
Structure Elucidation and Anti-Tumor Activities of Trichothecenes from Endophytic Fungus Fusariumsporotrichioides

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Instruments, Chemicals, and Biochemicals
2.2. Strain Isolation and Cultivation
2.3. Extraction and Isolation
2.4. Cell Lines and Cell Culture
2.5. Cell Viability Assay
2.6. Cell Cycle Assay
2.7. Cell Apoptosis Analysis
2.8. Mitochondrial Membrane Potential Analysis
2.9. Measurement of ROS Generation
2.10. Western Blot Analysis
2.11. Statistical Analysis
3. Results
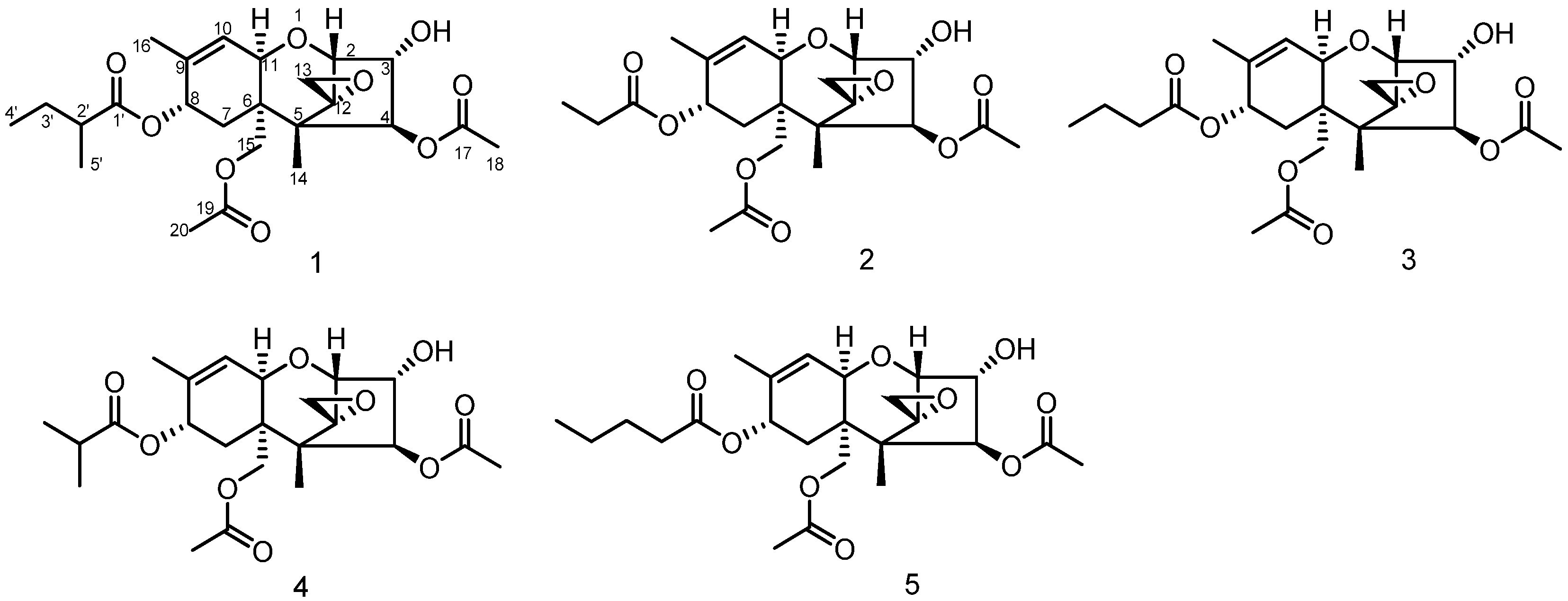
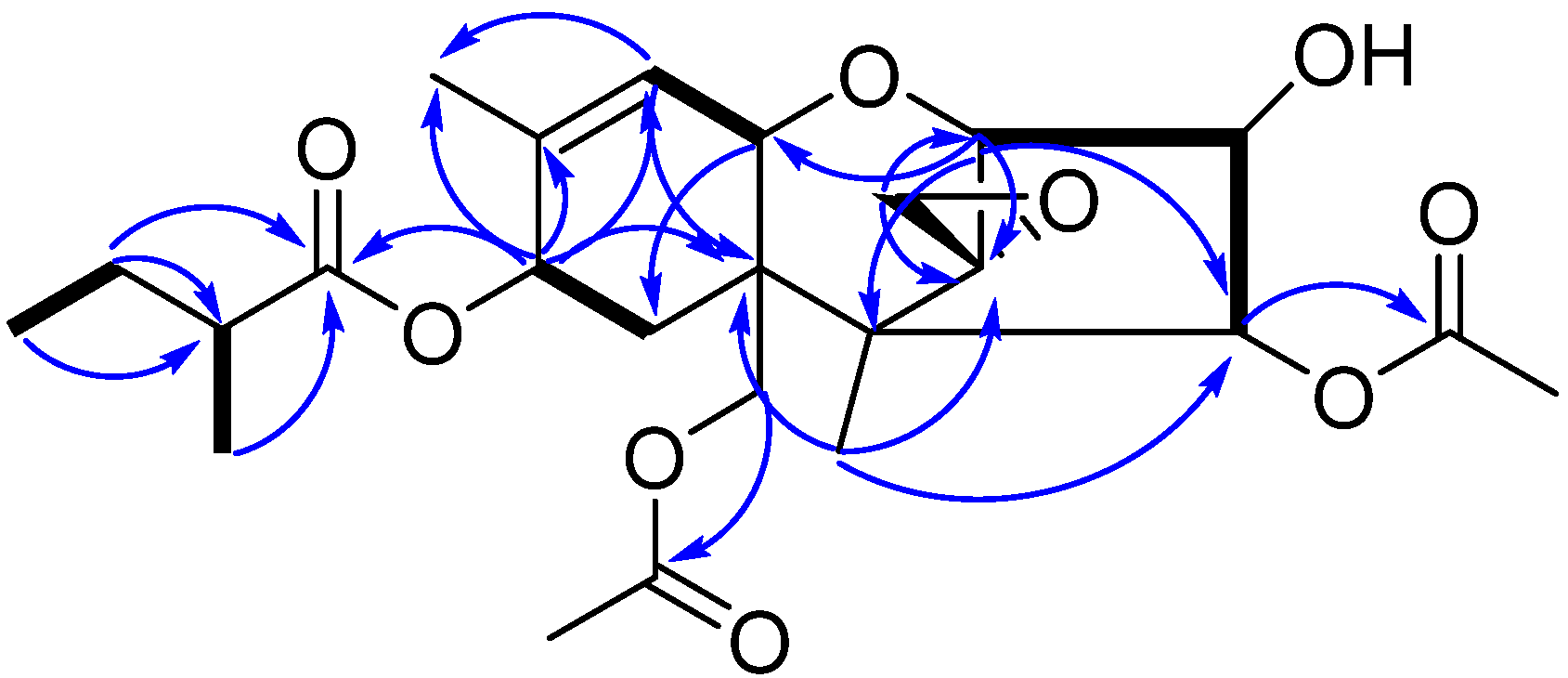
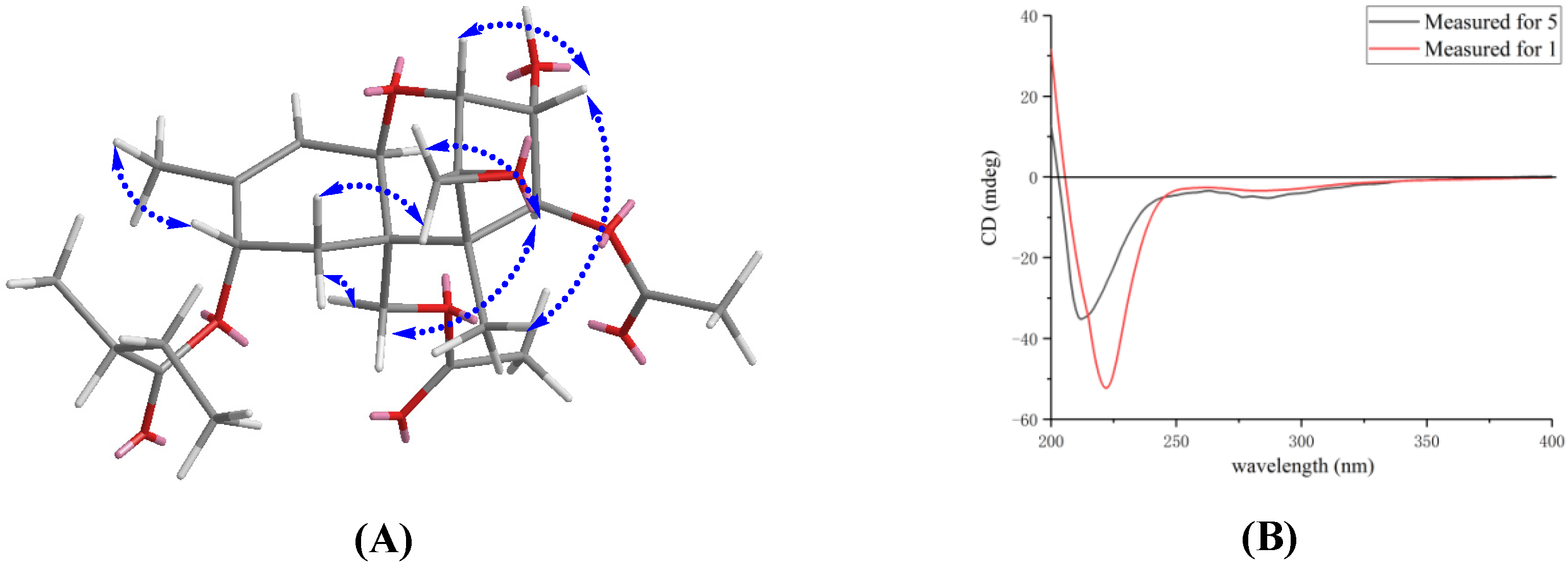
3.1. Structure Elucidation of Compound 1
3.2. Anti-Tumor Activities of Compounds 1–5
3.3. Effects of Compound 3 Inducing Cell Cycle Arrest in Huh-7 Cells
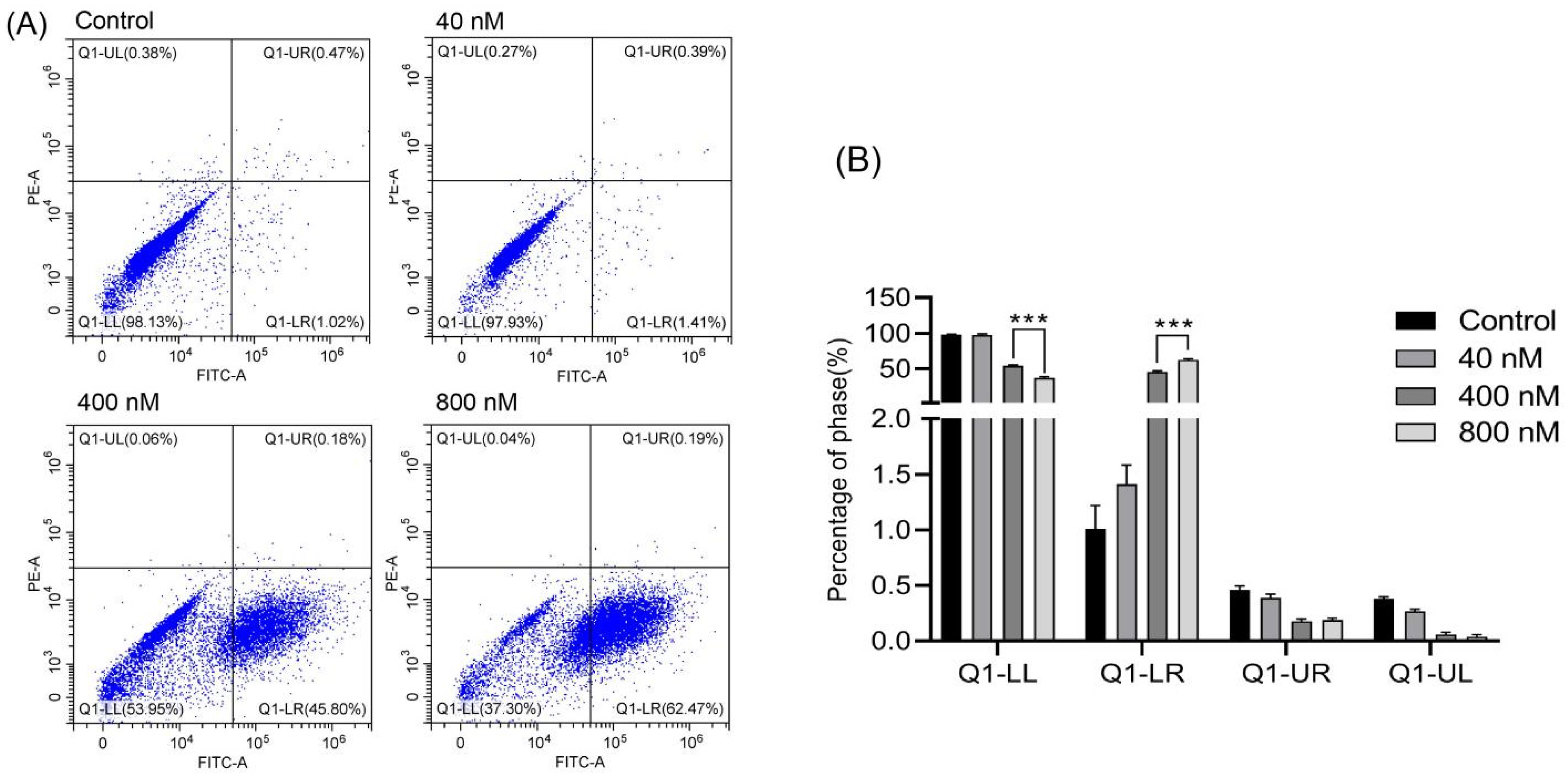
3.4. Effects of Compound 3 Inducing Apoptosis of Huh-7 Cells
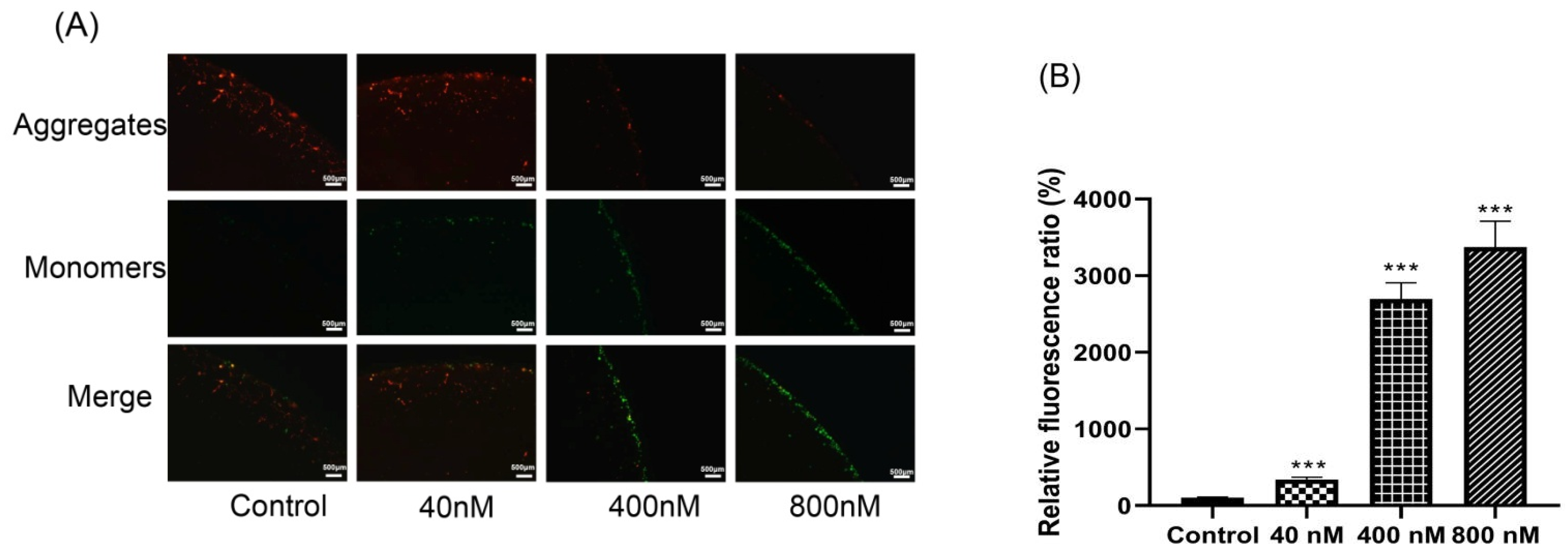
3.5. Effects of Compound 3 on Inducing Depolarization of Mitochondrial Membrane Potential and Generation of ROS
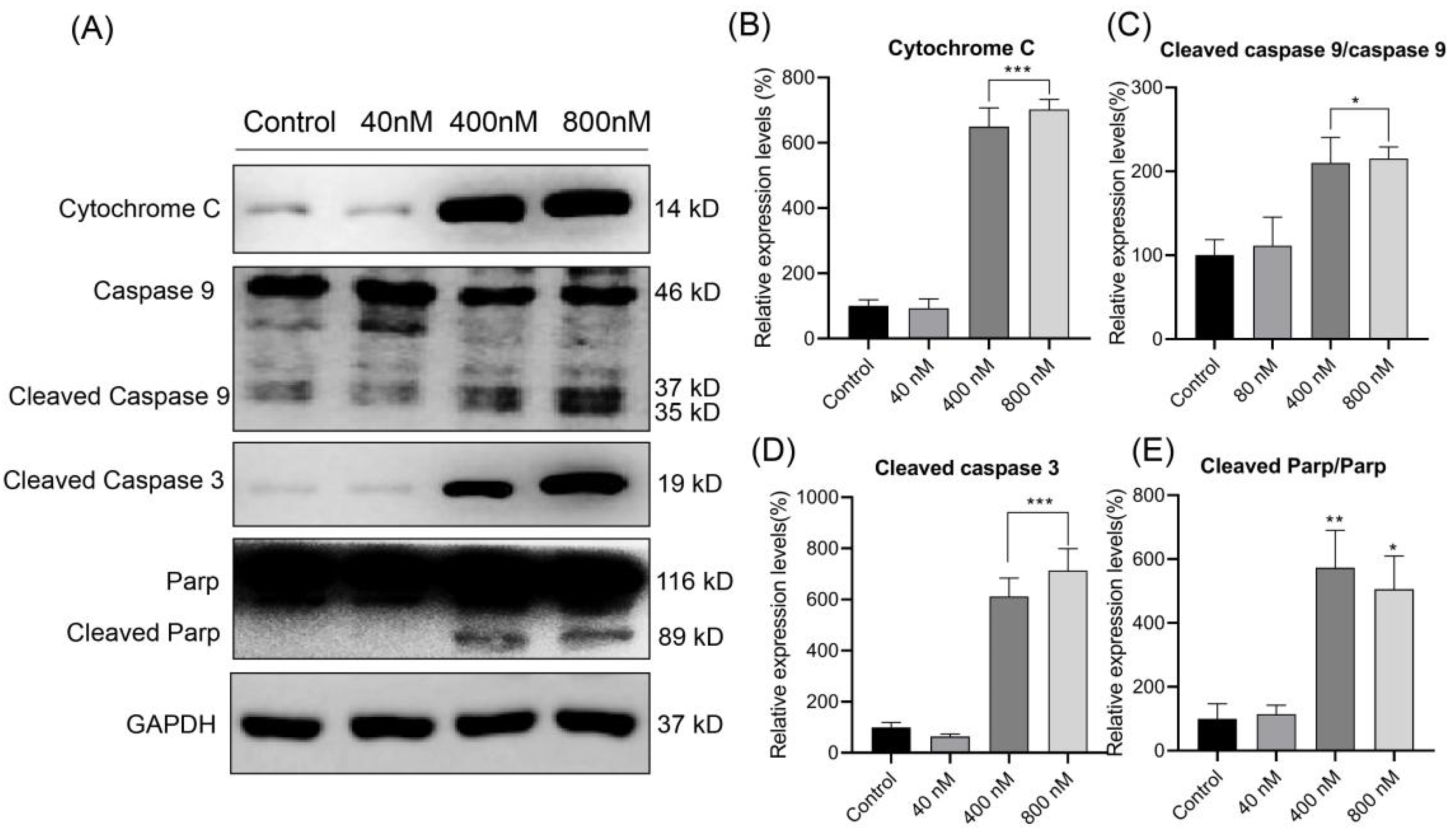
3.6. Effect of Compound 3 on the Expression of Apoptosis-Related Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Strobel, G.; Daisy, B.; Castillo, U.; Harper, J. Natural products from endophytic microorganisms. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar] [CrossRef]
