
Network Theoretical Approach to Explore Factors Affecting Signal Propagation and Stability in Dementia’s Protein-Protein Interaction Network

 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Acquisition of Data
2.2. Annotation Enrichment/Over-Representation Analysis
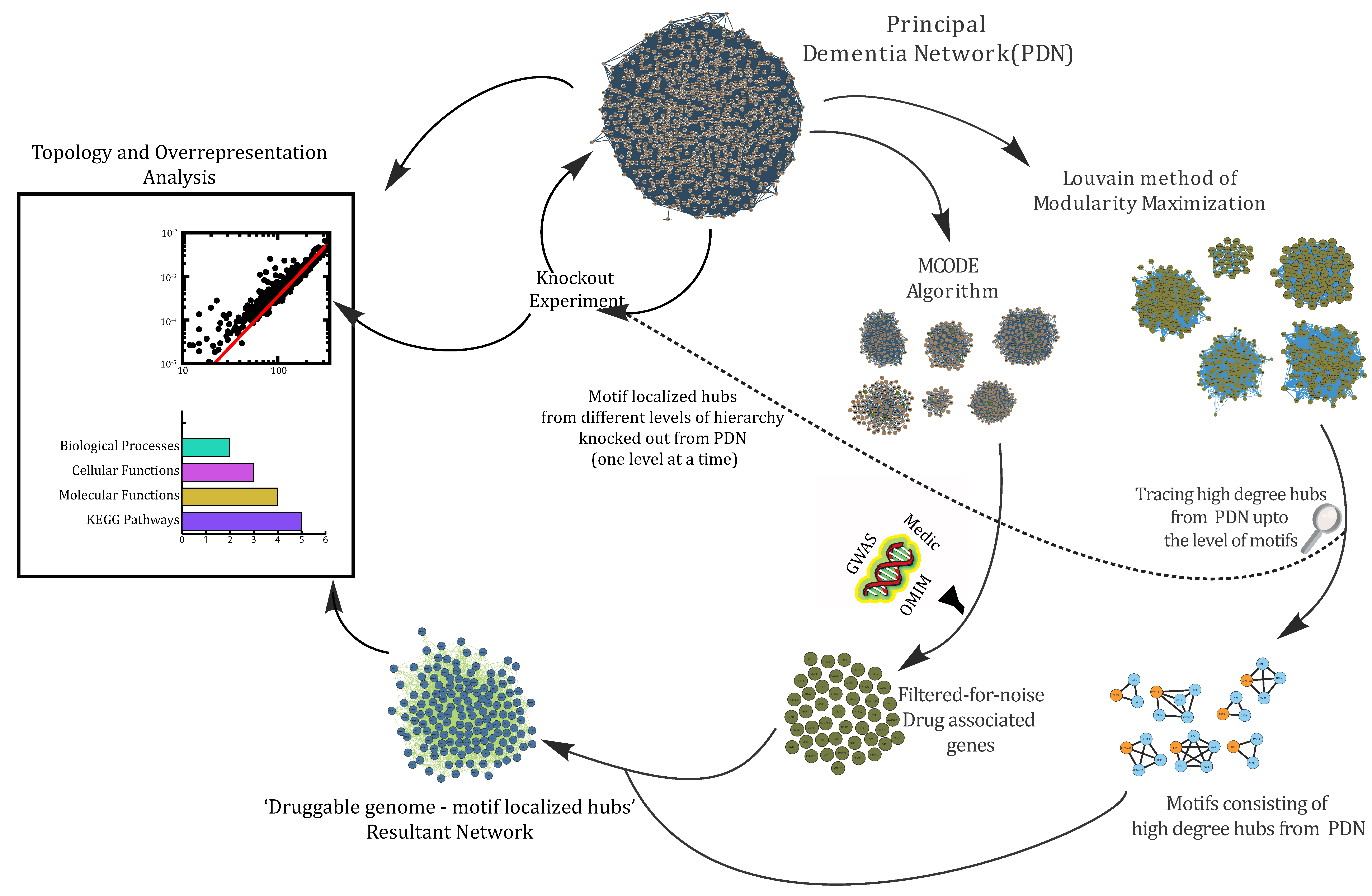
2.3. Construction of Protein–Protein/Gene–Gene Interaction (PPI) Networks
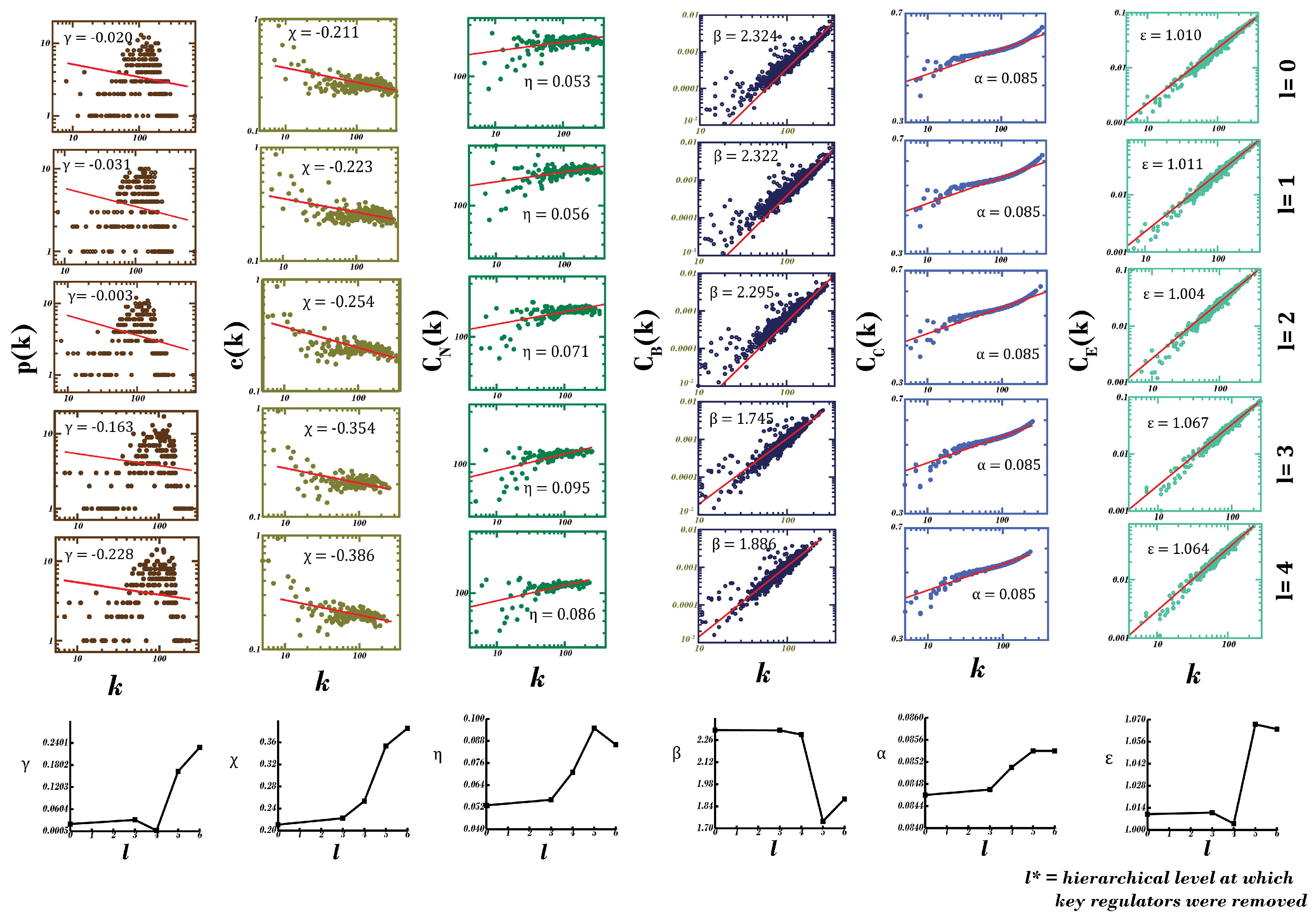
2.4. Characterization of Topological Properties of Networks
2.4.1. Degree Distribution, p(k)
2.4.2. Clustering Coefficient, c(k)
2.4.3. Neighborhood Connectivity Distribution, CN(k)
2.4.4. Closeness Centrality, CC(k)
2.4.5. Eigenvector Centrality, CE(k)
2.4.6. Betweenness Centrality, CB(k)
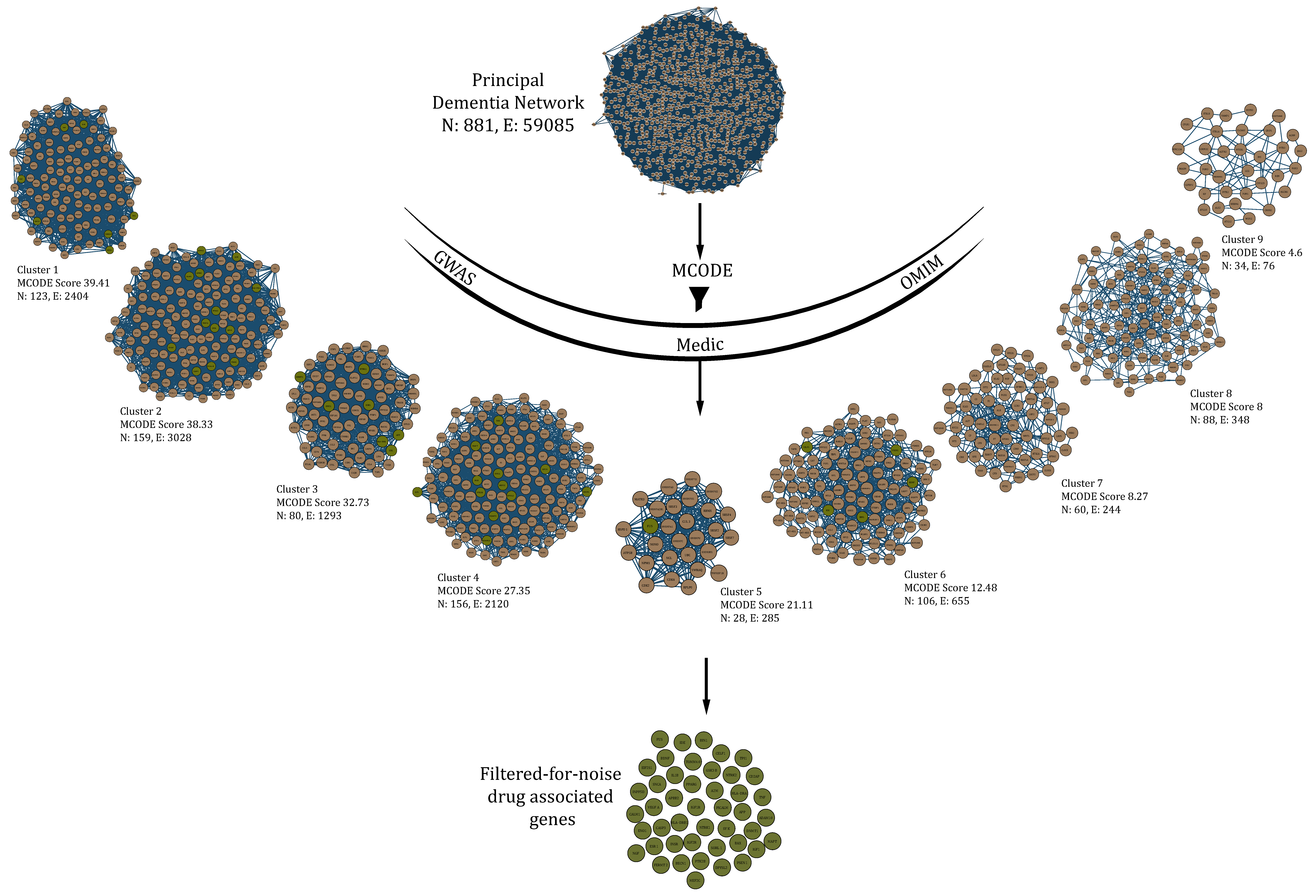
2.5. MCODE (Molecular Complex Detection)-Derived Protein Complexes to Filter Drug-Actionable Genes in the Network
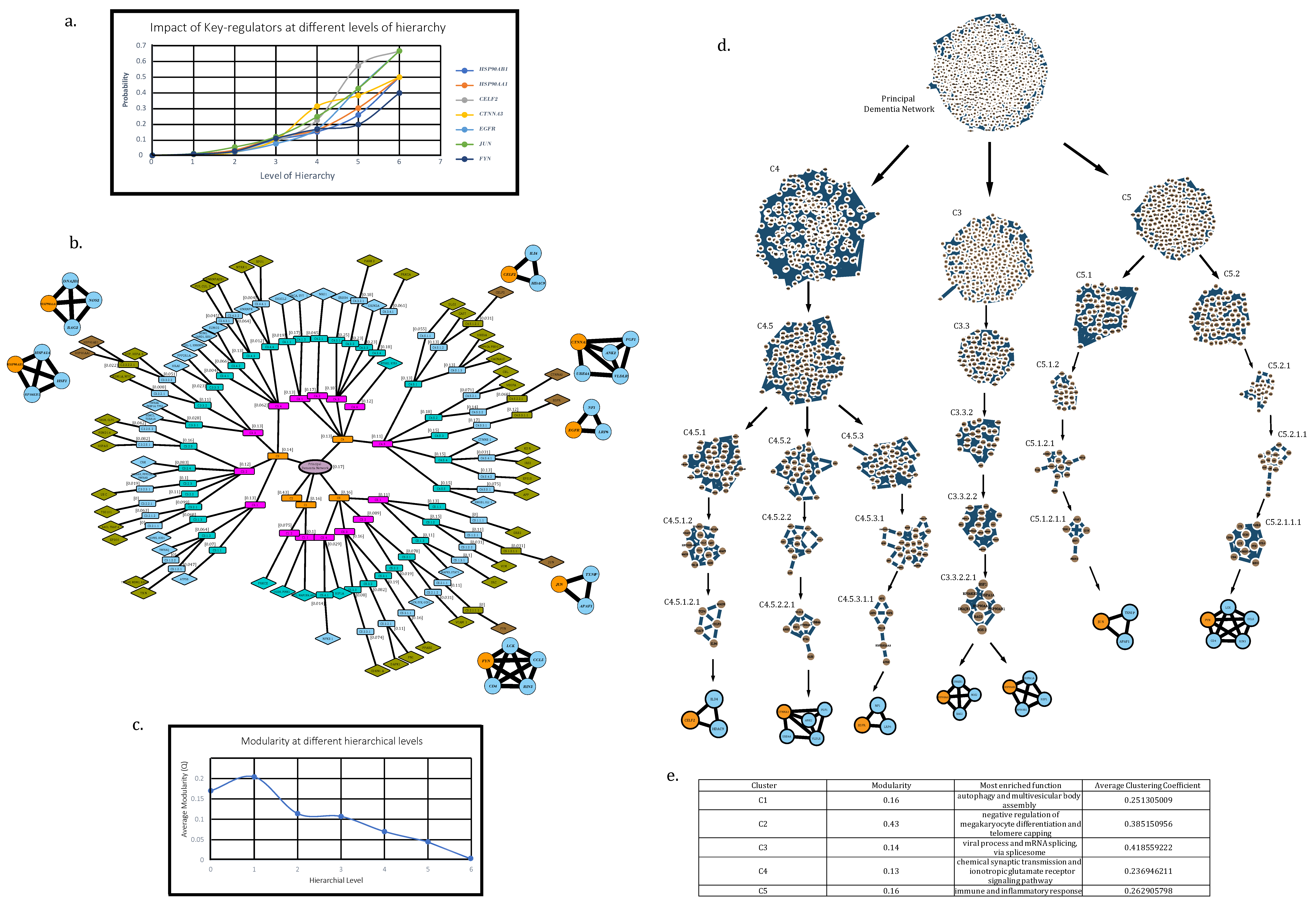
2.6. Detection of Key Regulators (KRs)
2.7. Knockout Experiment
2.8. Validation of Expression Patterns
3. Results
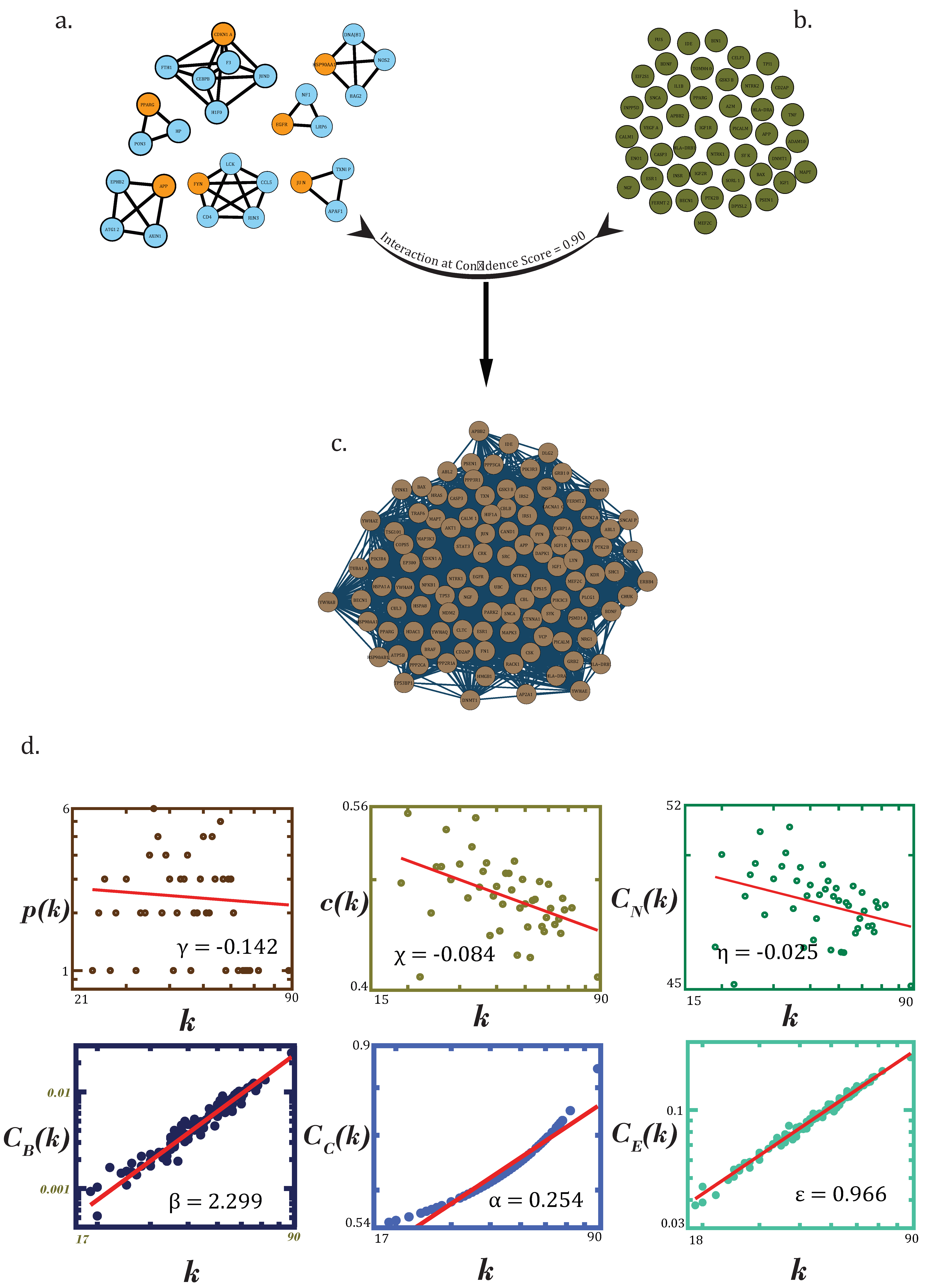
3.1. Data Acquisition and Principal Dementia Network
3.2. Gene-Ontology-Based Overrepresentation Analysis
3.3. Alzheimer’s Disease and Other Dementias’ PPI Networks Exhibit Hierarchical, Scale-free Topologies
3.4. Filtering Drug-Actionable Genes for Noise through Dense Clusters Obtained from the PDN
3.5. Key Regulators
3.6. Assessment of the Network’s Stability
3.7. Interaction Analysis of Druggable Genome and Network’s Stability
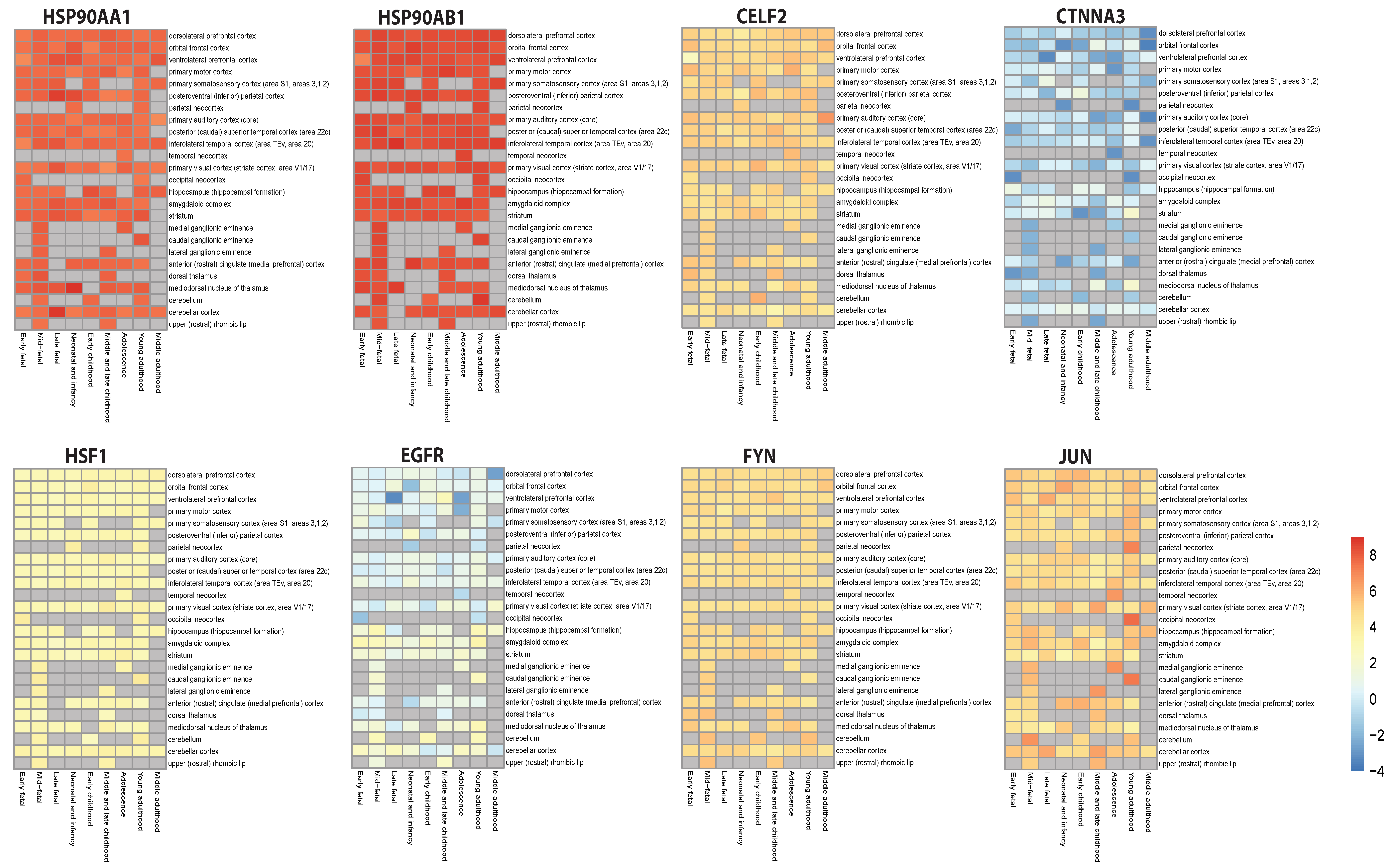
3.8. Validation of Key Regulators’ Expression Patterns
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD. Global, regional, and national burden of Alzheimer’s disease and other dementias, 1990-2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 88–106. [Google Scholar] [CrossRef] [Green Version]
- Korsnes, M.S.; Winkler, A.S. Global, regional, and national burden of dementia, 1990–2016: Predictions need local calibration. Neurology 2020, 94, 718–719. [Google Scholar] [CrossRef] [Green Version]
- Livingston, G.; Sommerlad, A.; Orgeta, V.; Costafreda, S.G.; Huntley, J.; Ames, D.; Ballard, C.; Banerjee, S.; Burns, A.; Cohen-Mansfield, J.; et al. Dementia prevention, intervention, and care. Lancet 2017, 390, 2673–2734. [Google Scholar] [CrossRef] [Green Version]
- Martins, I.J.; Berger, T.; Sharman, M.J.; Verdile, G.; Fuller, S.J.; Martins, R.N. Cholesterol metabolism and transport in the pathogenesis of Alzheimer’s disease. J. Neurochem. 2009, 111, 1275–1308. [Google Scholar] [CrossRef]
- Rogan, S.; Lippa, C.F. Alzheimer’s disease and other dementias: A review. Am. J. Alzheimers Dis. Other Demen. 2002, 17, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Paulson, H.L.; Igo, I. Genetics of dementia. Semin. Neurol. 2011, 31, 449–460. [Google Scholar] [CrossRef]
- Krisko, A.; Radman, M. Protein damage, ageing and age-related diseases. Open Biol. 2019, 9, 180249. [Google Scholar] [CrossRef] [Green Version]
- Guerreiro, R.; Bras, J. The age factor in Alzheimer’s disease. Genome Med. 2015, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.S.; Kågedal, K.; Halliday, G.M. Alpha-synuclein biology in Lewy body diseases. Alzheimers Res. Ther. 2014, 6, 73. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.T.; Grossman, M. TDP-43 and frontotemporal dementia. Curr. Neurol. Neurosci. Rep. 2009, 9, 353–358. [Google Scholar] [CrossRef]
- Neary, D.; Snowden, J.; Mann, D. Frontotemporal dementia. Lancet Neurol. 2005, 4, 771–780. [Google Scholar] [CrossRef]
- Neumann, M.; Rademakers, R.; Roeber, S.; Baker, M.; Kretzschmar, H.A.; Mackenzie, I.R. A new subtype of frontotemporal lobar degeneration with FUS pathology. Brain 2009, 132, 2922–2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pottier, C.; Ravenscroft, T.A.; Sanchez-Contreras, M.; Rademakers, R. Genetics of FTLD: Overview and what else we can expect from genetic studies. J. Neurochem. 2016, 138 (Suppl. 1), 32–53. [Google Scholar] [CrossRef] [PubMed]
- Rademakers, R.; Cruts, M.; van Broeckhoven, C. The role of tau (MAPT) in frontotemporal dementia and related tauopathies. Hum. Mutat. 2004, 24, 277–295. [Google Scholar] [CrossRef] [PubMed]
- Thinakaran, G. The role of presenilins in Alzheimer’s disease. J. Clin. Investig. 1999, 104, 1321–1327. [Google Scholar] [CrossRef] [Green Version]
