Abstract
Sugar transporters play an important role in the cellulase production of lignocellulose-degrading fungi. Nevertheless, the role and function of these transporters are still unclear. Here we first report intracellular sugar transporters assisting cellulase production in Trichoderma reesei (T. reesei) using lactose. The mRNA levels of sugar transporter genes mfs, gst, and lac1 were substantially upregulated in T. reesei cultivated on lactose, with the most abundant mRNA levels at 24 h as compared to glucose. Moreover, the individual deletion of these sugar transporters significantly inhibited cellulase production, solid cell growth, and sporulation of T. reesei, suggesting they play a supporting role in cellulase production when grown in lactose. Surprisingly, MFS, GST, and LAC1 were mainly localized in the cytoplasm, with MFS and LAC1 in the endoplasmic reticulum (ER), representing the first discovery of intracellular sugar transporters involved in cellulase biosynthesis in lactose culture. The absence of the gene lac1 noticeably inhibited most of the crucial genes related to cellulase production, including cellulase-encoding genes, transcription factors, and sugar transporters, at 24 h, which was fully relieved at 48 h or 72 h, indicating that lac1 affects cellulase production more at the early step. This research advances the understanding of the function of intracellular sugar transporters in fungi, particularly for fungal cellulase production.
1. Introduction
Trichoderma reesei (T. reesei) decays wood in nature, metabolizing the cellulose and hemicellulose of the plant biomass via cellulases and hemicellulases. Cellulase is a complicated, extracellular enzyme mixture, majorly comprised of three kinds of synergistic enzymes for cellulose degradation: endoglucanase (EG, EC3.2.1.4), exoglucanase (or cellobiohydrolases, CBH, EC 3.2.1.91), and β-glucosidase (BGL, EC 3.2.1.21) [1]. Cellulase is produced by microbial fermentation for applications in the biofuel and biorefinery industries using T. reesei RUT-C30 as the working horse. Lactose, the byproduct from the whey processing and cheese manufacturing industries, is inexpensive and economically feasible. Despite the fact that it is not as efficient as cellulose in cellulase induction, lactose is the only major soluble carbon source for cellulolytic enzyme fermentation by T. reesei on an industrial level [2]. Nevertheless, the mechanism behind the induction of cellulase by lactose is not understood, limiting the metabolic engineering efforts for improved cellulase induction efficiency of lactose.
Sugar transporters have crucial functions in sensing and transporting diverse sugars for cellulase production in lignocellulose-degrading fungi, such as Crt1 and Stp1 in T. reesei [3,4,5], cellodextrin transporters CDT-1, CDT-2, and CLP1 in Neurospora crassa [6,7], cellobiose transporters cltA and cltB in Aspergillus nidulans (A. nidulans) [8], and cellodextrin transporters CdtC, CdtD, and CdtG in Penicillium oxalicum [9]. In T. reesei, there is a large array of sugar transporters that are capable of effectively transporting the constituent sugars into the cell, enabling its access to sugars. In contrast to that, Saccharomyces cerevisiae has only 43 sugar transporters, while T. reesei has 113 putative sugar transporters in its in silico proteome [10]. Recently, intracellular sugar transporters have been reported for cellulase production in T. reesei on cellulose [11,12].
It is assumed that lactose can be hydrolyzed extracellularly by β-galactosidase (lactase, β-D-galactosidase galactohydrolase; E.C 3.2.1.23) to D-glucose and D-galactose, that are then taken up by cells through the respective permeases [13]. Alternatively, lactose itself can be taken up by specific lactose permeases and hydrolyzed intracellularly. The cellular uptake of these sugars is crucial for cell growth and is intricately linked to cellulase and hemicellulase production. Accordingly, sugar transporters are found to play a critical role in cellulase production using lactose. Transporters for cellular internalization of lactose were induced by cellulose-derived sugars [14]. Meanwhile, several key sugar transporters have been identified as involved in cellulase production from lactose. For instance, the lactose transporter Crt1 (Trire2: 3405) appears to be essential for growth on lactose, and its deletion prevents lactose-induced cellulase gene expression in T. reesei [3,5,15], which is proved to exist in the plasma membrane and the periphery of the nucleus [11]. A screening for lactose permeases in T. reesei PC-3-7 was performed, leading to the identification of two MFS sugar transporters (TR79202 and TR77517) whose deletion decreased lactose uptake, cell growth, and cellulase production on lactose with no significant effect on cellulase production in cellulose culture [4]. This demonstrated that the two identified sugar transporters are important for cellulase generation when using lactose as the carbon source. A lactose permease gene (lacpA) was found in another cellulase-producing fungus, A. nidulans, whose deletion impaired lactose uptake and cell growth [16]. Despite the great importance of sugar transporters in cellulase production, the molecular mechanism behind the function and role of sugar transporters in cellulase production remained undetermined.
Here, the function of three sugar transporters, MFS (M419DRAFT_137795), GST (M419DRAFT_138519), and LAC1 (M419DRAFT_127980), was investigated in cellulase production using lactose as the carbon source. We measured the mRNA levels of these sugar transporters and assayed their effect on cellulase activities by knocking them out in T. reesei, finding that they facilitate cellulase production. Furthermore, we investigated the impact of lac1 on the transcriptional levels of cellulase-related genes like cellulase-encoding genes, transcription factors, other sugar transporters, and the lactose metabolism enzyme, revealing that the function of lac1 in cellulase expression acted at the transcriptional levels at an early stage of cellulase production, which was mediated by the well-known cellulase regulators CRT1 and XYR1. Finally, we analyzed the cellular distribution of sugar transporters MFS, GST, and LAC1 by C-terminally tagging them with the red fluorescent protein DsRed, unraveling that they are localized in the cytoplasm, especially in the endoplasmic reticulum (ER). This study might offer new strategies to improve fungal cellulase production by engineering intracellular sugar transporters.
2. Materials and Methods
2.1. Strains and Cultivation Conditions
Escherichia coli (E. coli) DH5α was utilized to construct plasmids, and Agrobacterium tumefaciens (A. tumefaciens) AGL-1 was used for fungal transformation. Strain T. reesei Rut-C30 (CICC 13052, ATCC 56765) was used as a parental strain for gene overexpression, and strain Δku70 was used as a parental strain for gene knockout and in situ fluorescence protein tagging [17,18]. T. reesei Δku70, a derivate of T. reesei Rut-C30, created by deleting the gene ku70, was kindly provided by Professor Wei Wang from East China University of Science and Technology [19]. E. coli DH5α and A. tumefaciens were grown on Luria-Bertani medium at 37 °C and 28 °C, respectively. T. reesei were cultured in Sabouraud Dextrose Broth (SDB) and Trichoderma minimal media (TMM) [20] containing 2% lactose as the sole carbon source at 28 °C for the analysis of cellulase production and gene regulation, and cultured on potato dextrose agar (PDA) plates and TMM solid plates with lactose at 28 °C for the analysis of cell growth and sporulation ability.
