
Molecular Profiling of Tissue Samples with Chronic Rejection from Patients with Chronic Lung Allograft Dysfunction: A Pilot Study in Cystic Fibrosis Patients

 ,
,  , , ,
, , ,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Population
2.2. Sample Collection
2.3. RNA Extraction
2.4. RNA Sequencing and Differential Gene Expression Analysis
2.5. Ingenuity Pathway Analysis (IPA)
2.6. Quantitative Real-Time (qRT-PCR) Analysis
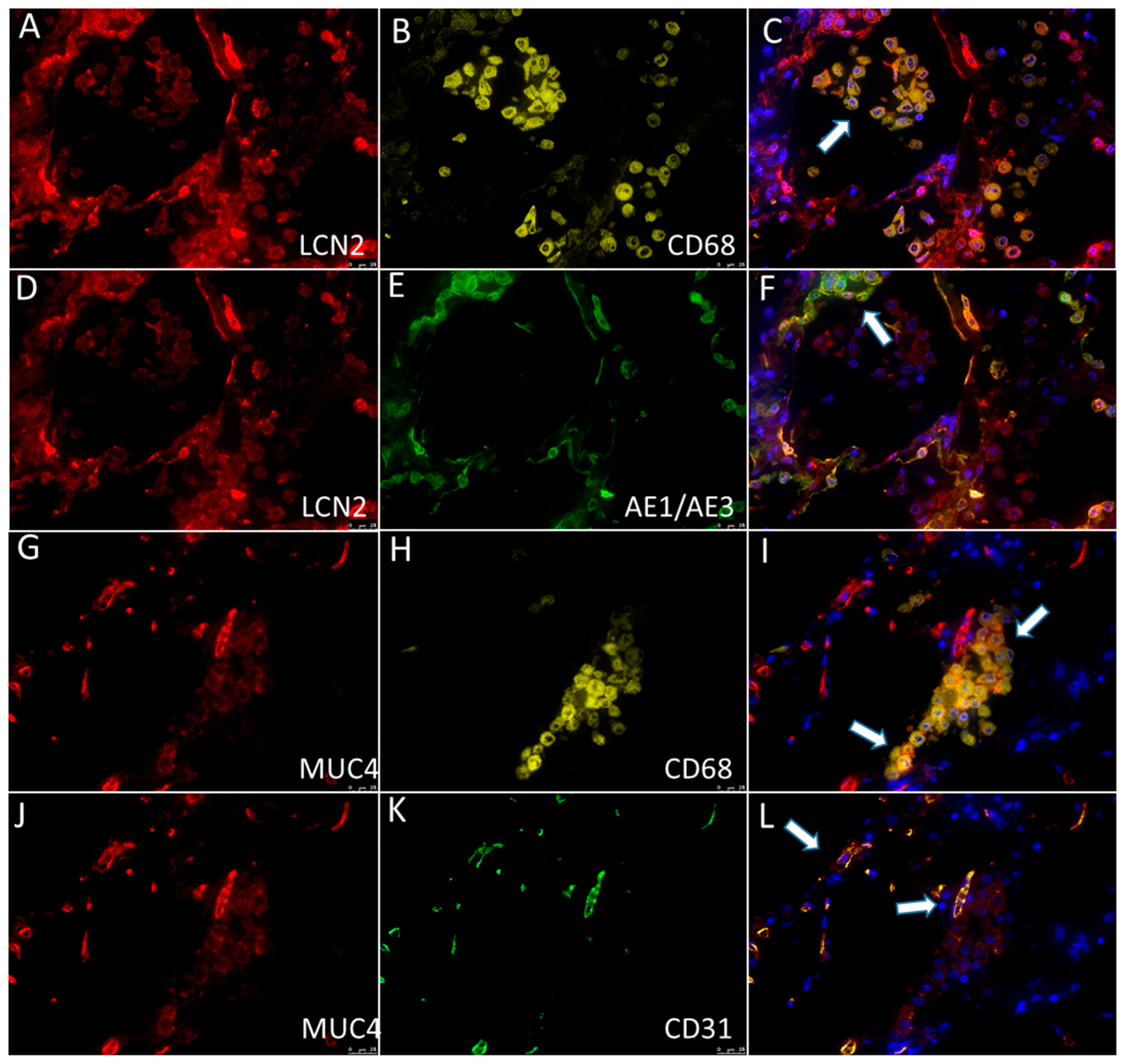
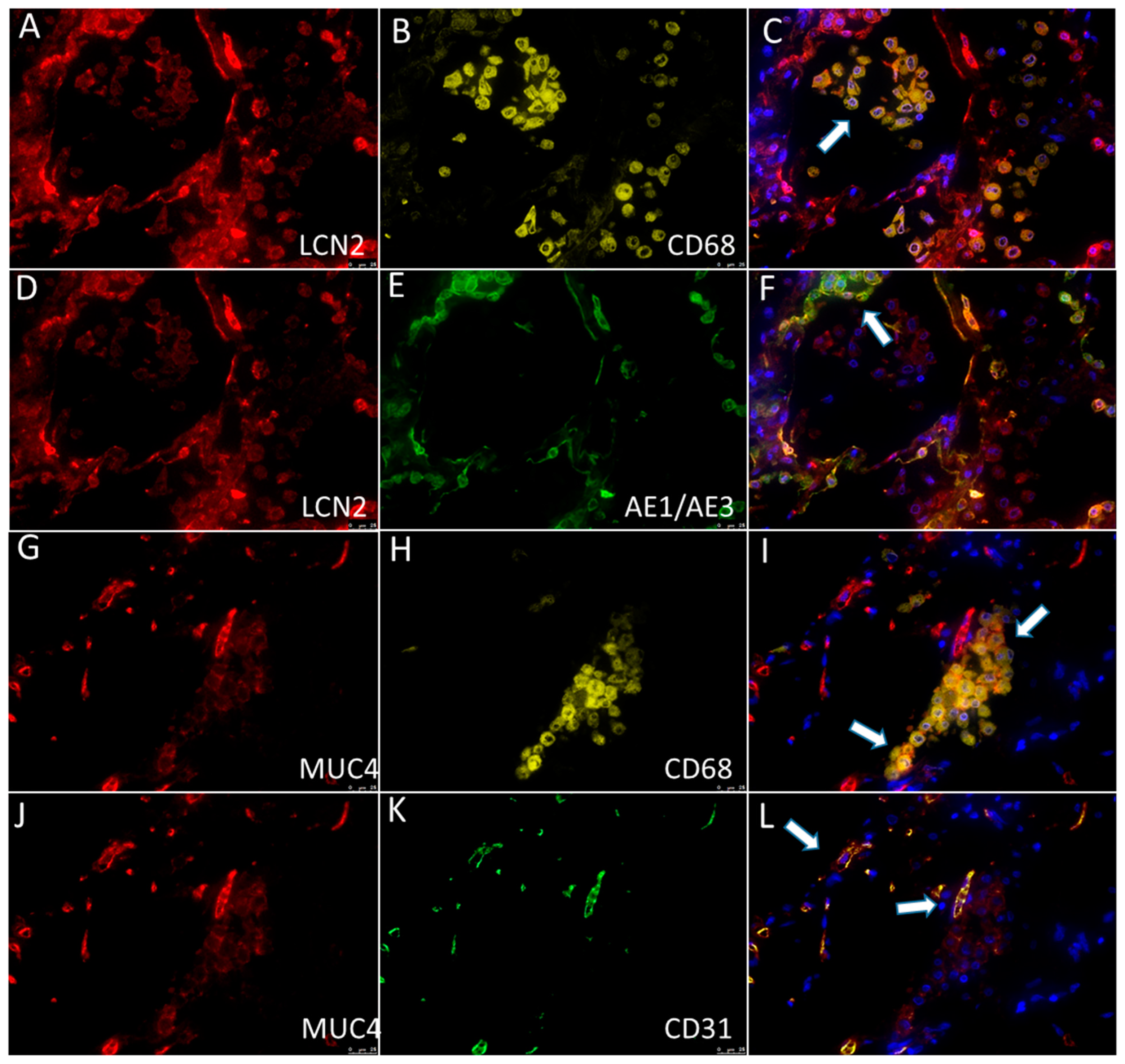
2.7. Immunohistochemistry and Multiplex Immunofluorescence
3. Results
3.1. Comprehensive Profiling of Biopsies Reveals Specific Alterations
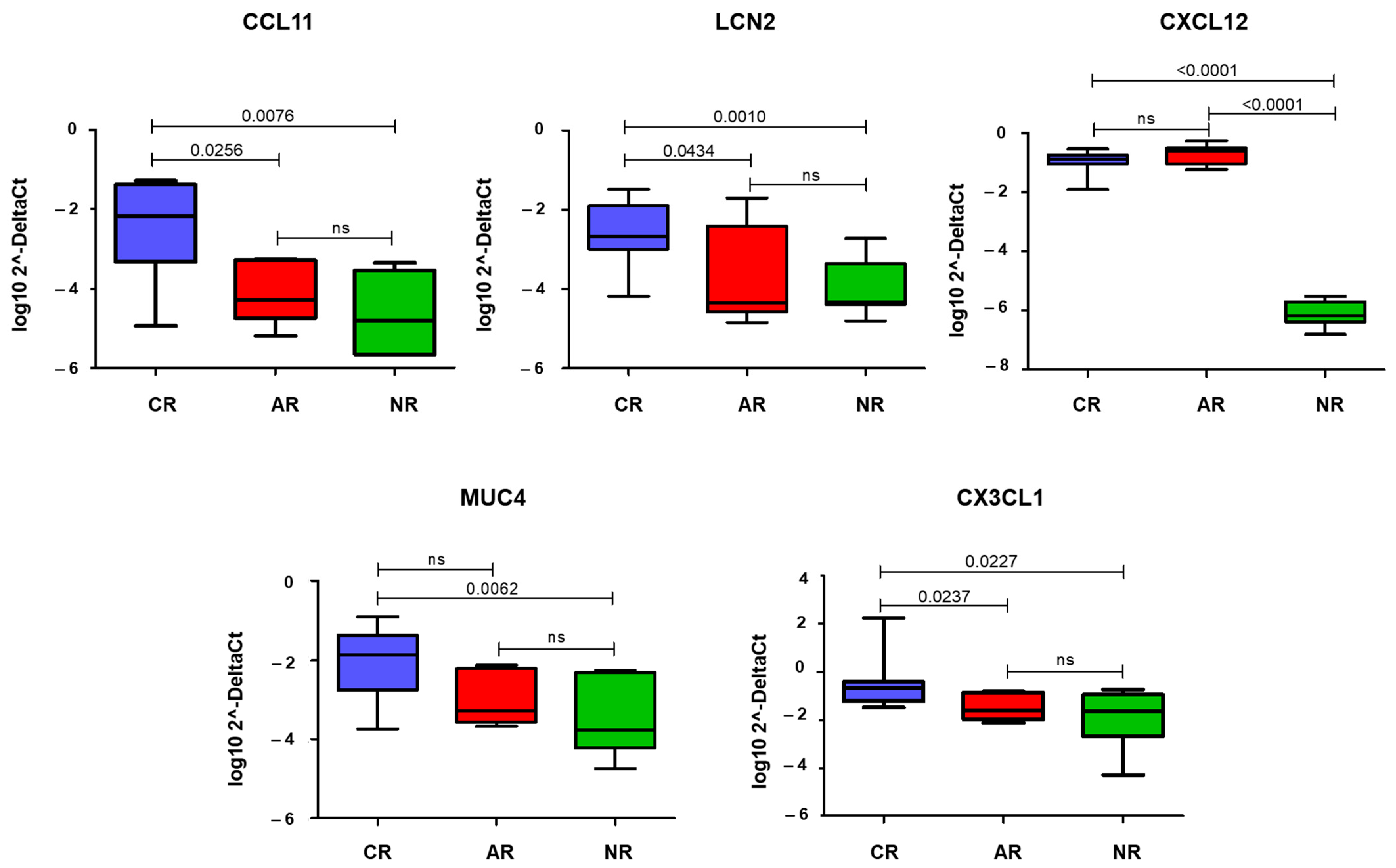
3.2. qRT-PCR and Immunohistochemistry
3.3. Finding of Longitudinal Cases
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation List
References
- Khush, K.K.; Cherikh, W.S.; Chambers, D.C.; Harhay, M.O.; Hayes, D.; Jr Hsich, E.; Meiser, B.; Potena, L.; Robinson, A.; Rossano, J.W.; et al. The International Thoracic Organ Transplant Registry of the International Society for Heart and Lung Transplantation: Thirty-sixth adult lung and heart-lung transplantation Report-2019; Focus theme: Donor and recipient size match. J. Heart Lung Trans. 2019, 38, 1042–1055. [Google Scholar] [CrossRef] [PubMed]