- Daley, S.K.; Cordell, G.A. Biologically significant and recently isolated alkaloids from endophytic fungi. J. Nat. Prod. 2021, 84, 871–897. [Google Scholar] [CrossRef]
- Zhao, H.M.; Yang, A.P.; Liu, J.; Bao, S.; Peng, R.; Hu, Y.; Yuan, T.J.; Hou, S.R.; Xie, T.; Zhang, Q.C.; et al. Chartspiroton, a Tetracyclic Spiro-naphthoquinone Derivative from a medicinal plant endophytic Streptomyces. Org. Lett. 2020, 22, 3739–3743. [Google Scholar] [CrossRef]
- Gakuubi, M.M.; Munusamy, M.; Liang, Z.X.; Ng, S.B. Fungal endophytes: A promising frontier for discovery of novel bioactive compounds. J. Fungi 2021, 7, 786. [Google Scholar] [CrossRef]
- Ai, H.L.; Shi, B.B.; Li, W.; He, J.; Li, Z.H.; Feng, T.; Liu, J.K. Bipolarithizole A, an antifungal phenylthiazole-sativene merosesquiterpenoid from the potato endophytic fungus Bipolaris eleusines. Org. Chem. Front. 2022, 9, 1814–1819. [Google Scholar] [CrossRef]
- Yang, H.X.; Xu, X.; Chi, M.J.; Li, Z.H.; Feng, T.; Ai, H.L.; Liu, J.K. Structure and cytotoxicity of trichothecenes produced by the potato-associated fungus Trichothecium crotocinigenum. Bioorg. Chem. 2021, 111, 104874. [Google Scholar] [CrossRef]
- Shi, Z.Z.; Liu, X.H.; Li, X.N.; Li, N.Y. Antifungal and antimicroalgal trichothecene sesquiterpenes from the marine algicolous fungus Trichoderma brevicompactum A-DL-9-2. J. Agric. Food Chem. 2020, 68, 15440–15448. [Google Scholar] [CrossRef]
