Green Version]
- Chantalat, E.; Valera, M.C.; Vaysse, C.; Noirrit, E.; Rusidze, M.; Weyl, A.; Vergriete, K.; Buscail, E.; Lluel, P.; Fontaine, C.; et al. Estrogen Receptors and Endometriosis. Int. J. Mol. Sci. 2020, 21, 2815. [Google Scholar] [CrossRef] [Green Version]
- Struthers, R.S.; Nicholls, A.J.; Grundy, J.; Chen, T.; Jimenez, R.; Yen, S.S.; Bozigian, H.P. Suppression of gonadotropins and estradiol in premenopausal women by oral administration of the nonpeptide gonadotropin-releasing hormone antagonist elagolix. J. Clin. Endocrinol. Metab. 2009, 94, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Ambacher, K.; Secter, M.; Sanders, A.P. The use of progestin subdermal implants in the management of endometriosis-related pain symptoms and quality of life: A systematic review. Curr. Med. Res. Opin. 2022, 38, 479–486. [Google Scholar] [CrossRef]
- Ashfaq, S.; Can, A.S. Danazol. In StatPearls; Copyright © 2022, StatPearls Publishing LLC.: Treasure Island, FL, USA, 2022. [Google Scholar]
- Zakhari, A.; Delpero, E.; McKeown, S.; Tomlinson, G.; Bougie, O.; Murji, A. Endometriosis recurrence following post-operative hormonal suppression: A systematic review and meta-analysis. Hum. Reprod. Update 2021, 27, 96–107. [Google Scholar] [CrossRef]
- Tosti, C.; Biscione, A.; Morgante, G.; Bifulco, G.; Luisi, S.; Petraglia, F. Hormonal therapy for endometriosis: From molecular research to bedside. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 209, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E. Endometriosis. N. Engl. J. Med. 2009, 360, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Halme, J.; Hammond, M.G.; Hulka, J.F.; Raj, S.G.; Talbert, L.M. Retrograde menstruation in healthy women and in patients with endometriosis. Obstet. Gynecol. 1984, 64, 151–154. [Google Scholar]
- Treloar, S.A.; O’Connor, D.T.; O’Connor, V.M.; Martin, N.G. Genetic influences on endometriosis in an Australian twin sample. Fertil. Steril. 1999, 71, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Saha, R.; Pettersson, H.J.; Svedberg, P.; Olovsson, M.; Bergqvist, A.; Marions, L.; Tornvall, P.; Kuja-Halkola, R. Heritability of endometriosis. Fertil. Steril. 2015, 104, 947–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, J.V.; Perini, J.A.; Machado, D.E.; Pinto, R.; Medeiros, R. Systematic review of genome-wide association studies on susceptibility to endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 255, 74–82. [Google Scholar] [CrossRef]
- Hata, Y.; Nakaoka, H.; Yoshihara, K.; Adachi, S.; Haino, K.; Yamaguchi, M.; Nishikawa, N.; Kashima, K.; Yahata, T.; Tajima, A.; et al. A nonsynonymous variant of IL1A is associated with endometriosis in Japanese population. J. Hum. Genet. 2013, 58, 517–520. [Google Scholar] [CrossRef] [Green Version]
- Adachi, S.; Tajima, A.; Quan, J.; Haino, K.; Yoshihara, K.; Masuzaki, H.; Katabuchi, H.; Ikuma, K.; Suginami, H.; Nishida, N.; et al. Meta-analysis of genome-wide association scans for genetic susceptibility to endometriosis in Japanese population. J. Hum. Genet. 2010, 55, 816–821. [Google Scholar] [CrossRef] [Green Version]
- Tapmeier, T.T.; Rahmioglu, N.; Lin, J.; De Leo, B.; Obendorf, M.; Raveendran, M.; Fischer, O.M.; Bafligil, C.; Guo, M.; Harris, R.A.; et al. Neuropeptide S receptor 1 is a nonhormonal treatment target in endometriosis. Sci. Transl. Med. 2021, 13, eabd6469. [Google Scholar] [CrossRef]
- Laitinen, T.; Polvi, A.; Rydman, P.; Vendelin, J.; Pulkkinen, V.; Salmikangas, P.; Makela, S.; Rehn, M.; Pirskanen, A.; Rautanen, A.; et al. Characterization of a common susceptibility locus for asthma-related traits. Science 2004, 304, 300–304. [Google Scholar] [CrossRef]