- Papakonstantinou, E.; Bacopoulou, F.; Brouzas, D.; Megalooikonomou, V.; D’Elia, D.; Bongcam-Rudloff, E.; Vlachakis, D. NOTCH3 and CADASIL syndrome: A genetic and structural overview. EMBnet J. 2019, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, P.; Zhao, Z.; Hulgan, T.; Bush, W.S.; Samuels, D.C.; Bloss, C.S.; Heaton, R.K.; Ellis, R.J.; Schork, N.; Marra, C.M.; et al. Genome-wide association study of HIV-associated neurocognitive disorder (HAND): A CHARTER group study. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2017, 174, 413–426. [Google Scholar] [CrossRef]
- O’Brien, R.J.; Wong, P.C. Amyloid precursor protein processing and Alzheimer’s disease. Annu. Rev. Neurosci. 2011, 34, 185–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohrer, J.D.; Guerreiro, R.; Vandrovcova, J.; Uphill, J.; Reiman, D.; Beck, J.; Isaacs, A.M.; Authier, A.; Ferrari, R.; Fox, N.C.; et al. The heritability and genetics of frontotemporal lobar degeneration. Neurology 2009, 73, 1451–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuang, D.W.; DiGiacomo, L.; Bird, T.D. Familial occurrence of dementia with Lewy bodies. Am. J. Geriatr. Psychiatry 2004, 12, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Clifford, D.B.; Ances, B.M. HIV-associated neurocognitive disorder. Lancet Infect. Dis. 2013, 13, 976–986. [Google Scholar] [CrossRef] [Green Version]
- Cho, D.-Y.; Kim, Y.-A.; Przytycka, T.M. Chapter 5: Network Biology Approach to Complex Diseases. PLoS Comput. Biol. 2012, 8, e1002820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverman, E.K.; Loscalzo, J. Network medicine approaches to the genetics of complex diseases. Discov. Med. 2012, 14, 143–152. [Google Scholar]
- Barabási, A.L.; Gulbahce, N.; Loscalzo, J. Network medicine: A network-based approach to human disease. Nat. Rev. Genet. 2011, 12, 56–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, J.A.; Bottero, V.; Potashkin, J.A. Transcriptomic and Network Analysis Highlight the Association of Diabetes at Different Stages of Alzheimer’s Disease. Front. Neurosci. 2019, 13, 1273. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Rocke, D.M.; Perry, G.; Ray, M. Differential network analyses of Alzheimer’s disease identify early events in Alzheimer’s disease pathology. Int. J. Alzheimers Dis. 2014, 2014, 721453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Ma, C.; Gearing, M.; Wang, P.G.; Chin, L.S.; Li, L. Integrated proteomics and network analysis identifies protein hubs and network alterations in Alzheimer’s disease. Acta Neuropathol. Commun. 2018, 6, 19. [Google Scholar] [CrossRef]
- Mizuno, S.; Ogishima, S.; Kitatani, K.; Kikuchi, M.; Tanaka, H.; Yaegashi, N.; Nakaya, J. Network Analysis of a Comprehensive Knowledge Repository Reveals a Dual Role for Ceramide in Alzheimer’s Disease. PLoS ONE 2016, 11, e0148431. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.; Lovering, R.C.; Hardy, J.; Lewis, P.A.; Manzoni, C. Weighted Protein Interaction Network Analysis of Frontotemporal Dementia. J. Proteome Res. 2017, 16, 999–1013. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amberger, J.; Bocchini, C.A.; Scott, A.F.; Hamosh, A. McKusick’s Online Mendelian Inheritance in Man (OMIM). Nucleic Acids Res. 2009, 37, D793–D796. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Wiegers, T.C.; Rosenstein, M.C.; Mattingly, C.J. MEDIC: A practical disease vocabulary used at the Comparative Toxicogenomics Database. Database 2012, 2012, bar065. [Google Scholar] [CrossRef]
- Wang, L.; Khankhanian, P.; Baranzini, S.E.; Mousavi, P. iCTNet: A Cytoscape plugin to produce and analyze integrative complex traits networks. BMC Bioinform. 2011, 12, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Taye, B.; Vaz, C.; Tanavde, V.; Kuznetsov, V.A.; Eisenhaber, F.; Sugrue, R.J.; Maurer-Stroh, S. Benchmarking selected computational gene network growing tools in context of virus-host interactions. Sci. Rep. 2017, 7, 5805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.K.; Carlin, D.E.; Yu, M.K.; Zhang, W.; Kreisberg, J.F.; Tamayo, P.; Ideker, T. Systematic Evaluation of Molecular Networks for Discovery of Disease Genes. Cell Syst. 2018, 6, 484–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiboonchoe, A.; Samarasinghe, S.; Kulasiri, D.; Salehi-Ashtiani, K. Integrated analysis of gene network in childhood leukemia from microarray and pathway databases. Biomed. Res. Int. 2014, 2014, 278748. [Google Scholar] [CrossRef] [PubMed]
