
Residues of Legume AG41 Peptide Crucial to Its Bio-Insecticidal Activity

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
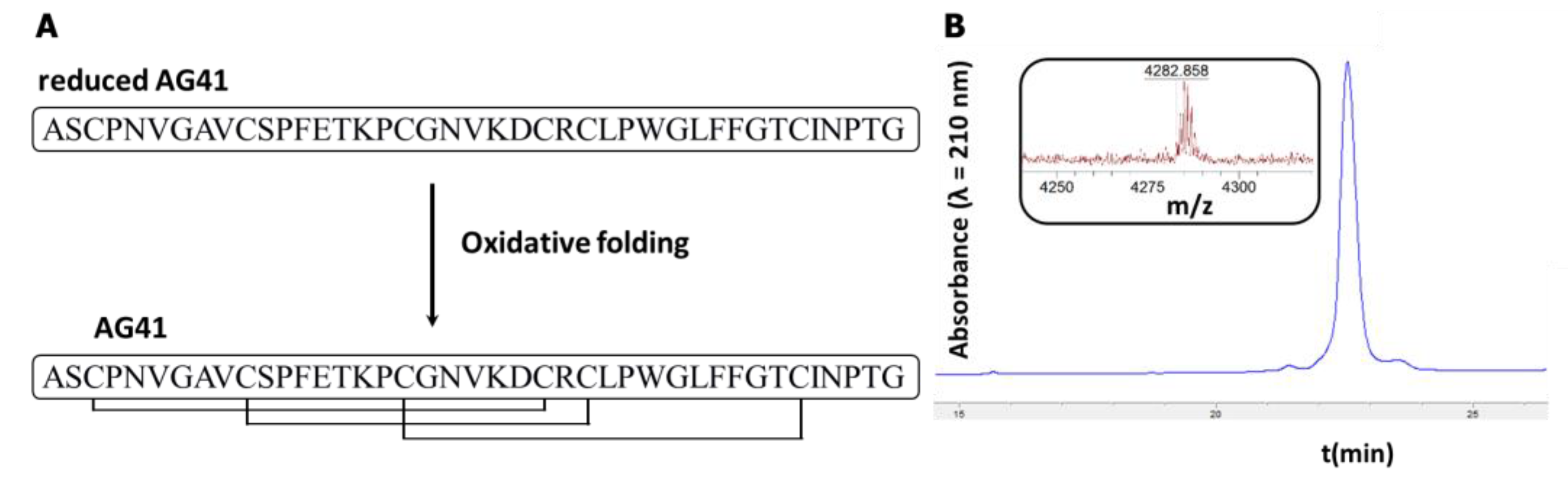
2.1. Synthesis and Purification of Peptides
2.2. NMR Sample Preparation
2.3. NMR Experiments
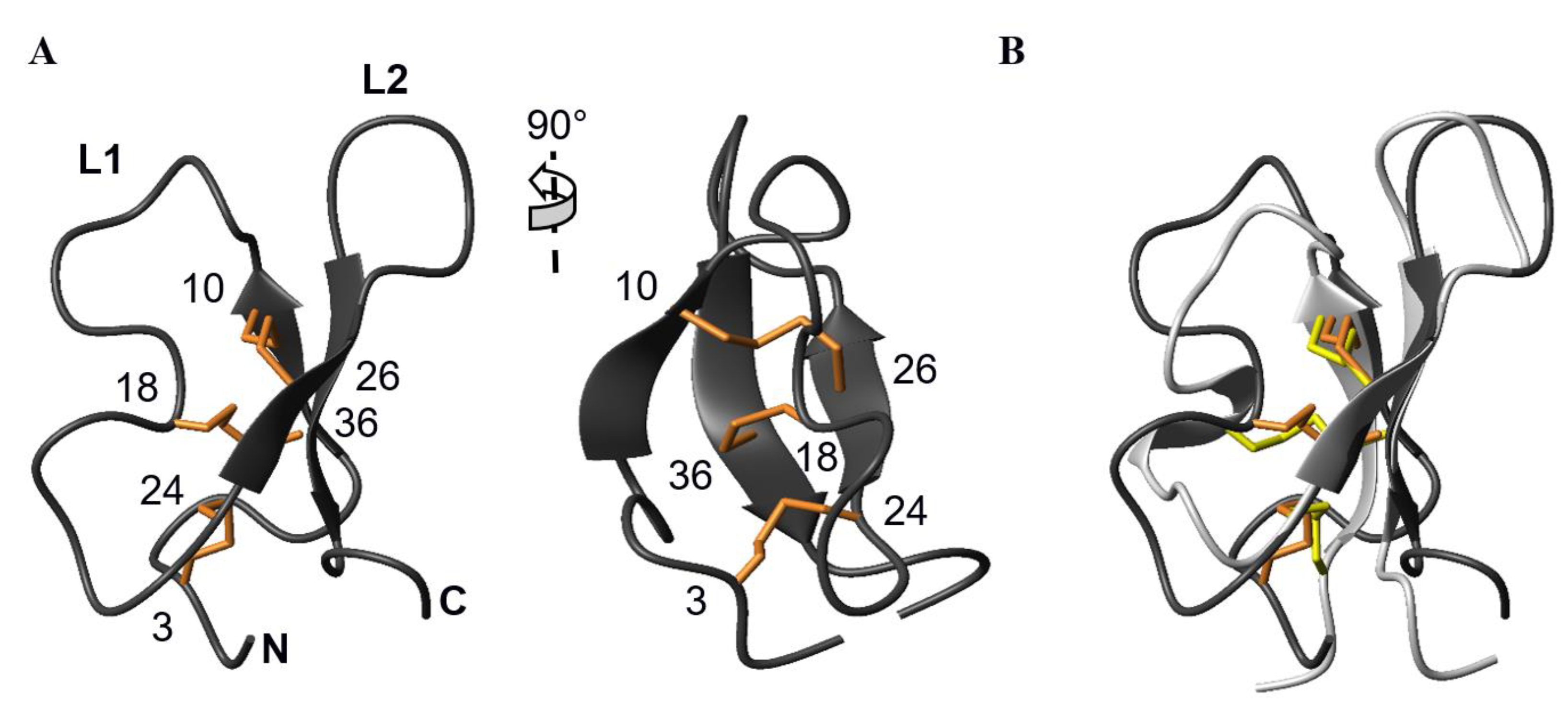
2.4. Structure Calculations
2.5. Bioassays
2.6. Biological Data Analyses
3. Results and Discussion
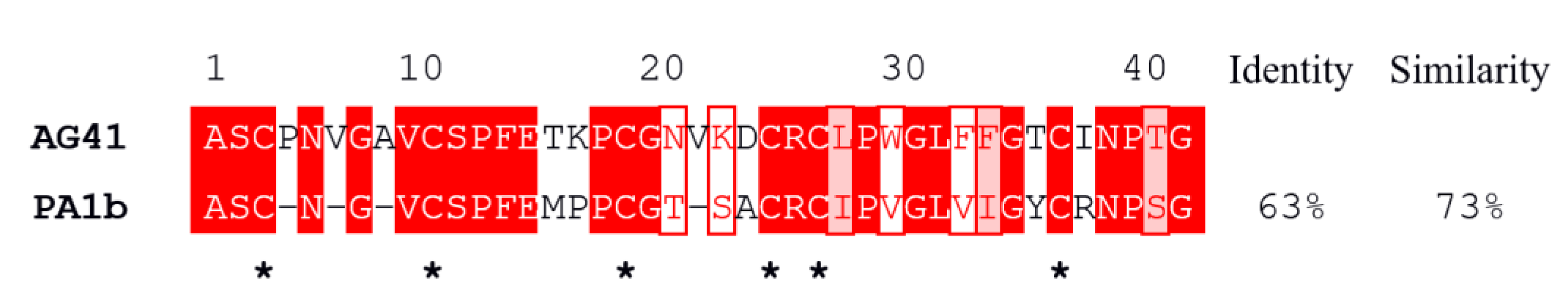
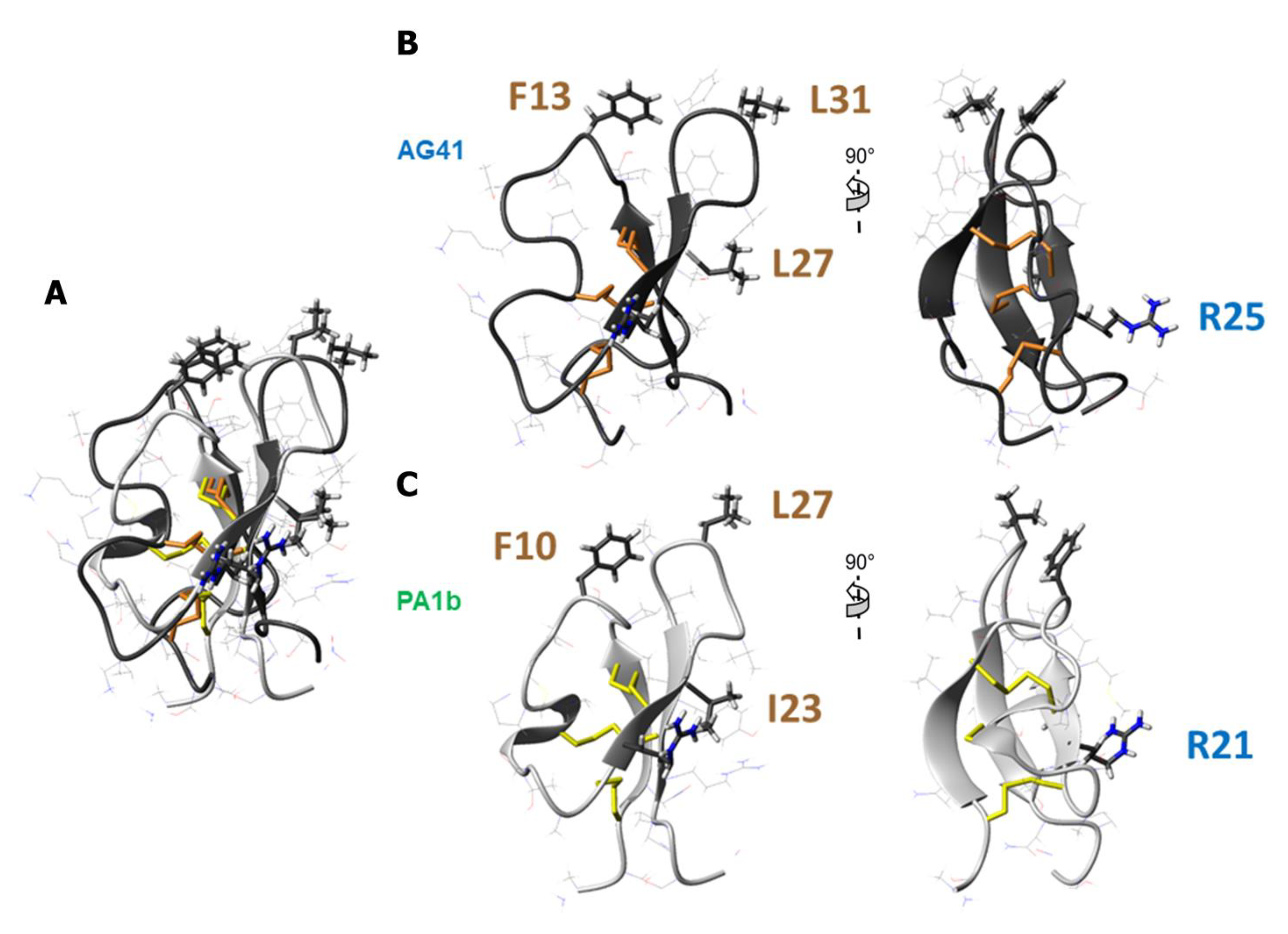
3.1. The AG41 Peptide Fold Is Similar to That of PA1b
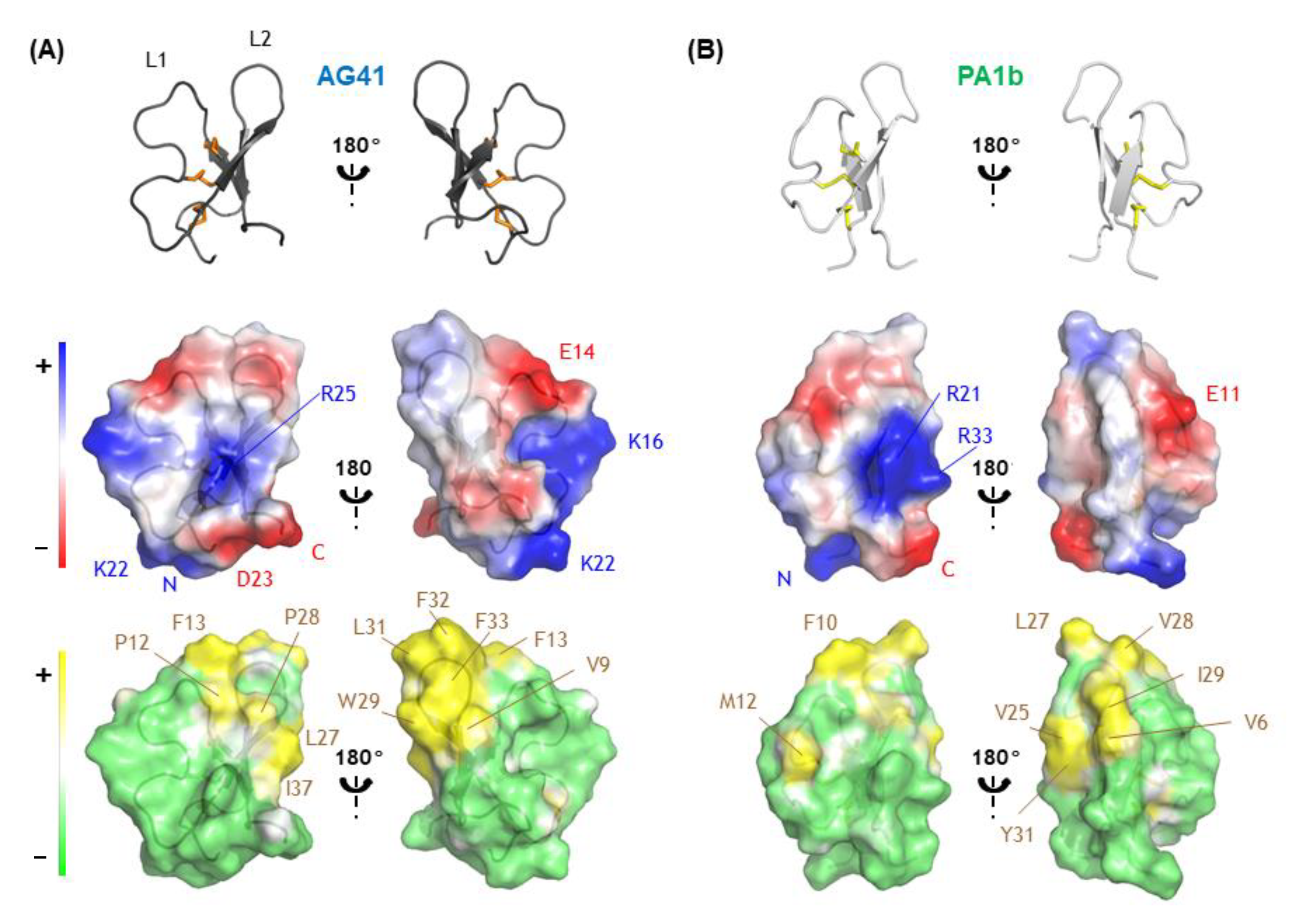
3.2. Structural Comparison Revealed the Potential Key Residues in the Activity of AG41
3.3. Synthesis and Structural Characterization of AG41 Mutants
3.4. Bioactivity of Alanine Mutants of AG41
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tudi, M.; Daniel Ruan, H.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D.T. Agriculture Development, Pesticide Application and Its Impact on the Environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef] [PubMed]
- Jacquet, F.; Jeuffroy, M.-H.; Jouan, J.; Le Cadre, E.; Litrico, I.; Malausa, T.; Reboud, X.; Huyghe, C. Pesticide-free agriculture as a new paradigm for research. Agron. Sustain. Dev. 2022, 42, 8. [Google Scholar] [CrossRef]
- Tang, F.H.M.; Lenzen, M.; McBratney, A.; Maggi, F. Risk of pesticide pollution at the global scale. Nat. Geosci. 2021, 14, 206–210. [Google Scholar] [CrossRef]