- D’Amato, M.; Bruce, S.; Bresso, F.; Zucchelli, M.; Ezer, S.; Pulkkinen, V.; Lindgren, C.; Astegiano, M.; Rizzetto, M.; Gionchetti, P.; et al. Neuropeptide s receptor 1 gene polymorphism is associated with susceptibility to inflammatory bowel disease. Gastroenterology 2007, 133, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Pulkkinen, V.; Majuri, M.L.; Wang, G.; Holopainen, P.; Obase, Y.; Vendelin, J.; Wolff, H.; Rytila, P.; Laitinen, L.A.; Haahtela, T.; et al. Neuropeptide S and G protein-coupled receptor 154 modulate macrophage immune responses. Hum. Mol. Genet. 2006, 15, 1667–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundman, L.; Saarialho-Kere, U.; Vendelin, J.; Lindfors, K.; Assadi, G.; Kaukinen, K.; Westerholm-Ormio, M.; Savilahti, E.; Maki, M.; Alenius, H.; et al. Neuropeptide S receptor 1 expression in the intestine and skin—Putative role in peptide hormone secretion. Neurogastroenterol. Motil. 2010, 22, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Y.; Zhao, L.; Wang, L.; Wu, Z.; Mei, Q.; Nie, J.; Li, X.; Li, Y.; Fu, X.; et al. Whole-exome sequencing of endometriosis identifies frequent alterations in genes involved in cell adhesion and chromatin-remodeling complexes. Hum. Mol. Genet. 2014, 23, 6008–6021. [Google Scholar] [CrossRef] [Green Version]
- Oosterlynck, D.J.; Cornillie, F.J.; Waer, M.; Vandeputte, M.; Koninckx, P.R. Women with endometriosis show a defect in natural killer activity resulting in a decreased cytotoxicity to autologous endometrium. Fertil. Steril. 1991, 56, 45–51. [Google Scholar] [CrossRef]
- Xie, Q.; He, H.; Wu, Y.H.; Zou, L.J.; She, X.L.; Xia, X.M.; Wu, X.Q. Eutopic endometrium from patients with endometriosis modulates the expression of CD36 and SIRP-alpha in peritoneal macrophages. J. Obs. Gynaecol. Res. 2019, 45, 1045–1057. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H.; Chen, Y.H.; Chang, H.Y.; Au, H.K.; Tzeng, C.R.; Huang, Y.H. Chronic Niche Inflammation in Endometriosis-Associated Infertility: Current Understanding and Future Therapeutic Strategies. Int. J. Mol. Sci. 2018, 19, 2385. [Google Scholar] [CrossRef] [Green Version]
- Malutan, A.M.; Drugan, T.; Costin, N.; Ciortea, R.; Bucuri, C.; Rada, M.P.; Mihu, D. Pro-inflammatory cytokines for evaluation of inflammatory status in endometriosis. Cent. Eur. J. Immunol. 2015, 40, 96–102. [Google Scholar] [CrossRef]
- Li, Y.Z.; Wang, L.J.; Li, X.; Li, S.L.; Wang, J.L.; Wu, Z.H.; Gong, L.; Zhang, X.D. Vascular endothelial growth factor gene polymorphisms contribute to the risk of endometriosis: An updated systematic review and meta-analysis of 14 case-control studies. Genet. Mol. Res. GMR 2013, 12, 1035–1044. [Google Scholar] [CrossRef]
- Hayrabedyan, S.; Kyurkchiev, S.; Kehayov, I. FGF-1 and S100A13 possibly contribute to angiogenesis in endometriosis. J. Reprod. Immunol. 2005, 67, 87–101. [Google Scholar] [CrossRef]
- KhoshdelRad, N.; Salehi, Z.; Mashayekhi, F.; Abbasi, O.; Mirzajani, E. Soluble c-Met expression in the peritoneal fluid and serum of patients with different stages of endometriosis. Arch. Gynecol. Obstet. 2014, 289, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- May, K.E.; Conduit-Hulbert, S.A.; Villar, J.; Kirtley, S.; Kennedy, S.H.; Becker, C.M. Peripheral biomarkers of endometriosis: A systematic review. Hum. Reprod. Update 2010, 16, 651–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psarras, A.; Emery, P.; Vital, E.M. Type I interferon–mediated autoimmune diseases: Pathogenesis, diagnosis and targeted therapy. Rheumatology 2017, 56, 1662–1675. [Google Scholar] [CrossRef] [Green Version]