- Vlasblom, J.; Zuberi, K.; Rodriguez, H.; Arnold, R.; Gagarinova, A.; Deineko, V.; Kumar, A.; Leung, E.; Rizzolo, K.; Samanfar, B.; et al. Novel function discovery with GeneMANIA: A new integrated resource for gene function prediction in Escherichia coli. Bioinformatics 2015, 31, 306–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef]
- Albert, R.; Barabási, A.-L. Statistical mechanics of complex networks. Rev. Mod. Phys. 2002, 74, 47–97. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Li, M.; Wang, J.; Pan, Y.; Wu, F.X. CytoNCA: A cytoscape plugin for centrality analysis and evaluation of protein interaction networks. Biosystems 2015, 127, 67–72. [Google Scholar] [CrossRef]
- Assenov, Y.; Ramirez, F.; Schelhorn, S.E.; Lengauer, T.; Albrecht, M. Computing topological parameters of biological networks. Bioinformatics 2008, 24, 282–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravasz, E. Detecting hierarchical modularity in biological networks. Methods Mol. Biol. 2009, 541, 145–160. [Google Scholar] [CrossRef]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of ‘small-world’ networks. Nature 1998, 393, 440–442. [Google Scholar] [CrossRef]
- Ravasz, E.; Barabasi, A.L. Hierarchical organization in complex networks. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2003, 67, 026112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerqueti, R.; Ferraro, G.; Iovanella, A. A new measure for community structures through indirect social connections. Expert Syst. Appl. 2018, 114, 196–209. [Google Scholar] [CrossRef] [Green Version]
- Maslov, S.; Sneppen, K. Specificity and stability in topology of protein networks. Science 2002, 296, 910–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor-Satorras, R.; Vázquez, A.; Vespignani, A. Dynamical and correlation properties of the internet. Phys. Rev. Lett. 2001, 87, 258701. [Google Scholar] [CrossRef] [Green Version]
- Barrat, A.; Barthélemy, M.; Vespignani, A. The Architecture of Complex Weighted Networks: Measurements and Models. In Large Scale Structure and Dynamics of Complex Networks; Caldarelli, G., Vespignani, A., Eds.; World Scientific: Singapore, 2007; Volume 2, pp. 67–92. [Google Scholar]
- Canright, G.; Engø-Monsen, K. Roles in networks. Sci. Comp. Program. 2004, 53, 195–214. [Google Scholar] [CrossRef] [Green Version]
- Canright, G.S.; Engø-Monsen, K. Spreading on Networks: A Topographic View. Complexus 2006, 3, 131–146. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Kim, P.M.; Sprecher, E.; Trifonov, V.; Gerstein, M. The importance of bottlenecks in protein networks: Correlation with gene essentiality and expression dynamics. PLoS Comput. Biol. 2007, 3, e59. [Google Scholar] [CrossRef]
- Freeman, L.C. A set of measures of centrality based on betweenness. Sociometry 1977, 40, 35–41. [Google Scholar] [CrossRef]
- Bader, G.D.; Hogue, C.W. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterJ. Complex syst. 2006, 1695, 1–9. [Google Scholar]
- Blondel, V.D.; Guillaume, J.-L.; Lambiotte, R.; Lefebvre, E. Fast unfolding of communities in large networks. J. Stat. Mech. Theory Exp. 2008, 2008, P10008. [Google Scholar] [CrossRef] [Green Version]
- Sunkin, S.M.; Ng, L.; Lau, C.; Dolbeare, T.; Gilbert, T.L.; Thompson, C.L.; Hawrylycz, M.; Dang, C. Allen Brain Atlas: An integrated spatio-temporal portal for exploring the central nervous system. Nucleic Acids Res. 2013, 41, D996–D1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Lin, W.; Zhang, Y.; Li, W.; Wang, J. BEST: A web server for brain expression Spatio-temporal pattern analysis. BMC Bioinform. 2019, 20, 632. [Google Scholar] [CrossRef] [Green Version]
- Clauset, A.; Shalizi, C.R.; Newman, M.E.J. Power-Law Distributions in Empirical Data. SIAM Rev. 2009, 51, 661–703. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F.K.; Deris, S.; Othman, N.H. The inference of breast cancer metastasis through gene regulatory networks. J. Biomed. Inform. 2012, 45, 350–362. [Google Scholar] [CrossRef] [Green Version]
- Barabási, A.L.; Oltvai, Z.N. Network biology: Understanding the cell’s functional organization. Nat. Rev. Genet. 2004, 5, 101–113. [Google Scholar] [CrossRef]
