

Fungal Laccases: Fundamentals, Engineering and Classification Update


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
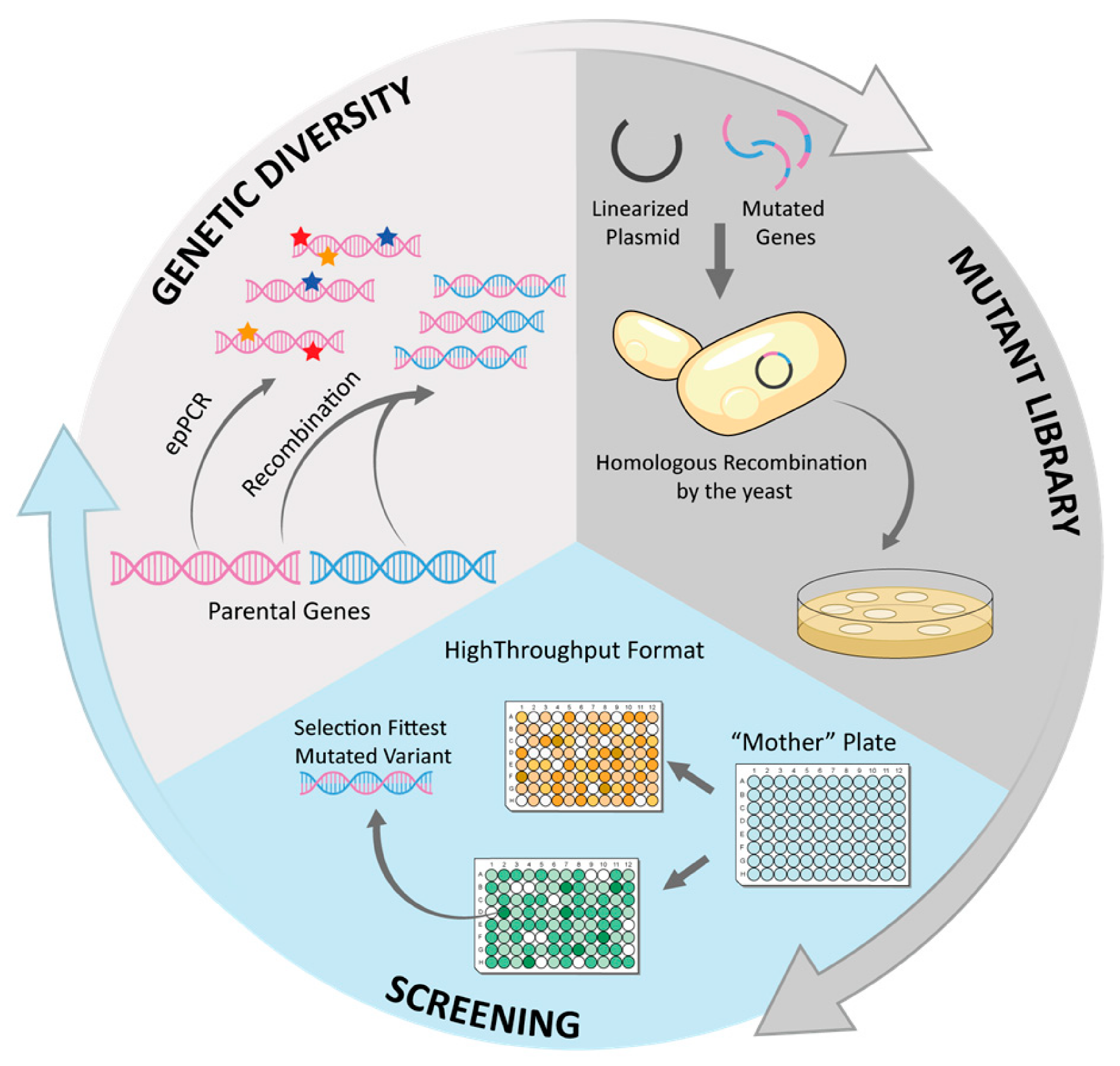{kind=link}
{kind=link}
Abstract
1. Laccases: General Aspects
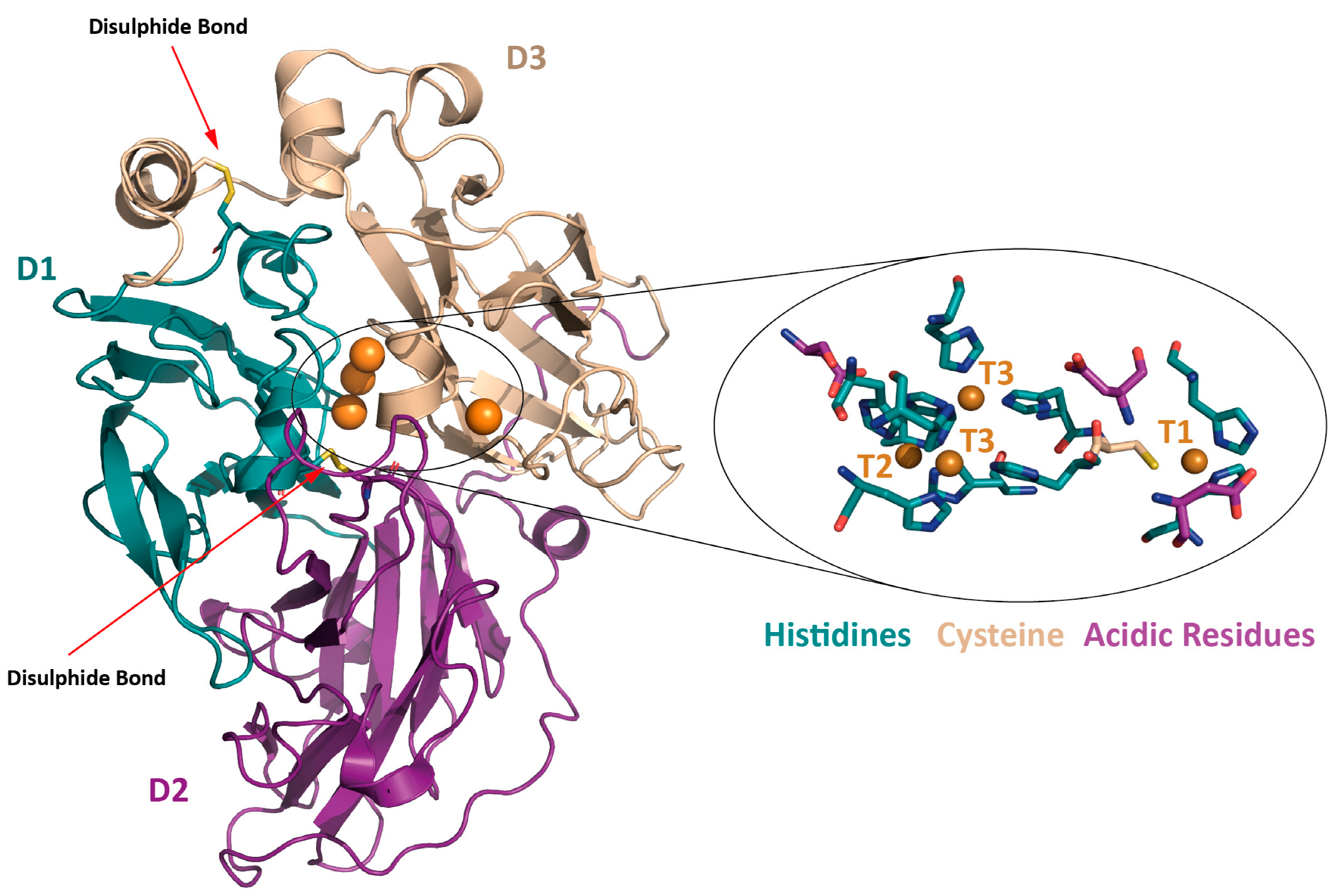
1.1. Structure
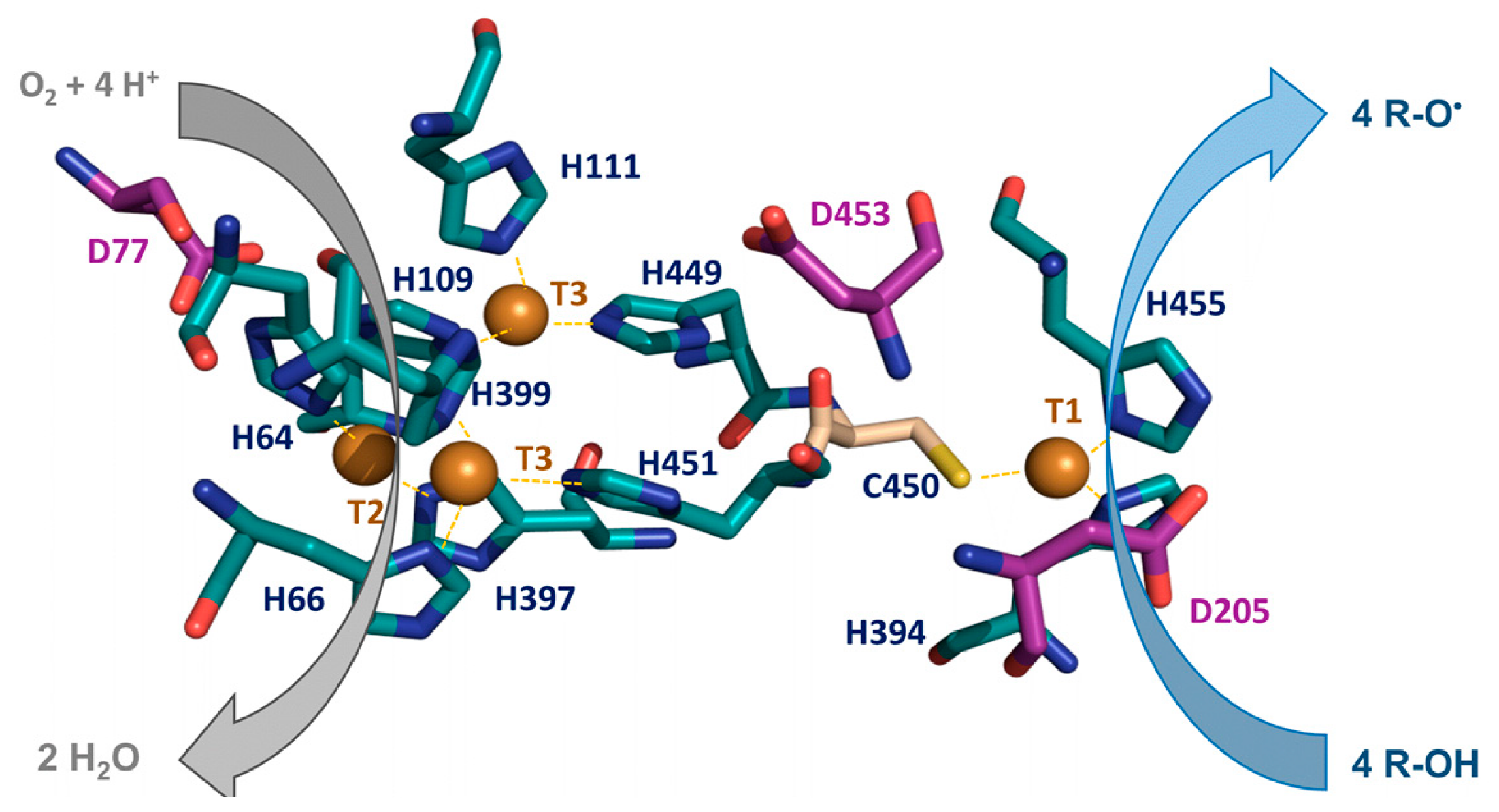
- Type 1 copper site (T1): located in D3, it is coordinated by two histidine residues and one cysteine residue in a trigonal coplanar arrangement. The Cu-S(Cys) bond is responsible for the typical blue color associated with these enzymes, resulting in a pronounced absorption in the visible region at 600 nm and a small parallel hyperfine coupling constant in electron paramagnetic resonance (EPR).
- Type 2 copper site (T2): composed of one copper coordinated by two histidines, it shows no absorption in the visible spectrum but reveals paramagnetic properties.
- Type 3 copper site (T3): this site is a binuclear center with two catalytic coppers coordinated by six histidine residues (three for each T3 copper atom). It is spectroscopically characterized by absorption at 330 nm and the absence of an EPR signal due to the antiferromagnetic coupling of the copper pair.
1.2. Catalytic Activity
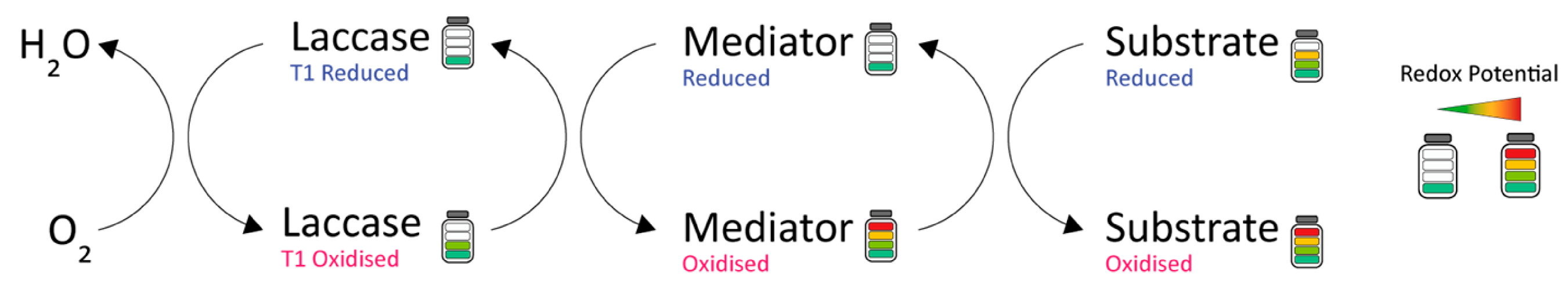
1.3. Redox Mediators
2. Multicopper Oxidases Reclassification
3. Biotechnological Applications
4. Laccase Engineering and Heterologous Production
4.1. Engineering of Fungal Laccases
4.2. Heterologous Expression
5. Challenges and Opportunities
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lindley, P.F. Handbook on Metalloproteins; Dekker, M., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2001; pp. 763–811. [Google Scholar]
- Yoshida, H. Chemistry of Lacquer (Urushi). J. Chem. Soc. 1883, 43, 472–486. [Google Scholar] [CrossRef]
- Bertrand, G. Sur La Presence Simultanee de La Laccase et de La Tyrosinase Dans Le Suc de Quelques Champignons. Comptes Rendus Hebd. Seances L’academie Sci. 1896, 123, 463–465. [Google Scholar] [CrossRef]
- Givaudan, A.; Effosse, A.; Faure, D.; Potier, P.; Bouillant, M.L.; Bally, R. Polyphenol Oxidase in Azospirillum Lipoferum Isolated from Rice Rhizosphere: Evidence for Laccase Activity in Non-Motile Strains of Azospirillum Lipoferum. FEMS Microbiol. Lett. 1993, 108, 205–210. [Google Scholar] [CrossRef]
- Dittmer, N.T.; Kanost, M.R. Insect Multicopper Oxidases: Diversity, Properties, and Physiological Roles. Insect Biochem. Mol. Biol. 2010, 40, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Janusz, G.; Pawlik, A.; Świderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkołazka, A.; Paszczyński, A. Laccase Properties, Physiological Functions, and Evolution. Int. J. Mol. Sci. 2020, 21, 966. [Google Scholar] [CrossRef] [PubMed]
- Thurston, C. The Structure and Function of Fungal Laccases. In Microbiology; Nomos Verlagsgesellschaft mbH & Co. KG: Baden-Baden, Germany, 1994; Volume 140, pp. 19–26. [Google Scholar]
- Claus, H. Laccases: Structure, Reactions, Distribution. Micron 2004, 35, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Martínez, Á.T.; Speranza, M.; Ruiz-Dueñas, F.J.; Ferreira, P.; Camarero, S.; Guillén, F.; Martínez, M.J.; Gutiérrez, A.; Del Río, J.C. Biodegradation of Lignocellulosics: Microbial, Chemical, and Enzymatic Aspects of the Fungal Attack of Lignin. Int. Microbiol. 2005, 8, 195–204. [Google Scholar] [CrossRef]
- De Salas, F.; Aza, P.; Gilabert, J.F.; Santiago, G.; Kilic, S.; Sener, M.E.; Vind, J.; Guallar, V.; Martínez, A.T.; Camarero, S. Engineering of a Fungal Laccase to Develop a Robust, Versatile and Highly-Expressed Biocatalyst for Sustainable Chemistry. Green Chem. 2019, 21, 5374–5385. [Google Scholar] [CrossRef]
- Escribano, D.R.; Magán, R.P.; Salas, F.D.; Aza, P.; Gentili, P.; Ihalainen, P.; Levée, T.; Meyer, V.; Conil, M.P.; Lingua, S.T.; et al. Tailor-Made Alkaliphilic and Thermostable Fungal Laccases for Industrial Wood Processing. Biotechnol. Biofuels Bioprod. 2022, 15, 149. [Google Scholar] [CrossRef]
- Baldrian, P. Fungal Laccases-Occurrence and Properties. FEMS Microbiol. Rev. 2006, 30, 215–242. [Google Scholar] [CrossRef]
- Zhukova, Y.N.; Lyashenko, A.V.; Lashkov, A.A.; Gur’Yanov, V.A.; Kobyl’skaya, Y.V.; Zhukhlistova, N.E.; Mikhailov, A.M. Atomic Structure of Unligated Laccase from Cerrena Maxima at 1.76 Å with Molecular Oxygen and Hydrogen Peroxide. Crystallogr. Rep. 2010, 55, 436–447. [Google Scholar] [CrossRef]
- Hakulinen, N.; Rouvinen, J. Three-Dimensional Structures of Laccases. Cell. Mol. Life Sci. 2015, 72, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Pardo, I.; Santiago, G.; Gentili, P.; Lucas, F.; Monza, E.; Medrano, F.J.; Galli, C.; Martínez, A.T.; Guallar, V.; Camarero, S. Re-Designing the Substrate Binding Pocket of Laccase for Enhanced Oxidation of Sinapic Acid. Catal. Sci. Technol. 2016, 6, 3900–3910. [Google Scholar] [CrossRef]
- Mot, A.C.; Silaghi-Dumitrescu, R. Laccases: Complex Architectures for One-Electron Oxidations. Biochemistry 2012, 77, 1395–1407. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Solomon, E.I. Electron Transfer and Reaction Mechanism of Laccases. Cell Mol Life Sci 2015, 72, 869–883. [Google Scholar] [CrossRef] [PubMed]
- Quintanar, L.; Yoon, J.; Aznar, C.P.; Palmer, A.E.; Andersson, K.K.; Britt, R.D.; Solomon, E.I. Spectroscopic and Electronic Structure Studies of the Trinuclear Cu Cluster Active Site of the Multicopper Oxidase Laccase: Nature of Its Coordination Unsaturation. J. Am. Chem. Soc. 2005, 127, 13832–13845. [Google Scholar] [CrossRef]