- Martensen, P.; Vestergaard, A.; Knudsen, U. Virus Infection and Type I Interferon in Endometriosis. Endometr.-Basic Concepts Curr. Res. Trends 2012, 245–262. [Google Scholar] [CrossRef] [Green Version]
- Nishida, M.; Nasu, K.; Ueda, T.; Fukuda, J.; Takai, N.; Miyakawa, I. Endometriotic cells are resistant to interferon-gamma-induced cell growth inhibition and apoptosis: A possible mechanism involved in the pathogenesis of endometriosis. Mol. Hum. Reprod. 2005, 11, 29–34. [Google Scholar] [CrossRef]
- Herington, J.L.; Bruner-Tran, K.L.; Lucas, J.A.; Osteen, K.G. Immune interactions in endometriosis. Expert Rev. Clin. Immunol. 2011, 7, 611–626. [Google Scholar] [CrossRef] [Green Version]
- Casazza, R.L.; Lazear, H.M.; Miner, J.J. Protective and Pathogenic Effects of Interferon Signaling During Pregnancy. Viral Immunol. 2020, 33, 3–11. [Google Scholar] [CrossRef]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef] [Green Version]
- Platanias, L.C. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nat. Rev. Immunol. 2005, 5, 375–386. [Google Scholar] [CrossRef]
- Wack, A.; Terczynska-Dyla, E.; Hartmann, R. Guarding the frontiers: The biology of type III interferons. Nat. Immunol. 2015, 16, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Prchal-Murphy, M.; Semper, C.; Lassnig, C.; Wallner, B.; Gausterer, C.; Teppner-Klymiuk, I.; Kobolak, J.; Müller, S.; Kolbe, T.; Karaghiosoff, M.; et al. TYK2 kinase activity is required for functional type I interferon responses in vivo. PloS One 2012, 7, e39141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazewski, C.; Perez, R.E.; Fish, E.N.; Platanias, L.C. Type I Interferon (IFN)-Regulated Activation of Canonical and Non-Canonical Signaling Pathways. Front. Immunol. 2020, 11, 606456. [Google Scholar] [CrossRef] [PubMed]
- Critchley, H.O.D.; Maybin, J.A.; Armstrong, G.M.; Williams, A.R.W. Physiology of the Endometrium and Regulation of Menstruation. Physiol. Rev. 2020, 100, 1149–1179. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.P.; Tayade, C.; Ashkar, A.A.; Hatta, K.; Zhang, J.; Croy, B.A. Interferon gamma in successful pregnancies. Biol. Reprod. 2009, 80, 848–859. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Kurohmaru, M.; Hayashi, Y. Localization of type I interferon in murine trophoblast and decidua during decidual formation. J. Reprod. Fertil. 1992, 95, 559–565. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.P.; Warnock, S.H.; Urry, R.L.; Edwin, S.S.; Mitchell, M.D. Immunosuppressive activity and alpha interferon concentrations in human embryo culture media as an index of potential for successful implantation. Fertil. Steril. 1992, 57, 637–640. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, M.; Kumar, S.; Zhu, L.J.; Chen, D.; Bagchi, M.K.; Bagchi, I.C. Identification and implantation stage-specific expression of an interferon-alpha-regulated gene in human and rat endometrium. Endocrinology 2001, 142, 2390–2400. [Google Scholar] [CrossRef]
- Kitaya, K.; Yasuda, J.; Fushiki, S.; Honjo, H. Localization of interferon regulatory factor-1 in human endometrium throughout the menstrual cycle. Fertil. Steril. 2001, 75, 992–996. [Google Scholar] [CrossRef]
- Jabbour, H.N.; Critchley, H.O.; Yu-Lee, L.Y.; Boddy, S.C. Localization of interferon regulatory factor-1 (IRF-1) in nonpregnant human endometrium: Expression of IRF-1 is up-regulated by prolactin during the secretory phase of the menstrual cycle. J. Clin. Endocrinol. Metab. 1999, 84, 4260–4265. [Google Scholar] [CrossRef]