- Goulson, D. Call to restrict neonicotinoids. Science 2018, 360, 973. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef]
- Da Silva, P. Use of Peptides as Insecticidal Agents. U.S. Patent US2021/0137126 A1, 13 May 2021. [Google Scholar]
- Loth, K.; Parisot, N.; Paquet, F.; Terrasson, H.; Sivignon, C.; Rahioui, I.; Ribeiro Lopes, M.; Gaget, K.; Duport, G.; Delmas, A.F.; et al. Aphid BCR4 Structure and Activity Uncover a New Defensin Peptide Superfamily. Int. J. Mol. Sci. 2022, 23, 12480. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-M.; Ye, D.-X.; Liu, Y.; Zhang, X.-Y.; Zhou, Y.-L.; Zhang, L.; Yang, X.-L. Peptides, new tools for plant protection in eco-agriculture. Adv. Agrochem 2023, in press. [CrossRef]
- Royer, C.; Da Silva, P.; Gressent, F.; Karaki, L.; Rahbe, Y. Entomotoxic Polypeptides. U.S. Patent US9,994,623, 18 June 2015. [Google Scholar]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Delobel, B.; Grenier, A.M.; Gueguen, J.; Ferrasson, E.; Mbaiguinam, M. Utilisation d’un Polypeptide Dérivé D’une Albumine PA1b de Légumineuse Comme Insecticide. French Patent Brevet-98/05877, 11 May 1998. [Google Scholar]
- Chouabe, C.; Eyraud, V.; Silva, P.; Rahioui, I.; Royer, C.; Soulage, C.; Bonvallet, R.; Huss, M.; Gressent, F. New Mode of Action for a Knottin Protein Bioinsecticide: Pea Albumin 1 Subunit b (PA1b) is the first Peptidic Inhibitor of V-ATPase*. J. Biol. Chem. 2011, 286, 36291–36296. [Google Scholar] [CrossRef]