- McCormick, S.P.; Stanley, A.M.; Stover, N.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, J.; Ruan, H.L. Trichothecenes from an endophytic Fungus Alternaria sp. sb23. Planta Med. 2020, 86, 976–982. [Google Scholar] [CrossRef]
- Magdalena, P.S.; Beata, P. Trichothecenes in food and feed, relevance to human and animal health and methods of detection: A Systematic Review. Molecules 2021, 26, 454. [Google Scholar]
- Zhu, M.; Cen, Y.; Ye, W.; Li, S.; Zhang, W. Recent advances on macrocyclic trichothecenes, their bioactivities and biosynthetic pathway. Toxins 2020, 12, 417. [Google Scholar] [CrossRef]
- Mccormick, S.P.; Price, N.P.; Kurtzman, C.P. Glucosylation and other biotransformations of T-2 toxin by yeasts of the trichomonascus clade. Appl. Environ. Microb. 2012, 78, 8694–8702. [Google Scholar] [CrossRef] [Green Version]
- Greenhalgh, R.; Fielder, D.A.; Blackwell, B.A.; Miller, J.D.; Charland, J.P.; ApSimon, J.W. Some minor secondary metabolites of Fusarium sporotrichioides DAOM 165006. J. Agric. Food Chem. 1990, 38, 1978–1984. [Google Scholar] [CrossRef]
- Bekele, E.; Rottinghaus, A.A.; Rottinghaus, G.E. Two new trichothecenes from Fusarium sporotrichioides. J. Nat. Prod. 1991, 54, 1303–1308. [Google Scholar] [CrossRef]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in cereal grains—An Update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef] [Green Version]