- Ozaki, T.; Takahashi, K.; Kanasaki, H.; Iida, K.; Miyazaki, K. Expression of the type I interferon receptor and the interferon-induced Mx protein in human endometrium during the menstrual cycle. Fertil. Steril. 2005, 83, 163–170. [Google Scholar] [CrossRef]
- Kashiwagi, A.; DiGirolamo, C.M.; Kanda, Y.; Niikura, Y.; Esmon, C.T.; Hansen, T.R.; Shioda, T.; Pru, J.K. The postimplantation embryo differentially regulates endometrial gene expression and decidualization. Endocrinology 2007, 148, 4173–4184. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.V.; Ghosh, M.; Fahey, J.V.; Wira, C.R. Uterine epithelial cells specifically induce interferon-stimulated genes in response to polyinosinic-polycytidylic acid independently of estradiol. PloS One 2012, 7, e35654. [Google Scholar] [CrossRef] [Green Version]
- Bourke, N.M.; Achilles, S.L.; Huang, S.U.; Cumming, H.E.; Lim, S.S.; Papageorgiou, I.; Gearing, L.J.; Chapman, R.; Thakore, S.; Mangan, N.E.; et al. Spatiotemporal regulation of human IFNepsilon and innate immunity in the female reproductive tract. JCI Insight 2022, 7, e135407. [Google Scholar] [CrossRef]
- Fung, K.Y.; Mangan, N.E.; Cumming, H.; Horvat, J.C.; Mayall, J.R.; Stifter, S.A.; De Weerd, N.; Roisman, L.C.; Rossjohn, J.; Robertson, S.A.; et al. Interferon-epsilon protects the female reproductive tract from viral and bacterial infection. Science 2013, 339, 1088–1092. [Google Scholar] [CrossRef] [Green Version]
- Nickodem, C.; Criscitiello, M.F.; Bazer, F.; Abiodun-Ojo, O.; Taylor, B.D. Interferon epsilon in the reproductive tract of healthy and genital herpes simplex virus-infected pregnant women: Results of a pilot study. Am. J. Reprod. Immunol. 2018, 80, e12995. [Google Scholar] [CrossRef]
- Miller, D.; Romero, R.; Kacerovsky, M.; Musilova, I.; Galaz, J.; Garcia-Flores, V.; Xu, Y.; Pusod, E.; Demery-Poulos, C.; Gutierrez-Contreras, P.; et al. Defining a role for Interferon Epsilon in normal and complicated pregnancies. Heliyon 2022, 8, e09952. [Google Scholar] [CrossRef]
- Paulesu, L.; Romagnoli, R.; Cintorino, M.; Ricci, M.G.; Garotta, G. First trimester human trophoblast expresses both interferon-gamma and interferon-gamma-receptor. J. Reprod. Immunol. 1994, 27, 37–48. [Google Scholar] [CrossRef]
- Hess, A.P.; Hamilton, A.E.; Talbi, S.; Dosiou, C.; Nyegaard, M.; Nayak, N.; Genbecev-Krtolica, O.; Mavrogianis, P.; Ferrer, K.; Kruessel, J.; et al. Decidual stromal cell response to paracrine signals from the trophoblast: Amplification of immune and angiogenic modulators. Biol. Reprod. 2007, 76, 102–117. [Google Scholar] [CrossRef] [Green Version]
- Platt, J.S.; Hunt, J.S. Interferon-gamma gene expression in cycling and pregnant mouse uterus: Temporal aspects and cellular localization. J. Leukoc. Biol. 1998, 64, 393–400. [Google Scholar] [CrossRef]
- Seaward, A.V.; Burke, S.D.; Croy, B.A. Interferon gamma contributes to preimplantation embryonic development and to implantation site structure in NOD mice. Hum. Reprod. (Oxf. Engl.) 2010, 25, 2829–2839. [Google Scholar] [CrossRef]
- Saito, S.; Nishikawa, K.; Morii, T.; Enomoto, M.; Narita, N.; Motoyoshi, K.; Ichijo, M. Cytokine production by CD16-CD56bright natural killer cells in the human early pregnancy decidua. Int. Immunol. 1993, 5, 559–563. [Google Scholar] [CrossRef]
- Tabibzadeh, S. Evidence of T-cell activation and potential cytokine action in human endometrium. J. Clin. Endocrinol. Metab. 1990, 71, 645–649. [Google Scholar] [CrossRef]
