Abstract
Common uterine diseases include endometriosis, uterine fibroids, endometrial polyps, endometrial hyperplasia, endometrial cancer, and endometrial dysfunction causing infertility. Patients with uterine diseases often suffer from abdominal pain, menorrhagia, infertility and other symptoms, which seriously impair their health and disturb their lives. Androgens play important roles in the normal physiological functions of the uterus and pathological progress of uterine diseases. Androgens in women are synthesized in the ovaries and adrenal glands. The action of androgens in the uterus is mainly mediated by its ligand androgen receptor (AR) that regulates transcription of the target genes. However, much less is known about the signaling pathways through which androgen functions in uterine diseases, and contradictory findings have been reported. This review summarizes and discusses the progress of research on androgens and the involvement of AR in uterine diseases. Future studies should focus on developing new therapeutic strategies that precisely target specific AR and their related signaling pathways in uterine diseases.
1. Introduction
The uterus, as an important reproductive organ of women, is extremely sensitive to the actions of sex steroids. There are a variety of uterine diseases, including endometriosis, fibroids, endometrial polyps, endometrial hyperplasia (EH), endometrial cancer (EC), and infertility due to endometrial dysfunction. Androgens play an important role in the regulation of uterus function in health and disease. Dehydroepiandrosterone sulfate (DHEAS), dehydroepiandrosterone (DHEA), androstenedione (A4), testosterone (T), and dihydrotestosterone (DHT) are the most common androgens in women [1]. The first three kinds of androgens cannot play their role directly. They must be transformed into T and DHT in the target organ before binding to the androgen receptor (AR) [2]. There is growing evidence that androgens regulate important functions of the uterus, including endometrial proliferation, preparation for pregnancy, and tissue repair during menstruation. Dysregulation of androgen action is associated with endometrial diseases such as endometriosis and endometrial cancer and may cause infertility in women due to endometrial dysfunction [3]. Compared with the effects of progesterone and estrogen on the physiological and pathological uterus, limited information on the roles of androgens in uterus lesion is available, and contradictory findings have also been reported. This article discusses the recent research progress of androgens in uterine diseases and explores the potential of AR as a therapeutic target for treating uterine diseases.
2. Androgen and Androgen Receptor
2.1. Metabolism and Biological Activity of Androgen
In females, androgens are mainly produced by the ovaries and adrenal glands, which are controlled by gonadotropin (Gn) and adrenocorticotropic hormone (ACTH). DHEA and T are regarded as indicators of androgen secretion by the adrenal glands and ovaries, respectively. After menopause, estrogen production is markedly reduced, but the ovaries continue to produce androgens, including DHEA, A4, and T. In postmenopausal women, DHEA is the main source of androgens and estrogens. Approximately 20% of circulating DHEA is produced by the ovaries, and the remaining 80% comes from the adrenal glands [4]. DHEA is converted to DHEAS by the enzyme steroid sulfotransferase type 2A1 (SULT2A1), or to A4 by the enzyme 3β-hydroxysteroid dehydrogenase type 2 (HSD3B2) [5]. A4 is further converted to T under the action of 17β-hydroxysteroid dehydrogenase type 5 (17β-HSD type 5), also known as aldo-keto reductase family 1 member 3 (AKR1C3) [6]. Finally, T is converted to DHT by the action of 5α-reductase (Figure 1). The formed DHEAS, DHEA, A4, and T enter the systemic circulation, of which the latter three are mainly bound to sex hormone-binding globulin (SHBG). In women, 80% of androgens bind to SHBG, 19% bind to serum albumin, and only 1% are free. Free androgens are the only active androgens. The rest of the bound androgens are used as a reservoir of circulating androgens. Biologically active androgens, such as T and DHT, exert their effects primarily by binding to AR to regulate the expression of downstream target genes. In addition, androgens are converted by aromatase to estrogens, which act primarily through the estrogen receptor (ER). A4 and T are converted to estrone (E1) and estradiol (E2), respectively [7,8].
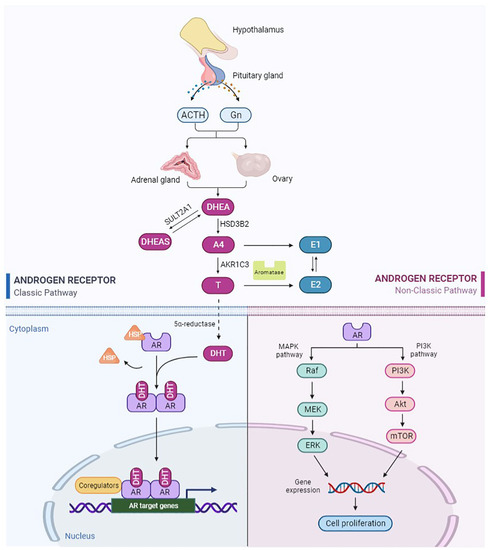
Figure 1.
Schematic of androgen production, metabolism, and classical and nonclassical signaling pathways of AR. The figure was created with https://biorender.com (accessed on 1 September 2022).
2.2. Structure and Function of AR
AR is a nuclear hormone receptor transcription factor [9]. AR in the uterus is mainly located in the endometrium, mesenchyme and myometrium. A higher level of AR expression has been reported in stromal cells than in epithelial cells [10]. Furthermore, AR expression varies with the menstrual cycle [11]. The structure of AR protein consists of three domains: transcriptional regulatory domain (N-terminal domain, NTD), DNA-binding domain (DBD), and ligand-binding domain (C-terminal domain, LBD) [12]. DBD is the most conserved region, and its main function is to bind DNA. LBD is the region that binds to AR to form a dimer, which is critical for AR activation. Androgens activate downstream target gene pathways mainly through the classical AR-binding pathway (Figure 1). First, androgens bind to AR. Then, AR is separated from heat shock proteins (HSP) and transferred to the nucleus. AR homodimers bind to the androgen response element (ARE) on the promoter region of the target gene and recruit coactivators, which finally regulate the transcription of target genes [13]. In addition, AR can be rapidly activated in the absence of ligands, known as the nonclassical pathway (Figure 1) [14]. This ligand-independent pathway may be related to AR phosphorylation or AR-related signaling factors [14]. This is a fast and short activation mode with a response time of only a few seconds to a few minutes. Noncanonical pathways are not involved in the regulation of transcription and translation of target genes. It includes the activation of the PI3K/AKT/mTOR (Phosphatidylinositide 3-kinases/Serine/threonine kinases of the AGC family/Mammalian target of rapamycin) signaling pathway, the MAPK/ERK (Mitogen-activated protein kinase/extracellularregulated protein kinase) signaling pathway, and these pathways interact with each other [15,16].
3. Androgen Signaling in Endometriosis
Endometriosis is a benign estrogen-dependent disease in which the ectopic endometrium implants in a location other than the endometrium [17]. The prevalence of endometriosis is 10% in women of childbearing age [18]. Severe endometriosis can result in extensive pelvic adhesions, which lead to pain and infertility. It has been reported that T levels in endometriosis lesions are 5–19 times higher than the corresponding serum levels [19]. Consistent with this finding, an increased concentration of T, but not DHT, was found in endometriosis lesions, suggesting that T is the predominant androgen in endometriosis [18]. In addition, Carneiro et al. found 5α-reductase in the cytoplasm of glandular and stromal cells of the ectopic endometrium. This enzyme may mediate the conversion of T to DHT to enhance AR stimulation [20]. These findings suggest that active androgens may be formed in endometriosis tissue and that both local and systemic androgens may act on ectopic endometrial cells. In addition, a study identified AR as an endometriosis-associated transcription factor, and 373 AR target genes were differentially expressed in endometriosis compared to normal endometrium [21]. What is more, AR gene polymorphism may be related to the pathogenesis of endometriosis. The AR gene has a polymorphic cytosine, adenine, and guanine (CAG) microsatellite in exon 1 that encodes a variable-length glutamine repeat in the amino-terminal domain of the AR protein [22]. Hsieh et al. found that individuals with the M genotype (21 CAG repeats) had a higher risk of developing endometriosis compared with individuals without the M genotype [23]. This suggests that the CAG repeat polymorphisms of AR may serve as useful markers to predict endometriosis.