- Foroud, N.A.; Eudes, F. Trichothecenes in cereal grains. Int. J. Mol. Sci. 2009, 10, 147–173. [Google Scholar] [CrossRef] [Green Version]
- Savard, M.E.; Blackwell, B.A.; Greenhalgh, R. An 1H nuclear magnetic resonance study of derivatives of 3-hydroxy-12,13-epoxytrichothec-9-enes. Can. J. Chem. 1987, 65, 2254–2262. [Google Scholar] [CrossRef]
- Yang, J.H.; Guo, W.B.; Wang, J.H.; Yang, X.L.; Zhang, Z.Q.; Zhao, Z.H. T-2 toxin-induced oxidative stress leads to imbalance of mitochondrial fission and fusion to activate cellular apoptosis in the human liver 7702 Cell Line. Toxins 2020, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Tang, M.; Pi, C.; Zheng, J.R.; Gao, S.X.; Chabanne, T.; Chauvin, R.; Cheng, W.Z.; Lin, H.J.; Xu, R.; et al. Novel ferrocene derivatives induce apoptosis through mitochondria-dependent and cell cycle arrest via PI3K/Akt/mTOR signaling pathway in T Cell acute lymphoblastic leukemia. Cancers 2021, 13, 4677. [Google Scholar] [CrossRef]
- Pucci, B.; Kasten, M.; Giordano, A. Cell cycle and apoptosis. Neoplasia 2000, 2, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, K.; Berneman, Z.N.; Van Bockstaele, D.R. Cell cycle and apoptosis. Cell Prolif. 2003, 36, 165–175. [Google Scholar] [CrossRef]