- Ashkar, A.A.; Di Santo, J.P.; Croy, B.A. Interferon gamma contributes to initiation of uterine vascular modification, decidual integrity, and uterine natural killer cell maturation during normal murine pregnancy. J. Exp. Med. 2000, 192, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.V.; Hopkins, D.C.; Barr, F.D.; Wira, C.R. Sex Hormones and Aging Modulate Interferon Lambda 1 Production and Signaling by Human Uterine Epithelial Cells and Fibroblasts. Front. Immunol. 2021, 12, 718380. [Google Scholar] [CrossRef]
- Dicitore, A.; Castiglioni, S.; Saronni, D.; Gentilini, D.; Borghi, M.O.; Stabile, S.; Vignali, M.; Di Blasio, A.M.; Persani, L.; Vitale, G. Effects of human recombinant type I IFNs (IFN-alpha2b and IFN-beta1a) on growth and migration of primary endometrial stromal cells from women with deeply infiltrating endometriosis: A preliminary study. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 230, 192–198. [Google Scholar] [CrossRef]
- Kao, L.C.; Germeyer, A.; Tulac, S.; Lobo, S.; Yang, J.P.; Taylor, R.N.; Osteen, K.; Lessey, B.A.; Giudice, L.C. Expression profiling of endometrium from women with endometriosis reveals candidate genes for disease-based implantation failure and infertility. Endocrinology 2003, 144, 2870–2881. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, S.; Canis, M.; Pouly, J.L.; Botchorishvili, R.; Dechelotte, P.J.; Mage, G. Differential expression of genes in eutopic and ectopic endometrium from patients with ovarian endometriosis. Fertil. Steril. 2006, 86, 548–553. [Google Scholar] [CrossRef]
- Ingelmo, J.M.; Quereda, F.; Acien, P. Intraperitoneal and subcutaneous treatment of experimental endometriosis with recombinant human interferon-alpha-2b in a murine model. Fertil. Steril. 1999, 71, 907–911. [Google Scholar] [CrossRef]
- Acien, P.; Quereda, F.; Campos, A.; Gomez-Torres, M.J.; Velasco, I.; Gutierrez, M. Use of intraperitoneal interferon alpha-2b therapy after conservative surgery for endometriosis and postoperative medical treatment with depot gonadotropin-releasing hormone analog: A randomized clinical trial. Fertil. Steril. 2002, 78, 705–711. [Google Scholar] [CrossRef]
- Podgaec, S.; Abrao, M.S.; Dias, J.A., Jr.; Rizzo, L.V.; de Oliveira, R.M.; Baracat, E.C. Endometriosis: An inflammatory disease with a Th2 immune response component. Hum. Reprod. (Oxf. Engl.) 2007, 22, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.H.; Yang, B.C.; Hsu, C.C.; Lee, Y.C.; Huang, K.E. The expression of soluble intercellular adhesion molecule-1 in endometriosis. Fertil. Steril. 1998, 70, 1139–1142. [Google Scholar] [CrossRef]
- Wu, M.H.; Yang, B.C.; Lee, Y.C.; Wu, P.L.; Hsu, C.C. The differential expression of intercellular adhesion molecule-1 (ICAM-1) and regulation by interferon-gamma during the pathogenesis of endometriosis. Am. J. Reprod. Immunol. 2004, 51, 373–380. [Google Scholar] [CrossRef]
- Delbandi, A.A.; Mahmoudi, M.; Shervin, A.; Heidari, S.; Kolahdouz-Mohammadi, R.; Zarnani, A.H. Evaluation of apoptosis and angiogenesis in ectopic and eutopic stromal cells of patients with endometriosis compared to non-endometriotic controls. BMC Womens Health 2020, 20, 3. [Google Scholar] [CrossRef]
- Tsai, M.H.; Pai, L.M.; Lee, C.K. Fine-Tuning of Type I Interferon Response by STAT3. Front. Immunol. 2019, 10, 1448. [Google Scholar] [CrossRef] [Green Version]