2.2. Construction of Recombinant T. reesei Strains
To knockout genes mfs, gst, and lac1, the 1.5-kilobase (KB) upstream and downstream fragments of these genes were amplified from the genomic DNA of Δku70 and then integrated into plasmid pXBthg, resulting in plasmids pXBthg-mfs, pXBthg-gst, and pXBthg-lac1. The recombinant plasmids were then transformed into parental strain Δku70, respectively, through the Agrobacterium-mediated transformation (AMT) method [21] to obtain the recombinant strains Δmfs, Δgst, and Δlac1.
For linking a red fluorescent protein DsRed at the C-terminus of the transporter to track its intracellular localization, the upstream (including the promoter and coding sequence) and downstream (including the terminator) fragments of mfs, gst, and lac1 amplified from the genomic DNA of Δku70 were ligated to the plasmid pXBred [22], respectively. These plasmids were transformed into the parental strain Δku70 by the AMT method, leading to the recombinant strains MFS-DsRed, GST-DsRed, and LAC1-DsRed, respectively. In addition, the fragment of gene lac1 was amplified from the genomic DNA of Rut-C30 and inserted into plasmid p-DsRed [22] at the XbaI site under a modified cbh1 promoter, leading to plasmid lac1-pDmred. Plasmid lac1-pDmred was transformed into parental strain Rut-C30 using the AMT method, generating the recombinant strain LAC1-DsRed-OE. All the gene modifications in the recombinant strains were confirmed by polymerase chain reaction (PCR) and sequencing of the PCR products at Sangon Biotech. The primers used for amplification and verification are listed in Table S1 (supplementary material).
2.3. The Shake Flask Culture of T. reesei
An amount of 0.5 mL of T. reesei conidia (107/mL) was seeded into 10 mL SDB and grown for 48 h at 28 °C with 200 rpm. Following this, 5 mL pre-grown mycelia (the wet weight of mycelia was about 0.6 g) were inoculated into a 250 mL conical flask containing 50 mL TMM media with 2% lactose, and cultured for 72 h at 28°C with 200 rpm. Samples were collected at different time points for cellulase and hemicellulase activity assays [22,23,24,25,26], confocal observation, and quantitative reverse transcription PCR (qRT-PCR) analysis. For analyzing the cell growth in liquid culture medium, 1 mL of a fungal culture solution containing diffuse mycelia was collected at 24 h, 48 h, and 72 h and diluted appropriately, and then the absorption value at 600 nm wavelength of the diluent was measured using an ultraviolet spectrophotometer (UV-2600, Shimadzu, Japan). In order to measure the growth ability of the recombinant strains on the solid medium, 10 μL of fresh spore solution was dropped into the center of the solid TMM plates. Then, the colony diameter was measured at 48 and 72 h, respectively.
2.4. qRT-PCR Analysis
Fresh mycelia were preserved at the indicated time points. Total RNA was extracted from mycelia cells using the Fungi RNA Kit (Omega Bio-Tek, Inc., USA, R6840) and then reverse transcribed into cDNA using PrimeScript™ RT reagent Kit with gDNA Eraser (Takara, Dalian, China, RR047Q). qRT-PCR was performed in an ABI 7500 real-time PCR system, following the protocol of TB Green Premix Ex Taq™ II (Takara, Dalian, China, RR820Q). The relative mRNA level of each tested gene was normalized to the housekeeping gene sar1 [27,28] and was calculated by the 2−ΔCt method, where ΔCt = Ct(test) − Ct(sar1). All the primers used are described in Table S1 (supplementary material). Three biological replicates were performed, with three technical replicates for each biological replicate.
2.5. Confocal Imaging
Confocal images of T. reesei strains DsRed [29], MFS-DsRed, GST-DsRed, LAC1-DsRed, and LAC1-DsRed-OE cultured on lactose were taken using a confocal laser scanning microscopy (CLSM) TCS SP8 with a 100× oil immersion objective. The 552 nm laser was selected to excite the samples, and the fluorescence emission was detected between 570 and 700 nm. T. reesei mycelium was collected at different time points through centrifuging at 8000 rpm for 15 min. For labeling ER, T. reesei mycelium was washed twice with Hank’s Balanced Salt Solution (HBSS) buffer and resuspended in 1 mL HBSS. Then ER-Green (KeyGEN BioTECH Co., Ltd., Jiangsu, China) was added into the suspension at a ratio of 1:1000 and incubated at 37 °C for 20 min. After incubation, the culture was washed twice with HBSS and resuspended in HBSS. Then, the images of stained samples were taken with an excitation wavelength of 488 nm and an emission wavelength range of 500–550 nm. For dying cell membranes, T. reesei mycelium was washed with Phosphate Buffered Saline (PBS) and then resuspended in staining solution GC-PEG-cholesterol-FITC and incubated in the dark at 37 °C for 10 min [22,30,31]. After incubation, the culture was washed with PBS twice and resuspended in PBS. The images of samples stained with cell membrane dye were taken with an excitation wavelength of 488 nm and an emission wavelength range of 500–550 nm.
2.6. Bioinformatic Analysis
To obtain the transmembrane domains of these sugar transporters, the amino acid sequences of these sugar transporters were downloaded from the EnsemblFungi database and then submitted to the web server Protter, respectively. To find the ER localization signal motif of sugar transporters, the amino acid sequences of these sugar transporters were submitted to the CLUSTALW online server, and the outcomes were then submitted to the ESPript online server.
3. Results
3.1. The Transcription Dynamics of Sugar Transporters MFS, GST, and LAC1 during the Cellulase Production on Lactose
To explore the role of sugar transporters MFS, GST, and LAC1 on cellulase generation, we measured the mRNA levels of these three genes during the whole process of cellulase production on lactose (cellulase-producing condition) by qRT-PCR in comparison to glucose (no cellulase-producing condition) (Figure 1). The transcriptional abundances of these sugar transporters on lactose were notably higher than those on glucose, similar to that of the well-known cellulase-essential sugar transporter gene crt1(M419DRAFT_109243). Particularly, the relative mRNA levels of mfs, gst, and lac1 were 8.81, 0.33, and 0.01 in T. reesei on lactose for 24 h, which were 11.40, 54.90, and 17.11 times that on glucose. Moreover, gene mfs possessed the highest mRNA abundance, followed by gst and lac1 in a decreasing order, but was lower than that of crt1. This remarkable upregulation in the gene expression of mfs, gst, and lac1 under lactose conditions is indicative of their importance for cellulase production when using lactose as the carbon source.