- Verleden, G.M.; Glanville, A.R.; Lease, E.D.; Fisher, A.J.; Calabrese, F.; Corris, P.A.; Ensor, C.R.; Gottlieb, J.; Hachem, R.R.; Lama, V.; et al. Chronic lung allograft dysfunction: Definition, diagnostic criteria, and approaches to treatment―A consensus report from the Pulmonary Council of the ISHLT. J. Heart Lung Trans. 2019, 38, 493–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotecha, S.; Paraskeva, M.A.; Levin, K.; Snell, G.I. An update on chronic lung allograft dysfunction. Ann. Trans. Med. 2020, 8, 417. [Google Scholar] [CrossRef] [PubMed]
- Glanville, A.R.; Verleden, G.M.; Todd, J.L.; Benden, C.; Calabrese, F.; Gottlieb, J.; Hachem, R.R.; Levine, D.; Meloni, F.; Palmer, S.M.; et al. Chronic lung allograft dysfunction: Definition and update of restrictive allograft syndrome―A consensus report from the Pulmonary Council of the ISHLT. J. Heart Lung Trans. 2019, 38, 483–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, P.M.; Aboyoun, C.L.; Chhajed, P.N.; Malouf, M.A.; Plit, M.L.; Rainer, S.P.; Glanville, A.R. Association of minimal rejection in lung transplant recipients with obliterative bronchiolitis. Am. J. Res. Crit. Care Med. 2004, 170, 1022–1026. [Google Scholar] [CrossRef] [PubMed]