- Ho, H.H.; Ivashkiv, L.B. Role of STAT3 in type I interferon responses. Negative regulation of STAT1-dependent inflammatory gene activation. J. Biol. Chem. 2006, 281, 14111–14118. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.H.; Lee, C.K. STAT3 Cooperates With Phospholipid Scramblase 2 to Suppress Type I Interferon Response. Front. Immunol. 2018, 9, 1886. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Zhu, F.; Zhang, M.; Li, Y.; Drennan, A.C.; Kimpara, S.; Rumball, I.; Selzer, C.; Cameron, H.; Kellicut, A.; et al. Gene regulation and suppression of type I interferon signaling by STAT3 in diffuse large B cell lymphoma. Proc. Natl. Acad. Sci. USA 2018, 115, E498–E505. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R.L.; Sharma, A.; Bai, S.; Heneidi, S.; Lee, T.J.; Kodeboyina, S.K.; Patel, N.; Sharma, S. Comparative STAT3-Regulated Gene Expression Profile in Renal Cell Carcinoma Subtypes. Front. Oncol. 2019, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.G.; Yoo, J.Y.; Kim, T.H.; Shin, J.H.; Langenheim, J.F.; Ferguson, S.D.; Fazleabas, A.T.; Young, S.L.; Lessey, B.A.; Jeong, J.W. Aberrant activation of signal transducer and activator of transcription-3 (STAT3) signaling in endometriosis. Hum. Reprod. (Oxf. Engl.) 2015, 30, 1069–1078. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.Y.; Jeong, J.W.; Fazleabas, A.T.; Tayade, C.; Young, S.L.; Lessey, B.A. Protein Inhibitor of Activated STAT3 (PIAS3) Is Down-Regulated in Eutopic Endometrium of Women with Endometriosis. Biol. Reprod. 2016, 95, 11. [Google Scholar] [CrossRef] [PubMed]
- Kashanian, M.; Sariri, E.; Vahdat, M.; Ahmari, M.; Moradi, Y.; Sheikhansari, N. A comparison between serum levels of interleukin-6 and CA125 in patients with endometriosis and normal women. Med. J. Islam. Repub. Iran. 2015, 29, 280. [Google Scholar] [PubMed]
- Wang, Y.; van Boxel-Dezaire, A.H.; Cheon, H.; Yang, J.; Stark, G.R. STAT3 activation in response to IL-6 is prolonged by the binding of IL-6 receptor to EGF receptor. Proc. Natl. Acad. Sci. USA 2013, 110, 16975–16980. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, S.; Pouly, J.L.; Canis, M. Persistent activation of signal transducer and activator of transcription 3 via interleukin-6 trans-signaling is involved in fibrosis of endometriosis. Hum. Reprod. (Oxf. Engl.) 2022, 37, 1489–1504. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nie, X.B.; Liu, S.J.; Liu, J.; Bian, W.H. Curcumol Attenuates Endometriosis by Inhibiting the JAK2/STAT3 Signaling Pathway. Med. Sci. Monit. 2022, 28, e934914. [Google Scholar] [CrossRef]
- Kotlyar, A.M.; Mamillapalli, R.; Flores, V.A.; Taylor, H.S. Tofacitinib alters STAT3 signaling and leads to endometriosis lesion regression. Mol. Hum. Reprod. 2021, 27, gaab016. [Google Scholar] [CrossRef]
- Maruyama, T. A Revised Stem Cell Theory for the Pathogenesis of Endometriosis. J. Pers. Med. 2022, 12, 216. [Google Scholar] [CrossRef] [PubMed]
- Sourial, S.; Tempest, N.; Hapangama, D.K. Theories on the pathogenesis of endometriosis. Int. J. Reprod. Med. 2014, 2014, 179515. [Google Scholar] [CrossRef] [Green Version]
- Leyendecker, G.; Herbertz, M.; Kunz, G.; Mall, G. Endometriosis results from the dislocation of basal endometrium. Hum. Reprod. (Oxf. Engl.) 2002, 17, 2725–2736. [Google Scholar] [CrossRef] [Green Version]
- Kao, A.P.; Wang, K.H.; Chang, C.C.; Lee, J.N.; Long, C.Y.; Chen, H.S.; Tsai, C.F.; Hsieh, T.H.; Tsai, E.M. Comparative study of human eutopic and ectopic endometrial mesenchymal stem cells and the development of an in vivo endometriotic invasion model. Fertil. Steril. 2011, 95, 1308–1315. [Google Scholar] [CrossRef]
- Sahraei, S.S.; Davoodi Asl, F.; Kalhor, N.; Sheykhhasan, M.; Fazaeli, H.; Moud, S.S.; Sheikholeslami, A. A Comparative Study of Gene Expression in Menstrual Blood-Derived Stromal Cells between Endometriosis and Healthy Women. BioMed. Res. Int. 2022, 2022, 7053521. [Google Scholar] [CrossRef] [PubMed]