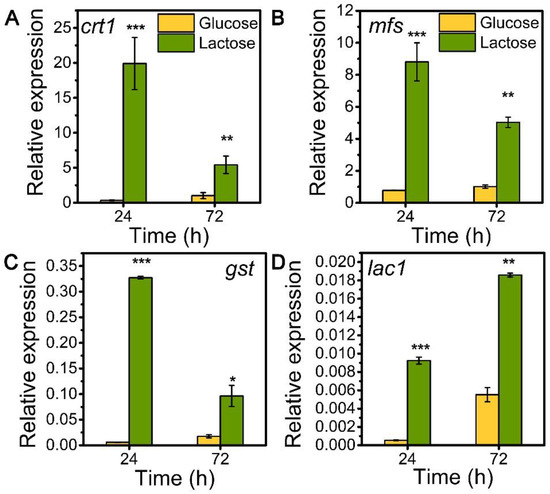
Figure 1.
Transcription levels of sugar transporter genes (A) crt1, (B) mfs, (C) gst, and (D) lac1 in T. reesei Rut-C30 cultured on TMM+2% lactose/glucose at 24 h and 72 h. Values are the means of three biological replicates, and error bars are the standard deviations of these three replicates. The gene expression level was calculated using the 2−ΔCt method. The relative mRNA level of each tested gene was normalized to the housekeeping gene sar1. * p < 0.05, ** p < 0.01, and *** p < 0.001.
3.2. Sugar Transporters MFS, GST, and LAC1 Facilitated Cellulase and Hemicellulase Production in T. reesei Grown on Lactose
Inspired by their notable transcriptional upregulation under lactose conditions, we individually knockout genes encoding putative sugar transporters MFS, GST, and LAC, in the parental strain Δku70 by homologous recombination, yielding strains Δmfs, Δgst, and Δlac1, respectively. The enzyme-producing ability of these deletion strains was evaluated by measuring cellulase and hemicellulase activities in their culture supernatant at varied time points using lactose as the carbon source, as compared to the starting strain Δku70 (Figure 2). The single deletion of sugar transporters MFS, GST, and LAC1, substantially inhibited the cellulase and hemicellulase production in T. reesei grown on lactose. When grown on lactose for 72 h, strain Δlac1 exhibited 1.25 IU/mL FPase activity, 4.02 IU/mL CMCase activity, 0.14 IU/mL pNPCase activity, 0.59 IU/mL pNPGase activity, and 0.11 IU/mL pNPxase activity, and a 3.23 mg/mL secreted protein concentration, only 30.54%, 66.81%, 31.24%, 63.68%, 43.44%, and 59.06% of that in strain Δku70, respectively. The other two knockout strains showed similar reduction degree to strain Δlac1. Clearly, the sugar transporters MFS, GST, and LAC1 are crucial for cellulase and hemicellulase production in T. reesei.
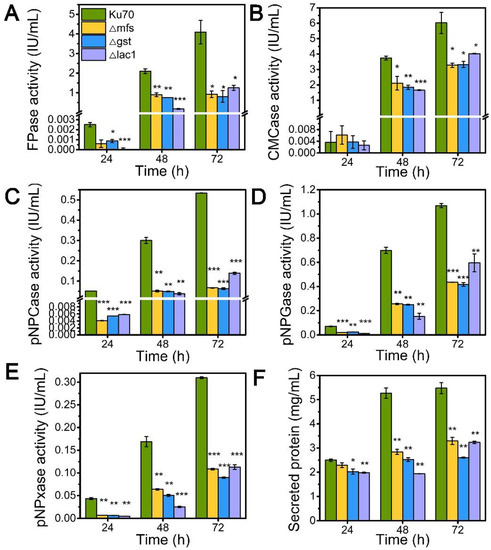
Figure 2.
The cellulase and hemicellulase activities and protein secretion of T. reesei Δku70, Δmfs, Δgst, and Δlac1 cultured in TMM+2% lactose at 24 h, 48 h, and 72 h, respectively. (A) FPase: the filter paper activity; (B) CMCase: the CMC activity; (C) pNPCase: the CBH activity; (D) pNPGase: the BGL activity; (E) pNPxase: the xylanse activity; (F) protein secretion. Values are the means of three biological replicates, and error bars are the standard deviations of these three replicates. Asterisks indicate significant differences (* p < 0.05, ** p < 0.01, and *** p < 0.001) as assessed by the Student’s t test.
3.3. The Knockout of Sugar Transporters MFS, GST, and LAC1 Reduced the Cell Growth and Spore Ability of T. reesei Cultivated on Lactose
When being cultured in TMM+2% lactose, the cell growth of strains Δmfs and Δgst was noticeably reduced at 24 h and 48 h but increased at 72 h, while strain Δlac1 showed decreased cell growth at all tested time points as compared to Δku70 (Figure 3A). When grown on TMM+2% lactose agar plates, the three deletion strains Δmfs, Δgst, and Δlac1 showed much smaller colony diameters than Δku70, only 86.7%, 84.9%, and 56.7% of that Δku70 at 72 h (Figure 3B,C). The single deletion of these transporter genes, mfs (Trire2: 79202), gst (Trire2: 50894), or lac1 (Trire2: 56684), in QM9414 also led to growth reduction using lactose as the carbon source [3]. Meanwhile, all three knockout strains showed a significantly reduced number of spores in comparison to Δku70 (Figure 3B,D). These data demonstrate that the sugar transporters MFS, GST, and LAC1 play a positive role in both cell growth and sporulation with T. reesei together in cellulase biosynthesis.
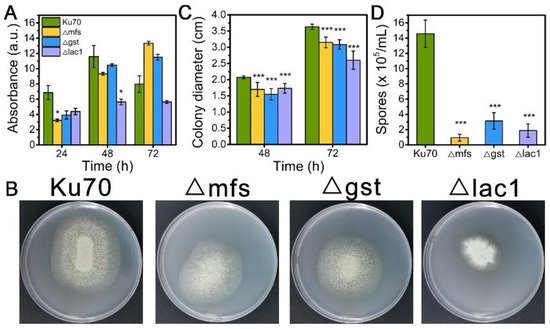
Figure 3.