The clinical therapy strategy of endometriosis is to remove or reduce ectopic endometrium. Treatments of endometriosis include surgical treatment and drug medication. Drug therapy mainly includes nonsteroidal anti-inflammatory drugs, progestins, androgens, oral contraceptives, and gonadotropin-releasing hormone (GnRH) analogues [24]. Androgen is regarded as an effective treatment for women who cannot tolerate oral contraceptives or high doses of progesterone. Danazol is a derivative of 17α-ethinyltestosterone that increases the serum level of free T [25]. Atrophy of the endometrium has been reported as a result of hyper-androgen status [26]. In addition, danazol decreases estrogen production by interfering with the secretion of follicle-stimulating hormone (FSH) and luteinizing hormone (LH) [26]. In this way, danazol produces a high androgenic and low estrogenic environment, thereby reducing the size of the lesions in endometriosis and relieving pain [27,28]. In 18 endometriosis patients implanted with an intrauterine device (IUD) containing 300–400 mg danazol, there were significant improvements in symptoms like dysmenorrhoea and dyspareunia [29]. Adenomyosis is a type of endometriosis characterized by the presence of ectopic endometrium in the myometrium and hyperplasia of adjacent smooth muscle [30]. Igarashi was the first to use an IUD containing 175 mg danazol for adenomyosis treatment. This treatment reduced the uterine size significantly, and 66.6% of patients became pregnant as a result [31]. A prospective study of women with adenomyosis found that after six months of danazol-loaded IUD treatment, 81% of the participants had complete remission of dysmenorrhea, and 76% of the participants had improved menstrual bleeding [32]. In a mouse model using an IUD containing danazol, the number of adenomyosis nodules decreased as the dose increased [33]. In terms of mechanism, danazol affects the proliferation of adenomyosis cells by inhibiting DNA synthesis and inducing apoptosis. In adenomyotic glands and stromal cells treated with danazol, ER and bcl-2 protein expression is decreased, and necrosis of apoptotic cells is increased [34]. Vaginal administration [35] or cervical injection of danazol [36] may also improve symptoms of pain, vaginal bleeding, and dyspareunia. However, the use of danazol is limited due to side effects such as liver dysfunction and weight gain.
4. Androgen Signaling in Uterine Fibroids
Uterine fibroids are benign tumors formed by the proliferation of smooth muscle tissue in the uterus. Depending on their location, uterine fibroids can be classified as intramural, submucosal, or subserosal types [37]. Uterine fibroids have a variety of clinical manifestations, including excessive menstruation, abdominal pain, anemia, and infertility, which affect the life quality of patients. High levels of estrogen in the body are a contributing factor to the development of uterine fibroids. Estrogen promotes the proliferation of uterine smooth muscle tissue by acting on ER [38]. Nevertheless, in vitro studies have shown that androgens are involved in the development of uterine fibroids as well (Figure 2) [39,40]. Wong et al. found that both high levels of T and E2 were associated with an increased risk of developing uterine fibroids [41]. Aromatase is overexpressed in uterine fibroids compared with normal myometrium, suggesting an interaction between androgen and estrogen in uterine fibroids [42]. The conversion of T to E2 might be the mechanism by which T promotes the growth of uterine fibroids [43].
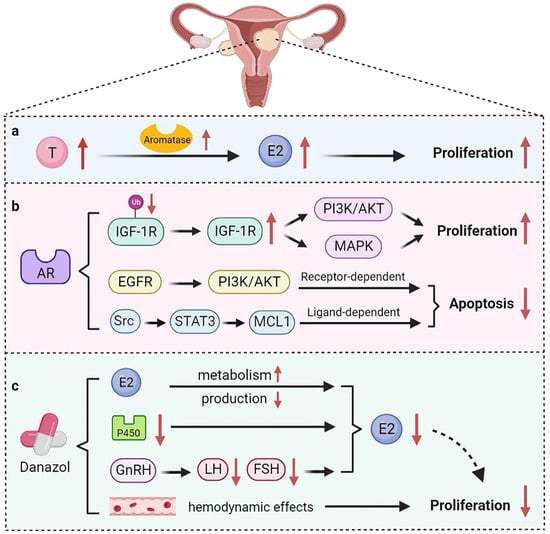
Figure 2.
The role of androgen in uterine fibroids. (a) Testosterone is converted into E2 under the action of aromatase, thereby promoting the growth of uterine fibroids; (b) AR maintains the stability of IGF-1R protein and triggers the PI3K/AKT and MAPK pathways to promote cell proliferation. AR also mediates anti-apoptotic functions through receptor-dependent and ligand-dependent pathways; (c) Danazol normalizes estrogen metabolism and affects the production of FSH and LH, ultimately inhibiting estrogen production. Danazol can also affect the hemodynamic effects of uterine fibroids.
AR was also found in uterine fibroids, suggesting a potential role for AR in the development of uterine fibroids [44]. Biopsies of 14 cases of fibroids and paired myometrial tissue revealed a 2–3-fold higher expression of AR and Ki-67 in fibroids than in normal myometrium [45]. AR exerts pro-proliferative and anti-apoptotic effects in myometrial cells through different signaling pathways (Figure 2). Insulin-like growth factor-1 (IGF-1) is an anabolic factor that regulates growth and differentiation. IGF-1 signaling has been reported to play a role in regulating proliferation of uterine leiomyomas [46]. AR inhibits the ubiquitination of the IGF-1 receptor (IGF-1R) protein, which, in turn, increases proliferation of myofibroblasts through the PI3K/AKT and MAPK pathways [47]. Furthermore, AR regulates the anti-apoptosis function of the downstream target gene MCL1 through receptor-dependent and ligand-dependent pathways [48]. In the receptor-dependent pathway, AR regulates the expression of MCL1 through the epidermal growth factor receptor (EGFR)/PI3K/AKT pathway. In the ligand-dependent pathway, AR triggers activation of Src kinase and the transcription factor STAT3, leading to increased expression of MCL1. These results suggest that AR is an important regulator of uterine fibroid growth. In addition, AR trinucleotide repeat polymorphisms are associated with susceptibility to uterine fibroids. Hsieh et al. has found that the distribution of CAG repeats of the AR gene differs between patients with uterine fibroids and normal individuals [49]. Another study demonstrated that Asian Taiwanese women with the S genotype (27 CAG repeats) have a higher risk of developing uterine fibroids [49]. However, women with 20 CAG repeats have more possibility of developing uterine fibroids in Asian Indian women [50]. Inversely, AR gene (CAG)n-repeat polymorphism was not associated with the risk of uterine fibroids in Brazilian women [51]. Therefore, AR gene polymorphism is inconsistent among different ethnic groups.
The treatments of uterine fibroids mainly include surgery and drug treatment. Myomectomy can radically remove the lesions. However, surgery also causes damage to the uterus and disrupts fertility [52]. Medicines commonly used include GnRH agonists (GnRH-a), danazol, progesterone, mifepristone, and some traditional Chinese medicines [53]. The mechanisms by which danazol inhibits the growth of fibroids include increasing the metabolism of estrogen and progesterone and inhibiting the production of both hormones (Figure 2). In patients with uterine fibroids, danazol normalizes estrogen metabolism by reducing the expression of aromatase cytochrome P450 [54]. In particular, danazol affects the production of FSH and LH by inhibiting the release of GnRH and ultimately inhibits the production of estrogen and progesterone [55]. A clinical study found that the volume of uterine fibroids in women decreased after danazol treatment, and the reduction persisted for six months or even longer after the end of the treatment [56]. In addition, danazol can prolong the therapeutic effect of GnRH-a on uterine fibroids. A cohort study that enrolled 21 patients treated with 100 mg danazol for six months after GnRH-a treatment found that the rebound of uterine fibroids was about 30% less than that of the control group [57]. During GnRH-a treatment, bone mineral content of patients decreased significantly, whereas this situation improved during danazol treatment. Additionally, danazol has been shown to affect the hemodynamic effects of uterine fibroids, with a positive correlation between therapeutic effects and increased uterine artery impedance [58].
5. Androgen Signaling in Endometrial Polyps
Endometrial polyps are one of the most common types of endometrial lesions [59]. Endometrial polyps are single or multiple smooth masses covered by epithelium due to the overgrowth of localized stromal cells. It is estimated that 8–12% of women suffered from endometrial polyps during their reproductive years [60]. The growth of endometrial polyps is stimulated by estrogen and can cause varying degrees of endometrial hyperplasia, which may progress to malignant diseases. Hysteroscopic resection is considered necessary for the treatment of endometrial polyps [61]. Several risk factors associated with endometrial polyps include obesity, late menopause, and the use of tamoxifen [62,63]. The effect of hormones on polyp formation is not well understood. In premenopausal women, decreased levels of estrogen receptors and progesterone receptors in polyp stromal cells may make polyps less sensitive to cyclic hormonal changes [64]. In one study, 13 postmenopausal women with benign endometrial lesions, including endometrial polyps, had significantly higher serum DHEA, DHEAS, and T levels than normal postmenopausal women [65]. It has been shown that endometrial polyps express the aromatase p450 enzyme, which stimulates the conversion of free T into E2, thereby inducing the proliferation of polyps [66]. This may be the reason why endometrial polyps are insensitive to T. Among 258 postmenopausal women who received estradiol and testosterone implants for the relief of menopausal symptoms, endometrial hyperplasia and endometrial polyps were found to be more prevalent [67]. Therefore, the combination of progestin and estrogen is superior to the combination of androgen and estrogen for postmenopausal women.