- Sancar, A.; Lindsey-Boltz, L.A.; Unsal-Kaçmaz, K.; Linn, S. Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu. Rev. Biochem. 2004, 73, 39–85. [Google Scholar] [CrossRef] [Green Version]
- Norbury, C.J.; Zhivotovsky, B. DNA damage-induced apoptosis. Oncogene 2004, 23, 2797–2808. [Google Scholar] [CrossRef] [Green Version]
- Zbakh, H.; Zubía, E.; Reyes, C.L.; Calderón-Montaño, J.M.; Motilva, V. Anticancer activities of meroterpenoids isolated from the brown Alga Cystoseira usneoides against the human colon cancer cells HT-29. Foods 2020, 9, 300. [Google Scholar] [CrossRef] [Green Version]
- El-Huneidi, W.; Shehab, N.G.; Bajbouj, K.; Vinod, A.; El-Serafi, A.; Shafarin, J.; Bou Malhab, L.J.; Abdel-Rahman, W.M.; Abu-Gharbieh, E. Micromeria fruticose induces cell cycle arrest and apoptosis in breast and colorectal cancer cells. Pharmaceuticals 2020, 13, 115. [Google Scholar] [CrossRef]
- Liu, T.; Zhu, W.; Yang, X.; Chen, L.; Yang, R.W.; Hua, Z.C.; Li, G.X. Detection of apoptosis based on the interaction between annexin V and phosphatidylserine. Anal. Chem. 2009, 81, 2410–2413. [Google Scholar] [CrossRef]
- Kupcho, K.; Shultz, J.; Hurst, R.; Hartnett, J.; Zhou, W.H.; Machleidt, T.; Grailer, J.; Worzella, T.; Riss, T.; Lazar, D.; et al. A real-time, bioluminescent annexin V assay for the assessment of apoptosis. Apoptosis 2019, 24, 184–197. [Google Scholar] [CrossRef] [Green Version]