(A) The growth, (B) plate growth pictures, (C) colony diameters, and (D) sporulation of T. reesei Ku70, Δmfs, Δgst, and Δlac1 grown on TMM with 2% lactose. Both the spore assay and the plate growth pictures were performed at 72 h. Values are the means of three biological replicates, and error bars are the standard deviations of these three replicates. Asterisks indicate significant differences (* p < 0.05, and *** p < 0.001) as assessed by the Student’s t test.
3.4. The Impact of lac1 Knockout on the mRNA Levels of Crucial Genes Involved in Cellulase Production
Though the decline of enzyme activities of these three constructed strains Δmfs, Δgst, and Δlac1, was similar, the growth of strain Δlac1 was more significantly repressed when cultured on lactose. Hence, we chose strain mutant Δlac1 to perform qRT-PCR analysis. We monitored the mRNA dynamics of key genes associated with (hemi)cellulase expression in strain Δlac1 cultivated on lactose, including genes encoding cellulase, which include β-glucosidase BGL1 (cel3a), the major cellobiohydrolase CBHI (cel7a), the major endoglucanase CMC (cel7b), hemicellulse β-xylosidase (bxl1), the well-known transcription factors xyr1 [32], ace3 [33], clr2 [34], clr3 [35,36], and xpp1 [37], sugar transporters crt1 and stp1 [5], and gene xyl1 in the lactose metabolism. Except genes xpp1 and stp1, which stayed unchanged, all the assayed genes were notably downregulated in strain Δlac1 at 24 h, as compared to Δku70. Nevertheless, the mRNA levels of these genes were comparable to or even higher than those in Δku70 at 48 h and/or 72 h (Figure 4). The transcriptional abundance of xpp1 and stp1 stayed almost constant in the first 48 h, but was noticeably reduced at 72 h.
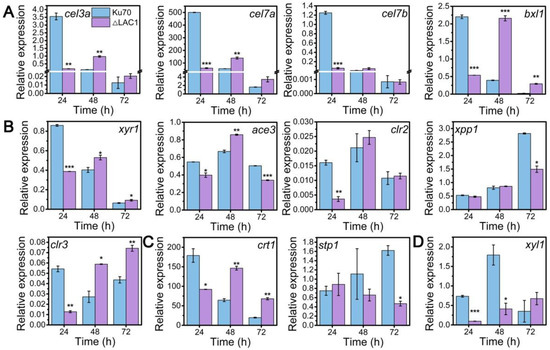
Figure 4.
qRT-PCR analysis of the transcript abundance of genes relevant to cellulase and hemicellulase production in T. reesei Δku70 and Δlac1 grown on 2% lactose for 72 h including genes involved in (A) cellulase and hemicellulase production, (B) transcription factors, (C) sugar transportation, and (D) lactose metabolism. Values are the means of three biological replicates, and error bars are the standard deviations of these three replicates. The gene expression level was calculated using the 2−ΔCt method. The relative mRNA level of each tested gene was normalized to the housekeeping gene sar1. * p < 0.05, ** p < 0.01, and *** p < 0.001.
3.5. Cellular Distribution of Sugar Transporters MFS, GST, and LAC1
Structure analysis showed that these transporters are transmembrane proteins belonging to MFSs and displaying characteristics of sugar transporters. There are 12 transmembrane domains in GST and 11 in MFS and LAC1 (Figure S1). Nevertheless, the cellular distribution of these sugar transporters is still unknown. To investigate the cellular distribution of sugar transporters MFS, GST, and LAC1, we tagged the fluorescence protein DsRed at their C termini, by integrating the DsRed gene sequence at their 3′ end through homologous recombination, obtaining recombinant strains MFS-DsRed, GST-DsRed, and LAC1-DsRed expressing the fusion proteins MFS-DsRed, GST-DsRed, and LAC1-DsRed. CLSM was utilized to track these red-emitting fusion proteins (Figure 5A and Figure S2). Strain MFS-DsRed started to show weak red fluorescence in the cytoplasm at 24 h, which became much stronger at 48 h and 72 h. At 48 h, the protein MFS-DsRed was dominantly localized to characteristic perinuclear ER rings, exhibiting bright red fluorescence. Nevertheless, red fluorescence was also found on the cell membrane, although it was not that significant, indicating that MFS-DsRed can also distribute to the cell membrane. No yellow fluorescence was found in the strain MFS-DsRed co-labelled with a green cell wall dye, GC-PEG-cholesterol-FITC (Figure S3), implying that MFS were on the cell membrane but not the cell wall. The red fluorescence in strain GST-DsRed was only observed at 72 h but not at 24 or 48 h, while no fluorescence was found in strain LAC1-DsRed at all studied time points. Unlike MFS-DsRed which was localized in perinuclear ER rings, dense red fluorescent punctuates were observed in DsRed. At 72 h, the fluorescence intensity in strain MFS-DsRed was the highest, followed by that in strains GST-DsRed and LAC1-DsRed, which was consistent with their transcriptional abundance order.
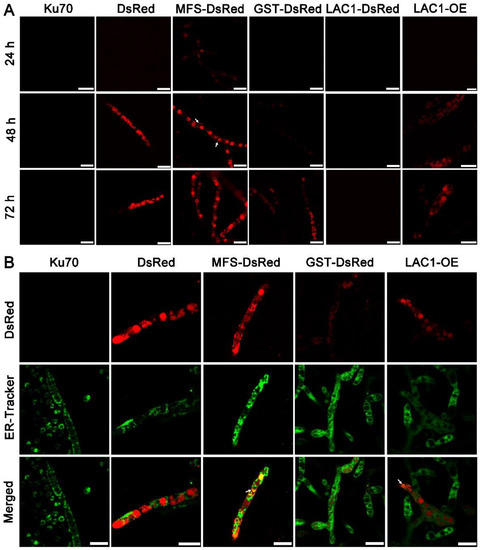
Figure 5.
(A) Cellular localization of the three sugar transporters in T. reesei cultured in TMM + 2% lactose at 24 h, 48 h, and 72 h, respectively. The white arrow indicates the localization of MFS-DsRed on the cell membrane. (B) Confocal images of strains Δku70, DsRed, MFS-DsRed, GST-DsRed, LAC1-DsRed, and LAC1-DsRed-OE (LAC1-OE) with the treatment of ER-Tracker (green). Strains MFS-DsRed and GST-DsRed were constructed under their own endogenous promoters, and strain LAC1-DsRed-OE was constructed under the modified strong promoter cbh1.The white arrow points to the overlap of the green ER dye with the red fluorescent protein fusion LAC1-DsRed-OE. Scale bar = 10 μm.