- Rahioui, I.; Eyraud, V.; Karaki, L.; Sasse, F.; Carre-Pierrat, M.; Qin, A.; Zheng, M.H.; Toepfer, S.; Sivignon, C.; Royer, C.; et al. Host range of the potential biopesticide Pea Albumin 1b (PA1b) is limited to insects. Toxicon Off. J. Int. Soc. Toxinology 2014, 89, 67–76. [Google Scholar] [CrossRef]
- Gressent, F.; Duport, G.; Rahioui, I.; Pauchet, Y.; Bolland, P.; Specty, O.; Rahbe, Y. Biological activity and binding site characteristics of the PA1b entomotoxin on insects from different orders. J. Insect Sci. 2007, 7, 12. [Google Scholar] [CrossRef]
- Muench, S.P.; Rawson, S.; Eyraud, V.; Delmas, A.F.; Da Silva, P.; Phillips, C.; Trinick, J.; Harrison, M.A.; Gressent, F.; Huss, M. PA1b Inhibitor Binding to Subunits c and e of the Vacuolar ATPase Reveals Its Insecticidal Mechanism. J. Biol. Chem. 2014, 289, 16399–16408. [Google Scholar] [CrossRef] [PubMed]
- Eyraud, V.; Balmand, S.; Karaki, L.; Rahioui, I.; Sivignon, C.; Delmas, A.F.; Royer, C.; Rahbe, Y.; Da Silva, P.; Gressent, F. The interaction of the bioinsecticide PA1b (Pea Albumin 1 subunit b) with the insect V-ATPase triggers apoptosis. Sci. Rep. 2017, 7, 4902. [Google Scholar] [CrossRef]
- Karaki, L.; Da Silva, P.; Rizk, F.; Chouabe, C.; Chantret, N.; Eyraud, V.; Gressent, F.; Sivignon, C.; Rahioui, I.; Kahn, D.; et al. Genome-wide analysis identifies gain and loss/change of function within the small multigenic insecticidal Albumin 1 family of Medicago truncatula. BMC Plant Biol. 2016, 16, 1–19. [Google Scholar] [CrossRef]
