
Molecular Mechanisms Driving and Regulating the AAA+ ATPase VCP/p97, an Important Therapeutic Target for Treating Cancer, Neurological and Infectious Diseases
 ,
,
Abstract
:1. Introduction
1.1. General Introduction
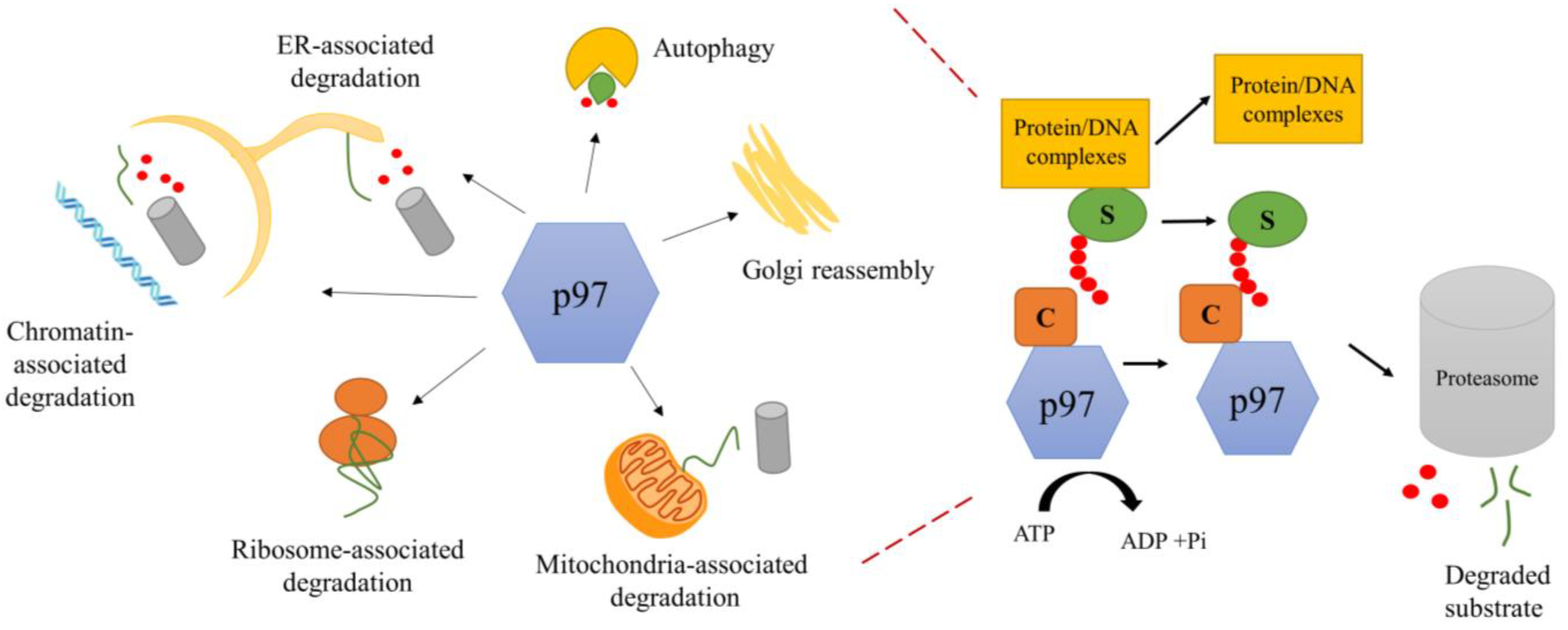
1.2. Biological Function of p97
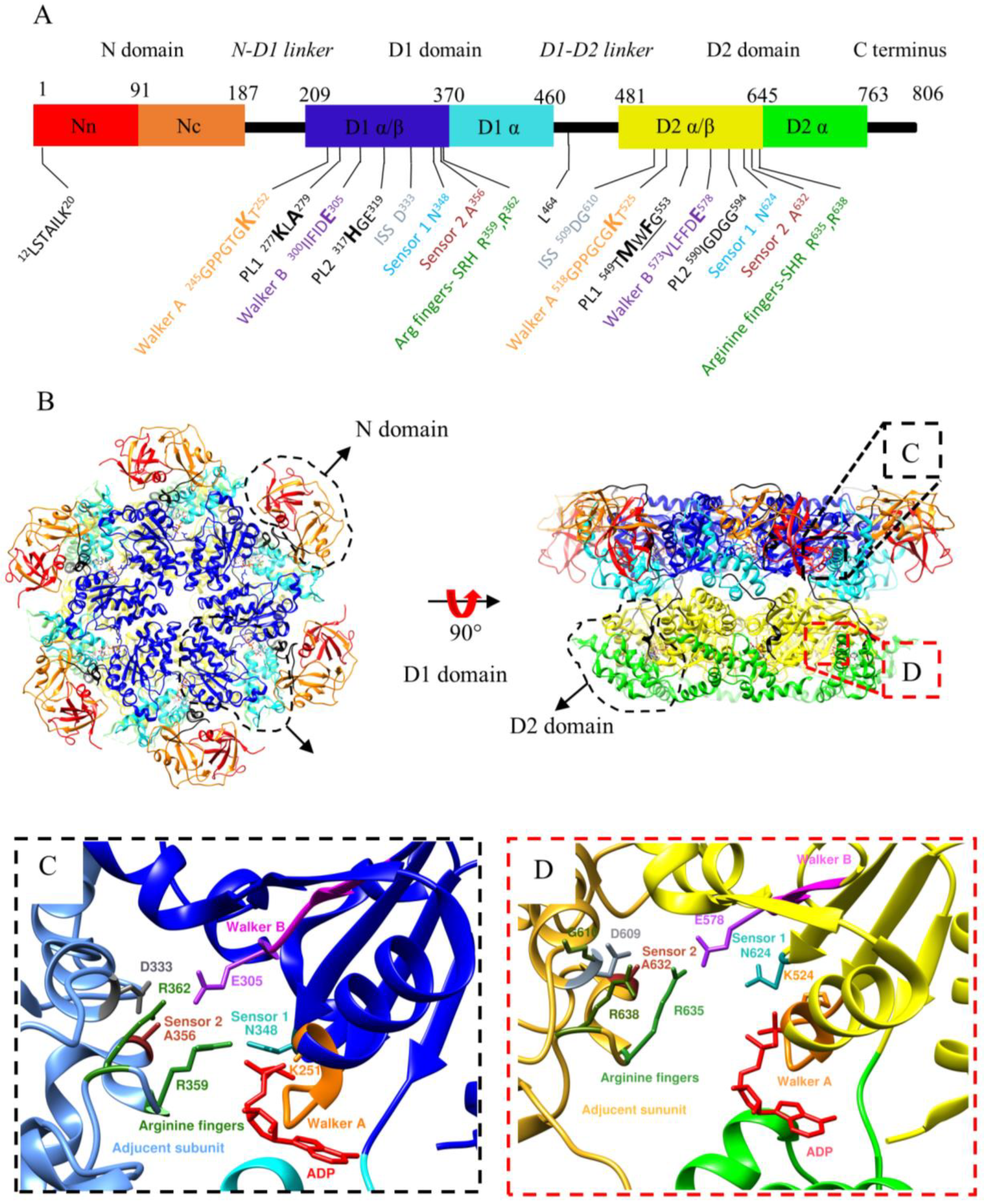
1.3. Structure of p97
2. The Mechanochemical Cycle of p97
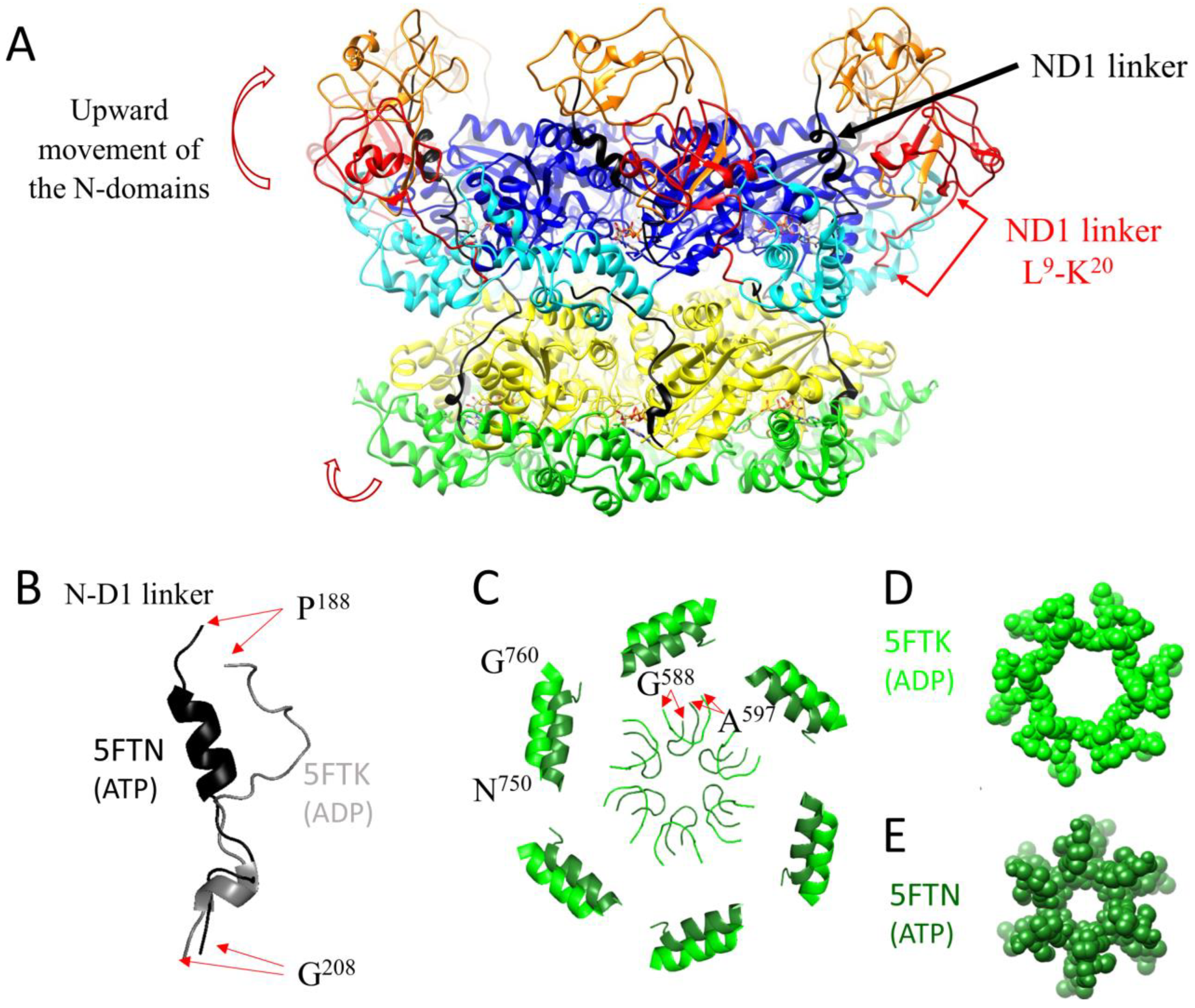
2.1. Conformational Changes during the ATPase Cycle of p97
2.2. Conformational Changes of p97 upon Substrate Binding
2.3. The Orthodox Model of Substrate Unfolding
3. Regulation of p97 Activity
3.1. Regulation of p97 Activity by Cofactors
3.2. Effect of Disease-Causing Mutations on the Structure and Function of p97
3.3. Regulating p97 Activity Using Small Molecules
3.3.1. ATP-Competitive Inhibitors
3.3.2. Allosteric Inhibitors
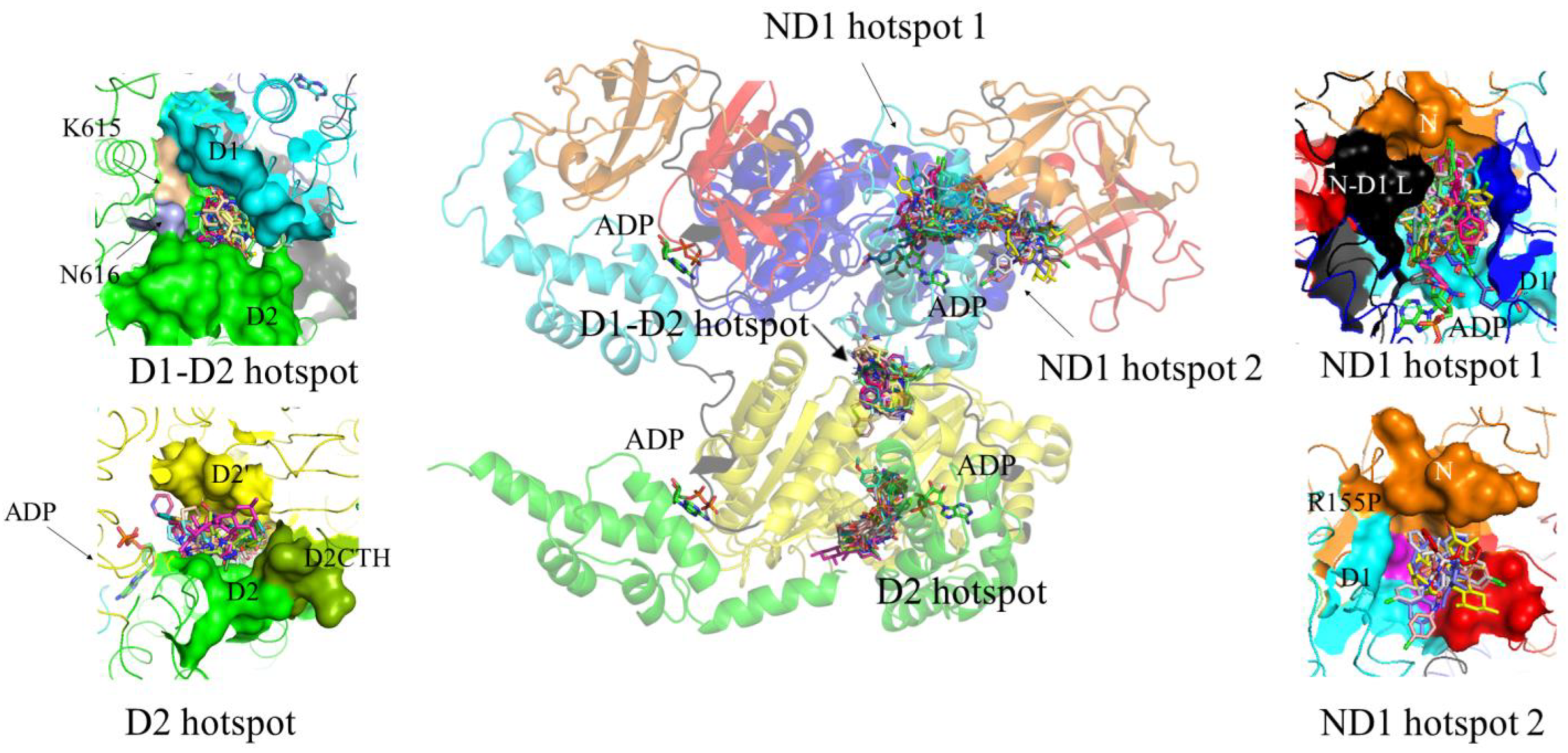
3.3.3. Potential Additional Binding Sites for Allosteric Inhibitors
4. Discussion and Future Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ogura, T.; Wilkinson, A.J. AAA+ superfamily ATPases: Common structure-diverse function. Genes Cells 2001, 6, 575–597. [Google Scholar] [CrossRef] [PubMed]
- Snider, J.; Thibault, G.; Houry, W.A. The AAA+ superfamily of functionally diverse proteins. Genome Biol. 2008, 9, 216. [Google Scholar] [CrossRef] [PubMed]
- Barthelme, D.; Sauer, R.T. Origin and Functional Evolution of the Cdc48/p97/VCP AAA+ Protein Unfolding and Remodeling Machine. J. Mol. Biol. 2016, 428 Pt B, 1861–1869. [Google Scholar] [CrossRef]
- Unciuleac, M.-C.; Smith, P.C.; Shuman, S. Crystal Structure and Biochemical Characterization of a Mycobacterium smegmatis AAA-Type Nucleoside Triphosphatase Phosphohydrolase (Msm0858). J. Bacteriol. 2016, 198, 1521–1533. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, B.G.; Padmanabhan, P.K.; Dumas, C.; Papadopoulou, B. Valosin-containing protein VCP/p97 is essential for the intracellular development of Leishmania and its survival under heat stress. Cell. Microbiol. 2018, 20, e12867. [Google Scholar] [CrossRef]