6. Androgen Signaling in Endometrial Hyperplasia
Endometrial hyperplasia is characterized by an increase in volume and structural changes in the endometrium, with a ratio of endometrial glands to mesenchyme greater than 1:1 [68]. EH results from chronic estrogen excess or progesterone deficiency [68]. There is strong evidence that EH increases the risk of concurrent endometrial cancer or endometrial cancer progression [69]. As defined by the World Health Organization (WHO) in 2014, EH includes hyperplasia with and without atypia [70]. Androgen plays an important role in the development of EH. The concentration of androgen in the peripheral blood of premenopausal women with EH was significantly higher than that of normal women [71]. Ito et al. found that the role of androgens in EH may be primarily regulated by serum T rather than DHT because of the lack of 5α-reductase [72]. Conservative medications for EH include progestins, GnRH-a, and danazol [73]. It is reported that danazol released from the danazol-releasing intrauterine device (D-IUD) has a direct therapeutic effect on EH through local diffusion and blood transmission (Figure 3) [73]. The mechanisms of action of danazol on EH include inhibition of endometrial cell proliferation and reduction of estrogen synthesis [74]. After treatment with danazol, pseudodecidual changes occurred. Endometrial glands atrophied, and further, they were transformed into normal secretory endometrium, thus improving the symptoms of patients with EH [73]. An animal experiment demonstrated that danazol significantly reduced the incidence of atypical EH by inhibiting estrogen-induced c-fos/jun expression [75].
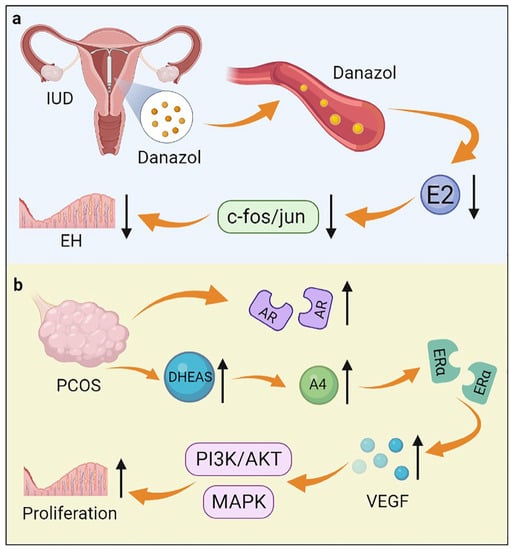
Figure 3.
The role of androgen in endometrial hyperplasia. (a) An IUD containing danazol inhibits estrogen-induced c-fos/jun expression by releasing danazol, thereby inhibiting endometrial hyperplasia; (b) DHEAS and AR are increased in women with PCOS. DHEAS is converted to A4, which increases the expression of VEGF through ERα. VEGF further activates the MAPK and PI3K/AKT pathways, thereby promoting the occurrence of endometrial hyperplasia.
Polycystic ovary syndrome (PCOS) is a common endocrine disorder that affects 5–8% of women of reproductive age. PCOS is characterized by polycystic ovaries, elevated androgen levels, and menstrual irregularities [76]. There is a high prevalence of endometrial lesions such as endometrial hyperplasia, endometrial cancer, and embryo implantation failure in women with PCOS [77]. Prenatal hyperandrogenization (PH) is considered to be one of the main factors in the development of PCOS. Ferreira et al. found that PH caused an increase in the total thickness of the uterus and disturbed the cell cycle, which led to the development of EH [78]. About 20–30% of PCOS patients have excess androgens in circulation, especially DHEAS [79]. In women with PCOS, DHEAS is converted to high levels of A4 by steroidogenic enzymes. Then, A4 acts through ERα to increase the expression of vascular endothelial growth factor (VEGF), which activates the MAPK and PI3K/AKT pathways in an autocrine/paracrine manner, thereby promoting the development of EH (Figure 3) [80]. In addition, AR expression increased in the endometrium during the development of EH in women with PCOS [81]. It is confirmed that androgens induce the development of EH in PCOS patients through AR-mediated AMPK (Adenosine 5’-monophosphate (AMP)-activated protein kinase) -α activation [82]. In addition, Li et al. reported that the combination therapy of Diane-35 and metformin could treat EH and early-stage endometrial cancer in women with PCOS [83]. The exact mechanism is not clear but may be caused by the anti-androgenic properties of Diane-35 and inhibition of endometrial AR signaling.
7. Androgen Signaling in Endometrial Cancer
7.1. Dual Effects of Androgens
Endometrial cancer is one of the most common gynecological malignancies in women and is mainly classified into type I and type II [84]. Nearly 80% of patients have type I EC, which is mainly endometrioid adenocarcinoma and has a better prognosis [85]. Type II EC consists of serous, clear cell, carcinosarcoma, and undifferentiated carcinoma, as well as other rarer types [86]. Several studies have reported an association between elevated serum androgen levels and the risk of EC. In postmenopausal patients with EC, elevated levels of DHEA, DHEAS, A4, and T were found in patients with EC compared to the healthy population [87]. Tanaka et al. found that patients with endometrioid adenocarcinoma had an 8-fold higher DHT concentration in the tissue/serum ratio than normal controls [88]. A limitation of these studies is that they only considered patients’ hormone levels over a single period. In recent years, two random Mendelian studies genetically analyzed the hormone levels of over 12,000 EC patients over their lifetimes, and they found that free T was associated with EC side effects [89,90]. In terms of mechanism, a high level of androgens is converted into estrogens by aromatase, resulting in increased levels of estrogen in tissues and promoting tumor cell growth (Figure 4) [91]. Aromatase is expressed not only in cancer cells, but also in stromal cells [92]. High expression of aromatase in stromal cells is associated with poor prognosis, suggesting an interaction between the tumor cells and stromal cells [93]. Qiu et al. found that AR bound to forkhead box A1 (FOXA1) and activated the Notch signaling pathway, thereby promoting EC cell proliferation [94]. Besides, androgens and AR have been shown to enhance cancer cell migration, epithelial–mesenchymal transition (EMT), and the number of EC stem cells [95]. Androgen can also affect the drug resistance of EC cells to cisplatin chemotherapy by increasing the expression of CD133 [95].
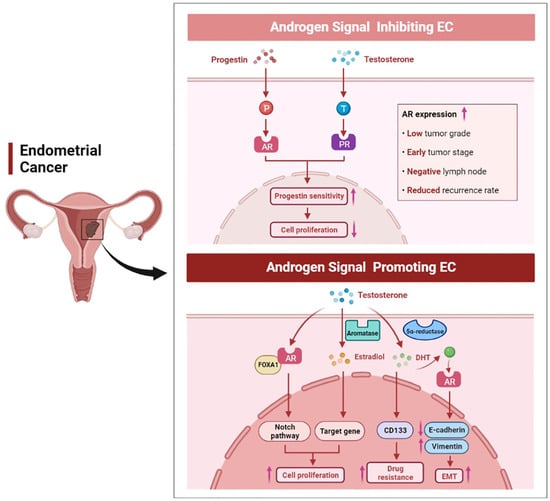
Figure 4.
The dual role of androgen signaling in endometrial cancer. High AR expression was positively correlated with a lower grade, early stage, negative lymph node, and lower recurrence rate in EC. Progestin can inhibit tumor growth through AR signaling, and androgen can also bind to PR, thereby enhancing the progestin sensitivity of EC. On the other hand, androgen can promote EC cell proliferation, enhance EMT, and affect the resistance of EC cells to cisplatin.
However, the effects of androgens on EC cells have inconsistent results compared to studies that directly measure the risk of androgens on EC. In vitro studies have shown that androgens, particularly A4, can inhibit the growth of endometrial epithelial cells [96]. It has been shown that the levels of androgens decrease with age in women, while the incidence of EC increases simultaneously [97]. Based on the action of aromatase, the decrease in androgen levels directly causes a corresponding decrease in the levels of estrogen in menopausal women. However, a decrease in estrogen levels does not reduce the incidence of EC. This may be because when androgen levels decrease, the inhibitory effect of androgen on EC cell proliferation is weakened [97]. This is consistent with the findings that androgen can inhibit the proliferation of Ishikawa cells, which are well-differentiated EC cells [98]. In addition, there is a correlation between an androgen signaling pathway and a progestin signaling pathway. It has been demonstrated that androgens increase PR expression in EC, thus exogenous androgen therapy may be an innovative therapy for EC patients who are insensitive to progestin treatment (Figure 4) [99]. What is more, progestin can reduce the stimulation of estrogen signaling and inhibit the proliferation of EC cells via upregulating the expression of AR [100,101]. It has been reported that MFE-296 endometrial cancer cells express AR in vitro, and both progestin and DHT treatment can inhibit the proliferation of MFE-296 cells [102]. In mammalian cells expressing exogenous or endogenous AR, medroxyprogesterone acetate (MPA) exerts a pronounced androgen agonistic effect. A significant increase in AR transcriptional activity was observed in the COS-1 cell line after MPA treatment in vitro [103]. In conclusion, due to hormone interaction, cell specificity, androgen type, androgen exposure time and other uncertain factors, it may be difficult to determine the role of androgen in EC [104].