- Da Silva, P.; Rahioui, I.; Laugier, C.; Jouvensal, L.; Meudal, H.; Chouabe, C.; Delmas, A.F.; Gressent, F. Molecular Requirements for the Insecticidal Activity of the Plant Peptide Pea Albumin 1 Subunit b (PA1b). J. Biol. Chem. 2010, 285, 32689–32694. [Google Scholar] [CrossRef]
- Da Silva, P.; Strzepa, A.; Jouvensal, L.; Rahioui, I.; Gressent, F.; Delmas, A.F. A folded and functional synthetic PA1b: An interlocked entomotoxic miniprotein. Biopolymers 2009, 92, 436–444. [Google Scholar] [CrossRef]
- Vranken, W.F.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, P.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins 2005, 59, 687–696. [Google Scholar] [CrossRef]
- Brunger, A.T. Version 1.2 of the Crystallography and NMR system. Nat. Protoc. 2007, 2, 2728–2733. [Google Scholar] [CrossRef]
- Brunger, A.T.; Adams, P.D.; Clore, G.M.; DeLano, W.L.; Gros, P.; Grosse-Kunstleve, R.W.; Jiang, J.S.; Kuszewski, J.; Nilges, M.; Pannu, N.S.; et al. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr. Sect. D Biol. Crystallogr. 1998, 54, 905–921. [Google Scholar] [CrossRef]
- Rieping, W.; Habeck, M.; Bardiaux, B.; Bernard, A.; Malliavin, T.E.; Nilges, M. ARIA2: Automated NOE assignment and data integration in NMR structure calculation. Bioinformatics 2007, 23, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.S.; Maguire, M.L.; Stevens, T.J.; Broadhurst, R.W. DANGLE: A Bayesian inferential method for predicting protein backbone dihedral angles and secondary structure. J. Magn. Reson 2010, 202, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; Rullmann, J.A.C.; MacArthur, M.W.; Kaptein, R.; Thornton, J.M. AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 1996, 8, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, E.G.; Thornton, J.M. PROMOTIF—A program to identify and analyze structural motifs in proteins. Protein Sci. 1996, 5, 212–220. [Google Scholar] [CrossRef]
- Koradi, R.; Billeter, M.; Wüthrich, K. MOLMOL: A program for display and analysis of macromolecular structures. J. Mol. Graph. 1996, 14, 51–55. [Google Scholar] [CrossRef]
- De Lano, W.L. Pymol; De Lano Scientific: South San Francisco, CA, USA, 2002. [Google Scholar]
- Gressent, F.; Rahioui, I.; Rahbe, Y. Characterization of a high-affinity binding site for the pea albumin 1b entomotoxin in the weevil Sitophilus. Eur. J. Biochem. 2003, 270, 2429–2435. [Google Scholar] [CrossRef]
- Rahioui, I.; Laugier, C.; Balmand, S.; Silva, P.; Rahbé, Y.; Gressent, F. Toxicity, binding and internalization of the pea-A1b entomotoxin in Sf9 cells. Biochimie 2007, 89, 1539–1543. [Google Scholar] [CrossRef] [PubMed]
- Holzhütter, H.G.; Colosimo, A. SIMFIT: A microcomputer software-toolkit for modelistic studies in biochemistry. Bioinformatics 1990, 6, 23–28. [Google Scholar] [CrossRef]
- Gelly, J.C.; Gracy, J.; Kaas, Q.; Le-Nguyen, D.; Heitz, A.; Chiche, L. The KNOTTIN website and database: A new information system dedicated to the knottin scaffold. Nucleic Acids Res. 2004, 32, D156–D159. [Google Scholar] [CrossRef]