- Abomaray, F.; Gidlof, S.; Gotherstrom, C. Mesenchymal Stromal Cells Are More Immunosuppressive In Vitro If They Are Derived from Endometriotic Lesions than from Eutopic Endometrium. Stem Cells Int. 2017, 2017, 3215962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.S.; Jang, I.K.; Lee, M.W.; Ko, Y.J.; Lee, D.H.; Lee, J.W.; Sung, K.W.; Koo, H.H.; Yoo, K.H. Enhanced Immunosuppressive Properties of Human Mesenchymal Stem Cells Primed by Interferon-γ. EBioMedicine 2018, 28, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Guan, Q.; Ezzati, P.; Spicer, V.; Krokhin, O.; Wall, D.; Wilkins, J.A. Interferon γ induced compositional changes in human bone marrow derived mesenchymal stem/stromal cells. Clin. Proteom. 2017, 14, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meligy, F.Y.; Elgamal, D.A.; Abdelzaher, L.A.; Khashbah, M.Y.; El-Mokhtar, M.A.; Sayed, A.A.; Refaiy, A.M.; Othman, E.R. Adipose tissue-derived mesenchymal stem cells reduce endometriosis cellular proliferation through their anti-inflammatory effects. Clin. Exp. Reprod. Med. 2021, 48, 322–336. [Google Scholar] [CrossRef]
- Hirakawa, T.; Yotsumoto, F.; Shirasu, N.; Kiyoshima, C.; Urushiyama, D.; Yoshikawa, K.; Miyata, K.; Kurakazu, M.; Koga, K.A.; Aoki, M.; et al. Trophic and immunomodulatory effects of adipose tissue derived stem cells in a preclinical murine model of endometriosis. Sci. Rep. 2022, 12, 8031. [Google Scholar] [CrossRef]
- Abomaray, F.; Gidlof, S.; Bezubik, B.; Engman, M.; Gotherstrom, C. Mesenchymal Stromal Cells Support Endometriotic Stromal Cells In Vitro. Stem Cells Int. 2018, 2018, 7318513. [Google Scholar] [CrossRef] [Green Version]
- Lupatov, A.Y.; Saryglar, R.Y.; Vtorushina, V.V.; Poltavtseva, R.A.; Bystrykh, O.A.; Chuprynin, V.D.; Krechetova, L.V.; Pavlovich, S.V.; Yarygin, K.N.; Sukhikh, G.T. Mesenchymal Stromal Cells Isolated from Ectopic but Not Eutopic Endometrium Display Pronounced Immunomodulatory Activity In Vitro. Biomedicines 2021, 9, 1286. [Google Scholar] [CrossRef]
- Guan, Q.; Li, Y.; Shpiruk, T.; Bhagwat, S.; Wall, D.A. Inducible indoleamine 2,3-dioxygenase 1 and programmed death ligand 1 expression as the potency marker for mesenchymal stromal cells. Cytotherapy 2018, 20, 639–649. [Google Scholar] [CrossRef]
- De Pedro, M.A.; Gomez-Serrano, M.; Marinaro, F.; Lopez, E.; Pulido, M.; Preusser, C.; Pogge von Strandmann, E.; Sanchez-Margallo, F.M.; Alvarez, V.; Casado, J.G. IFN-Gamma and TNF-Alpha as a Priming Strategy to Enhance the Immunomodulatory Capacity of Secretomes from Menstrual Blood-Derived Stromal Cells. Int. J. Mol. Sci. 2021, 22, 12177. [Google Scholar] [CrossRef]
- Bulun, S.E.; Cheng, Y.H.; Pavone, M.E.; Xue, Q.; Attar, E.; Trukhacheva, E.; Tokunaga, H.; Utsunomiya, H.; Yin, P.; Luo, X.; et al. Estrogen receptor-beta, estrogen receptor-alpha, and progesterone resistance in endometriosis. Semin. Reprod. Med. 2010, 28, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Han, S.J.; Jung, S.Y.; Wu, S.; Hawkins, S.M.; Park, M.J.; Kyo, S.; Qin, J.; Lydon, J.P.; Tsai, S.Y.; Tsai, M.J.; et al. Estrogen Receptor β modulates apoptosis complexes and the inflammasome to drive the pathogenesis of endometriosis. Cell 2015, 163, 960–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Q.; Lin, Z.; Cheng, Y.H.; Huang, C.C.; Marsh, E.; Yin, P.; Milad, M.P.; Confino, E.; Reierstad, S.; Innes, J.; et al. Promoter methylation regulates estrogen receptor 2 in human endometrium and endometriosis. Biol. Reprod. 2007, 77, 681–687. [Google Scholar] [CrossRef]