- Borgatti, S.P.; Everett, M.G. A Graph-theoretic perspective on centrality. Soc. Netw. 2006, 28, 466–484. [Google Scholar] [CrossRef]
- Milo, R.; Shen-Orr, S.; Itzkovitz, S.; Kashtan, N.; Chklovskii, D.; Alon, U. Network motifs: Simple building blocks of complex networks. Science 2002, 298, 824–827. [Google Scholar] [CrossRef] [Green Version]
- Stone, L.; Simberloff, D.; Artzy-Randrup, Y. Network motifs and their origins. PLoS Comput. Biol. 2019, 15, e1006749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoter, A.; El-Sabban, M.E.; Naim, H.Y. The HSP90 Family: Structure, Regulation, Function, and Implications in Health and Disease. Int. J. Mol. Sci. 2018, 19, 2560. [Google Scholar] [CrossRef] [Green Version]
- Haase, M.; Fitze, G. HSP90AB1: Helping the good and the bad. Gene 2016, 575, 171–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackie, R.E.; Maciejewski, A.; Ostapchenko, V.G.; Marques-Lopes, J.; Choy, W.Y.; Duennwald, M.L.; Prado, V.F.; Prado, M.A.M. The Hsp70/Hsp90 Chaperone Machinery in Neurodegenerative Diseases. Front. Neurosci. 2017, 11, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClellan, A.J.; Xia, Y.; Deutschbauer, A.M.; Davis, R.W.; Gerstein, M.; Frydman, J. Diverse cellular functions of the Hsp90 molecular chaperone uncovered using systems approaches. Cell 2007, 131, 121–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schopf, F.H.; Biebl, M.M.; Buchner, J. The HSP90 chaperone machinery. Nat. Rev. Mol. Cell Biol. 2017, 18, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, G.G. Tauopathies. Handb. Clin. Neurol. 2017, 145, 355–368. [Google Scholar] [CrossRef]
- Alvarez, G.; Muñoz-Montaño, J.R.; Satrústegui, J.; Avila, J.; Bogónez, E.; Díaz-Nido, J. Lithium protects cultured neurons against beta-amyloid-induced neurodegeneration. FEBS Lett. 1999, 453, 260–264. [Google Scholar] [CrossRef] [Green Version]
- Avila, J.; Wandosell, F.; Hernández, F. Role of glycogen synthase kinase-3 in Alzheimer’s disease pathogenesis and glycogen synthase kinase-3 inhibitors. Expert Rev. Neurother. 2010, 10, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, M.A.; Acker, C.M.; Davies, P. Tau phosphorylated at tyrosine 394 is found in Alzheimer’s disease tangles and can be a product of the Abl-related kinase, Arg. J. Alzheimers Dis. 2010, 19, 721–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavallini, A.; Brewerton, S.; Bell, A.; Sargent, S.; Glover, S.; Hardy, C.; Moore, R.; Calley, J.; Ramachandran, D.; Poidinger, M.; et al. An unbiased approach to identifying tau kinases that phosphorylate tau at sites associated with Alzheimer disease. J. Biol. Chem. 2013, 288, 23331–23347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, S.D.; Whitehead, S.N.; Swayne, L.A.; Moffat, T.C.; Hou, W.; Ethier, M.; Bourgeois, A.J.; Rashidian, J.; Blanchard, A.P.; Fraser, P.E.; et al. Amyloid-beta42 signals tau hyperphosphorylation and compromises neuronal viability by disrupting alkylacylglycerophosphocholine metabolism. Proc. Natl. Acad. Sci. USA 2009, 106, 20936–20941. [Google Scholar] [CrossRef] [Green Version]
- Hernández, F.; Gómez de Barreda, E.; Fuster-Matanzo, A.; Lucas, J.J.; Avila, J. GSK3: A possible link between beta amyloid peptide and tau protein. Exp. Neurol. 2010, 223, 322–325. [Google Scholar] [CrossRef] [PubMed]
- Tamagno, E.; Robino, G.; Obbili, A.; Bardini, P.; Aragno, M.; Parola, M.; Danni, O. H2O2 and 4-hydroxynonenal mediate amyloid beta-induced neuronal apoptosis by activating JNKs and p38MAPK. Exp. Neurol. 2003, 180, 144–155. [Google Scholar] [CrossRef]
- Mucke, L.; Selkoe, D.J. Neurotoxicity of amyloid β-protein: Synaptic and network dysfunction. Cold Spring Harb. Perspect. Med. 2012, 2, a006338. [Google Scholar] [CrossRef] [Green Version]