- Roggy, J.L.; Bangs, J.D. Molecular cloning and biochemical characterization of a VCP homolog in African trypanosomes. Mol. Biochem. Parasitol. 1999, 98, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pamnani, V.; Tamura, T.; Lupas, A.; Peters, J.; Cejka, Z.; Ashraf, W.; Baumeister, W. Cloning, sequencing and expression of VAT, a CDC48/p97 ATPase homologue from the archaeon Thermoplasma acidophilum. FEBS Lett. 1997, 404, 263–268. [Google Scholar] [CrossRef]
- León, A.; McKearin, D. Identification of TER94, an AAA ATPase protein, as a Bam-dependent component of the Drosophila fusome. Mol. Biol. Cell 1999, 10, 3825–3834. [Google Scholar] [CrossRef] [PubMed]
- Kobakhidze, G.; Sethi, A.; Valimehr, S.; Ralph, S.A.; Rouiller, I. The AAA+ ATPase p97 as a novel parasite and tuberculosis drug target. Trends Parasitol. 2022, 38, 572–590. [Google Scholar] [CrossRef]
- Huryn, D.M.; Kornfilt, D.J.P.; Wipf, P. p97: An Emerging Target for Cancer, Neurodegenerative Diseases, and Viral Infections. J. Med. Chem. 2019, 63, 1892–1907. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Dudley, J.P. How Viruses Use the VCP/p97 ATPase Molecular Machine. Viruses 2021, 13, 1881. [Google Scholar] [CrossRef] [PubMed]
- Buchberger, A.; Schindelin, H.; Hänzelmann, P. Control of p97 function by cofactor binding. FEBS Lett. 2015, 589, 2578–2589. [Google Scholar] [CrossRef]
- Zhang, X.; Gui, L.; Zhang, X.; Bulfer, S.L.; Sanghez, V.; Wong, D.E.; Lee, Y.; Lehmann, L.; Lee, J.S.; Shih, P.Y.; et al. Altered cofactor regulation with disease-associated p97/VCP mutations. Proc. Natl. Acad. Sci. USA 2015, 112, E1705–E1714. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.; Maksim, N.; De la Cruz, F.; La Clair, J.J. Inhibitors of the AAA+ chaperone p97. Molecules 2015, 20, 3027–3049. [Google Scholar] [CrossRef]