- Perry, S.W.; Norman, J.P.; Barbieri, J.; Brown, E.B.; Gelbard, H.A. Mitochondrial membrane potential probes and the proton gradient: A practical usage guide. BioTechniques 2011, 50, 98–115. [Google Scholar] [CrossRef]
- Horbay, R.; Bilyy, R. Mitochondrial dynamics during cell cycling. Apoptosis 2016, 21, 1327–1335. [Google Scholar] [CrossRef]
- Bock, F.J.; Tait, S. Mitochondria as multifaceted regulators of cell death. Nat. Rev. Mol. Cell Biol. 2020, 21, 85–100. [Google Scholar] [CrossRef]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (deltapsi(m)) in apoptosis; an update. Apoptosis 2003, 8, 115–128. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [Green Version]
- Tajeddine, N. How do reactive oxygen species and calcium trigger mitochondrial membrane permeabilisation. Biochim. Biophys. Acta 2016, 1860, 1079–1088. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhong, J.; Bi, Y.; Liu, Y.; Liu, Y.; Guo, J.; Pan, L.; Tan, Y.; Yu, X. Gambogenic acid induces Noxa-mediated apoptosis in colorectal cancer through ROS-dependent activation of IRE1α/JNK. Phytomedicine 2020, 78, 153306. [Google Scholar] [CrossRef]
- Kiraz, Y.; Adan, A.; Kartal Yandim, M.; Baran, Y. Major apoptotic mechanisms and genes involved in apoptosis. Tumour Biol. 2016, 37, 8471–8486. [Google Scholar] [CrossRef] [Green Version]