7.2. AR Expression in EC
AR signaling in EC has both oncogenic and tumor suppressive effects, which may depend on the different stages of EC (Figure 4). In a mouse model of type I EC, a third-generation AR antagonist, enzalutamide inhibited EC cell proliferation and increased cancer cell apoptosis in a dose-dependent manner. However, enzalutamide increased tumor invasion and metastasis in advanced tumors [105]. A number of studies showed that AR expression was significantly increased in type I EC and was associated with favorable prognostic factors such as early stage of cancer, low grade, lymph node negativity, and reduced tumor recurrence rate [106,107,108]. Besides, decreased AR expression has been reported to lead to methylation of the MLH1 gene, which, in turn, causes mismatch repair (MMR) deficiency and promotes endometrial carcinogenesis [109]. Taken together, these data suggest that AR may play different roles in the development of EC. AR expression levels can also be used to predict the occurrence of EC. The N-terminal domain of the human AR gene contains CAG repeats that affect transcriptional efficiency, and the length of the CAG sequence is inversely related to AR activity [110]. It was shown that the CAG sequence was longer in patients with EC than in normal people, thus resulting in reduced AR activity. The decrease of AR activity reduced the anti-proliferative effect on endometrial cells and promoted endometrial cell carcinogenesis [110]. Thus, the susceptibility of an individual to EC can be predicted based on the length of the CAG repeat sequence in the AR gene. Furthermore, hypermethylation of CpG islands on AR genes leads to AR inactivation and is associated with the development of stage III and IV EC [111].
Previous studies have focused on endometrioid adenocarcinoma, with fewer studies on high-grade EC. Undifferentiated endometrial cancer (UC) and dedifferentiated endometrial cancer (DEAC) are high-grade endometrial cancers with a lower incidence and a greater capacity for invasion and metastasis [112]. AR is reported to be highly expressed in UC/DEAC (63%), serous carcinomas (88%), and carcinosarcomas (80%) [113]. In addition, AR is positive in these high-grade endometrial cancers lacking ER or PR expression [113]. These findings suggest a potential therapeutic role for androgen inhibitors in the treatment of patients with these tumors. In prostate cancer and AR-positive triple-negative breast cancer, androgen antagonists have shown good therapeutic effects [114,115]. AR immunostaining allows screening of a group of AR-positive EC that would benefit from anti-androgen therapy. Therefore, androgen antagonists are novel therapeutic agents for high-grade non-endometrioid cancers that currently lack effective endocrine therapy [116].
8. Androgen Signaling in Infertility Associated with Endometrial Dysfunction
Androgens have a beneficial effect on the regulation of pregnancy. It is reported that androgens regulate endometrial decidualization and embryo implantation [117]. Endometrial decidualization is the proliferation and differentiation of endometrial mesenchymal cells, which occurs during pregnancy [118]. Blocking androgen signaling leads to a significant reduction in the expression of key indicators of decidualization such as PRL and IGFBP1 [119]. In human embryonic stem cells, the expression and activity of the androgen synthesis-related enzymes AKR1C3 and SRD5A1 are associated with decidualization in vitro, suggesting that these enzymes may be targets for the treatment of infertility associated with endometrial dysfunction [119]. Androgens also regulate the expression of endometrial receptivity-related markers in human embryonic stem cells [119]. Gibson et al. reported that supplementation with DHEA could improve the decidualization of the endometrium and promote the initiation of pregnancy [120]. Interestingly, treatment with supraphysiological levels of DHEA is not effective and decidualization is weakened [121]. Therefore, the therapeutic effect of DHEA may be dose-dependent, and it is suggested to use a wider therapeutic window for treatment. Additionally, in mouse models, androgen deficiency delays embryo implantation, while excess androgens result in abnormal gene expression at the site of implantation [122].
Low androgen levels have been reported to be associated with decreased ovarian reserve (DOR) and lower pregnancy rates [123]. Supplementation with androgens or androgen precursors may improve ovarian function, thereby increasing embryo fertilization rates. In 89 patients with DOR treated with DHEA before in vitro fertilization (IVF), clinical pregnancy rates were significantly increased [124]. However, in a study of DHEA supplementation in women with DOR undergoing IVF or intracytoplasmic single sperm injection (ICSI), a higher pregnancy rate was found in the control group than in the DHEA-treated women [125]. Although DHEA supplementation in these trials was intended to support ovarian function, DHEA did not appear to improve ovarian function in terms of ovulation rates or the number of embryos fertilized [124]. Moreover, the group supplemented with DHEA had a lower miscarriage rate, and almost half of the pregnancies occurred before IVF [124]. It is more likely that the ability of DHEA to increase pregnancy rates is due to the improved condition of endometrium. Therefore, androgens can be used in assisted reproduction techniques to improve pregnancy outcomes of patients, but more research is needed to investigate the effects of androgens on ovary andendometrium. However, many women are intolerant to the side effects produced by androgens. The selective androgen receptor modulator (SARM) and new enzyme selective modulator such as AKR1C3 provide a more targeted treatment to reduce the side effects [117]. Targeted androgen therapy can also be performed through intrauterine devices to provide an effective treatment for improving pregnancy outcomes.
9. Conclusions
Androgen signaling is involved in many uterine physiological processes, and there are varying degrees of evidence for its role in benign uterine lesions and malignant progression. Androgens have been shown to be effective in many uterine diseases; however, the side effects of androgens have limited their use in women’s diseases. SARM is a new class of compounds that exhibit agonist and antagonist effects on AR in target tissues with good efficacy and few side effects [126]. SARMs have been used to treat breast cancer, but there have been no clinical trials for endometrial cancer, endometriosis, and other uterine-related diseases, so further research is needed [127]. A growing number of epidemiological and clinical trials support the further exploration of AR-targeted drugs in a variety of uterine-related diseases (Table 1). The next generation of AR-targeted drugs will play a broad therapeutic role in uterine-related diseases in the coming decades.

Table 1.
Ongoing studies testing androgen-related therapies in uterine diseases.
Author Contributions