- Dou, F.; Chang, X.; Ma, D. Hsp90 Maintains the Stability and Function of the Tau Phosphorylating Kinase GSK3β. Int. J. Mol. Sci. 2007, 8, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, B.; Liu, D.; Li, J.J.; Xue, Y.; Sakata, K.; Zhu, L.Q.; Heldt, S.A.; Xu, H.; Liao, F.F. Hsp90 chaperone inhibitor 17-AAG attenuates Aβ-induced synaptic toxicity and memory impairment. J. Neurosci. 2014, 34, 2464–2470. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, Y.; Huang, L.; Chen, J.; Li, J.J.; Wang, R.; Kim, E.; Chen, Y.; Justicia, C.; Sakata, K.; et al. A CNS-permeable Hsp90 inhibitor rescues synaptic dysfunction and memory loss in APP-overexpressing Alzheimer’s mouse model via an HSF1-mediated mechanism. Mol. Psychiatry 2017, 22, 990–1001. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Rodina, A.; Chiosis, G. Heat shock protein 90: Translation from cancer to Alzheimer’s disease treatment? BMC Neurosci. 2008, 9 (Suppl. 2), S7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spillantini, M.G.; Van Swieten, J.C.; Goedert, M. Tau gene mutations in frontotemporal dementia and parkinsonism linked to chromosome 17 (FTDP-17). Neurogenetics 2000, 2, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Neumann, M.; Sampathu, D.M.; Kwong, L.K.; Truax, A.C.; Micsenyi, M.C.; Chou, T.T.; Bruce, J.; Schuck, T.; Grossman, M.; Clark, C.M.; et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 2006, 314, 130–133. [Google Scholar] [CrossRef] [Green Version]
- Mackenzie, I.R.; Bigio, E.H.; Ince, P.G.; Geser, F.; Neumann, M.; Cairns, N.J.; Kwong, L.K.; Forman, M.S.; Ravits, J.; Stewart, H.; et al. Pathological TDP-43 distinguishes sporadic amyotrophic lateral sclerosis from amyotrophic lateral sclerosis with SOD1 mutations. Ann. Neurol. 2007, 61, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Sreedharan, J.; Blair, I.P.; Tripathi, V.B.; Hu, X.; Vance, C.; Rogelj, B.; Ackerley, S.; Durnall, J.C.; Williams, K.L.; Buratti, E.; et al. TDP-43 mutations in familial and sporadic amyotrophic lateral sclerosis. Science 2008, 319, 1668–1672. [Google Scholar] [CrossRef]
- Brettschneider, J.; Del Tredici, K.; Toledo, J.B.; Robinson, J.L.; Irwin, D.J.; Grossman, M.; Suh, E.; Van Deerlin, V.M.; Wood, E.M.; Baek, Y.; et al. Stages of pTDP-43 pathology in amyotrophic lateral sclerosis. Ann. Neurol. 2013, 74, 20–38. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Gendron, T.F.; Xu, Y.F.; Ko, L.W.; Yen, S.H.; Petrucelli, L. Phosphorylation regulates proteasomal-mediated degradation and solubility of TAR DNA binding protein-43 C-terminal fragments. Mol. Neurodegener. 2010, 5, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliday, G.M.; Holton, J.L.; Revesz, T.; Dickson, D.W. Neuropathology underlying clinical variability in patients with synucleinopathies. Acta Neuropathol. 2011, 122, 187–204. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Pozo, A.; Frosch, M.P.; Masliah, E.; Hyman, B.T. Neuropathological alterations in Alzheimer disease. Cold Spring Harb. Perspect. Med. 2011, 1, a006189. [Google Scholar] [CrossRef]
- Ingelsson, M. Alpha-Synuclein Oligomers-Neurotoxic Molecules in Parkinson’s Disease and Other Lewy Body Disorders. Front. Neurosci. 2016, 10, 408. [Google Scholar] [CrossRef] [Green Version]
- Uryu, K.; Richter-Landsberg, C.; Welch, W.; Sun, E.; Goldbaum, O.; Norris, E.H.; Pham, C.T.; Yazawa, I.; Hilburger, K.; Micsenyi, M.; et al. Convergence of heat shock protein 90 with ubiquitin in filamentous alpha-synuclein inclusions of alpha-synucleinopathies. Am. J. Pathol. 2006, 168, 947–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falsone, S.F.; Kungl, A.J.; Rek, A.; Cappai, R.; Zangger, K. The molecular chaperone Hsp90 modulates intermediate steps of amyloid assembly of the Parkinson-related protein alpha-synuclein. J. Biol. Chem. 2009, 284, 31190–31199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.S.; Chung, J.H.; Oh, B.H.; Hong, C.H. Increased plasma levels of heat shock protein 70 in patients with vascular mild cognitive impairment. Neurosci. Lett. 2008, 436, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Sun, W.; Taldone, T.; Rodina, A.; Chiosis, G. Heat shock protein 90 in neurodegenerative diseases. Mol. Neurodegener. 2010, 5, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filone, C.M.; Caballero, I.S.; Dower, K.; Mendillo, M.L.; Cowley, G.S.; Santagata, S.; Rozelle, D.K.; Yen, J.; Rubins, K.H.; Hacohen, N.; et al. The master regulator of the cellular stress response (HSF1) is critical for orthopoxvirus infection. PLoS Pathog. 2014, 10, e1003904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, C. The heat-shock, or HSF1-mediated proteotoxic stress, response in cancer: From proteomic stability to oncogenesis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20160525. [Google Scholar] [CrossRef]