- Bertheloot, D.; Latz, E.; Franklin, B.S. Necroptosis, pyroptosis and apoptosis: An intricate game of cell death. Cell Mol. Immunol. 2021, 18, 1106–1121. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A. Apoptosis: A target for anticancer therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [Green Version]
- Kesavardhana, S.; Malireddi, R.; Kanneganti, T.D. Caspases in cell death, inflammation, and pyroptosis. Annu. Rev. Immunol. 2020, 38, 567–595. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y. Mechanisms of caspase activation and inhibition during apoptosis. Mol. Cell. 2002, 9, 459–470. [Google Scholar] [CrossRef]
- Nagata, S. Apoptosis and clearance of apoptotic cells. Annu. Rev. Immunol. 2018, 36, 489–517. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | δH J (Hz) | δC | No. | δH J (Hz) | δC |
|---|---|---|---|---|---|
| 2 | 3.54, d, 4.9 | 80.4, CH | 14 | 0.72, s | 6.7, CH3 |
| 3 | 4.22, dd, 4.9, 3.1 | 77.8, CH | 15 | 4.34, d, 12.5 | 65.9, CH2 |
| 4 | 5.78, d, 3.1 | 83.9, CH | 4.12, d, 12.5 | ||
| 5 | 49.8, C | 16 | 1.74, s | 20.2, CH3 | |
| 6 | 44.0, C | 17 | 172.3, C=O | ||
| 7 | 2.43, dd, 15.3, 6.0 | 29.0, CH2 | 18 | 2.08, s | 20.6, CH3 |
| 1.85, d, 15.3 | 19 | 172.1, C=O | |||
| 8 | 5.34, d, 6.0 | 69.1, CH | 20 | 2.05, s | 20.9, CH3 |
| 9 | 137.1, C | 1’ | 177.3, C=O | ||