- Latterich, M.; Fröhlich, K.-U.; Schekman, R. Membrane fusion and the cell cycle: Cdc48p participates in the fusion of ER membranes. Cell 1995, 82, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, K.U.; Fries, H.W.; Rüdiger, M.; Erdmann, R.; Botstein, D.; Mecke, D. Yeast cell cycle protein CDC48p shows full-length homology to the mammalian protein VCP and is a member of a protein family involved in secretion, peroxisome formation, and gene expression. J. Cell Biol. 1991, 114, 443–453. [Google Scholar] [CrossRef]
- Rabouille, C.; Levine, T.P.; Peters, J.-M.; Warren, G. An NSF-like ATPase, p97, and NSF mediate cisternal regrowth from mitotic golgi fragments. Cell 1995, 82, 905–914. [Google Scholar] [CrossRef]
- Xu, S.; Peng, G.; Wang, Y.; Fang, S.; Karbowski, M. The AAA-ATPase p97 is essential for outer mitochondrial membrane protein turnover. Mol. Biol. Cell 2011, 22, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Vaz, B.; Halder, S.; Ramadan, K. Role of p97/VCP (Cdc48) in genome stability. Front Genet. 2013, 4, 60. [Google Scholar] [CrossRef] [PubMed]
- Kochenova, O.V.; Mukkavalli, S.; Raman, M.; Walter, J.C. Cooperative assembly of p97 complexes involved in replication termination. Nat. Commun. 2022, 13, 6591. [Google Scholar] [CrossRef]
- Rabinovich, E.; Kerem, A.; Fröhlich, K.U.; Diamant, N.; Bar-Nun, S. AAA-ATPase p97/Cdc48p, a cytosolic chaperone required for endoplasmic reticulum-associated protein degradation. Mol. Cell. Biol. 2002, 22, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Halawani, D.; Latterich, M. p97: The cell’s molecular purgatory? Mol. Cell 2006, 22, 713–717. [Google Scholar] [CrossRef]
- Alexandru, G.; Graumann, J.; Smith, G.T.; Kolawa, N.J.; Fang, R.; Deshaies, R.J. UBXD7 Binds Multiple Ubiquitin Ligases and Implicates p97 in HIF1α Turnover. Cell 2008, 134, 804–816. [Google Scholar] [CrossRef]
- Bodnar, N.; Rapoport, T. Toward an understanding of the Cdc48/p97 ATPase. F1000Research 2017, 6, 1318. [Google Scholar] [CrossRef]