- Klettner, A. The induction of heat shock proteins as a potential strategy to treat neurodegenerative disorders. Drug News Perspect. 2004, 17, 299–306. [Google Scholar] [CrossRef]
- Brown, I.R. Heat shock proteins and protection of the nervous system. Ann. N. Y. Acad. Sci. 2007, 1113, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Muchowski, P.J.; Wacker, J.L. Modulation of neurodegeneration by molecular chaperones. Nat. Rev. Neurosci. 2005, 6, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Anckar, J.; Sistonen, L. Heat shock factor 1 as a coordinator of stress and developmental pathways. Adv. Exp. Med. Biol. 2007, 594, 78–88. [Google Scholar] [CrossRef]
- Hietakangas, V.; Ahlskog, J.K.; Jakobsson, A.M.; Hellesuo, M.; Sahlberg, N.M.; Holmberg, C.I.; Mikhailov, A.; Palvimo, J.J.; Pirkkala, L.; Sistonen, L. Phosphorylation of serine 303 is a prerequisite for the stress-inducible SUMO modification of heat shock factor 1. Mol. Cell Biol. 2003, 23, 2953–2968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelin, E.; Freeman, B.C. Lysine deacetylases regulate the heat shock response including the age-associated impairment of HSF1. J. Mol. Biol. 2015, 427, 1644–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purwana, I.; Liu, J.J.; Portha, B.; Buteau, J. HSF1 acetylation decreases its transcriptional activity and enhances glucolipotoxicity-induced apoptosis in rat and human beta cells. Diabetologia 2017, 60, 1432–1441. [Google Scholar] [CrossRef]
- Hilgarth, R.S.; Hong, Y.; Park-Sarge, O.K.; Sarge, K.D. Insights into the regulation of heat shock transcription factor 1 SUMO-1 modification. Biochem. Biophys. Res. Commun. 2003, 303, 196–200. [Google Scholar] [CrossRef]
- Dewji, N.N.; Do, C. Heat shock factor-1 mediates the transcriptional activation of Alzheimer’s beta-amyloid precursor protein gene in response to stress. Brain Res. Mol. Brain Res. 1996, 35, 325–328. [Google Scholar] [CrossRef]
- Kourtis, N.; Tavernarakis, N. Cellular stress response pathways and ageing: Intricate molecular relationships. EMBO J. 2011, 30, 2520–2531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Guo, Y.; Guettouche, T.; Smith, D.F.; Voellmy, R. Repression of heat shock transcription factor HSF1 activation by HSP90 (HSP90 complex) that forms a stress-sensitive complex with HSF1. Cell 1998, 94, 471–480. [Google Scholar] [CrossRef] [Green Version]
- Bandyopadhyay, U.; Kaushik, S.; Varticovski, L.; Cuervo, A.M. The chaperone-mediated autophagy receptor organizes in dynamic protein complexes at the lysosomal membrane. Mol. Cell Biol. 2008, 28, 5747–5763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Martinez-Vicente, M.; Krüger, U.; Kaushik, S.; Wong, E.; Mandelkow, E.M.; Cuervo, A.M.; Mandelkow, E. Tau fragmentation, aggregation and clearance: The dual role of lysosomal processing. Hum. Mol. Genet. 2009, 18, 4153–4170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, L.; Zhang, X.D. Role of chaperone-mediated autophagy in degrading Huntington’s disease-associated huntingtin protein. Acta Biochim. Biophys. Sin. 2014, 46, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Xilouri, M.; Brekk, O.R.; Landeck, N.; Pitychoutis, P.M.; Papasilekas, T.; Papadopoulou-Daifoti, Z.; Kirik, D.; Stefanis, L. Boosting chaperone-mediated autophagy in vivo mitigates α-synuclein-induced neurodegeneration. Brain 2013, 136, 2130–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, S.; Fang, M.; Ni, B.; Lu, D.; Martin, B.; Maudsley, S. Central role of the EGF receptor in neurometabolic aging. Int. J. Endocrinol. 2012, 2012, 739428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Chiang, H.C.; Wu, W.; Liang, B.; Xie, Z.; Yao, X.; Ma, W.; Du, S.; Zhong, Y. Epidermal growth factor receptor is a preferred target for treating amyloid-β-induced memory loss. Proc. Natl. Acad. Sci. USA 2012, 109, 16743–16748. [Google Scholar] [CrossRef] [Green Version]
- Wong, R.W.; Guillaud, L. The role of epidermal growth factor and its receptors in mammalian CNS. Cytokine Growth Factor Rev. 2004, 15, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Mazzucchelli, C.; Brambilla, R. Ras-related and MAPK signalling in neuronal plasticity and memory formation. Cell Mol. Life Sci. 2000, 57, 604–611. [Google Scholar] [CrossRef]
- Chiang, H.C.; Wang, L.; Xie, Z.; Yau, A.; Zhong, Y. PI3 kinase signaling is involved in Abeta-induced memory loss in Drosophila. Proc. Natl. Acad. Sci. USA 2010, 107, 7060–7065. [Google Scholar] [CrossRef] [Green Version]