To visualize the protein LAC1 in T. reesei, the overexpression of LAC1-DsRed under the strong promoter cbh1 was carried out, leading to the recombinant strain LAC1-DsRed-OE. Strain LAC1-DsRed-OE displayed red fluorescence in perinuclear ER rings and vacuoles, similar to strain MFS-DsRed. This suggests that LAC1 was distributed in perinuclear ER rings and vacuoles in T. reesei grown on lactose. Moreover, the red fluorescence at the apical regions of the recombinant strains MFS-DsRed, GST-DsRed, and LAC1-DsRed-OE was not stronger than that at the subapical areas, showing that all three sugar transporters were not accumulated at the apical regions (Figure S4).
To check whether the sugar transporters are ER-targeted, mutant strains DsRed, MFS-DsRed, GST-DsRed, and LAC1-DsRed-OE were labeled with a green-emissive ER-tracker (Figure 5B). The red fluorescence of MFS-DsRed and LAC1-DsRed-OE was overlapped with the green fluorescence of the ER tracker, leading to the yellow fluorescence in the merged figure, showing that MFS-DsRed and LAC1-DsRed-OE were localized in ER. In contrast, no yellow fluorescence was found in the merged picture derived from the pictures of GST-DsRed and ER tracker, indicating that GST-DsRed was not distributed in the ER. Moreover, no yellow fluorescence was found in strain DsRed, indicating that the fluorescence protein DsRed alone was not distributed in the ER. Bioinformatics analysis showed that there was an ER localization signal KKXX (Lys-Lys-X-X) [38,39] in the amino acid sequence of LAC1 (Figure S5). However, interestingly, there was no such ER localization signal sequence was found in transporters MFS and GST (Figure S5).
4. Discussion and Conclusions
Currently, much of the research efforts on cellulase production using lactose as the carbon source has focused on the lactose metabolism [1,40,41,42,43,44,45]. On the contrary, the importance of sugar transporters on cellulase formation by lactose has not received widespread interest, although some efforts have been made. For instance, Ivanova et al. performed a system analysis of the T. reesei transcriptome when using lactose as the carbon source and identified a lactose permease that is essential for cellulase induction [3]. Herein, the function and cellular distribution of three predicted sugar transporters (MFS, GST, and LAC1) were investigated under cellulase-generating conditions when using lactose as the carbon source. First, the mRNA levels of these three sugar transporters were much higher in Rut-C30 grown on lactose than those on glucose. Furthermore, cellulase production was much lower in the deletion strains Δmfs, Δgst, and Δlac1 than in the parental strain Ku70. Both these results demonstrate that sugar transporters MFS, GST, and LAC1 facilitate cellulase production using lactose (Figure 6). Several putative lactose permeases have been found to work in cellulase production in lignocellulosic fungi, such as the proposed lactose transporter Crt1 [3,5,15], two MFS sugar transporters (TR79202 and TR77517) [4] in T. reesei, and the lactose permease gene (lacpA) in A. nidulans [16]. Just like MFS, GST, and LAC1, cellulose response transporter 1 (CRT1) belongs to the MFS transporter and is initially considered a lactose permease. Crt1 is required for cellulase biosynthesis on lactose, cellulose, or sophorose, and is highly expressed in T. reesei cultivated in lactose [4], cellulose [4], and wheat straw [46]. Different from, where the single knockout of crt1 was enough to fully repress cellulase formation in T. reesei cultivated on lactose [3,5], the sole deletion of mfs, gst, and lac1 severely but not completely repressed cellulose production in T. reesei, showing that these sugar transporters play a supporting rather than an essential role in cellulase production. Another possible explanation for this might be that there are other functionally equivalent sugar transporters to MFS, GST, or LAC1 in T. reesei that can circumvent the loss of these sugar transporters and allow for cellulase production.
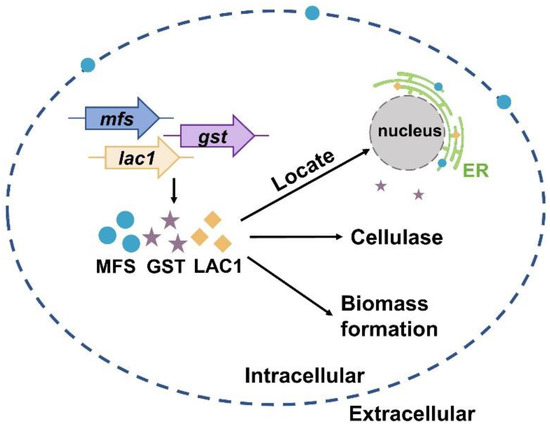
Figure 6.
A scheme of the function and cellular distribution of three intracellular sugar transporters in T. reesei when cultured in lactose.
The time-course analysis of the mRNA levels of critical genes for cellulase expression in strain Δlac1 cultured in lactose for cellulase generation revealed that the deletion of LAC1 notably reduced the transcriptional abundances of these genes except xpp1 and stp1 at 24 h. The severe reduction of transcriptional levels of cellulase-encoding genes cel3a, cel7a, and cel7b was consistent with the notable inhibition of cellulase activities (Figure 2B–D). Meanwhile, the mRNA levels of the well-known cellulase transcriptional activator XYR1 and sugar transporter CRT1, both of which are essential for cellulase formation, were significantly reduced. Nevertheless, this transcriptional suppression was recovered at 48 h and/or 72 h. It seems that LAC1 works at the transcriptional level at the early stage of cellulase production, which is crucial for the initiation of cellulase production with lactose.