- Olszewski, M.M.; Williams, C.; Dong, K.C.; Martin, A. The Cdc48 unfoldase prepares well-folded protein substrates for degradation by the 26S proteasome. Commun. Biol. 2019, 2, 29. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, N.O.; Rapoport, T.A. Molecular Mechanism of Substrate Processing by the Cdc48 ATPase Complex. Cell 2017, 169, 722–735.e9. [Google Scholar] [CrossRef]
- Blythe, E.E.; Olson, K.C.; Chau, V.; Deshaies, R.J. Ubiquitin- and ATP-dependent unfoldase activity of P97/VCP*NPLOC4*UFD1L is enhanced by a mutation that causes multisystem proteinopathy. Proc. Natl. Acad. Sci. USA 2017, 114, E4380–E4388. [Google Scholar] [CrossRef]
- Rouiller, I.; Butel, V.M.; Latterich, M.; Milligan, R.A.; Wilson-Kubalek, E.M. A Major Conformational Change in p97 AAA ATPase upon ATP Binding. Mol. Cell 2000, 6, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- Rouiller, I.; DeLaBarre, B.; May, A.P.; Weis, W.I.; Brunger, A.T.; Milligan, R.A.; Wilson-Kubalek, E.M. Conformational changes of the multifunction p97 AAA ATPase during its ATPase cycle. Nat. Struct. Biol. 2002, 9, 950–957. [Google Scholar] [CrossRef]
- DeLaBarre, B.; Brunger, A.T. Complete structure of p97/valosin-containing protein reveals communication between nucleotide domains. Nat. Struct. Biol. 2003, 10, 856–863. [Google Scholar] [CrossRef]
- Banerjee, S.; Bartesaghi, A.; Merk, A.; Rao, P.; Bulfer, S.L.; Yan, Y.; Green, N.; Mroczkowski, B.; Neitz, R.J.; Wipf, P.; et al. 2.3 Å resolution cryo-EM structure of human p97 and mechanism of allosteric inhibition. Science 2016, 351, 871–875. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.M.; Brunger, A.; Weis, W.I. Improved Structures of Full-Length p97, an AAA ATPase: Implications for Mechanisms of Nucleotide-Dependent Conformational Change. Structure 2008, 16, 715–726. [Google Scholar] [CrossRef]
- Hänzelmann, P.; Schindelin, H. Structural Basis of ATP Hydrolysis and Intersubunit Signaling in the AAA+ ATPase p97. Structure 2016, 24, 127–139. [Google Scholar] [CrossRef]