- Larrondo, L.F.; Salas, L.; Melo, F.; Vicuña, R.; Cullen, D. A Novel Extracellular Multicopper Oxidase from Phanerochaete Chrysosporium with Ferroxidase Activity. Appl. Environ. Microbiol. 2003, 69, 6257–6263. [Google Scholar] [CrossRef]
- Quintanar, L.; Stoj, C.; Taylor, A.B.; Hart, P.J.; Kosman, D.J.; Solomon, E.I. Shall We Dance? How a Multicopper Oxidase Chooses Its Electron Transfer Partner. Acc. Chem. Res. 2007, 40, 445–452. [Google Scholar] [CrossRef]
- Mehra, R.; Muschiol, J.; Meyer, A.S.; Kepp, K.P. A Structural-Chemical Explanation of Fungal Laccase Activity. Sci. Rep. 2018, 8, 17285. [Google Scholar] [CrossRef]
- Xu, F.; Shin, W.; Brown, S.H.; Wahleithner, J.A.; Sundaram, U.M.; Solomon, E.I. A Study of a Series of Recombinant Fungal Laccases and Bilirubin Oxidase That Exhibit Significant Differences in Redox Potential, Substrate Specificity, and Stability. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1996, 1292, 303–311. [Google Scholar] [CrossRef]
- Xu, F. Oxidation of Phenols, Anilines, and Benzenethiols by Fungal Laccases: Correlation between Activity and Redox Potentials as Well as Halide Inhibition. Biochemistry 1996, 35, 7608–7614. [Google Scholar] [CrossRef]
- Zille, A.; Gornacka, B.; Rehorek, A.; Cavaco-Paulo, A. Degradation of Azo Dyes by Trametes Villosa Laccase over Long Periods of Oxidative Conditions. Appl. Environ. Microbiol. 2005, 71, 6711–6718. [Google Scholar] [CrossRef]
- de Salas, F.; Camarero, S. Fungal Laccases as Biocatalysts for Wide Range Applications. Encycl. Mycol. 2021, 2, 233–238. [Google Scholar] [CrossRef]
- Galli, C.; Madzak, C.; Vadalà, R.; Jolivalt, C.; Gentili, P. Concerted Electron/Proton Transfer Mechanism in the Oxidation of Phenols by Laccase. ChemBioChem 2013, 14, 2500–2505. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.J.; Lancaster, K.M.; Richards, J.H.; Gray, H.B. Inner- and Outer-Sphere Metal Coordination in Blue Copper Proteins. J. Inorg. Biochem. 2012, 115, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Polyakov, K.M.; Gavryushov, S.; Ivanova, S.; Fedorova, T.V.; Glazunova, O.A.; Popov, A.N.; Koroleva, O.V. Structural Study of the X-Ray-Induced Enzymatic Reduction of Molecular Oxygen to Water by Steccherinum Murashkinskyi Laccase: Insights into the Reaction Mechanism. Acta Crystallogr. Sect. D Struct. Biol. 2017, 73, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Xu, F. Effects of Redox Potential and Hydroxide Inhibition on the PH Activity Profile of Fungal Laccases. J. Biol. Chem. 1997, 272, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Morozova, O.V.; Shumakovich, G.P.; Gorbacheva, M.A.; Shleev, S.V.; Yaropolov, A.I. “Blue” Laccases. Biochemistry 2007, 72, 1136–1150. [Google Scholar] [CrossRef]
- Sekretaryova, A.; Jones, S.M.; Solomon, E.I. O2 Reduction to Water by High Potential Multicopper Oxidases: Contributions of the T1 Copper Site Potential and the Local Environment of the Trinuclear Copper Cluster. J. Am. Chem. Soc. 2019, 141, 11304–11314. [Google Scholar] [CrossRef]
- Hall, J.F.; Kanbi, L.D.; Strange, R.W.; Hasnain, S.S. Role of the Axial Ligand in Type 1 Cu Centers Studied by Point Mutations of Met148 in Rusticyanin. Biochemistry 1999, 38, 12675–12680. [Google Scholar] [CrossRef]
- Xu, F.; Palmer, A.E.; Yaver, D.S.; Berka, R.M.; Gambetta, G.A.; Brown, S.H.; Solomon, E.I. Targeted Mutations in a Trametes Villosa Laccase: Axial Perturbations of the T1 Copper. J. Biol. Chem. 1999, 274, 12372–12375. [Google Scholar] [CrossRef] [PubMed]
- Hakulinen, N.; Kiiskinen, L.L.; Kruus, K.; Saloheimo, M.; Paanen, A.; Koivula, A.; Rouvinen, J. Crystal Structure of a Laccase from Melanocarpus Albomyces with an Intact Trinuclear Copper Site. Nat. Struct. Biol. 2002, 9, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Piontek, K.; Antorini, M.; Choinowski, T. Crystal Structure of a Laccase from the Fungus Trametes Versicolor at 1.90-Å Resolution Containing a Full Complement of Coppers. J. Biol. Chem. 2002, 277, 37663–37669. [Google Scholar] [CrossRef] [PubMed]
- Cambria, M.T.; Gullotto, D.; Garavaglia, S.; Cambria, A. In Silico Study of Structural Determinants Modulating the Redox Potential of Rigidoporus Lignosus and Other Fungal Laccases. J. Biomol. Struct. Dyn. 2012, 30, 89–101. [Google Scholar] [CrossRef]
- Nersissian, A.M.; Shipp, E.L. Blue Copper-Binding Domains. Adv. Protein Chem. 2002, 60, 271–340. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, S.; Cambria, M.T.; Miglio, M.; Ragusa, S.; Iacobazzi, V.; Palmieri, F.; D’Ambrosio, C.; Scaloni, A.; Rizzi, M. The Structure of Rigidoporus Lignosus Laccase Containing a Full Complement of Copper Ions, Reveals an Asymmetrical Arrangement for the T3 Copper Pair. J. Mol. Biol. 2004, 342, 1519–1531. [Google Scholar] [CrossRef]
- Baiocco, P.; Barreca, A.M.; Fabbrini, M.; Galli, C.; Gentili, P. Promoting Laccase Activity towards Non-Phenolic Substrates: A Mechanistic Investigation with Some Laccase-Mediator Systems. Org. Biomol. Chem. 2003, 1, 191–197. [Google Scholar] [CrossRef]
- Kunamneni, A.; Camarero, S.; García-Burgos, C.; Plou, F.J.; Ballesteros, A.; Alcalde, M. Engineering and Applications of Fungal Laccases for Organic Synthesis. Microb. Cell Fact. 2008, 7, 32. [Google Scholar] [CrossRef]
- Bourbonnais, R.; Paice, M.G. Oxidation of Non-Phenolic Substrates. An Expanded Role for Laccase in Lignin Biodegradation. FEBS Lett. 1990, 267, 99–102. [Google Scholar] [CrossRef]