- Almeida, C.G.; Takahashi, R.H.; Gouras, G.K. Beta-amyloid accumulation impairs multivesicular body sorting by inhibiting the ubiquitin-proteasome system. J. Neurosci. 2006, 26, 4277–4288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Repetto, E.; Yoon, I.S.; Zheng, H.; Kang, D.E. Presenilin 1 regulates epidermal growth factor receptor turnover and signaling in the endosomal-lysosomal pathway. J. Biol. Chem. 2007, 282, 31504–31516. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.W.; Wang, R.; Liu, Q.; Zhang, H.; Liao, F.F.; Xu, H. Presenilin/gamma-secretase-dependent processing of beta-amyloid precursor protein regulates EGF receptor expression. Proc. Natl. Acad. Sci. USA 2007, 104, 10613–10618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Keeffe, G.C.; Tyers, P.; Aarsland, D.; Dalley, J.W.; Barker, R.A.; Caldwell, M.A. Dopamine-induced proliferation of adult neural precursor cells in the mammalian subventricular zone is mediated through EGF. Proc. Natl. Acad. Sci. USA 2009, 106, 8754–8759. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Name | Gene Ontology Annotation | Degree (k) | c(k) | CN(k) | CB(k) | CC(k) | CE(k) |
|---|---|---|---|---|---|---|---|---|
| UBC | ubiquitin C | protease binding | 88 | 0.419801 | 45.125 | 0.025 | 0.84375 | 0.171251 |
| EGFR | epidermal growth factor receptor | identical protein binding and protein kinase activity | 72 | 0.475352 | 48.08333 | 0.013 | 0.75 | 0.148904 |
| APP | amyloid beta precursor protein | identical protein binding and enzyme binding | 66 | 0.454079 | 47.07576 | 0.012 | 0.72 | 0.133501 |
| CTNNB1 | catenin beta 1 | DNA-binding transcription factor activity and binding | 68 | 0.473661 | 47.98529 | 0.012 | 0.72973 | 0.140163 |
| NTRK1 | neurotrophic receptor tyrosine kinase 1 | protein homodimerization activity and protein kinase activity | 65 | 0.465865 | 47.30769 | 0.012 | 0.715232 | 0.132686 |
| FN1 | fibronectin 1 | heparin binding and protease binding | 63 | 0.461342 | 47.28571 | 0.011 | 0.705882 | 0.128571 |
| HSP90AA1 | heat shock protein 90kDa alpha family class A member 1 | identical protein binding | 67 | 0.483492 | 48.20896 | 0.010 | 0.724832 | 0.139365 |
| MDM2 | MDM2 proto-oncogene | identical protein binding and ligase activity | 59 | 0.456458 | 46.86441 | 0.010 | 0.687898 | 0.119216 |
| VCP | valosin-containing protein | signaling receptor binding | 61 | 0.477049 | 47.95082 | 0.010 | 0.696774 | 0.125994 |
| CTNNA1 | catenin alpha 1 | actin filament binding | 58 | 0.455535 | 47.2069 | 0.010 | 0.683544 | 0.117217 |
| GRB2 | growth factor receptor-bound protein 2 | protein kinase binding | 61 | 0.472678 | 47.77049 | 0.010 | 0.696774 | 0.125459 |
| Gene | Name | Gene Ontology Annotation | Degree (k) | c(k) | CN(k) | CB(k) | CC(k) | CE(k) |
|---|---|---|---|---|---|---|---|---|
| ANK2 | ankyrin 2, neuronal | protein kinase binding and structural constituent of cytoskeleton | 153 | 0.23787 | 164.797 | 9.84 × 10−4 | 0.54624 | 0.03467 |
| APAF1 | apoptotic peptidase activating factor 1 | identical protein binding and ADP binding | 131 | 0.23864 | 163.863 | 7.37 × 10−4 | 0.53922 | 0.02975 |
| BAG2 | BCL2 associated athanogene 2 | identical protein binding and chaperone binding | 137 | 0.25537 | 165.263 | 8.11 × 10−4 | 0.54121 | 0.03184 |
| CCL5 | C-C motif chemokine ligand 5 | protein homodimerization activity and chemokine activity | 126 | 0.36648 | 158.738 | 4.94 × 10−4 | 0.53528 | 0.0271 |
| CD4 | CD4 molecule | protein homodimerization activity and enzyme binding | 148 | 0.25896 | 162.304 | 0.00104 | 0.54321 | 0.03311 |
| CELF2 | CUGBP, Elav-like family member 2 | nucleic acid binding and RNA binding | 278 | 0.23663 | 163.838 | 0.00327 | 0.59259 | 0.06322 |
| CTNNA3 | catenin alpha 3 | structural molecule activity and beta–catenin binding | 205 | 0.24017 | 161.654 | 0.0022 | 0.56374 | 0.04577 |
| DNAJB1 | DnaJ heat shock protein family (Hsp40) member B1 | unfolded protein binding and ATPase binding | 125 | 0.25639 | 163.832 | 6.87 × 10−4 | 0.5379 | 0.02853 |
| EGFR | epidermal growth factor receptor | identical protein binding and protein kinase activity | 316 | 0.23984 | 165.997 | 0.004 | 0.609 | 0.07338 |