- Gracy, J.; Le-Nguyen, D.; Gelly, J.-C.; Kaas, Q.; Heitz, A.; Chiche, L. KNOTTIN: The knottin or inhibitor cystine knot scaffold in 2007. Nucleic Acids Res. 2008, 36, D314–D319. [Google Scholar] [CrossRef]
- Jouvensal, L.; Quillien, L.; Ferrasson, E.; Rahbe, Y.; Gueguen, J.; Vovelle, F. PA1b, an insecticidal protein extracted from pea seeds (Pisum sativum): 1H-2-D NMR study and molecular modeling. Biochemistry 2003, 42, 11915–11923. [Google Scholar] [CrossRef]
- Coste, F.; Chaume, G.; Madinier, J.B.; Rahioui, I.; Castaing, B.; Da Silva, P.; Brigaud, T.; Aucagne, V.; Delmas, A.F. Incorporation of trifluoromethylated pseudoproline in an entomotoxic knottin: Effects on oxidative folding, 3D structure, and biological activity. J. Pept. Sci. 2018, 24, S119. [Google Scholar]
- Pyrkov, T.V.; Chugunov, A.O.; Krylov, N.A.; Nolde, D.E.; Efremov, R.G. PLATINUM: A web tool for analysis of hydrophobic/hydrophilic organization of biomolecular complexes. Bioinformatics 2009, 25, 1201–1202. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wong, K.H.; Tay, S.V.; Serra, A.; Sze, S.K.; Tam, J.P. Astratides: Insulin-Modulating, Insecticidal, and Antifungal Cysteine-Rich Peptides from Astragalus membranaceus. J. Nat. Prod. 2019, 82, 194–204. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NMR Restraints | ||
|---|---|---|
| Distance Restraints | ||
| Total NOE | 920 | |
| Unambiguous | 845 | |
| Ambiguous | 75 | |
| Hydrogen bonds | 8 | |
| Dihedral Angle Restraints | 56 | |