- Bourbonnais, R.; Paice, M.G.; Freiermuth, B.; Bodie, E.; Borneman, S. Reactivities of Various Mediators and Laccases with Kraft Pulp and Lignin Model Compounds. Appl. Environ. Microbiol. 1997, 63, 4627–4632. [Google Scholar] [CrossRef]
- Li, K.; Xu, F.; Eriksson, K.-E.L. Comparison of Fungal Laccases and Redox Mediators in Oxidation of a Nonphenolic Lignin Model Compound. Appl. Environ. Microbiol. 1999, 65, 2654–2660. [Google Scholar] [CrossRef]
- Fabbrini, M.; Galli, C.; Gentili, P.; Macchitella, D. An Oxidation of Alcohols by Oxygen with the Enzyme Laccase and Mediation by TEMPO. Tetrahedron Lett. 2001, 43, 7551–7553. [Google Scholar] [CrossRef]
- Majcherczyk, A.; Johannes, C.; Hüttermann, A. Oxidation of Polycyclic Aromatic Hydrocarbons (PAH) by Laccase of Trametes Versicolor. Enzym. Microb. Technol. 1998, 22, 335–341. [Google Scholar] [CrossRef]
- Ibarra, D.; Romero, J.; Martínez, M.J.; Martínez, A.T.; Camarero, S. Exploring the Enzymatic Parameters for Optimal Delignification of Eucalypt Pulp by Laccase-Mediator. Enzym. Microb. Technol. 2006, 39, 1319–1327. [Google Scholar] [CrossRef]
- Quintana, E.; Roncero, M.B.; Vidal, T.; Valls, C. Cellulose Oxidation by Laccase-TEMPO Treatments. Carbohydr. Polym. 2017, 157, 1488–1495. [Google Scholar] [CrossRef] [PubMed]
- Hilgers, R.; Van Dam, A.; Zuilhof, H.; Vincken, J.P.; Kabel, M.A. Controlling the Competition: Boosting Laccase/HBT-Catalyzed Cleavage of a β-O-4′ Linked Lignin Model. ACS Catal. 2020, 10, 8650–8659. [Google Scholar] [CrossRef]
- Eggert, C.; Temp, U.; Dean, J.F.D.; Eriksson, K.-E.L. A Fungal Metabolite Mediates Degradation of Non-Phenolic Lignin Structures and Synthetic Lignin by Laccase. FEBS Lett. 1996, 391, 144–148. [Google Scholar] [CrossRef]
- Johannes, C.; Majcherczyk, A. Natural Mediators in the Oxidation of Polycyclic Aromatic Hydrocarbons by Laccase Mediator Systems. Appl. Environ. Microbiol. 2000, 66, 524–528. [Google Scholar] [CrossRef]
- Cañas, A.I.; Camarero, S. Laccases and Their Natural Mediators: Biotechnological Tools for Sustainable Eco-Friendly Processes. Biotechnol. Adv. 2010, 28, 694–705. [Google Scholar] [CrossRef]
- Camarero, S.; Ibarra, D.; Martínez, Á.T.; Romero, J.; Gutiérrez, A.; del Río, J.C. Paper Pulp Delignification Using Laccase and Natural Mediators. Enzym. Microb. Technol. 2007, 40, 1264–1271. [Google Scholar] [CrossRef]
- Camarero, S.; Ibarra, D.; Martínez, M.J.; Martínez, Á.T. Lignin-Derived Compounds as Efficient Laccase Mediators for Decolorization of Different Types of Recalcitrant Dyes. Appl. Environ. Microbiol. 2005, 71, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Cañas, A.I.; Alcalde, M.; Plou, F.; Martínez, M.J.; Martínez, Á.T.; Camarero, S. Transformation of Polycyclic Aromatic Hydrocarbons by Laccase Is Strongly Enhanced by Phenolic Compounds Present in Soil. Environ. Sci. Technol. 2007, 41, 2964–2971. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.V.S.; Phale, P.S.; Durani, S.; Wangikar, P.P. Combined Sequence and Structure Analysis of the Fungal Laccase Family. Biotechnol. Bioeng. 2003, 83, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Go, N. Function and Molecular Evolution of Multicopper Blue Proteins. Cell. Mol. Life Sci. 2005, 62, 2050–2066. [Google Scholar] [CrossRef] [PubMed]
- Eggert, C.; Lafayette, P.R.; Temp, U.; Eriksson, K.-E.L.; Dean, J.F.D. Molecular Analysis of a Laccase Gene from the White Rot Fungus Pycnoporus Cinnabarinus. Appl. Environ. Microbiol. 1998, 64, 1766–1772. [Google Scholar] [CrossRef] [PubMed]
- Reiss, R.; Ihssen, J.; Richter, M.; Eichhorn, E.; Schilling, B.; Thöny-Meyer, L. Laccase versus Laccase-Like Multi-Copper Oxidase: A Comparative Study of Similar Enzymes with Diverse Substrate Spectra. PLoS ONE 2013, 8, e65633. [Google Scholar] [CrossRef] [PubMed]
- Hoegger, P.J.; Kilaru, S.; James, T.Y.; Thacker, J.R.; Kües, U. Phylogenetic Comparison and Classification of Laccase and Related Multicopper Oxidase Protein Sequences. FEBS J. 2006, 273, 2308–2326. [Google Scholar] [CrossRef]
- Valderrama, B.; Oliver, P.; Medrano-Soto, A.; Vazquez-Duhalt, R. Evolutionary and Structural Diversity of Fungal Laccases. Antonie van Leeuwenhoek, Int. J. Gen. Mol. Microbiol. 2003, 84, 289–299. [Google Scholar] [CrossRef]
- Moiseenko, K.V.; Maloshenok, L.G.; Vasina, D.V.; Bruskin, S.A.; Tyazhelova, T.V.; Koroleva, O.V. Laccase Multigene Families in Agaricomycetes. J. Basic Microbiol. 2016, 56, 1392–1397. [Google Scholar] [CrossRef]
- Savinova, O.S.; Moiseenko, K.V.; Vavilova, E.A.; Chulkin, A.M.; Fedorova, T.V.; Tyazhelova, T.V.; Vasina, D.V. Evolutionary Relationships between the Laccase Genes of Polyporales: Orthology-Based Classification of Laccase Isozymes and Functional Insight from Trametes Hirsuta. Front. Microbiol. 2019, 10, 152. [Google Scholar] [CrossRef]
- Ruiz-Dueñas, F.J.; Barrasa, J.M.; Sánchez-García, M.; Camarero, S.; Miyauchi, S.; Serrano, A.; Linde, D.; Babiker, R.; Drula, E.; Ayuso-Fernández, I.; et al. Genomic Analysis Enlightens Agaricales Lifestyle Evolution and Increasing Peroxidase Diversity. Mol. Biol. Evol. 2021, 38, 1428–1446. [Google Scholar] [CrossRef]
- De Tullio, M.C.; Guether, M.; Balestrini, R. Ascorbate Oxidase Is the Potential Conductor of a Symphony of Signaling Pathways. Plant Signal. Behav. 2013, 8, 23213. [Google Scholar] [CrossRef] [PubMed]
- Braunschmid, V.; Fuerst, S.; Perz, V.; Zitzenbacher, S.; Hoyo, J.; Fernandez-Sanchez, C.; Tzanov, T.; Steinkellner, G.; Gruber, K.; Nyanhongo, G.S.; et al. A Fungal Ascorbate Oxidase with Unexpected Laccase Activity. Int. J. Mol. Sci. 2020, 21, 5754. [Google Scholar] [CrossRef]