| 10 | 5.80, d, 5.8 | 124.8, CH | 2’ | 2.33, m | 42.3, CH |
| 11 | 4.41, d, 5.8 | 68.2, CH | 3’ | 1.65, dq, 13.6, 7.4 | 27.6, CH2 |
| 12 | 65.3, C | 1.50, dq, 13.6, 7.4 | |||
| 13 | 3.02, d, 3.9 | 47.7, CH2 | 4’ | 0.92, t, 7.4 | 11.7, CH3 |
| 2.85, d, 3.9 | 5’ | 1.14, d, 7.0 | 16.5, CH3 |
| Cell Lines | IC50 (nM) | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | T-2 Toxin | |
| Huh-7 | 2202.7 | 890.7 | 265.9 | 479.7 | 676.4 | 357.1 |
| MRMT-1 | 1739.0 | 679.8 | 279.4 | 462.4 | 577.4 | 122.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-J.; Ma, N.; Lu, Y.-F.; Dai, S.-Y.; Song, X.; Li, C.; Sun, Y.; Pei, Y.-H. Structure Elucidation and Anti-Tumor Activities of Trichothecenes from Endophytic Fungus Fusariumsporotrichioides. Biomolecules 2022, 12, 778. https://doi.org/10.3390/biom12060778
Wang Y-J, Ma N, Lu Y-F, Dai S-Y, Song X, Li C, Sun Y, Pei Y-H. Structure Elucidation and Anti-Tumor Activities of Trichothecenes from Endophytic Fungus Fusariumsporotrichioides. Biomolecules. 2022; 12(6):778. https://doi.org/10.3390/biom12060778
Chicago/Turabian StyleWang, Ya-Jing, Nan Ma, Yong-Fu Lu, Si-Yang Dai, Xue Song, Chang Li, Yi Sun, and Yue-Hu Pei. 2022. "Structure Elucidation and Anti-Tumor Activities of Trichothecenes from Endophytic Fungus Fusariumsporotrichioides" Biomolecules 12, no. 6: 778. https://doi.org/10.3390/biom12060778