Conceptualization, Z.Z. and H.L.; writing—original draft preparation, M.L., J.Y. and Y.H.; writing—review and editing, J.M. and J.X.; visualization, Y.W. and L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (grants 81872111 and 81672562), the National Key Technology R&D Program of China (2019YFC1005200 and 2019YFC1005201), the Shanghai Municipal Science and Technology Committee of Shanghai outstanding academic leaders plan (19XD1423100), the project of Outstanding Medical Doctor for ZZ, the Shanghai Municipal Education Commission—Gaofeng Clinical Medicine Grant Support (20181713, 20181714), the Shanghai Talent Development Foundation (2019121), the Shanghai Municipal Science and Technology Committee of Multi-center Clinical Study Project (20Z21900400), the Shanghai Collaborative Innovation Center of Translational Medicine (TM202015), the Health care project of Shanghai Pudong New Area Science and Technology Development Fund for Livelihood Research (PKJ2021-Y05), and the scientific research project of the Shanghai Municipal Health Commission (202140279).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Allen, N.E.; Key, T.J.; Dossus, L.; Rinaldi, S.; Cust, A.; Lukanova, A.; Peeters, P.H.; Onland-Moret, N.C.; Lahmann, P.H.; Berrino, F.; et al. Endogenous sex hormones and endometrial cancer risk in women in the European Prospective Investigation into Cancer and Nutrition (EPIC). Endocr. -Relat. Cancer 2008, 15, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Burger, H.G. Androgen production in women. Fertil. Steril. 2002, 4, S3–S5. [Google Scholar] [CrossRef]
- Simitsidellis, I.; Saunders, P.T.K.; Gibson, D.A. Androgens and endometrium: New insights and new targets. Mol. Cell. Endocrinol. 2018, 465, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Labrie, F.; Martel, C.; Bélanger, A.; Pelletier, G. Androgens in women are essentially made from DHEA in each peripheral tissue according to intracrinology. J. Steroid Biochem. Mol. Biol. 2017, 168, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Labrie, F.; Simard, J.; Luu-The, V.; Bélanger, A.; Pelletier, G. Structure, function and tissue-specific gene expression of 3β-hydroxysteroid dehydrogenase/5-ene-4-ene isomerase enzymes in classical and peripheral intracrine steroidogenic tissues. J. Steroid Biochem. Mol. Biol. 1992, 43, 805–826. [Google Scholar] [CrossRef]
- Nakamura, Y.; Hornsby, P.J.; Casson, P.; Morimoto, R.; Satoh, F.; Xing, Y.; Kennedy, M.R.; Sasano, H.; Rainey, W.E. Type 5 17beta-hydroxysteroid dehydrogenase (AKR1C3) contributes to testosterone production in the adrenal reticularis. J. Clin. Endocrinol. Metab. 2009, 94, 2192–2198. [Google Scholar] [CrossRef]
- Sinnesael, M.; Claessens, F.; Boonen, S.; Vanderschueren, D. Novel insights in the regulation and mechanism of androgen action on bone. Curr. Opin. Endocrinol. Diabetes Obes. 2013, 20, 240–244. [Google Scholar] [CrossRef]
- Bulun, S.E.; Lin, Z.; Imir, G.; Amin, S.; Demura, M.; Yilmaz, B.; Martin, R.; Utsunomiya, H.; Thung, S.; Gurates, B.; et al. Regulation of aromatase expression in estrogen-responsive breast and uterine disease: From bench to treatment. Pharmacol. Rev. 2005, 57, 359–383. [Google Scholar] [CrossRef]
- Schweizer, M.T.; Yu, E.Y. Persistent androgen receptor addiction in castration-resistant prostate cancer. J. Hematol. Oncol. 2015, 8, 128. [Google Scholar] [CrossRef]
- Apparao, K.B.; Lovely, L.P.; Gui, Y.; Lininger, R.A.; Lessey, B.A. Elevated endometrial androgen receptor expression in women with polycystic ovarian syndrome. Biol. Reprod. 2002, 66, 297–304. [Google Scholar] [CrossRef]
- Mertens, H.J.; Heineman, M.J.; Theunissen, P.H.; de Jong, F.H.; Evers, J.L. Androgen, estrogen and progesterone receptor expression in the human uterus during the menstrual cycle. Eur. J. Obstet. Gynecol. Reprod. Biol. 2001, 98, 58–65. [Google Scholar] [CrossRef]
- Lallous, N.; Dalal, K.; Cherkasov, A.; Rennie, P.S. Targeting alternative sites on the androgen receptor to treat castration-resistant prostate cancer. Int. J. Mol. Sci. 2013, 14, 12496–12519. [Google Scholar] [CrossRef] [PubMed]
- Koryakina, Y.; Ta, H.Q.; Gioeli, D. Androgen receptor phosphorylation: Biological context and functional consequences. Endocr. -Relat. Cancer 2014, 21, T131–T145. [Google Scholar] [CrossRef] [PubMed]
- Zamagni, A.; Cortesi, M.; Zanoni, M.; Tesei, A. Non-nuclear AR Signaling in Prostate Cancer. Front. Chem. 2019, 7, 651. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Mawji, N.R.; Bruchovsky, N.; Sadar, M.D. Ligand-independent activation of the androgen receptor by interleukin-6 and the role of steroid receptor coactivator-1 in prostate cancer cells. J. Biol. Chem. 2002, 277, 38087–38094. [Google Scholar] [CrossRef]
- Bonaccorsi, L.; Marchiani, S.; Ferruzzi, P.; Muratori, M.; Crescioli, C.; Forti, G.; Maggi, M.; Baldi, E. Non-genomic effects of the androgen receptor and vitamin D agonist are involved in suppressing invasive phenotype of prostate cancer cells. Steroids 2006, 71, 304–309. [Google Scholar] [CrossRef]
- Giudice, L.C.; Kao, L.C. Endometriosis. Lancet 2004, 364, 1789–1799. [Google Scholar] [CrossRef]
- Giudice, L.C. Clinical practice. Endometriosis. N. Engl. J. Med. 2010, 362, 2389–2398. [Google Scholar] [CrossRef]
- Huhtinen, K.; Saloniemi-Heinonen, T.; Keski-Rahkonen, P.; Desai, R.; Laajala, D.; Ståhle, M.; Häkkinen, M.R.; Awosanya, M.; Suvitie, P.; Kujari, H.; et al. Intra-tissue steroid profiling indicates differential progesterone and testosterone metabolism in the endometrium and endometriosis lesions. J. Clin. Endocrinol. Metab. 2014, 99, E2188–E2197. [Google Scholar] [CrossRef]
- Carneiro, M.M.; Morsch, D.M.; Camargos, A.F.; Reis, F.M.; Spritzer, P.M. Androgen receptor and 5alpha-reductase are expressed in pelvic endometriosis. BJOG 2008, 115, 113–117. [Google Scholar] [CrossRef]
- Yang, H.; Kang, K.; Cheng, C.; Mamillapalli, R.; Taylor, H.S. Integrative Analysis Reveals Regulatory Programs in Endometriosis. Reprod. Sci. 2015, 22, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Mitsumori, K.; Terai, A.; Oka, H.; Segawa, T.; Ogura, K.; Yoshida, O.; Ogawa, O. Androgen receptor CAG repeat length polymorphism in benign prostatic hyperplasia (BPH): Correlation with adenoma growth. Prostate 1999, 41, 253–257. [Google Scholar] [CrossRef]
- Hsieh, Y.Y.; Chang, C.C.; Tsai, F.J.; Wu, J.Y.; Tsai, C.H.; Tsai, H.D. Androgen receptor trinucleotide polymorphism in endometriosis. Fertil. Steril. 2001, 76, 412–413. [Google Scholar] [CrossRef]
- Vannuccini, S.; Luisi, S.; Tosti, C.; Sorbi, F.; Petraglia, F. Role of medical therapy in the management of uterine adenomyosis. Fertil. Steril. 2018, 109, 398–405. [Google Scholar] [CrossRef]
- Barbieri, R.L. Danazol: Molecular, endocrine, and clinical pharmacology. Prog. Clin. Biol. Res. 1990, 323, 241–252. [Google Scholar]
- Barbieri, R.L. Endometriosis 1990. Current treatment approaches. Drugs 1990, 39, 502–510. [Google Scholar] [CrossRef]
- Selak, V.; Farquhar, C.; Prentice, A.; Singla, A. Danazol for pelvic pain associated with endometriosis. Cochrane Database Syst. Rev. 2007, 4, Cd000068. [Google Scholar]
- Ferrero, S.; Tramalloni, D.; Venturini, P.L.; Remorgida, V. Vaginal danazol for women with rectovaginal endometriosis and pain symptoms persisting after insertion of a levonorgestrel-releasing intrauterine device. Int. J. Gynaecol. Obstet. Off. Organ Int. Fed. Gynaecol. Obstet. 2011, 113, 116–119. [Google Scholar] [CrossRef]
- Cobellis, L.; Razzi, S.; Fava, A.; Severi, F.M.; Igarashi, M.; Petraglia, F. A danazol-loaded intrauterine device decreases dysmenorrhea, pelvic pain, and dyspareunia associated with endometriosis. Fertil. Steril. 2004, 82, 239–240. [Google Scholar] [CrossRef]
- Farquhar, C.; Brosens, I. Medical and surgical management of adenomyosis. Best Pract. Res. Clin. Obstet. Gynaecol. 2006, 20, 603–616. [Google Scholar] [CrossRef]
- Igarashi, M. A new therapy for pelvic endometriosis and uterine adenomyosis: Local effect of vaginal and intrauterine danazol application. Asia-Ocean. J. Obstet. Gynaecol. 1990, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shawki, O.A. Danazol loaded intrauterine device (D-IUD): A novel conservative management for uterine adenomyosis. Middle East Fertil. J. 2002, 7, 214–220. [Google Scholar]