- Pignocchi, C.; Foyer, C.H. Apoplastic Ascorbate Metabolism and Its Role in the Regulation of Cell Signalling. Curr. Opin. Plant Biol. 2003, 6, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Stearman, R.; Yuan, D.S.; Yamaguchi-Iwai, Y.; Klausner, R.D.; Dancis, A. A Permease-Oxidase Complex Involved in High-Affinity Iron Uptake in Yeast. Science 1996, 271, 1552–1557. [Google Scholar] [CrossRef]
- Taylor, A.B.; Stoj, C.S.; Ziegler, L.; Kosman, D.J.; Hart, P.J. The Copper-Iron Connection in Biology: Structure of the Metallo-Oxidase Fet3p. Proc. Natl. Acad. Sci. USA 2005, 102, 15459–15464. [Google Scholar] [CrossRef] [PubMed]
- Stoj, C.S.; Augustine, A.J.; Zeigler, L.; Solomon, E.I.; Kosman, D.J. Structural Basis of the Ferrous Iron Specificity of the Yeast Ferroxidase, Fet3p. Biochemistry 2006, 45, 12741–12749. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Tewari, R.P.; Williamson, P.R. Laccase Protects Cryptococcus neoformans from Antifungal Activity of Alveolar Macrophages. Infect. Immun. 1999, 67, 6034–6039. [Google Scholar] [CrossRef]
- Williamson, P.R. Biochemical and Molecular Characterization of the Diphenol Oxidase of Cryptococcus neoformans: Identification as a Laccase. J. Bacteriol. 1994, 176, 656–664. [Google Scholar] [CrossRef]
- Rodríguez-Rincón, F.; Suarez, A.; Lucas, M.; Larrondo, L.F.; De La Rubia, T.; Polaina, J.; Martínez, J. Molecular and Structural Modeling of the Phanerochaete Flavido-Alba Extracellular Laccase Reveals Its Ferroxidase Structure. Arch. Microbiol. 2010, 192, 883–892. [Google Scholar] [CrossRef]
- Aza, P.; Molpeceres, G.; Vind, J.; Camarero, S. Multicopper Oxidases with Laccase-Ferroxidase Activity: Classification and Study of Ferroxidase Activity Determinants in a Member from Heterobasidion annosum s. l. Comput. Struct. Biotechnol. J. 2023, 21, 1041–1053. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, T.; Jolivalt, C.; Briozzo, P.; Caminade, E.; Joly, N.; Madzak, C.; Mougin, C. Crystal Structure of a Four-Copper Laccase Complexed with an Arylamine: Insights into Substrate Recognition and Correlation with Kinetics. Biochemistry 2002, 41, 7325–7333. [Google Scholar] [CrossRef] [PubMed]
- Faraco, V.; Ercole, C.; Festa, G.; Giardina, P.; Piscitelli, A.; Sannia, G. Heterologous Expression of Heterodimeric Laccase from Pleurotus ostreatus in Kluyveromyces lactis. Appl. Microbiol. Biotechnol. 2008, 77, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yuan, C.; Wang, F.; Xu, S.; Li, Y.; Shi, G.; Ding, Z. Roles of Small Subunits of Laccase (SsPOXA3a/b) in Laccase Production by Pleurotus eryngii var. ferulae. J. Agric. Food Chem. 2021, 69, 13113–13124. [Google Scholar] [CrossRef]
- Aza, P.; Linde, D.; Molpeceres, G.; Vind, J.; Medrano, F.J.; Camarero, S. Role and Structure of the Small Subunit Forming Heterodimers with Laccase-like Enzymes. Protein Sci. 2023, 32, 4734. [Google Scholar] [CrossRef] [PubMed]
- Valls, C.; Roncero, M.B. Using Both Xylanase and Laccase Enzymes for Pulp Bleaching. Bioresour. Technol. 2009, 100, 2032–2039. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, P.; Shrivastava, R.; Agrawal, P.K. Bioprospecting and Biotechnological Applications of Fungal Laccase. 3 Biotech 2016, 6, 15. [Google Scholar] [CrossRef]
- Moreno, A.D.; Ibarra, D.; Eugenio, M.E.; Tomás-Pejó, E. Laccases as Versatile Enzymes: From Industrial Uses to Novel Applications. J. Chem. Technol. Biotechnol. 2020, 95, 481–494. [Google Scholar] [CrossRef]
- Almeida, N.; Meyer, V.; Burnet, A.; Boucher, J.; Talens-Perales, D.; Pereira, S.; Ihalainen, P.; Levée, T.; Polaina, J.; Petit-Conil, M.; et al. Use of a Novel Extremophilic Xylanase for an Environmentally Friendly Industrial Bleaching of Kraft Pulps. Int. J. Mol. Sci. 2022, 23, 3423. [Google Scholar] [CrossRef]
- Fillat, U.; Prieto, A.; Camarero, S.; Martínez, Á.T.; Martínez, M.J. Biodeinking of Flexographic Inks by Fungal Laccases Using Synthetic and Natural Mediators. Biochem. Eng. J. 2012, 67, 97–103. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Del Río, J.C.; Ibarra, D.; Rencoret, J.; Romero, J.; Speranza, M.; Camarero, S.; Martínez, M.J.; Martínez, Á.T. Enzymatic Removal of Free and Conjugated Sterols Forming Pitch Deposits in Environmentally Sound Bleaching of Eucalypt Paper Pulp. Environ. Sci. Technol. 2006, 40, 3416–3422. [Google Scholar] [CrossRef] [PubMed]
- Virk, A.P.; Sharma, P.; Capalash, N. Use of Laccase in Pulp and Paper Industry. Biotechnol. Prog. 2012, 28, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Escribano, D.; Salas, F.D.; Marques, G.; Lev, T.; Suonpää, A.; Guti, A.; Mart, Á.T.; Ihalainen, P.; Rencoret, J.; Camarero, S. Depolymerisation of Kraft Lignin by Tailor-Made Alkaliphilic. Polymers 2023, 15, 4433. [Google Scholar] [CrossRef]
- Pazarlioǧlu, N.K.; Sariişik, M.; Telefoncu, A. Laccase: Production by Trametes versicolor and Application to Denim Washing. Process Biochem. 2005, 40, 1673–1678. [Google Scholar] [CrossRef]
- Pezzella, C.; Giacobbe, S.; Giacobelli, V.G.; Guarino, L.; Kylic, S.; Sener, M.; Sannia, G.; Piscitelli, A. Green Routes towards Industrial Textile Dyeing: A Laccase Based Approach. J. Mol. Catal. B Enzym. 2016, 134, 274–279. [Google Scholar] [CrossRef]
- Su, J.; Shim, E.; Noro, J.; Fu, J.; Wang, Q.; Kim, H.R.; Silva, C.; Cavaco-Paulo, A. Conductive Cotton by in Situ Laccase-Polymerization of Aniline. Polymers 2018, 10, 1023. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Noro, J.; Silva, S.; Fu, J.; Wang, Q.; Ribeiro, A.; Silva, C.; Cavaco-Paulo, A. Antimicrobial Coating of Textiles by Laccase in Situ Polymerization of Catechol and P-Phenylenediamine. React. Funct. Polym. 2019, 136, 25–33. [Google Scholar] [CrossRef]