- Wendler, P.; Ciniawsky, S.; Kock, M.; Kube, S. Structure and function of the AAA+ nucleotide binding pocket. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2012, 1823, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.; Carter, A.P. Review: Structure and mechanism of the dynein motor ATPase. Biopolymers 2016, 105, 557–567. [Google Scholar] [CrossRef]
- Huang, C.; Li, G.; Lennarz, W.J. Dynamic flexibility of the ATPase p97 is important for its interprotomer motion transmission. Proc. Natl. Acad. Sci. USA 2012, 109, 9792–9797. [Google Scholar] [CrossRef]
- DeLaBarre, B.; Christianson, J.C.; Kopito, R.R.; Brunger, A.T. Central pore residues mediate the p97/VCP activity required for ERAD. Mol. Cell 2006, 22, 451–462. [Google Scholar] [CrossRef]
- Cooney, I.; Han, H.; Stewart, M.G.; Carson, R.H.; Hansen, D.T.; Iwasa, J.H.; Price, J.C.; Hill, C.P.; Shen, P.S. Structure of the Cdc48 segregase in the act of unfolding an authentic substrate. Science 2019, 365, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Twomey, E.C.; Ji, Z.; Wales, T.E.; Bodnar, N.O.; Ficarro, S.B.; Marto, J.A.; Engen, J.R.; Rapoport, T.A. Substrate processing by the Cdc48 ATPase complex is initiated by ubiquitin unfolding. Science 2019, 365, eaax1033. [Google Scholar] [CrossRef]
- Pan, M.; Yu, Y.; Ai, H.; Zheng, Q.; Xie, Y.; Liu, L.; Zhao, M. Mechanistic insight into substrate processing and allosteric inhibition of human p97. Nat. Struct. Mol. Biol. 2021, 28, 614–625. [Google Scholar] [CrossRef]
- Song, C.; Wang, Q.; Li, C.-C.H. ATPase Activity of p97-Valosin-containing Protein (VCP). J. Biol. Chem. 2003, 278, 3648–3655. [Google Scholar] [CrossRef]
- Briggs, L.C.; Baldwin, G.; Miyata, N.; Kondo, H.; Zhang, X.; Freemont, P.S. Analysis of Nucleotide Binding to P97 Reveals the Properties of a Tandem AAA Hexameric ATPase. J. Biol. Chem. 2008, 283, 13745–13752. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Huang, C.; Zhao, G.; Lennarz, W.J. Interprotomer motion-transmission mechanism for the hexameric AAA ATPase p97. Proc. Natl. Acad. Sci. USA 2012, 109, 3737–3741. [Google Scholar] [CrossRef]