- Girnita, A.L.; Duquesnoy, R.; Yousem, S.A.; Iacono, A.T.; Corcoran, T.E.; Buzoianu, M.; Johnson, B.; Spichty, K.J.; Dauber, J.H.; Burckart, G.; et al. HLA-specific antibodies are risk factors for lymphocytic bronchiolitis and chronic lung allograft dysfunction. Am. J. Trans. 2005, 5, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Jonigk, D.; Izykowski, N.; Rische, J.; Braubach, P.; Kühnel, M.; Warnecke, G.; Lippmann, T.; Kreipe, H.; Haverich, A.; Welte, T.; et al. Molecular profiling in lung biopsies of human pulmonary allografts to predict chronic lung allograft dysfunction. Am. J. Pathol. 2015, 185, 3178–3188. [Google Scholar] [CrossRef] [PubMed]
- Halloran, K.M.; Parkes, M.D.; Chang, J.; Timofte, I.L.; Snell, G.I.; Westall, G.P.; Hachem, R.; Kreisel, D.; Trulock, E.; Roux, A.; et al. Molecular assessment of rejection and injury in lung transplant biopsies. J. Heart Lung Trans. 2019, 38, 504–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacreas, A.; Yang, J.; Vanaudenaerde, B.M.; Sigdel, T.K.; Liberto, J.M.; Damm, I.; Verleden, G.M.; Vos, R.; Verleden, S.E.; Sarwal, M.M. The common rejection module in chronic rejection post lung transplantation. PLoS ONE 2018, 13, e0205107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkes, M.D.; Halloran, K.; Hirji, A.; Pon, S.; Weinkauf, J.; Timofte, I.L.; Snell, G.I.; Westall, G.P.; Havlin, J.; Lischke, R.; et al. Transcripts associated with chronic lung allograft dysfunction in transbronchial biopsies of lung transplants. Am. J. Trans. 2022, 22, 1054–1072. [Google Scholar] [CrossRef] [PubMed]