More importantly, the cellular distribution analysis by CLSM showed that MFS, GST, and LAC1 were dominantly localized in the cytoplasm, though a minor fraction of MFS was sometimes was found on the cell membrane (Figure 6). They were distributed evenly throughout the whole hyphal, without accumulation in the apical regions. As far as we know, this is the first report on intracellular sugar transporters for cellulase production induced by lactose. Previous studies focused on the membrane-bound sugar transporters for cellulase production in lignocellulosic fungi, like Crt1 and Stp1 in T. reesei [5], cellodextrin transporters CDT-1, CDT-2, and CLP1 in Neurospora crassa [6,7], and cellobiose transporters cltA and cltB in Aspergillus nidulans [8]. The localization of MFS, GST, and LAC1 in cytoplasm and ER implies they may function in sugar sensing, transporting, and/or utilization inside the cells after its cellular uptake. Particularly, MFS and LAC1 were majorly distributed to ER (Figure 6), as indicated by a large number of perinuclear ER rings found in strains MFS-DsRed and LAC1-DsRED-OE (Figure 5A), and the red fluorescence of proteins MFS-DsRed and LAC1-DsRED-OE was overlapped with the green fluorescence of the ER tracker (Figure 5B). ER is a well-known place for the glycosylation of proteins and lipids, requiring a regular replenishment of cytosolically synthesized nucleotide sugars, which are transported by a group of dedicated nucleotide sugar transporters (NST) [47,48]. There is a possibility that MFS and LAC1 are involved in protein glycosylation in the ER, acting as nucleotide sugar transporters, endomembrane sugar transporters, or sugar transceptors. Similarly, these three sugar transporters were also localized intracellularly in T. reesei grown on cellulose [12]. Moreover, the well-known lactose permease Crt1, which is essential for cellulase production on lactose [15] and had been considered to be on the cell membrane for a long time, was recently found to be intracellular, mostly at the periphery of the nucleus [11]. Nevertheless, the detailed roles of these intracellular sugar transporters remain unknown, which is worth exploring in the future. There is a possibility that the localization of MFS and GST in ER is an artifact of stressful conditions, which we believe is probably not the case in our study. If the localization of MFS and GST in the ER was due to the ER stress, where ER is overloaded with misfolded proteins, it would probably cause the fluorescence quenching of DsRed. Then, no fluorescence would be observed in the ER of strains MFS-DsRed and LAC1-DsRed, which would be contrary to our result (Figure 5). In addition, through a literature search and amino acid sequence analysis, we found that some of the analyzed sugar transporters contained an ER localization signal (Figure S5). There is one ER localization signal sequence reported in the literature: KKXX (Lys-Lys-X-X) [38,39] which existed in LAC1. Interestingly, no ER localization signal sequence was observed in MFS or GST. Nevertheless, further experimentation is required to completely exclude this possibility. For example, protein expression can be performed under ER stress caused by adding drugs like tunicamycin. As the first-ever report on the discovery and function of intracellular sugar transporters for cellulase production in lignocellulosic fungi that we know of, this study deepens our understanding of fungal intracellular sugar transporters and cellulase production mechanisms.
In summary, we reported that sugar transporters MFS and LAC1 were localized in the ER, while GST was in the cytoplasm. They have a positive role in cellulase production and biomass formation in T. reesei when cultured in lactose (Figure 6).
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/biom13020295/s1, Figure S1: Bioinformatic analysis of transmembrane domains of sugar transporters MFS, GST, and LAC1 obtained from the webserver Protter (http://wlab.ethz.ch/protter/#). The green square represents N-glycosylation motif and red circle represents the signal peptide; Figure S2: Brightfield images of T. reesei strains Δku70, DsRed, MFS-DsRed, GST-DsRed, LAC1-DsRed, and LAC1-DsRed-OE (LAC1-OE) cultured in TMM+2% lactose at 24 h, 48 h, and 72 h, respectively. Scale bar = 10 μm; Figure S3: Confocal images of strain MFS-DsRed stained with GC-PEG-cholesterol-FITC, a green fluorescence dye for cell walls. Strain MFS-DsRed was grown on lactose for 72 h. Scale bar = 10 μm; Figure S4: Cellular location of MFS-DsRed, GST-DsRed, and LAC1-DsRed-OE at the apical regions of recombinant strains MFS-DsRed, GST-DsRed, and LAC1-DsRed-OE cultured on lactose for 72 h. Scale bar = 10μm. Table S1: Primers used for gene amplification, verification, and qRT-PCR; Figure S5: Bioinformatic analysis of ER localization signal motif of sugar transporters MFS, GST, and LAC1. The ER localization signal motifs KKXX were highlighted with blue background. The amino acids in red background and white character are strictly identical, in red character are similar in protein sequence, and in blue frame are similar across protein sequences.
Author Contributions
Conceptualization, H.W. and F.L.; formal analysis, H.W.; funding acquisition, A.-P.P. and F.L.; methodology, H.W., A.-P.P., B.L. and F.L.; project administration, A.-P.P. and F.L.; visualization, H.W. and F.L.; writing—original draft, H.W. and F.L.; writing—review & editing, H.W., L.H., F.-G.W. and F.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32200037), State Key Laboratory of Microbial Technology Open Project Fund (No. M2022-12) of Food Nutrition and Safety (No. SKLFNS-KF-202214), and the Open Project Program of State Key Laboratory of Food Nutrition and Safety (No. SKLFNS-KF-202214).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are contained within the article and Supplementary Materials.
Acknowledgments
The authors would like to thank Wei Wang from East China University of Science and Technology and Zhihua Zhou from Chinese Academy of Sciences for kindly providing the T. reesei Δku70 strain and pXBthg plasmid, respectively.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kubicek, C.P.; Mikus, M.; Schuster, A.; Schmoll, M.; Seiboth, B. Metabolic engineering strategies for the improvement of cellulase production by Hypocrea jecorina. Biotechnol. Biofuels 2009, 2, 19. [Google Scholar] [CrossRef]