- Zhang, X.; Yuan, H.; Deng, L.; Hu, F.; Ma, J.; Lin, J. Evaluation of the efficacy of a danazol-loaded intrauterine contraceptive device on adenomyosis in an ICR mouse model. Hum. Reprod. 2008, 23, 2024–2030. [Google Scholar] [CrossRef] [PubMed]
- Ueki, K.; Kumagai, K.; Yamashita, H.; Li, Z.L.; Ueki, M.; Otsuki, Y. Expression of apoptosis-related proteins in adenomyotic uteri treated with danazol and GnRH agonists. Int. J. Gynaecol. Obstet. Off. Organ Int. Fed. Gynaecol. Obstet. 2004, 23, 248–258. [Google Scholar] [CrossRef]
- Luisi, S.; Razzi, S.; Lazzeri, L.; Bocchi, C.; Severi, F.M.; Petraglia, F. Efficacy of vaginal danazol treatment in women with menorrhagia during fertile age. Fertil. Steril. 2009, 92, 1351–1354. [Google Scholar] [CrossRef] [PubMed]
- Takebayashi, T.; Fujino, Y.; Umesaki, N.; Ogita, S. Danazol suspension injected into the uterine cervix of patients with adenomyosis and myoma. Prelim. Study Gynecol. Obstet. Investig. 1995, 39, 207–211. [Google Scholar] [CrossRef]
- Friend, D.R. Drug delivery for the treatment of endometriosis and uterine fibroids. Drug Deliv. Transl. Res. 2017, 7, 829–839. [Google Scholar] [CrossRef]
- Moravek, M.B.; Yin, P.; Ono, M.; Coon, J.S.t.; Dyson, M.T.; Navarro, A.; Marsh, E.E.; Chakravarti, D.; Kim, J.J.; Wei, J.J.; et al. Ovarian steroids, stem cells and uterine leiomyoma: Therapeutic implications. Hum. Reprod. Update 2015, 21, 1–12. [Google Scholar] [CrossRef]
- Taylor, D.K.; Leppert, P.C. Treatment for Uterine Fibroids: Searching for Effective Drug Therapies. Drug Discov. Today Ther. Strateg. 2012, 9, e41–e49. [Google Scholar] [CrossRef]
- Ke, L.Q.; Yang, K.; Li, J.; Li, C.M. Danazol for uterine fibroids. Cochrane Database Syst. Rev. 2009, 2009, Cd007692. [Google Scholar]
- Wong, J.Y.; Gold, E.B.; Johnson, W.O.; Lee, J.S. Circulating Sex Hormones and Risk of Uterine Fibroids: Study of Women’s Health Across the Nation (SWAN). J. Clin. Endocrinol. Metab. 2016, 101, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E.; Imir, G.; Utsunomiya, H.; Thung, S.; Gurates, B.; Tamura, M.; Lin, Z. Aromatase in endometriosis and uterine leiomyomata. J. Steroid Biochem. Mol. Biol. 2005, 95, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Moroni, R.; Vieira, C.; Ferriani, R.; Candido-Dos-Reis, F.; Brito, L. Pharmacological treatment of uterine fibroids. Ann. Med. Health Sci. Res. 2014, 4, S185–S192. [Google Scholar] [CrossRef] [PubMed]
- Horie, K.; Takakura, K.; Imai, K.; Liao, S.; Mori, T. Immunohistochemical localization of androgen receptor in the human endometrium, decidua, placenta and pathological conditions of the endometrium. Hum. Reprod. 1992, 7, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Lan, M.; Li, H.; Bao, L.; Li, M.; Lye, S.; Dong, X. In Vivo Evidence of the Androgen Receptor in Association With Myometrial Cell Proliferation and Apoptosis. Reprod. Sci. 2016, 23, 264–271. [Google Scholar] [CrossRef]
- Gao, Z.; Matsuo, H.; Wang, Y.; Nakago, S.; Maruo, T. Up-regulation by IGF-I of proliferating cell nuclear antigen and Bcl-2 protein expression in human uterine leiomyoma cells. J. Clin. Endocrinol. Metab. 2001, 86, 5593–5599. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Y.; Xie, N.; Shynlova, O.; Challis, J.R.; Slater, D.; Lye, S.; Dong, X. Proliferative action of the androgen receptor in human uterine myometrial cells--a key regulator for myometrium phenotype programming. J. Clin. Endocrinol. Metab. 2013, 98, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, Y.; Morin, D.; Plymate, S.; Lye, S.; Dong, X. The androgen receptor mediates antiapoptotic function in myometrial cells. Cell Death Dis. 2014, 5, e1338. [Google Scholar] [CrossRef]
- Hsieh, Y.Y.; Chang, C.C.; Tsai, F.J.; Lin, C.C.; Yeh, L.S.; Peng, C.T. Androgen receptor trinucleotide polymorphism in leiomyoma. J. Assist. Reprod. Genet. 2004, 21, 453–457. [Google Scholar] [CrossRef][Green Version]
- Shaik, N.A.; Govindan, S.; Kodati, V.; Rao, K.P.; Hasan, Q. Polymorphic (CAG)n repeats in the androgen receptor gene: A risk marker for endometriosis and uterine leiomyomas. Hematol. Oncol. Stem Cell Ther. 2009, 2, 289–293. [Google Scholar] [CrossRef]
- Rosa, F.E.; Rde, A.C.; Ambrosio, E.P.; Cirilo, P.D.R.; Pontes, A.; Rainho, C.A.; Rogatto, S.R. Polymorphisms of CYP17A1, CYP19, and androgen in Brazilian women with uterine leiomyomas. Clin. Chem. Lab. Med. 2008, 46, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Munro, M.G. Uterine polyps, adenomyosis, leiomyomas, and endometrial receptivity. Fertil. Steril. 2019, 111, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.K.; Kor, C.T.; Chen, C.P.; Chen, H.T.; Yang, P.T.; Tsai, C.D.; Huang, C.H. Increased Risk of Venous Thromboembolism in Women with Uterine Leiomyoma: A Nationwide, Population-Based Case-Control Study. Acta Cardiol. Sin. 2018, 34, 66–76. [Google Scholar] [PubMed]
- Ishihara, H.; Kitawaki, J.; Kado, N.; Koshiba, H.; Fushiki, S.; Honjo, H. Gonadotropin-releasing hormone agonist and danazol normalize aromatase cytochrome P450 expression in eutopic endometrium from women with endometriosis, adenomyosis, or leiomyomas. Fertil. Steril. 2003, 79 (Suppl. S1), 735–742. [Google Scholar] [CrossRef]
- Xuan, J.; Deng, G.; Liu, R.; Chen, X.; Zheng, Y. Analysis of medication data of women with uterine fibroids based on data mining technology. J. Infect. Public Health 2020, 13, 1513–1516. [Google Scholar] [CrossRef]
- De Leo, V.; la Marca, A.; Morgante, G. Short-term treatment of uterine fibromyomas with danazol. Gynecol. Obstet. Investig. 1999, 47, 258–262. [Google Scholar] [CrossRef]
- De Leo, V.; Morgante, G.; Lanzetta, D.; D’Antona, D.; Bertieri, R.S. Danazol administration after gonadotrophin-releasing hormone analogue reduces rebound of uterine myomas. Hum. Reprod. 1997, 12, 357–360. [Google Scholar] [CrossRef][Green Version]
- la Marca, A.; Musacchio, M.C.; Morgante, G.; Petraglia, F.; de Leo, V. Hemodynamic effect of danazol therapy in women with uterine leiomyomata. Fertil. Steril. 2003, 79, 1240–1242. [Google Scholar] [CrossRef]
- Lethaby, A.; Suckling, J.; Barlow, D.; Farquhar, C.M.; Jepson, R.G.; Roberts, H. Hormone replacement therapy in postmenopausal women: Endometrial hyperplasia and irregular bleeding. Cochrane Database Syst. Rev. 2004, Cd000402. [Google Scholar] [CrossRef]
- Dreisler, E.; Sorensen, S.S.; Ibsen, P.H.; Lose, G. Prevalence of endometrial polyps and abnormal uterine bleeding in a Danish population aged 20–74 years. Ultrasound Obstet. Gynecol. 2009, 33, 102–108. [Google Scholar] [CrossRef]
- Maia, H., Jr.; Maltez, A.; Fahel, P.; Coutinho, E. Hysteroscopic and immunohistochemical findings in type I and type II endometrial carcinomas. J. Am. Assoc. Gynecol. Laparosc. 2001, 8, 222–230. [Google Scholar] [CrossRef]
- Cohen, I. Endometrial pathologies associated with postmenopausal tamoxifen treatment. Gynecol. Oncol. 2004, 94, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Chalas, E.; Costantino, J.P.; Wickerham, D.L.; Wolmark, N.; Lewis, G.C.; Bergman, C.; Runowicz, C.D. Benign gynecologic conditions among participants in the Breast Cancer Prevention Trial. Am. J. Obstet. Gynecol. 2005, 192, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.J.; Jackson, T.L.; Reid, J.G.; Duffy, S.R. The differential expression of oestrogen receptors, progesterone receptors, Bcl-2 and Ki67 in endometrial polyps. BJOG 2003, 110, 794–798. [Google Scholar] [CrossRef] [PubMed]
- Brody, S.; Carlström, K.; von Uexküll, A.K.; Lagrelius, A.; Lunell, N.O.; Rosenborg, L. Peripheral hormone levels and the endometrial condition in postmenopausal women. Acta Obstet. Gynecol. Scand. 1983, 62, 525–529. [Google Scholar] [CrossRef]