- Greenland, J.R.; Wang, P.; Brotman, J.J.; Ahuja, R.; Chong, T.A.; Kleinhenz, M.E.; Leard, L.E.; Golden, J.A.; Hays, S.R.; Kukreja, J.; et al. Gene signatures common to allograft rejection are associated with lymphocytic bronchitis. Clin. Trans. 2019, 33, e13515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugger, D.T.; Fung, M.; Hays, S.R.; Singer, J.P.; Kleinhenz, M.E.; Leard, L.E.; Golden, J.A.; Shah, R.J.; Lee, J.S.; Deiter, F.; et al. Chronic lung allograft dysfunction small airways reveal a lymphocytic inflammation gene signature. Am. J. Trans. 2021, 21, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Lehto, J.T.; Koskinen, P.K.; Anttila, V.J.; Lautenschlager, I.; Lemström, K.; Sipponen, J.; Tukiainen, P.; Halme, M. Bronchoscopy in the diagnosis and surveillance of respiratory infections in lung and heart-lung transplant recipients. Trans. Int. 2005, 18, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 20 May 2022).
- Kechin, A.; Boyarskikh, U.; Kel, A.; Filipenko, M. cutPrimers: A New Tool for Accurate Cutting of Primers from Reads of Targeted Next Generation Sequencing. J. Comput. Biol. 2017, 24, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.gencodegenes.org/ (accessed on 20 May 2022).