- Schultz, A.; Jonas, U.; Hammer, E.; Schauer, F. Dehalogenation of Chlorinated Hydroxybiphenyls by Fungal Laccase. Appl. Environ. Microbiol. 2001, 67, 4377–4381. [Google Scholar] [CrossRef]
- Teng, C.; Wu, S.; Gong, G. Bio-Removal of Phenanthrene, 9-Fluorenone and Anthracene-9,10-Dione by Laccase from Aspergillus niger in Waste Cooking Oils. Food Control 2019, 105, 219–225. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Yi, X.Y.; Zhang, M.; Liu, L. Fundamental Study of Degradation of Dichlorodiphenyltrichloroethane in Soil by Laccase from White Rot Fungi. Int. J. Environ. Sci. Technol. 2010, 7, 359–366. [Google Scholar] [CrossRef]
- Shi, H.; Peng, J.; Li, J.; Mao, L.; Wang, Z.; Gao, S. Laccase-Catalyzed Removal of the Antimicrobials Chlorophene and Dichlorophen from Water: Reaction Kinetics, Pathway and Toxicity Evaluation. J. Hazard. Mater. 2016, 317, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Limon, A.; García Suárez, P.C.; Arellano-García, E.; Contreras, O.E.; Aguila, S.A. Enhanced Degradation of Pesticide Dichlorophen by Laccase Immobilized on Nanoporous Materials: A Cytotoxic and Molecular Simulation Investigation. Bioconjug. Chem. 2018, 29, 1073–1080. [Google Scholar] [CrossRef]
- Mayolo-Deloisa, K.; González-González, M.; Rito-Palomares, M. Laccases in Food Industry: Bioprocessing, Potential Industrial and Biotechnological Applications. Front. Bioeng. Biotechnol. 2020, 8, 222. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, W.; Ng, T.B.; Deng, X.; Lin, J.; Ye, X. Laccases: Production, Expression Regulation, and Applications in Pharmaceutical Biodegradation. Front. Microbiol. 2017, 8, 832. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, C.E.; Marco-Urrea, E.; Caminal, G. Degradation of Naproxen and Carbamazepine in Spiked Sludge by Slurry and Solid-Phase Trametes versicolor Systems. Bioresour. Technol. 2010, 101, 2259–2266. [Google Scholar] [CrossRef]
- Janusz, G.; Skwarek, E.; Pawlik, A. Potential of Laccase as a Tool for Biodegradation of Wastewater Micropollutants. Water 2023, 15, 3770. [Google Scholar] [CrossRef]
- Polak, J.; Jarosz-Wilkolazka, A. Fungal Laccases as Green Catalysts for Dye Synthesis. Process Biochem. 2012, 47, 1295–1307. [Google Scholar] [CrossRef]
- Kim, S.; Silva, C.; Evtuguin, D.V.; Gamelas, J.A.F.; Cavaco-Paulo, A. Polyoxometalate/Laccase-Mediated Oxidative Polymerization of Catechol for Textile Dyeing. Appl. Microbiol. Biotechnol. 2011, 89, 981–987. [Google Scholar] [CrossRef]
- Giacobelli, V.G.; Pezzella, C.; Sannia, G.; Olivieri, G.; Fontanarosa, C.; Amoresano, A.; Piscitelli, A. Laccase-Based Synthesis of SIC-RED: A New Dyeing Product for Protein Gel Staining. Biocatal. Agric. Biotechnol. 2018, 15, 270–276. [Google Scholar] [CrossRef]
- Aktaş, N.; Tanyolaç, A. Reaction Conditions for Laccase Catalyzed Polymerization of Catechol. Bioresour. Technol. 2003, 87, 209–214. [Google Scholar] [CrossRef]
- De Salas, F.; Pardo, I.; Salavagione, H.J.; Aza, P.; Amougi, E.; Vind, J.; Martínez, A.T.; Camarero, S. Advanced Synthesis of Conductive Polyaniline Using Laccase as Biocatalyst. PLoS ONE 2016, 11, e0164958. [Google Scholar] [CrossRef]
- Feng, X.M.; Li, R.M.; Ma, Y.W.; Chen, R.F.; Shi, N.E.; Fan, Q.L.; Huang, W. One-Step Electrochemical Synthesis of Graphene/Polyaniline Composite Film and Its Applications. Adv. Funct. Mater. 2011, 21, 2989–2996. [Google Scholar] [CrossRef]
- Hu, L.; Ren, Y.; Yang, H.; Xu, Q. Fabrication of 3D Hierarchical MoS2/Polyaniline and MoS 2/C Architectures for Lithium-Ion Battery Applications. ACS Appl. Mater. Interfaces 2014, 6, 14644–14652. [Google Scholar] [CrossRef] [PubMed]
- Osiadacz, J.; Al-Adhami, A.J.H.; Bajraszewska, D.; Fischer, P.; Peczyñska-Czoch, W. On the Use of Trametes versicolor Laccase for the Conversion of 4-Methyl-3-Hydroxyanthranilic Acid to Actinocin Chromophore. J. Biotechnol. 1999, 72, 141–149. [Google Scholar] [CrossRef]
- Burton, S. Laccases and Phenol Oxidases in Organic Synthesis—A Review. Curr. Org. Chem. 2005, 7, 1317–1331. [Google Scholar] [CrossRef]
- Sagui, F.; Chirivì, C.; Fontana, G.; Nicotra, S.; Passarella, D.; Riva, S.; Danieli, B. Laccase-Catalyzed Coupling of Catharanthine and Vindoline: An Efficient Approach to the Bisindole Alkaloid Anhydrovinblastine. Tetrahedron 2009, 65, 312–317. [Google Scholar] [CrossRef]
- Nicotra, S.; Intra, A.; Ottolina, G.; Riva, S.; Danieli, B. Laccase-Mediated Oxidation of the Steroid Hormone 17β-Estradiol in Organic Solvents. Tetrahedron Asymmetry 2004, 15, 2927–2931. [Google Scholar] [CrossRef]
- Nicotra, S.; Cramarossa, M.R.; Mucci, A.; Pagnoni, U.M.; Riva, S.; Forti, L. Biotransformation of Resveratrol: Synthesis of Trans-Dehydrodimers Catalyzed by Laccases from Myceliophtora thermophyla and from Trametes pubescens. Tetrahedron 2004, 60, 595–600. [Google Scholar] [CrossRef]
- Vittorio, O.; Cojoc, M.; Curcio, M.; Spizzirri, U.G.; Hampel, S.; Nicoletta, F.P.; Iemma, F.; Dubrovska, A.; Kavallaris, M.; Cirillo, G. Polyphenol Conjugates by Immobilized Laccase: The Green Synthesis of Dextran-Catechin. Macromol. Chem. Phys. 2016, 217, 1488–1492. [Google Scholar] [CrossRef]
- Fürtges, L.; Obermaier, S.; Thiele, W.; Foegen, S.; Müller, M. Diversity in Fungal Intermolecular Phenol Coupling of Polyketides: Regioselective Laccase-Based Systems. ChemBioChem 2019, 20, 1928–1932. [Google Scholar] [CrossRef]