- Warzywoda, M.; Ferre, V.; Pourquie, J. Development of a culture medium for large-scale production of cellulolytic enzymes by Trichoderma reesei. Biotechnol. Bioeng. 1983, 25, 3005–3011. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, C.; Bååth, J.A.; Seiboth, B.; Kubicek, C.P. Systems analysis of lactose metabolism in Trichoderma reesei identifies a lactose permease that is essential for cellulase induction. PLoS ONE 2013, 8, e62631. [Google Scholar] [CrossRef] [PubMed]
- Porciuncula, J.d.O.; Furukawa, T.; Shida, Y.; Mori, K.; Kuhara, S.; Morikawa, Y.; Ogasawara, W. Identification of major facilitator transporters involved in cellulase production during lactose culture of Trichoderma reesei PC-3-7. Biosci. Biotechnol. Biochem. 2013, 77, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Kou, Y.; Xu, J.; Cao, Y.; Zhao, G.; Shao, J.; Wang, H.; Wang, Z.; Bao, X.; Chen, G. Two major facilitator superfamily sugar transporters from Trichoderma reesei and their roles in induction of cellulase biosynthesis. J. Biol. Chem. 2013, 288, 32861–32872. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Wang, B.; Ji, J.; Jiang, Y.; Wan, L.; Tian, C.; Ma, Y. The putative cellodextrin transporter-like protein CLP1 is involved in cellulase induction in Neurospora crassa. J. Biol. Chem. 2015, 290, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Znameroski, E.A.; Li, X.; Tsai, J.C.; Galazka, J.M.; Glass, N.L.; Cate, J.H. Evidence for transceptor function of cellodextrin transporters in Neurospora crassa. J. Biol. Chem. 2014, 289, 2610–2619. [Google Scholar] [CrossRef] [PubMed]
- Dos Reis, T.F.; de Lima, P.B.A.; Parachin, N.S.; Mingossi, F.B.; de Castro Oliveira, J.V.; Ries, L.N.A.; Goldman, G.H. Identification and characterization of putative xylose and cellobiose transporters in Aspergillus nidulans. Biotechnol. Biofuels 2016, 9, 204. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, G.; Chen, M.; Li, Z.; Qin, Y.; Qu, Y. Cellodextrin transporters play important roles in cellulase induction in the cellulolytic fungus Penicillium oxalicum. Appl. Microbiol. Biotechnol. 2013, 97, 10479–10488. [Google Scholar] [CrossRef] [PubMed]
- Sloothaak, J.; Tamayo-Ramos, J.A.; Odoni, D.I.; Laothanachareon, T.; Derntl, C.; Mach-Aigner, A.R.; Martins dos Santos, V.A.; Schaap, P.J. Identification and functional characterization of novel xylose transporters from the cell factories Aspergillus niger and Trichoderma reesei. Biotechnol. Biofuels 2016, 9, 148. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, R.; Lv, W.; Zhang, W.; Meng, X.; Liu, W. Functional characterization of sugar transporter CRT1 reveals differential roles of its C-terminal region in sugar transport and cellulase induction in Trichoderma reesei. Microbiol. Spectr. 2022, 10, e00872-00822. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Pang, A.-P.; Wang, W.; Li, B.; Li, C.; Wu, F.-G.; Lin, F. Discovery of ER-localized sugar transporters for cellulase production with lac1 being essential. Biotechnol. Biofuels 2022, 15, 132. [Google Scholar] [CrossRef] [PubMed]
- Seiboth, B.; Hartl, L.; Salovuori, N.; Lanthaler, K.; Robson, G.D.; Vehmaanperä, J.; Penttilä, M.E.; Kubicek, C.P. Role of the bga1-encoded extracellular β-galactosidase of Hypocrea jecorina in cellulase induction by lactose. Appl. Environ. Microbiol. 2005, 71, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, Y.; Ohashi, T.; Mantani, O.; Okada, H. Cellulase induction by lactose in Trichoderma reesei PC-3-7. Appl. Microbiol. Biotechnol. 1995, 44, 106–111. [Google Scholar] [CrossRef]
- Havukainen, S.; Valkonen, M.; Koivuranta, K.; Landowski, C.P. Studies on sugar transporter CRT1 reveal new characteristics that are critical for cellulase induction in Trichoderma reesei. Biotechnol. Biofuels 2020, 13, 158. [Google Scholar] [CrossRef] [PubMed]
- Fekete, E.; Karaffa, L.; Seiboth, B.; Fekete, É.; Kubicek, C.P.; Flipphi, M. Identification of a permease gene involved in lactose utilisation in Aspergillus nidulans. Fungal Genet. Biol. 2012, 49, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Guangtao, Z.; Hartl, L.; Schuster, A.; Polak, S.; Schmoll, M.; Wang, T.; Seidl, V.; Seiboth, B. Gene targeting in a nonhomologous end joining deficient Hypocrea jecorina. J. Biotechnol. 2009, 139, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, X.; Zhang, G.; Zhang, J.; Wang, X.; Zhang, S.; Wang, W.; Wei, D. Light-inducible genetic engineering and control of non-homologous end-joining in industrial eukaryotic microorganisms: LML 3.0 and OFN 1.0. Sci. Rep. 2016, 6, 20761. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, Y.; Huang, X.; Sun, S.; Luo, J.; Lu, Z.; Wang, W.; Ma, Y. An efficient shortened genetic transformation strategy for filamentous fungus Trichoderma reesei. J. Gen. Appl. Microbiol. 2019, 65, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Minty, J.J.; Singer, M.E.; Scholz, S.A.; Bae, C.-H.; Ahn, J.-H.; Foster, C.E.; Liao, J.C.; Lin, X.N. Design and characterization of synthetic fungal-bacterial consortia for direct production of isobutanol from cellulosic biomass. Proc. Natl. Acad. Sci. USA 2013, 110, 14592–14597. [Google Scholar] [CrossRef]
- Zhong, Y.H.; Wang, X.L.; Wang, T.H.; Jiang, Q. Agrobacterium-mediated transformation (AMT) of Trichoderma reesei as an efficient tool for random insertional mutagenesis. Appl. Microbiol. Biotechnol. 2007, 73, 1348–1354. [Google Scholar] [CrossRef] [PubMed]
- Pang, A.-P.; Wang, H.; Luo, Y.; Yang, Z.; Liu, Z.; Wang, Z.; Li, B.; Yang, S.; Zhou, Z.; Lu, X. Dissecting Cellular Function and Distribution of β-Glucosidases in Trichoderma reesei. Mbio 2021, 12, e03671-20. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lin, F.; Li, Y.; Wei, W.; Wang, H.; Qin, L.; Zhou, Z.; Li, B.; Wu, F.; Chen, Z. A β-glucosidase hyper-production Trichoderma reesei mutant reveals a potential role of cel3D in cellulase production. Microb. Cell Factories 2016, 15, 151. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lin, F.; Zhou, L.; Qin, L.; Li, B.; Zhou, Z.; Jin, M.; Chen, Z. Cellulase hyper-production by Trichoderma reesei mutant SEU-7 on lactose. Biotechnol. Biofuels 2017, 10, 228. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Storms, R.; Tsang, A. Microplate-based filter paper assay to measure total cellulase activity. Biotechnol. Bioeng. 2004, 88, 832–837. [Google Scholar] [CrossRef]