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general-purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: Rsubread/Subread%20Users%20Guide.%20Rsubread%20v2.6.2/Subread%20v2.0.2. (accessed on 20 May 2022).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Geno. Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soneson, C.; Delorenzi, M. A comparison of methods for differential expression analysis of RNA-seq data. BMC Bioinform. 2013, 14, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ENCODE Guidelines and Best Practices for RNA-Seq. Available online: https://www.encodeproject.org/documents/cede0cbe-d324-4ce7-ace4-f0c3eddf5972/@@download/attachment/ENCODE%20Best%20Practices%20for%20RNA_v2.pdf (accessed on 20 May 2022).
- Cowland, J.B.; Borregaard, N. Molecular characterization and pattern of tissue expression of the gene for neutrophil gelatinase-associated lipocalin from humans. Genomics 1997, 45, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Veraar, C.; Kliman, J.; Benazzo, A.; Oberndorfer, F.; Laggner, M.; Hacker, P.; Raunegger, T.; Janik, S.; Jaksch, P.; Klepetko, W.; et al. Potential novel biomarkers for chronic lung allograft dysfunction and azithromycin responsive allograft dysfunction. Sci. Rep. 2021, 11, 6799. [Google Scholar] [CrossRef] [PubMed]
- Milara, J.; Ballester, B.; Safont, M.J.; Artigues, E.; Escrivá, J.; Morcillo, E.; Cortijo, J. MUC4 is overexpressed in idiopathic pulmonary fibrosis and collaborates with transforming growth factor β inducing fibrotic responses. Mucosal. Immunol. 2021, 14, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Perez, A.; Yasin, M.; Soto, P.; Rong, M.; Theodoropoulos, G.; Carothers Carraway, C.A.; Carraway, K.L. Presence of MUC4 in human milk and at the luminal surfaces of blood vessels. J. Cell Physiol. 2005, 204, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Carraway, C.A.; Carraway, K.L. Muc4 expression during blood vessel formation in damaged rat cornea. Curr. Eye Res. 2006, 31, 1011–1014. [Google Scholar] [CrossRef] [PubMed]
- Xu-Dubois, Y.C.; Peltier, J.; Brocheriou, I.; Suberbielle-Boissel, C.; Djamali, A.; Reese, S.; Mooney, N.; Keuylian, Z.; Lion, J.; Ouali, N.; et al. Markers of Endothelial-to-Mesenchymal Transition: Evidence for Antibody-Endothelium Interaction during Antibody-Mediated Rejection in Kidney Recipients. J. Am. Soc. Nephrol. 2016, 27, 324–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Sharma, N.; Krishn, S.R.; Lakshmanan, I.; Rachagani, S.; Baine, M.J.; Smith, L.M.; Lele, S.M.; Sasson, A.R.; Guha, S.; et al. MUC4-Mediated Regulation of Acute Phase Protein Lipocalin 2 through HER2/AKT/NF-κB Signaling in Pancreatic Cancer. Clin. Cancer Res. 2014, 20, 688–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Niu, Y.; Yu, L.; Lv, W.; Xu, H.; Abuduwufuer, A.; Cao, J.; Hu, J. The role of epithelial-mesenchymal transition in the post-lung transplantation bronchiolitis obliterans. J. Cardiothorac. Surg. 2017, 12, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: https://string-db.org/ (accessed on 20 May 2022).