- Osma, J.F.; Toca-Herrera, J.L.; Rodríguez-Couto, S. Uses of Laccases in the Food Industry. Enzym. Res. 2010, 2010, 918761. [Google Scholar] [CrossRef] [PubMed]
- Minussi, R.C.; Pastore, G.M.; Durán, N. Potential Applications of Laccase in the Food Industry. Trends Food Sci. Technol. 2002, 13, 205–216. [Google Scholar] [CrossRef]
- Kunamneni, A.; Plou, F.; Ballesteros, A.; Alcalde, M. Laccases and Their Applications: A Patent Review. Recent Pat. Biotechnol. 2008, 2, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Talens-Perales, D.; Nicolau-Sanus, M.; Polaina, P.; Daròs, J.A. Phylogenetic, Functional and Structural Characterization of a GH10 Xylanase Active at Extreme Conditions of Temperature and Alkalinity. Sci. Rep. 2022, 21, 19774–19775. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.M.; Han, S.S.; Kim, H.S. Industrial Applications of Enzyme Biocatalysis: Current Status and Future Aspects. Biotechnol. Adv. 2015, 33, 1443–1454. [Google Scholar] [CrossRef]
- Arnold, F.H.; Volkov, A.A. Directed Evolution of Biocatalysts. Curr. Opin. Chem. Biol. 1999, 3, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Tracewell, C.A.; Arnold, F.H. Directed Enzyme Evolution: Climbing Fitness Peaks One Amino Acid at a Time. Curr. Opin. Chem. Biol. 2009, 13, 3–9. [Google Scholar] [CrossRef]
- Brandenberg, O.F.; Fasan, R.; Arnold, F.H. Exploiting and Engineering Hemoproteins for Abiological Carbene and Nitrene Transfer Reactions. Curr. Opin. Biotechnol. 2017, 47, 102–111. [Google Scholar] [CrossRef]
- Sebestova, E.; Bendl, J.; Brezovsky, J.; Damborsky, J. Computational tools for designing smart libraries. In Directed Evolution Library Creation. Methods in Molecular Biology; Gillam, E., Copp, J., Ackerley, D., Eds.; Springer: New York, NY, USA, 2014; Volume 1179. [Google Scholar] [CrossRef]
- Huang, P.S.; Boyken, S.E.; Baker, D. The Coming of Age of de Novo Protein Design. Nature 2016, 537, 320–327. [Google Scholar] [CrossRef]
- Markel, U.; Essani, K.D.; Besirlioglu, V.; Schiffels, J.; Streit, W.R.; Schwaneberg, U. Advances in Ultrahigh-Throughput Screening for Directed Enzyme Evolution. Chem. Soc. Rev. 2020, 49, 233–262. [Google Scholar] [CrossRef]
- Morrison, M.S.; Podracky, C.J.; Liu, D.R. The Developing Toolkit of Continuous Directed Evolution. Nat. Chem. Biol. 2020, 16, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Romero, P.A.; Arnold, F.H. Exploring Protein Fitness Landscapes by Directed Evolution. Nat. Rev. Mol. Cell Biol. 2009, 10, 866–876. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xue, P.; Cao, M.; Yu, T.; Lane, S.T.; Zhao, H. Directed Evolution: Methodologies and Applications. Chem. Rev. 2021, 121, 12384–12444. [Google Scholar] [CrossRef]
- Bulter, T.; Alcalde, M.; Sieber, V.; Meinhold, P.; Schlachtbauer, C.; Arnold, F.H. Functional Expression of a Fungal Laccase in Saccharomyces Cerevisiae by Directed Evolution. Appl. Environ. Microbiol. 2003, 69, 987–995. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zumárraga, M.; Bulter, T.; Shleev, S.; Polaina, J.; Martínez-Arias, A.; Plou, F.J.; Ballesteros, A.; Alcalde, M. In Vitro Evolution of a Fungal Laccase in High Concentrations of Organic Cosolvents. Chem. Biol. 2007, 14, 1052–1064. [Google Scholar] [CrossRef] [PubMed]
- Festa, G.; Autore, F.; Fraternali, F.; Giardina, P.; Sannia, G. Development of New Laccases by Directed Evolution: Functional and Computational Analyses. Proteins Struct. Funct. Genet. 2008, 72, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Miele, A.; Giardina, P.; Sannia, G.; Faraco, V. Random Mutants of a Pleurotus ostreatus Laccase as New Biocatalysts for Industrial Effluents Bioremediation. J. Appl. Microbiol. 2010, 108, 998–1006. [Google Scholar] [CrossRef]
- Hu, M.R.; Chao, Y.P.; Zhang, G.Q.; Yang, X.Q.; Xue, Z.Q.; Qian, S.J. Molecular Evolution of Fome lignosus Laccase by Ethyl Methane Sulfonate-Based Random Mutagenesis in Vitro. Biomol. Eng. 2007, 24, 619–624. [Google Scholar] [CrossRef]
- Maté, D.; García-Burgos, C.; García-Ruiz, E.; Ballesteros, A.O.; Camarero, S.; Alcalde, M. Laboratory Evolution of High-Redox Potential Laccases. Chem. Biol. 2010, 17, 1030–1041. [Google Scholar] [CrossRef]
- Camarero, S.; Pardo, I.; Cañas, A.I.; Molina, P.; Record, E.; Martínez, A.T.; Martínez, M.J.; Alcalde, M. Engineering Platforms for Directed Evolution of Laccase from Pycnoporus cinnabarinus. Appl. Environ. Microbiol. 2012, 78, 1370–1384. [Google Scholar] [CrossRef]
- Abécassis, V.; Pompom, D.; Truan, G. Producing chimeric genes by CLERY. In Directed Evolution Library Creation. Methods in Molecular Biology; Arnold, F.H., Georgiou, G., Eds.; Humana Press: Totowa, NY, USA, 2003; Volume 231, pp. 3–11. [Google Scholar]
- Pardo, I.; Vicente, A.I.; Mate, D.M.; Alcalde, M.; Camarero, S. Development of Chimeric Laccases by Directed Evolution. Biotechnol. Bioeng. 2012, 109, 2978–2986. [Google Scholar] [CrossRef] [PubMed]
- Pardo, I.; Rodríguez-Escribano, D.; Aza, P.; de Salas, F.; Martínez, A.T.; Camarero, S. A Highly Stable Laccase Obtained by Swapping the Second Cupredoxin Domain. Sci. Rep. 2018, 8, 15669. [Google Scholar] [CrossRef]
- Santiago, G.; De Salas, F.; Lucas, M.F.; Monza, E.; Acebes, S.; Martinez, Á.T.; Camarero, S.; Guallar, V. Computer-Aided Laccase Engineering: Toward Biological Oxidation of Arylamines. ACS Catal. 2016, 6, 5415–5423. [Google Scholar] [CrossRef]