- Tang, W.K.; Xia, D. Role of the D1-D2 Linker of Human VCP/p97 in the Asymmetry and ATPase Activity of the D1-domain. Sci. Rep. 2016, 6, 20037. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.K.; Li, D.; Li, C.C.; Esser, L.; Dai, R.; Guo, L.; Xia, D. A novel ATP-dependent conformation in p97 N–D1 fragment revealed by crystal structures of disease-related mutants. EMBO J. 2010, 29, 2217–2229. [Google Scholar] [CrossRef]
- Schuller, J.M.; Beck, F.; Lössl, P.; Heck, A.J.R.; Förster, F. Nucleotide-dependent conformational changes of the AAA+ ATPase p97 revisited. FEBS Lett. 2016, 590, 595–604. [Google Scholar] [CrossRef]
- Conicella, A.E.; Huang, R.; Ripstein, Z.A.; Nguyen, A.; Wang, E.; Löhr, T.; Schuck, P.; Vendruscolo, M.; Rubinstein, J.L.; Kay, L.E. An intrinsically disordered motif regulates the interaction between the p47 adaptor and the p97 AAA+ ATPase. Proc. Natl. Acad. Sci. USA 2020, 117, 26226–26236. [Google Scholar] [CrossRef]
- Yu, G.; Bai, Y.; Li, K.; Amarasinghe, O.; Jiang, W.; Zhang, Z.-Y. Cryo-electron microscopy structures of VCP/p97 reveal a new mechanism of oligomerization regulation. iScience 2021, 24, 103310. [Google Scholar] [CrossRef]
- Gao, H.; Li, F.; Ji, Z.; Shi, Z.; Li, Y.; Yu, H. Cryo-EM structures of human p97 double hexamer capture potentiated ATPase-competent state. Cell Discov. 2022, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Han, H.; Cooney, I.; Guo, Y.; Moran, N.G.; Zuniga, N.R.; Price, J.C.; Hill, C.P.; Shen, P.S. Active conformation of the p97-p47 unfoldase complex. Nat. Commun. 2022, 13, 2640. [Google Scholar] [CrossRef]
- Gates, S.N.; Martin, A. Stairway to translocation: AAA+ motor structures reveal the mechanisms of ATP-dependent substrate translocation. Protein Sci. 2020, 29, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Yeung, H.O.; Kloppsteck, P.; Niwa, H.; Isaacson, R.L.; Matthews, S.; Zhang, X.; Freemont, P.S. Insights into adaptor binding to the AAA protein p97. Biochem. Soc. Trans. 2008, 36, 62–67. [Google Scholar] [CrossRef]