- Maia, H., Jr.; Pimentel, K.; Silva, T.M.; Freitas, L.A.; Zausner, B.; Athayde, C.; Coutinho, E.M. Aromatase and cyclooxygenase-2 expression in endometrial polyps during the menstrual cycle. Gynecol. Endocrinol. 2006, 22, 219–224. [Google Scholar] [CrossRef]
- Filho, A.M.; Barbosa, I.C.; Maia, H., Jr.; Genes, C.C.; Coutinho, E.M. Effects of subdermal implants of estradiol and testosterone on the endometrium of postmenopausal women. Gynecol. Endocrinol. 2007, 23, 511–517. [Google Scholar] [CrossRef]
- Horn, L.C.; Schnurrbusch, U.; Bilek, K.; Hentschel, B.; Einenkel, J. Risk of progression in complex and atypical endometrial hyperplasia: Clinicopathologic analysis in cases with and without progestogen treatment. Int. J. Gynecol. Cancer 2004, 14, 348–353. [Google Scholar] [CrossRef]
- Nees, L.K.; Heublein, S.; Steinmacher, S.; Juhasz-Böss, I.; Brucker, S.; Tempfer, C.B.; Wallwiener, M. Endometrial hyperplasia as a risk factor of endometrial cancer. Arch. Gynecol. Obstet. 2022, 306, 407–421. [Google Scholar] [CrossRef]
- Zaino, R.; Carinelli, S.; Eng, C.; Kurman, R.; Carcangiu, M.; Herrington, C.; Young, R. Tumours of the uterine corpus. In WHO Classification of Tumours of Female Reproductive Organs; World Health Organization Classification of Tumours; IARC Press: Lyon, France, 2014. [Google Scholar]
- Vitoratos, N.; Gregoriou, O.; Hassiakos, D.; Zourlas, P.A. The role of androgens in the late-premenopausal woman with adenomatous hyperplasia of the endometrium. Int. J. Gynaecol. Obstet. 1991, 34, 157–161. [Google Scholar] [CrossRef]
- Ito, K.; Suzuki, T.; Akahira, J.; Moriya, T.; Kaneko, C.; Utsunomiya, H.; Yaegashi, N.; Okamura, K.; Sasano, H. Expression of androgen receptor and 5alpha-reductases in the human normal endometrium and its disorders. Int. J. Cancer 2002, 99, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Tamaoka, Y.; Orikasa, H.; Sumi, Y.; Sakakura, K.; Kamei, K.; Nagatani, M.; Ezawa, S. Treatment of endometrial hyperplasia with a danazol-releasing intrauterine device: A prospective study. Gynecol. Obstet. Investig. 2004, 58, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Rigano, A.; Sturlese, E.; Rigano, M.; Baviera, G. Endocrine changes in postmenopausal women after high-dose danazol therapy. Panminerva Med. 1999, 41, 139–142. [Google Scholar] [PubMed]
- Niwa, K.; Hashimoto, M.; Morishita, S.; Yokoyama, Y.; Lian, Z.; Tagami, K.; Mori, H.; Tamaya, T. Preventive effects of danazol on endometrial carcinogenesis in mice. Cancer Lett. 2000, 158, 133–139. [Google Scholar] [CrossRef]
- Azziz, R.; Woods, K.S.; Reyna, R.; Key, T.J.; Knochenhauer, E.S.; Yildiz, B.O. The prevalence and features of the polycystic ovary syndrome in an unselected population. J. Clin. Endocrinol. Metab. 2004, 89, 2745–2749. [Google Scholar] [CrossRef]
- Pillay, O.C.; Fong, L.F.T.; Crow, J.C.; Benjamin, E.; Mould, T.; Atiomo, W.; Menon, P.A.; Leonard, A.J.; Hardiman, P. The association between polycystic ovaries and endometrial cancer. Hum. Reprod. 2006, 21, 924–929. [Google Scholar] [CrossRef]
- Ferreira, S.R.; Goyeneche, A.A.; Heber, M.F.; Abruzzese, G.A.; Telleria, C.M.; Motta, A.B. Prenatally androgenized female rats develop uterine hyperplasia when adult. Mol. Cell. Endocrinol. 2020, 499, 110610. [Google Scholar] [CrossRef]
- Pinola, P.; Piltonen, T.T.; Puurunen, J.; Vanky, E.; Sundström-Poromaa, I.; Stener-Victorin, E.; Ruokonen, A.; Puukka, K.; Tapanainen, J.S.; Morin-Papunen, L.C. Androgen Profile Through Life in Women With Polycystic Ovary Syndrome: A Nordic Multicenter Collaboration Study. J. Clin. Endocrinol. Metab. 2015, 100, 3400–3407. [Google Scholar] [CrossRef]
- Plaza-Parrochia, F.; Romero, C.; Valladares, L.; Vega, M. Endometrium and steroids, a pathologic overview. Steroids 2017, 126, 85–91. [Google Scholar] [CrossRef]
- Shang, K.; Jia, X.; Qiao, J.; Kang, J.; Guan, Y. Endometrial abnormality in women with polycystic ovary syndrome. Reprod. Sci. 2012, 19, 674–683. [Google Scholar] [CrossRef]
- Li, X.; Pishdari, B.; Cui, P.; Hu, M.; Yang, H.P.; Guo, Y.R.; Jiang, H.Y.; Feng, Y.; Billig, H.; Shao, R. Regulation of Androgen Receptor Expression Alters AMPK Phosphorylation in the Endometrium: In Vivo and In Vitro Studies in Women with Polycystic Ovary Syndrome. Int. J. Biol. Sci. 2015, 11, 1376–1389. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Guo, Y.R.; Lin, J.F.; Feng, Y.; Billig, H.; Shao, R. Combination of Diane-35 and Metformin to Treat Early Endometrial Carcinoma in PCOS Women with Insulin Resistance. J. Cancer 2014, 5, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Bokhman, J.V. Two pathogenetic types of endometrial carcinoma. Gynecol. Oncol. 1983, 15, 10–17. [Google Scholar] [CrossRef]
- Wei, J.J.; Paintal, A.; Keh, P. Histologic and immunohistochemical analyses of endometrial carcinomas: Experiences from endometrial biopsies in 358 consultation cases. Arch. Pathol. Lab. Med. 2013, 137, 1574–1583. [Google Scholar] [CrossRef] [PubMed]
- Cree, I.A.; White, V.A.; Indave, B.I.; Lokuhetty, D. Revising the WHO classification: Female genital tract tumours. Histopathology 2020, 76, 151–156. [Google Scholar] [CrossRef]
- Audet-Walsh, E.; Lépine, J.; Grégoire, J.; Plante, M.; Caron, P.; Têtu, B.; Ayotte, P.; Brisson, J.; Villeneuve, L.; Bélanger, A.; et al. Profiling of endogenous estrogens, their precursors, and metabolites in endometrial cancer patients: Association with risk and relationship to clinical characteristics. J. Clin. Endocrinol. Metab. 2011, 96, E330–E339. [Google Scholar] [CrossRef]
- Tanaka, S.; Miki, Y.; Hashimoto, C.; Takagi, K.; Doe, Z.; Li, B.; Yaegashi, N.; Suzuki, T.; Ito, K. The role of 5α-reductase type 1 associated with intratumoral dihydrotestosterone concentrations in human endometrial carcinoma. Mol. Cell. Endocrinol. 2015, 401, 56–64. [Google Scholar] [CrossRef]
- Ruth, K.S.; Day, F.R.; Tyrrell, J.; Thompson, D.J.; Wood, A.R.; Mahajan, A.; Beaumont, R.N.; Wittemans, L.; Martin, S.; Busch, A.S. Using human genetics to understand the disease impacts of testosterone in men and women. Nat. Med. 2020, 26, 252–258. [Google Scholar] [CrossRef]
- Mullee, A.; Dimou, N.; Allen, N.; O’Mara, T.; Gunter, M.J.; Murphy, N. Testosterone, sex hormone-binding globulin, insulin-like growth factor-1 and endometrial cancer risk: Observational and Mendelian randomization analyses. Br. J. Cancer 2021, 125, 1308–1317. [Google Scholar] [CrossRef]
- Siiteri, P.K. Adipose tissue as a source of hormones. Am. J. Clin. Nutr. 1987, 45, 277–282. [Google Scholar] [CrossRef]
- Che, Q.; Liu, B.Y.; Liao, Y.; Zhang, H.J.; Yang, T.T.; He, Y.Y.; Xia, Y.H.; Lu, W.; He, X.Y.; Chen, Z.; et al. Activation of a positive feedback loop involving IL-6 and aromatase promotes intratumoral 17β-estradiol biosynthesis in endometrial carcinoma microenvironment. Int. J. Cancer 2014, 135, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Segawa, T.; Shozu, M.; Murakami, K.; Kasai, T.; Shinohara, K.; Nomura, K.; Ohno, S.; Inoue, M. Aromatase expression in stromal cells of endometrioid endometrial cancer correlates with poor survival. Clin. Cancer Res. 2005, 11, 2188–2194. [Google Scholar] [CrossRef] [PubMed]
- Qiu, M.; Bao, W.; Wang, J.; Yang, T.; He, X.; Liao, Y.; Wan, X. FOXA1 promotes tumor cell proliferation through AR involving the Notch pathway in endometrial cancer. BMC Cancer 2014, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chang, W.C.; Hung, Y.C.; Chang, Y.Y.; Bao, B.Y.; Huang, H.C.; Chung, W.M.; Shyr, C.R.; Ma, W.L. Androgen receptor increases CD133 expression and progenitor-like population that associate with cisplatin resistance in endometrial cancer cell line. Reprod. Sci. 2014, 21, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Tuckerman, E.M.; Okon, M.A.; Li, T.; Laird, S.M. Do androgens have a direct effect on endometrial function? An in vitro study. Fertil. Steril. 2000, 74, 771–779. [Google Scholar] [CrossRef]