- Mate, D.M.; Gonzalez-Perez, D.; Falk, M.; Kittl, R.; Pita, M.; De Lacey, A.L.; Ludwig, R.; Shleev, S.; Alcalde, M. Blood Tolerant Laccase by Directed Evolution. Chem. Biol. 2013, 20, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhu, L.; Bocola, M.; Chen, N.; Spiess, A.C.; Schwaneberg, U. Directed Laccase Evolution for Improved Ionic Liquid Resistance. Green Chem. 2013, 15, 1348–1355. [Google Scholar] [CrossRef]
- Torres-Salas, P.; Mate, D.M.; Ghazi, I.; Plou, F.J.; Ballesteros, A.O.; Alcalde, M. Widening the PH Activity Profile of a Fungal Laccase by Directed Evolution. ChemBioChem 2013, 14, 934–937. [Google Scholar] [CrossRef] [PubMed]
- Scheiblbrandner, S.; Breslmayr, E.; Csarman, F.; Paukner, R.; Führer, J.; Herzog, P.L.; Shleev, S.V.; Osipov, E.M.; Tikhonova, T.V.; Popov, V.O.; et al. Evolving Stability and PH-Dependent Activity of the High Redox Potential Botrytis aclada Laccase for Enzymatic Fuel Cells. Sci. Rep. 2017, 7, 13688. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, F.; Zhang, X.; Geng, A. Directed Evolution of a Homodimeric Laccase from Cerrena unicolor BBP6 by Random Mutagenesis and in Vivo Assembly. Int. J. Mol. Sci. 2018, 19, 2989. [Google Scholar] [CrossRef]
- Yin, Q.; Zhou, G.; Peng, C.; Zhang, Y.; Kües, U.; Liu, J.; Xiao, Y.; Fang, Z. The First Fungal Laccase with an Alkaline PH Optimum Obtained by Directed Evolution and Its Application in Indigo Dye Decolorization. AMB Express 2019, 9, 151. [Google Scholar] [CrossRef]
- Su, X.; Yang, J.; Yuan, H.; Liu, C.; Tu, R.; Liu, P.; Wang, Q.; Zhu, L. Directed Evolution of Laccase for Improved Thermal Stability Facilitated by Droplet-Based Microfluidic Screening System. J. Agric. Food Chem. 2022, 70, 13700–13708. [Google Scholar] [CrossRef]
- Rogalski, J.; Lundell, T.K.; Leonowicz, A.; Hatakka, A.I. Influence of Aromatic Compounds and Lignin on Production of Ligninolytic Enzymes by Phlebia radiata. Phytochemistry 1991, 30, 2869–2872. [Google Scholar] [CrossRef]
- Piscitelli, A.; Giardina, P.; Lettera, V.; Pezzella, C.; Sannia, G.; Faraco, V. Induction and Transcriptional Regulation of Laccases in Fungi. Curr. Genom. 2011, 12, 104–112. [Google Scholar] [CrossRef]
- Wang, F.; Xu, L.; Zhao, L.; Ding, Z.; Ma, H.; Terry, N. Fungal Laccase Production from Lignocellulosic Agricultural Wastes by Solid-State Fermentation: A Review. Microorganisms 2019, 7, 665. [Google Scholar] [CrossRef]
- Pinheiro, V.E.; Michelin, M.; Vici, A.C.; de Almeida, P.Z.; Teixeira de Moraes Polizeli, M.d.L. Trametes versicolor Laccase Production Using Agricultural Wastes: A Comparative Study in Erlenmeyer Flasks, Bioreactor and Tray. Bioprocess Biosyst. Eng. 2020, 43, 507–514. [Google Scholar] [CrossRef]
- Kajita, S.; Sugawara, S.; Miyazaki, Y.; Nakamura, M.; Katayama, Y.; Shishido, K.; Iimura, Y. Overproduction of Recombinant Laccase Using a Homologous Expression System in Coriolus versicolor. Appl. Microbiol. Biotechnol. 2004, 66, 194–199. [Google Scholar] [CrossRef]
- Kilaru, S.; Hoegger, P.J.; Majcherczyk, A.; Burns, C.; Shishido, K.; Bailey, A.; Foster, G.D.; Kües, U. Expression of Laccase Gene Lcc1 in Coprinopsis cinerea under Control of Various Basidiomycetous Promoters. Appl. Microbiol. Biotechnol. 2006, 71, 200–210. [Google Scholar] [CrossRef]
- Arimoto, M.; Yamagishi, K.; Wang, J.; Tanaka, K.; Miyoshi, T.; Kamei, I.; Kondo, R.; Mori, T.; Kawagishi, H.; Hirai, H. Molecular Breeding of Lignin-Degrading Brown-Rot Fungus Gloeophyllum trabeum by Homologous Expression of Laccase Gene. AMB Express 2015, 5, 81. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.M.C.R.; Record, E.; Lomascolo, A.; Scholtmeijer, K.; Asther, M.; Wessels, J.G.H.; Wösten, M.A.B. Highly Efficient Production of Laccase by the Basidiomycete Pycnoporus cinnabarinus. Appl. Environ. Microbiol. 2004, 70, 6379–6384. [Google Scholar] [CrossRef] [PubMed]
- Salony; Garg, N.; Baranwal, R.; Chhabra, M.; Mishra, S.; Chaudhuri, T.K.; Bisaria, V.S. Laccase of Cyathus Bulleri: Structural, Catalytic Characterization and Expression in Escherichia Coli. Biochim. Biophys. Acta Proteins Proteomics 2008, 1784, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Lao, H.I.; Au, S.W.; Li, I.C.; Hu, J.; Yuen, H.M.; Cheong, W.M.; Lo, O.L.I.; Seak, L.C.U. Expression, Secretion and Functional Characterization of Three Laccases in E. Coli. Synth. Syst. Biotechnol. 2022, 7, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Sahdev, S.; Khattar, S.K.; Saini, K.S. Production of Active Eukaryotic Proteins through Bacterial Expression Systems: A Review of the Existing Biotechnology Strategies. Mol. Cell. Biochem. 2008, 307, 249–264. [Google Scholar] [CrossRef]
- Piscitelli, A.; Pezzella, C.; Giardina, P.; Faraco, V.; Sannia, G. Heterologous Laccase Production and Its Role in Industrial Applications. Bioeng. Bugs 2010, 1, 252–262. [Google Scholar] [CrossRef]
- Deshpande, N.; Wilkins, M.R.; Packer, N.; Nevalainen, H. Protein Glycosylation Pathways in Filamentous Fungi. Glycobiology 2008, 18, 626–637. [Google Scholar] [CrossRef]
- Record, E.; Punt, P.J.; Chamkha, M.; Labat, M.; Van Den Hondel, C.A.M.J.J.; Asther, M. Expression of the Pycnoporus cinnabarinus Laccase Gene in Aspergillus niger and Characterization of the Recombinant Enzyme. Eur. J. Biochem. 2002, 269, 602–609. [Google Scholar] [CrossRef]
- Otterbein, L.; Record, E.; Longhi, S.; Asther, M.; Moukha, S. Molecular Cloning of the CDNA Encoding Laccase from Pycnoporus cinnabarinus I-937 and Expression in Pichia pastoris. Eur. J. Biochem. 2000, 267, 1619–1625. [Google Scholar] [CrossRef]