- Hänzelmann, P.; Buchberger, A.; Schindelin, P.H.B. Hierarchical Binding of Cofactorsto the AAA ATPase p97. Structure 2011, 19, 833–843. [Google Scholar] [CrossRef]
- Kern, M.; Fernandez-Sáiz, V.; Schäfer, Z.; Buchberger, A. UBXD1 binds p97 through two independent binding sites. Biochem. Biophys. Res. Commun. 2009, 380, 303–307. [Google Scholar] [CrossRef]
- Bruderer, R.M.; Brasseur, C.; Meyer, H.H. The AAA ATPase p97/VCP interacts with its alternative co-factors, Ufd1-Npl4 and p47, through a common bipartite binding mechanism. J. Biol. Chem. 2004, 279, 49609–49616. [Google Scholar] [CrossRef] [PubMed]
- Beuron, F.; Dreveny, I.; Yuan, X.; Pye, V.E.; McKeown, C.; Briggs, L.C.; Cliff, M.; Kaneko, Y.; Wallis, R.; Isaacson, R.L.; et al. Conformational changes in the AAA ATPase p97–p47 adaptor complex. EMBO J. 2006, 25, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Rabouille, C.; Newman, R.; Levine, T.P.; Pappin, D.; Freemont, P.; Warren, G. p47 is a cofactor for p97-mediated membrane fusion. Nature 1997, 388, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Madsen, L.; Andersen, K.M.; Prag, S.; Moos, T.; Semple, C.A.; Seeger, M.; Hartmann-Petersen, R. Ubxd1 is a novel co-factor of the human p97 ATPase. Int. J. Biochem. Cell Biol. 2008, 40, 2927–2942. [Google Scholar] [CrossRef]
- Schuetz, A.K.; Kay, L.E. A Dynamic molecular basis for malfunction in disease mutants of p97/VCP. eLife 2016, 5, e20143. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.; Zheng, Q.; Yu, Y.; Ai, H.; Xie, Y.; Zeng, X.; Wang, C.; Liu, L.; Zhao, M. Seesaw conformations of Npl4 in the human p97 complex and the inhibitory mechanism of a disulfiram derivative. Nat. Commun. 2021, 12, 121. [Google Scholar] [CrossRef]