- Jamison, P.M.; Altekruse, S.F.; Chang, J.T.; Zahn, J.; Lee, R.; Noone, A.M.; Barroilhet, L. Site-specific factors for cancer of the corpus uteri from SEER registries: Collaborative stage data collection system, version 1 and version 2. Cancer 2014, 120 (Suppl. 23), 3836–3845. [Google Scholar] [CrossRef]
- Lovely, L.P.; Rao, K.B.A.; Gui, Y.; Lessey, B.A. Characterization of androgen receptors in a well-differentiated endometrial adenocarcinoma cell line (Ishikawa). J. Steroid Biochem. Mol. Biol. 2000, 74, 235–241. [Google Scholar] [CrossRef]
- Park, C.; Babayev, S.; Carr, B.R.; Keller, P.W.; Word, R.A.; Bukulmez, O. Androgen regulation of progesterone receptor (PR) expression in endometrium: Implications for endometriosis. Fertil. Steril. 2014, 102, e79–e80. [Google Scholar] [CrossRef]
- Brenner, R.M.; Slayden, O.D. Progesterone receptor antagonists and the endometrial antiproliferative effect. Semin. Reprod. Med. 2005, 23, 74–81. [Google Scholar] [CrossRef]
- Hackenberg, R.; Schulz, K.D. Androgen receptor mediated growth control of breast cancer and endometrial cancer modulated by antiandrogen- and androgen-like steroids. J. Steroid Biochem. Mol. Biol. 1996, 56, 113–117. [Google Scholar] [CrossRef]
- Hackenberg, R.; Beck, S.; Filmer, A.; Nia, A.H.; Kunzmann, R.; Koch, M.; Slater, E.P.; Schulz, K.D. Androgen responsiveness of the new human endometrial cancer cell line MFE-296. Int. J. Cancer 1994, 57, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Sasagawa, S.; Shimizu, Y.; Kami, H.; Takeuchi, T.; Mita, S.; Imada, K.; Kato, S.; Mizuguchi, K. Dienogest is a selective progesterone receptor agonist in transactivation analysis with potent oral endometrial activity due to its efficient pharmacokinetic profile. Steroids 2008, 73, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.X.; Yeap, B.B. Dihydrotestosterone and cancer risk. Curr. Opin. Endocrinol. Diabetes Obes. 2018, 25, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Koivisto, C.S.; Parrish, M.; Bonala, S.B.; Ngoi, S.; Torres, A.; Gallagher, J.; Sanchez-Hodge, R.; Zeinner, V.; Nahhas, G.J.; Liu, B.; et al. Evaluating the efficacy of enzalutamide and the development of resistance in a preclinical mouse model of type-I endometrial carcinoma. Neoplasia 2020, 22, 484–496. [Google Scholar] [CrossRef]
- Mahdi, Z.; Abdulfatah, E.; Pardeshi, V.; Hassan, O.; Schultz, D.; Morris, R.; Cote, M.L.; Elshaikh, M.A.; Bandyopadhyay, S.; Ali-Fehmi, R. The Impact of Androgen Receptor Expression on Endometrial Carcinoma Recurrence and Survival. Int. J. Gynecol. Pathol. 2017, 36, 405–411. [Google Scholar] [CrossRef]
- Kamal, A.M.; Bulmer, J.N.; DeCruze, S.B.; Stringfellow, H.F.; Martin-Hirsch, P.; Hapangama, D.K. Androgen receptors are acquired by healthy postmenopausal endometrial epithelium and their subsequent loss in endometrial cancer is associated with poor survival. Br. J. Cancer 2016, 114, 688–696. [Google Scholar] [CrossRef]
- Wu, X.; Zhong, X.; Huo, X.; Zhang, J.; Yang, X.; Zhang, Y. The Clinicopathological Significance and Prognostic Value of Androgen Receptor in Endometrial Carcinoma: A Meta-Analysis. Front. Oncol. 2022, 12, 905809. [Google Scholar] [CrossRef]
- Gan, Q.; Crumley, S.; Broaddus, R.R. Molecular Modifiers of Hormone Receptor Action: Decreased Androgen Receptor Expression in Mismatch Repair Deficient Endometrial Endometrioid Adenocarcinoma. Int. J. Gynecol. Pathol. 2019, 38, 44–51. [Google Scholar] [CrossRef]
- Sasaki, M.; Sakuragi, N.; Dahiya, R. The CAG repeats in exon 1 of the androgen receptor gene are significantly longer in endometrial cancer patients. Biochem. Biophys. Res. Commun. 2003, 305, 1105–1108. [Google Scholar] [CrossRef]
- Sasaki, M.; Oh, B.R.; Dharia, A.; Fujimoto, S.; Dahiya, R. Inactivation of the human androgen receptor gene is associated with CpG hypermethylation in uterine endometrial cancer. Mol. Carcinog. 2000, 29, 59–66. [Google Scholar] [CrossRef]
- Murali, R.; Davidson, B.; Fadare, O.; Carlson, J.A.; Crum, C.P.; Gilks, C.B.; Irving, J.A.; Malpica, A.; Matias-Guiu, X.; McCluggage, W.G.; et al. High-grade Endometrial Carcinomas: Morphologic and Immunohistochemical Features, Diagnostic Challenges and Recommendations. Int. J. Gynecol. Pathol. 2019, 38 (Suppl. 1), S40–S63. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Hacking, S.; Chavarria-Bernal, H.D.; Bhuiya, T.A.; Khutti, S. Androgen Receptor Immunohistochemical Expression in Undifferentiated/Dedifferentiated Endometrial Carcinoma. Int. J. Gynecol. Pathol. 2022, 41, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Munoz, J.; Wheler, J.J.; Kurzrock, R. Androgen receptors beyond prostate cancer: An old marker as a new target. Oncotarget 2015, 6, 592–603. [Google Scholar] [CrossRef]
- Mills, A.M.; Gottlieb, C.E.; Wendroth, S.M.; Brenin, C.M.; Atkins, K.A. Pure Apocrine Carcinomas Represent a Clinicopathologically Distinct Androgen Receptor-Positive Subset of Triple-Negative Breast Cancers. Am. J. Surg. Pathol. 2016, 40, 1109–1116. [Google Scholar] [CrossRef]
- Zadeh, S.L.; Duska, L.R.; Mills, A.M. Androgen Receptor Expression in Endometrial Carcinoma. Int. J. Gynecol. Pathol. 2018, 37, 167–173. [Google Scholar] [CrossRef]
- Gibson, D.A.; Simitsidellis, I.; Saunders, P.T. Regulation of androgen action during establishment of pregnancy. J. Mol. Endocrinol. 2016, 57, R35–R47. [Google Scholar] [CrossRef]
- Gellersen, B.; Brosens, J.J. Cyclic decidualization of the human endometrium in reproductive health and failure. Endocr. Rev. 2014, 35, 851–905. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.A.; Simitsidellis, I.; Cousins, F.L.; Critchley, H.O.; Saunders, P.T. Intracrine Androgens Enhance Decidualization and Modulate Expression of Human Endometrial Receptivity Genes. Sci. Rep. 2016, 6, 19970. [Google Scholar] [CrossRef]
- Gibson, D.A.; Simitsidellis, I.; Kelepouri, O.; Critchley, H.O.D.; Saunders, P.T.K. Dehydroepiandrosterone enhances decidualization in women of advanced reproductive age. Fertil. Steril. 2018, 109, 728–734.e722. [Google Scholar] [CrossRef]
- Young, S.L. Androgens and endometrium: New lessons from the corpus luteum via the adrenal cortex? Fertil. Steril. 2018, 109, 623–624. [Google Scholar] [CrossRef]
- Diao, H.L.; Su, R.W.; Tan, H.N.; Li, S.J.; Lei, W.; Deng, W.B.; Yang, Z.M. Effects of androgen on embryo implantation in the mouse delayed-implantation model. Fertil. Steril. 2008, 90, 1376–1383. [Google Scholar] [CrossRef]
- Gleicher, N.; Kim, A.; Weghofer, A.; Kushnir, V.A.; Shohat-Tal, A.; Lazzaroni, E.; Lee, H.J.; Barad, D.H. Hypoandrogenism in association with diminished functional ovarian reserve. Hum. Reprod. 2013, 28, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Barad, D.; Brill, H.; Gleicher, N. Update on the use of dehydroepiandrosterone supplementation among women with diminished ovarian function. J. Assist. Reprod. Genet. 2007, 24, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Kara, M.; Aydin, T.; Aran, T.; Turktekin, N.; Ozdemir, B. Does dehydroepiandrosterone supplementation really affect IVF-ICSI outcome in women with poor ovarian reserve? Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 173, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Burris, T.P.; Solt, L.A.; Wang, Y.; Crumbley, C.; Banerjee, S.; Griffett, K.; Lundasen, T.; Hughes, T.; Kojetin, D.J. Nuclear receptors and their selective pharmacologic modulators. Pharmacol. Rev. 2013, 65, 710–778. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, R.; Coss, C.C.; Dalton, J.T. Development of selective androgen receptor modulators (SARMs). Mol. Cell. Endocrinol. 2018, 465, 134–142. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).