| FGF1 | fibroblast growth factor 1 | growth factor activity and Hsp70 protein binding | 153 | 0.2254 | 158.529 | 0.00113 | 0.54591 | 0.03315 |
| FYN | FYN proto-oncogene, Src family tyrosine kinase | transferase activity, transferring phosphorus-containing groups and protein tyrosine kinase activity | 231 | 0.26934 | 172.139 | 0.00204 | 0.57516 | 0.05493 |
| HDAC9 | histone deacetylase 9 | transcription factor binding and histone deacetylase binding | 122 | 0.27476 | 176.23 | 5.79 × 10−4 | 0.53528 | 0.02986 |
| HSF1 | heat shock transcription factor 1 | DNA-binding transcription factor activity and chromatin binding | 92 | 0.3022 | 166.761 | 2.95 × 10−4 | 0.52569 | 0.02156 |
| HSP90AA1 | heat shock protein 90kDa alpha family class A member 1 | identical protein binding | 293 | 0.27411 | 172.635 | 0.00305 | 0.59823 | 0.07174 |
| HSP90AB1 | heat shock protein 90kDa alpha family class B member 1 | protein kinase binding | 207 | 0.30772 | 176.865 | 0.0015 | 0.56519 | 0.05223 |
| HSPA1A | heat shock protein family A (Hsp70) member 1A | ubiquitin protein ligase binding | 108 | 0.35722 | 193.324 | 3.41 × 10−4 | 0.53108 | 0.02952 |
| IL34 | interleukin 34 | cytokine activity and macrophage colony-stimulating factor receptor binding | 32 | 0.22379 | 160.719 | 4.95 × 10−5 | 0.50372 | 0.00696 |
| JUN | Jun proto-oncogene, AP-1 transcription factor subunit | sequence-specific DNA binding | 298 | 0.23117 | 161.597 | 0.00383 | 0.59986 | 0.06713 |
| LCK | LCK proto-oncogene, Src family tyrosine kinase | identical protein binding and protein kinase activity | 150 | 0.30318 | 174.307 | 7.34 × 10−4 | 0.54422 | 0.03627 |
| LRP6 | LDL receptor related protein 6 | protein homodimerization activity and signaling receptor binding | 98 | 0.23312 | 159.214 | 7.21 × 10−4 | 0.52695 | 0.02125 |
| NF1 | neurofibromin 1 | binding and phosphatidylcholine binding | 119 | 0.2286 | 162.832 | 6.87 × 10−4 | 0.53495 | 0.02656 |
| NOS2 | nitric oxide synthase 2 | protein homodimerization activity and oxidoreductase activity | 107 | 0.23911 | 165.486 | 6.23 × 10−4 | 0.53012 | 0.02444 |
| RIN3 | Ras and Rab interactor 3 | GTPase activator activity and Rab guanyl–nucleotide exchange factor activity | 43 | 0.29236 | 158.86 | 7.03 × 10−5 | 0.50286 | 0.00939 |
| RPS6KB2 | ribosomal protein S6 kinase B2 | transferase activity, transferring phosphorus-containing groups and protein tyrosine kinase activity | 112 | 0.27622 | 163.482 | 4.95 × 10−4 | 0.52916 | 0.02593 |
| TXNIP | thioredoxin interacting protein | ubiquitin protein ligase binding and enzyme inhibitor activity | 166 | 0.25936 | 171.554 | 9.43 × 10−4 | 0.55103 | 0.03948 |
| UBE4A | ubiquitination factor E4A | ligase activity and ubiquitin–ubiquitin ligase activity | 188 | 0.23603 | 158.957 | 0.00151 | 0.55802 | 0.04172 |
| VLDLR | very low-density lipoprotein receptor | calcium ion binding | 139 | 0.23762 | 162.698 | 8.79 × 10−4 | 0.54087 | 0.03118 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lalwani, A.K.; Krishnan, K.; Bagabir, S.A.; Alkhanani, M.F.; Almalki, A.H.; Haque, S.; Sharma, S.K.; Singh, R.K.B.; Malik, M.Z. Network Theoretical Approach to Explore Factors Affecting Signal Propagation and Stability in Dementia’s Protein-Protein Interaction Network. Biomolecules 2022, 12, 451. https://doi.org/10.3390/biom12030451
Lalwani AK, Krishnan K, Bagabir SA, Alkhanani MF, Almalki AH, Haque S, Sharma SK, Singh RKB, Malik MZ. Network Theoretical Approach to Explore Factors Affecting Signal Propagation and Stability in Dementia’s Protein-Protein Interaction Network. Biomolecules. 2022; 12(3):451. https://doi.org/10.3390/biom12030451
Chicago/Turabian StyleLalwani, Amit Kumar, Kushagra Krishnan, Sali Abubaker Bagabir, Mustfa F. Alkhanani, Atiah H. Almalki, Shafiul Haque, Saurabh Kumar Sharma, R. K. Brojen Singh, and Md. Zubbair Malik. 2022. "Network Theoretical Approach to Explore Factors Affecting Signal Propagation and Stability in Dementia’s Protein-Protein Interaction Network" Biomolecules 12, no. 3: 451. https://doi.org/10.3390/biom12030451
APA StyleLalwani, A. K., Krishnan, K., Bagabir, S. A., Alkhanani, M. F., Almalki, A. H., Haque, S., Sharma, S. K., Singh, R. K. B., & Malik, M. Z. (2022). Network Theoretical Approach to Explore Factors Affecting Signal Propagation and Stability in Dementia’s Protein-Protein Interaction Network. Biomolecules, 12(3), 451. https://doi.org/10.3390/biom12030451