- Sigoillot, C.; Record, E.; Belle, V.; Robert, J.L.; Levasseur, A.; Punt, P.J.; Van Den Hondel, C.A.M.J.J.; Fournel, A.; Sigoillot, J.C.; Asther, M. Natural and Recombinant Fungal Laccases for Paper Pulp Bleaching. Appl. Microbiol. Biotechnol. 2004, 64, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Mekmouche, Y.; Zhou, S.; Cusano, A.M.; Record, E.; Lomascolo, A.; Robert, V.; Simaan, A.J.; Rousselot-Pailley, P.; Ullah, S.; Chaspoul, F.; et al. Gram-Scale Production of a Basidiomycetous Laccase in Aspergillus niger. J. Biosci. Bioeng. 2014, 117, 25–27. [Google Scholar] [CrossRef] [PubMed]
- Kiiskinen, L.L.; Kruus, K.; Bailey, M.; Ylösmäki, E.; Siika-aho, M.; Saloheimo, M. Expression of Melanocarpus albomyces Laccase in Trichoderma reesei and Characterization of the Purified Enzyme. Microbiology 2004, 150, 3065–3074. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.J.O.; White, T.C. Expression of Laccase I and IV Genes from Trametes versicolor in Trichoderma reesei. ACS Symp. Ser. 2001, 785, 413–425. [Google Scholar] [CrossRef]
- Saloheimo, M.; Niku-Paavola, M.L.; Knowles, J.K.C. Isolation and Structural Analysis of the Laccase Gene from the Lignin-Degrading Fungus Phlebia radiata. J. Gen. Microbiol. 1991, 137, 1537–1544. [Google Scholar] [CrossRef] [PubMed]
- Pourmir, A.; Johannes, T.W. Directed Evolution: Selection of the Host Organism. Comput. Struct. Biotechnol. J. 2012, 2, e201209012. [Google Scholar] [CrossRef] [PubMed]
- Gellissen, G.; Hollenberg, C.P. Application of Yeasts in Gene Expression Studies: A Comparison of Saccharomyces cerevisiae, Hansenula Polymorpha and Kluyveromyces lactis—A Review. Gene 1997, 190, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Mai, J.; Ding, Y.; Wei, Y.; Ledesma-Amaro, R.; Ji, X.J. Improving the homologous recombination efficiency of Yarrowia lipolytica by grafting heterologous component from Saccharomyces cerevisiae. Metab. Eng. Commun. 2020, 11, e00152. [Google Scholar] [CrossRef]
- Alcalde, M.; Zumarraga, M.; Polaina, J.; Ballesteros, A.; Plou, F. Combinatorial Saturation Mutagenesis by In Vivo Overlap Extension for the Engineering of Fungal Laccases. Comb. Chem. High Throughput Screen. 2006, 9, 719–727. [Google Scholar] [CrossRef][Green Version]
- Zumarraga, M.; Dominguez, C.; Camarero, S.; Shleev, S.; Polaina, J.; Martinez-Arias, A.; Ferrer, M.; De Lacey, A.; Fernandez, V.; Ballesteros, A.; et al. Combinatorial Saturation Mutagenesis of the Myceliophthora thermophila Laccase T2 Mutant: The Connection between the C-Terminal Plug and the Conserved VSG Tripeptide. Comb. Chem. High Throughput Screen. 2008, 11, 807–816. [Google Scholar] [CrossRef]
- Cereghino, J.L.; Cregg, J.M. Heterologous Protein Expression in the Methylotrophic Yeast Pichia pastoris. FEMS Microbiol. Rev. 2000, 24, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Colao, M.C.; Lupino, S.; Garzillo, A.M.; Buonocore, V.; Ruzzi, M. Heterologous Expression of Lcc1 Gene from Trametes trogii in Pichia pastoris and Characterization of the Recombinant Enzyme. Microb. Cell Fact. 2006, 5, 31. [Google Scholar] [CrossRef]
- Zhou, H.M.; Honh, Y.Z.; Xiao, Y.Z.; Cui, T.J.; Wang, X.T.; Pu, C.L. High Output of a Trametes Laccase in Pichia pastoris and Characterization of Recombinant Enzymes. Chin. J. Biotechnol. 2007, 23, 1055–1059. [Google Scholar] [CrossRef]
- Kittl, R.; Gonaus, C.; Pillei, C.; Haltrich, D.; Ludwig, R. Constitutive Expression of Botrytis aclada Laccase in Pichia pastoris. Bioengineered 2012, 3, 232–235. [Google Scholar] [CrossRef]
- Hong, F.; Meinander, N.Q.; Jönsson, L.J. Fermentation Strategies for Improved Heterologous Expression of Laccase in Pichia pastoris. Biotechnol. Bioeng. 2002, 79, 438–449. [Google Scholar] [CrossRef]
- Mate, D.M.; Gonzalez-Perez, D.; Kittl, R.; Ludwig, R.; Alcalde, M. Functional Expression of a Blood Tolerant Laccase in Pichia pastoris. BMC Biotechnol. 2013, 13, 38. [Google Scholar] [CrossRef] [PubMed]
- Owji, H.; Nezafat, N.; Negahdaripour, M.; Hajiebrahimi, A.; Ghasemi, Y. A Comprehensive Review of Signal Peptides: Structure, Roles, and Applications. Eur. J. Cell Biol. 2018, 97, 422–441. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Ohira, T.; Igarashi, K.; Nagasawa, H.; Aida, K.; Hallberg, B.M.; Divne, C.; Nishino, T.; Samejima, M. Production and Characterization of Recombinant Phanerochaete Chrysosporium Cellobiose Dehydrogenase in the Methylotrophic Yeast Pichia pastoris. Biosci. Biotechnol. Biochem. 2001, 65, 2050–2057. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Anumanthan, A.; Gao, X.G.; Ilangovan, K.; Suzara, V.V.; Düzgüneş, N.; Renugopalakrishnan, V. Expression of Recombinant Proteins in Pichia pastoris. Appl. Biochem. Biotechnol. 2007, 142, 105–124. [Google Scholar] [CrossRef]
- Aza, P.; Molpeceres, G.; de Salas, F.; Camarero, S. Design of an Improved Universal Signal Peptide Based on the α-Factor Mating Secretion Signal for Enzyme Production in Yeast. Cell. Mol. Life Sci. 2021, 78, 3691–3707. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aza, P.; Camarero, S. Fungal Laccases: Fundamentals, Engineering and Classification Update. Biomolecules 2023, 13, 1716. https://doi.org/10.3390/biom13121716
Aza P, Camarero S. Fungal Laccases: Fundamentals, Engineering and Classification Update. Biomolecules. 2023; 13(12):1716. https://doi.org/10.3390/biom13121716
Chicago/Turabian StyleAza, Pablo, and Susana Camarero. 2023. "Fungal Laccases: Fundamentals, Engineering and Classification Update" Biomolecules 13, no. 12: 1716. https://doi.org/10.3390/biom13121716
APA StyleAza, P., & Camarero, S. (2023). Fungal Laccases: Fundamentals, Engineering and Classification Update. Biomolecules, 13(12), 1716. https://doi.org/10.3390/biom13121716