- Xiao, Z.; Storms, R.; Tsang, A. Microplate-based carboxymethylcellulose assay for endoglucanase activity. Anal. Biochem. 2005, 342, 176–178. [Google Scholar] [CrossRef]
- Derntl, C.; Mach, R.L.; Mach-Aigner, A.R. Fusion transcription factors for strong, constitutive expression of cellulases and xylanases in Trichoderma reesei. Biotechnol. Biofuels 2019, 12, 231. [Google Scholar] [CrossRef] [PubMed]
- Steiger, M.G.; Mach, R.L.; Mach-Aigner, A.R. An accurate normalization strategy for RT-qPCR in Hypocrea jecorina (Trichoderma reesei). J. Biotechnol. 2010, 145, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Pang, A.-P.; Yang, H.; Lv, R.; Zhou, Z.; Wu, F.-G.; Lin, F. Tracking localization and secretion of cellulase spatiotemporally and directly in living Trichoderma reesei. Biotechnol. Biofuels 2019, 12, 200. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Hua, X.-W.; Jia, H.-R.; Li, C.; Lin, F.; Chen, Z.; Wu, F.-G. Universal cell surface imaging for mammalian, fungal, and bacterial cells. ACS Biomater. Sci. Eng. 2016, 2, 987–997. [Google Scholar] [CrossRef] [PubMed]
- Pang, A.-P.; Luo, Y.; Hu, X.; Zhang, F.; Wang, H.; Gao, Y.; Durrani, S.; Li, C.; Shi, X.; Wu, F.-G. Transmembrane transport process and endoplasmic reticulum function facilitate the role of gene cel1b in cellulase production of Trichoderma reesei. Microb. Cell Factories 2022, 21, 90. [Google Scholar] [CrossRef] [PubMed]
- Mello-de-Sousa, T.M.; Rassinger, A.; Derntl, C.; Poças-Fonseca, M.J.; Mach, R.L.; Mach-Aigner, A.R. The relation between promoter chromatin status, Xyr1 and cellulase expression in Trichoderma reesei. Curr. Genom. 2016, 17, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Häkkinen, M.; Valkonen, M.J.; Westerholm-Parvinen, A.; Aro, N.; Arvas, M.; Vitikainen, M.; Penttilä, M.; Saloheimo, M.; Pakula, T.M. Screening of candidate regulators for cellulase and hemicellulase production in Trichoderma reesei and identification of a factor essential for cellulase production. Biotechnol. Biofuels 2014, 7, 14. [Google Scholar] [CrossRef]
- Coradetti, S.T.; Craig, J.P.; Xiong, Y.; Shock, T.; Tian, C.; Glass, N.L. Conserved and essential transcription factors for cellulase gene expression in ascomycete fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 7397–7402. [Google Scholar] [CrossRef] [PubMed]
- Xiong, B.; Wei, L.; Wang, Y.; Li, J.; Liu, X.; Zhou, Y.; Du, P.; Fang, H.; Liesche, J.; Wei, Y. Parallel proteomic and phosphoproteomic analyses reveal cellobiose-dependent regulation of lignocellulase secretion in the filamentous fungus Neurospora crassa. GCB Bioenergy 2021, 13, 1372–1387. [Google Scholar] [CrossRef]
- Wang, B.-T.; Hu, S.; Yu, X.-Y.; Jin, L.; Zhu, Y.-J.; Jin, F.-J. Studies of cellulose and starch utilization and the regulatory mechanisms of related enzymes in fungi. Polymers 2020, 12, 530. [Google Scholar] [CrossRef] [PubMed]
- Derntl, C.; Kluger, B.; Bueschl, C.; Schuhmacher, R.; Mach, R.L.; Mach-Aigner, A.R. Transcription factor Xpp1 is a switch between primary and secondary fungal metabolism. Proc. Natl. Acad. Sci. USA 2017, 114, E560–E569. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.R.; Nilsson, T.; Peterson, P.A. Identification of a consensus motif for retention of transmembrane proteins in the endoplasmic reticulum. EMBO J. 1990, 9, 3153–3162. [Google Scholar] [CrossRef]
- Stornaiuolo, M.; Lotti, L.V.; Borgese, N.; Torrisi, M.-R.; Mottola, G.; Martire, G.; Bonatti, S. KDEL and KKXX retrieval signals appended to the same reporter protein determine different trafficking between endoplasmic reticulum, intermediate compartment, and Golgi complex. Mol. Biol. Cell. 2003, 14, 889–902. [Google Scholar] [CrossRef]
- Seiboth, B.; Hartl, L.; Pail, M.; Fekete, E.; Karaffa, L.; Kubicek, C.P. The galactokinase of Hypocrea jecorina is essential for cellulase induction by lactose but dispensable for growth on d-galactose. Mol. Microbiol. 2004, 51, 1015–1025. [Google Scholar] [CrossRef]
- Seiboth, B.; Hofmann, G.; Kubicek, C. Lactose metabolism and cellulase production in Hypocrea jecorina: The gal7 gene, encoding galactose-1-phosphate uridylyltransferase, is essential for growth on galactose but not for cellulase induction. Mol. Genet. Genom. 2002, 267, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Fekete, E.; Seiboth, B.; Kubicek, C.P.; Szentirmai, A.; Karaffa, L. Lack of aldose 1-epimerase in Hypocrea jecorina (anamorph Trichoderma reesei): A key to cellulase gene expression on lactose. Proc. Natl. Acad. Sci. USA 2008, 105, 7141–7146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, Y.; Wu, C.; Liu, P.; Wang, W.; Wei, D. The transcription factor ACE3 controls cellulase activities and lactose metabolism via two additional regulators in the fungus Trichoderma reesei. J. Biol. Chem. 2019, 294, 18435–18450. [Google Scholar] [CrossRef] [PubMed]
- Bischof, R.; Fourtis, L.; Limbeck, A.; Gamauf, C.; Seiboth, B.; Kubicek, C.P. Comparative analysis of the Trichoderma reesei transcriptome during growth on the cellulase inducing substrates wheat straw and lactose. Biotechnol. Biofuels 2013, 6, 127. [Google Scholar] [CrossRef] [PubMed]
- Seiboth, B.; Gamauf, C.; Pail, M.; Hartl, L.; Kubicek, C.P. The d-xylose reductase of Hypocrea jecorina is the major aldose reductase in pentose and d-galactose catabolism and necessary for β-galactosidase and cellulase induction by lactose. Mol. Microbiol. 2007, 66, 890–900. [Google Scholar] [CrossRef]
- Ries, L.; Pullan, S.T.; Delmas, S.; Malla, S.; Blythe, M.J.; Archer, D.B. Genome-wide transcriptional response of Trichoderma reesei to lignocellulose using RNA sequencing and comparison with Aspergillus niger. BMC Genom. 2013, 14, 541. [Google Scholar] [CrossRef]
- Banfield, D.K. Mechanisms of protein retention in the Golgi. Cold Spring Harb. Perspect. Biol. 2011, 3, a005264. [Google Scholar] [CrossRef]
- Orellana, A.; Moraga, C.; Araya, M.; Moreno, A. Overview of nucleotide sugar transporter gene family functions across multiple species. J. Mol. Biol. 2016, 428, 3150–3165. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).