- Han, S.J.; Lee, J.E.; Cho, Y.J.; Park, M.J.; O’Malley, B.W. Genomic Function of Estrogen Receptor beta in Endometriosis. Endocrinology 2019, 160, 2495–2516. [Google Scholar] [CrossRef] [PubMed]
- De Ziegler, D.; Borghese, B.; Chapron, C. Gonadotropin-releasing hormone agonists for endometriosis. N. Engl. J. Med. 2008, 359, 2844. [Google Scholar] [CrossRef]
- Słopień, R.; Męczekalski, B. Aromatase inhibitors in the treatment of endometriosis. Prz. Menopauzalny 2016, 15, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Mehta, J.; Kling, J.M.; Manson, J.E. Risks, Benefits, and Treatment Modalities of Menopausal Hormone Therapy: Current Concepts. Front. Endocrinol. 2021, 12, 564781. [Google Scholar] [CrossRef]
- Badawy, S.Z.; Etman, A.; Cuenca, V.; Montante, A.; Kaufman, L. Effect of interferon alpha-2b on endometrioma cells in vitro. Obstet. Gynecol. 2001, 98, 417–420. [Google Scholar] [CrossRef]
- Ingelmo, J.M.; Quereda, F.; Acien, P. Effect of human interferon-alpha-2b on experimental endometriosis in rats: Comparison between short and long series of treatment. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 167, 190–193. [Google Scholar] [CrossRef]
- Ali, A.F.M.; Fateen, B.; Ezzet, A.; Badawy, H.; Ramadan, A.; El-tobge, A. Laparoscopic intraperitoneal injection of human interferon-α2b in the treatment of pelvic endometriosis: A new modality. Obstet. Gynecol. 2000, 95, S47–S48. [Google Scholar] [CrossRef]
- McComb, S.; Cessford, E.; Alturki, N.A.; Joseph, J.; Shutinoski, B.; Startek, J.B.; Gamero, A.M.; Mossman, K.L.; Sad, S. Type-I interferon signaling through ISGF3 complex is required for sustained Rip3 activation and necroptosis in macrophages. Proc. Natl. Acad. Sci. USA 2014, 111, E3206–E3213. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Xu, X.; Li, T.; Yu, Q.; Wang, J.; Chen, Y.; Ding, S.; Zhu, L.; Zou, G.; Zhang, X. NLRP3 Inflammasome Activation of Mast Cells by Estrogen via the Nuclear-Initiated Signaling Pathway Contributes to the Development of Endometriosis. Front. Immunol. 2021, 12, 749979. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, J.; Zhang, X. An Update on the Multifaceted Role of NF-kappaB in Endometriosis. Int. J. Biol. Sci. 2022, 18, 4400–4413. [Google Scholar] [CrossRef] [PubMed]
- Koninckx, P.R.; Craessaerts, M.; Timmerman, D.; Cornillie, F.; Kennedy, S. Anti-TNF-alpha treatment for deep endometriosis-associated pain: A randomized placebo-controlled trial. Hum. Reprod. 2008, 23, 2017–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, D.; Song, H.; Shi, G. Anti-TNF-α treatment for pelvic pain associated with endometriosis. Cochrane Database Syst. Rev. 2013, 3, Cd008088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grivennikov, S.I.; Karin, M. Dangerous liaisons: STAT3 and NF-kappaB collaboration and crosstalk in cancer. Cytokine Growth Factor Rev. 2010, 21, 11–19. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.; Han, S.J. Interferon Signaling in the Endometrium and in Endometriosis. Biomolecules 2022, 12, 1554. https://doi.org/10.3390/biom12111554
Park Y, Han SJ. Interferon Signaling in the Endometrium and in Endometriosis. Biomolecules. 2022; 12(11):1554. https://doi.org/10.3390/biom12111554
Chicago/Turabian StylePark, Yuri, and Sang Jun Han. 2022. "Interferon Signaling in the Endometrium and in Endometriosis" Biomolecules 12, no. 11: 1554. https://doi.org/10.3390/biom12111554
APA StylePark, Y., & Han, S. J. (2022). Interferon Signaling in the Endometrium and in Endometriosis. Biomolecules, 12(11), 1554. https://doi.org/10.3390/biom12111554