| Covalent disulfide bridges | Cys3-Cys24 Cys10-Cys26 Cys18-Cys36 | |
| Structural Statistics for the final 10 models of AG41 | ||
| Average number of violations per structure | ||
| NOEs ≥ 0.3 Å | 0 | |
| Hydrogen bonds ≥ 0.5 Å; ≥ 0.3 Å | 0; 0.1 | |
| Dihedrals ≥ 10°; ≥ 5° | 0; 0.9 | |
| Average RMSD (pairwise, Å) a,b | bb (N-Cα-C’) | All heavy atoms |
| Whole (2–41) | 0.46 ± 0.09 | 0.92 ± 0.11 |
| Triple-stranded β-sheet (8–9/25–27/35–37) | 0.13 ± 0.04 | 0.62 ± 0.23 |
| Loop L1 (10–24) | 0.39 ± 0.13 | 1.02 ± 0.20 |
| Hairpin loop L2 (28–34) | 0.09 ± 0.04 | 0.67 ± 0,28 |
| Ramachandran Analysis | ||
| Most favored region and allowed region | 96.0 | |
| Generously allowed | 4.0 | |
| Disallowed | 0.0 | |
| Energies (kcal.mol −1) a | ||
| Electrostatic | −1313.83 ± 22.11 | |
| Van der Walls | −306.91 ± 3.89 | |
| Total energy | −1204.00 ± 17.30 | |
| Residual NOE energy | 26.83 ± 2.21 | |
| AG41 and Its Mutants (Name and Sequence) a | Ki b (nM) | LD50 c (nM) | |
|---|---|---|---|
| AG41 | ASCPNVGAVCSPFETKPCGNVKDCRCLPWGLFFGTCINPTG | 1.3 ± 0.6 | 5.6 ± 1.8 |
| F13A | ASCPNVGAVCSPAETKPCGNVKDCRCLPWGLFFGTCINPTG | - | - |
| E14A | ASCPNVGAVCSPFATKPCGNVKDCRCLPWGLFFGTCINPTG | 1.2 ± 1.3 | 5.6 ± 1.8 |
| K16A | ASCPNVGAVCSPFETAPCGNVKDCRCLPWGLFFGTCINPTG | 3.6 ± 1.6 | 10.0 ± 4.0 |
| K22A | ASCPNVGAVCSPFETKPCGNVADCRCLPWGLFFGTCINPTG | 3.2 ± 1.7 | 14.2 ± 6.3 |
| D23A | ASCPNVGAVCSPFETKPCGNVKACRCLPWGLFFGTCINPTG | 1.2 ± 1.6 | 24 ± 9 |
| R25A | ASCPNVGAVCSPFETKPCGNVKDCACLPWGLFFGTCINPTG | - | - |