- Sallusto, F.; Mackay, C.R.; Lanzavecchia, A. The Role of Chemokine Receptors in Primary, Effector, and Memory Immune Responses. Annu. Rev. Immunol. 2000, 18, 593–620. [Google Scholar] [CrossRef] [PubMed]
- Nelson, P.J.; Krensky, A.M. Chemokines, Chemokine Receptors, and Allograft Rejection. Immunity 2001, 14, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-Sánchez, G.S.; Pita-Grisanti, V.; Quiñones-Díaz, B.; Gumpper, K.; Cruz-Monserrate, Z.; Vivas-Mejía, P.E. Biological Functions and Therapeutic Potential of Lipocalin 2 in Cancer. Int. J. Mol. Sci. 2020, 21, 4365. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Population (18 Patients) | |
|---|---|
| Age (years) [mean ± SD] | 26.1 ± 10.1 |
| Sex (males:females) | 6:12 |
| BMI (kg/m2) | 19.0 ± 3.0 |
| Colonization at the time of transplantation (yes:no) | 14:4 |
| FEV1 (L) | 1.0 ± 0.3 |
| Donor age (years) [mean ± SD] | 32.0 ± 12.8 |
| Donor sex (males:females) | 9:9 |
| Mean ischemic time (min) [mean ± SD] | 326.7 ± 89.9 |
| CMV status [N (%)] | |
| D+/− R+ | 14 (78%) |
| D+ R− | 0 (0%) |
| D− R− | 4 (22%) |
| CLAD [N (%)] | 12 (67%) |
| Status (dead:alive) | 6:12 |
| Survival (years) [mean ± SD] | 6.4 ± 3.2 |
| Comparison | Total DEGs | Down-Regulated Genes | Up-Regulated Genes |
|---|---|---|---|
| D vs. NR | 15,444 | 11,680 | 3764 |
| D vs. AR | 18,452 | 13,808 | 4644 |
| D vs. CR | 16,643 | 11,198 | 5445 |
| CR vs. NR | 114 | 37 | 77 |
| CR vs. AR | 499 | 275 | 224 |
| AR vs. NR | 94 | 28 | 66 |
| Gene Symbol | Gene Description | Relative Expression | ||
|---|---|---|---|---|
| Log2 Fold Change | p-Value | padj | ||
| CCL11 | C-C Motif Chemokine Ligand 11 | 3.2 | 8.6 × 10−8 | 4.8 × 10−5 |
| LCN2 | Lipocalin 2 | 2.7 | 1.0 × 10−4 | 8.0 × 10−3 |
| MT-CO2 | Mitochondrially Encoded Cytochrome C Oxidase II | 2.8 | 7.3 × 10−4 | 2.9 × 10−2 |
| SCARA-3 | Scavenger Receptor Class A Member 3 | 1.3 | 4.6 × 10−6 | 9.2 × 10−4 |
| CX3CL1 | C-X3-C Motif Chemokine Ligand 1 | 1.5 | 8.0 × 10−5 | 6.7 × 10−3 |
| CXCL12 | C-X-C Motif Chemokine Ligand 12 | 1.2 | 1.1 ×10−5 | 1.6 × 10−3 |
| MUC4 | Mucin 4 | 2.7 | 7.4 × 10−6 | 1.3 × 10−3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lunardi, F.; Abbrescia, D.I.; Vedovelli, L.; Pezzuto, F.; Fortarezza, F.; Comacchio, G.M.; Guzzardo, V.; Ferrigno, P.; Loy, M.; Giraudo, C.; et al. Molecular Profiling of Tissue Samples with Chronic Rejection from Patients with Chronic Lung Allograft Dysfunction: A Pilot Study in Cystic Fibrosis Patients. Biomolecules 2023, 13, 97. https://doi.org/10.3390/biom13010097
Lunardi F, Abbrescia DI, Vedovelli L, Pezzuto F, Fortarezza F, Comacchio GM, Guzzardo V, Ferrigno P, Loy M, Giraudo C, et al. Molecular Profiling of Tissue Samples with Chronic Rejection from Patients with Chronic Lung Allograft Dysfunction: A Pilot Study in Cystic Fibrosis Patients. Biomolecules. 2023; 13(1):97. https://doi.org/10.3390/biom13010097
Chicago/Turabian StyleLunardi, Francesca, Daniela Isabel Abbrescia, Luca Vedovelli, Federica Pezzuto, Francesco Fortarezza, Giovanni Maria Comacchio, Vincenza Guzzardo, Pia Ferrigno, Monica Loy, Chiara Giraudo, and et al. 2023. "Molecular Profiling of Tissue Samples with Chronic Rejection from Patients with Chronic Lung Allograft Dysfunction: A Pilot Study in Cystic Fibrosis Patients" Biomolecules 13, no. 1: 97. https://doi.org/10.3390/biom13010097
APA StyleLunardi, F., Abbrescia, D. I., Vedovelli, L., Pezzuto, F., Fortarezza, F., Comacchio, G. M., Guzzardo, V., Ferrigno, P., Loy, M., Giraudo, C., Fraia, A. S., Faccioli, E., Braccioni, F., Cozzi, E., Gregori, D., Verleden, G. M., Calabrese, F., Schena, F. P., & Rea, F. (2023). Molecular Profiling of Tissue Samples with Chronic Rejection from Patients with Chronic Lung Allograft Dysfunction: A Pilot Study in Cystic Fibrosis Patients. Biomolecules, 13(1), 97. https://doi.org/10.3390/biom13010097