| L27A | ASCPNVGAVCSPFETKPCGNVKDCRCAPWGLFFGTCINPTG | 486 ± 11 | 907 ± 46 |
| W29A | ASCPNVGAVCSPFETKPCGNVKDCRCLPAGLFFGTCINPTG | 36.3 ± 6.7 | 89.0 ± 3.7 |
| L31A | ASCPNVGAVCSPFETKPCGNVKDCRCLPWGAFFGTCINPTG | 263 ± 8 | - |
| F32A | ASCPNVGAVCSPFETKPCGNVKDCRCLPWGLAFGTCINPTG | 39.8 ± 15.0 | 122 ± 12 |
| F33A | ASCPNVGAVCSPFETKPCGNVKDCRCLPWGLFAGTCINPTG | 15.0 ± 4.8 | 210 ± 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diya, F.; Jouvensal, L.; Rahioui, I.; Loth, K.; Sivignon, C.; Karaki, L.; Kfoury, L.; Rizk, F.; Da Silva, P. Residues of Legume AG41 Peptide Crucial to Its Bio-Insecticidal Activity. Biomolecules 2023, 13, 446. https://doi.org/10.3390/biom13030446
Diya F, Jouvensal L, Rahioui I, Loth K, Sivignon C, Karaki L, Kfoury L, Rizk F, Da Silva P. Residues of Legume AG41 Peptide Crucial to Its Bio-Insecticidal Activity. Biomolecules. 2023; 13(3):446. https://doi.org/10.3390/biom13030446
Chicago/Turabian StyleDiya, Fatima, Laurence Jouvensal, Isabelle Rahioui, Karine Loth, Catherine Sivignon, Lamis Karaki, Linda Kfoury, Francine Rizk, and Pedro Da Silva. 2023. "Residues of Legume AG41 Peptide Crucial to Its Bio-Insecticidal Activity" Biomolecules 13, no. 3: 446. https://doi.org/10.3390/biom13030446
APA StyleDiya, F., Jouvensal, L., Rahioui, I., Loth, K., Sivignon, C., Karaki, L., Kfoury, L., Rizk, F., & Da Silva, P. (2023). Residues of Legume AG41 Peptide Crucial to Its Bio-Insecticidal Activity. Biomolecules, 13(3), 446. https://doi.org/10.3390/biom13030446