- Stach, L.; Freemont, P.S. The AAA+ ATPase p97, a cellular multitool. Biochem. J. 2017, 474, 2953–2976. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Meyer, H.H.; Rapoport, T.A. The AAA ATPase Cdc48/p97 and its partners transport proteins from the ER into the cytosol. Nature 2001, 414, 652–656. [Google Scholar] [CrossRef]
- Meyer, H.H.; Wang, Y.; Warren, G. Direct binding of ubiquitin conjugates by the mammalian p97 adaptor complexes, p47 and Ufd1–Npl4. EMBO J. 2002, 21, 5645–5652. [Google Scholar] [CrossRef] [PubMed]
- Weith, M.; Seiler, J.; van den Boom, J.; Kracht, M.; Hülsmann, J.; Primorac, I.; Garcia, J.D.P.; Kaschani, F.; Kaiser, M.; Musacchio, A.; et al. Ubiquitin-Independent Disassembly by a p97 AAA-ATPase Complex Drives PP1 Holoenzyme Formation. Mol. Cell 2018, 72, 766–777.e6. [Google Scholar] [CrossRef]
- Hanzelmann, P.; Schindelin, H. The structural and functional basis of the p97/valosin-containing protein (VCP)-interacting motif (VIM): Mutually exclusive binding of cofactors to the N-terminal domain of p97. J. Biol. Chem. 2011, 286, 38679–38690. [Google Scholar] [CrossRef]
- Dreveny, I.; Kondo, H.; Uchiyama, K.; Shaw, A.; Zhang, X.; Freemont, P.S. Structural basis of the interaction between the AAA ATPase p97/VCP and its adaptor protein p47. EMBO J. 2004, 23, 1030–1039. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Yang, J.K. Crystallization and preliminary X-ray crystallographic analysis of the hexameric human p97/VCP ND1 fragment in complex with the UBX domain of human FAF1. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67 Pt 10, 1199–1202. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.S.; Miller, S.E.; Hanson, P.I.; Weihl, C.C. Impaired protein aggregate handling and clearance underlie the pathogenesis of p97/VCP-associated disease. J. Biol. Chem. 2008, 283, 30289–30299. [Google Scholar] [CrossRef] [PubMed]
- Al-Tahan, S.; Al-Obeidi, E.; Yoshioka, H.; Lakatos, A.; Weiss, L.; Grafe, M.; Palmio, J.; Wicklund, M.; Harati, Y.; Omizo, M.; et al. Novel valosin-containing protein mutations associated with multisystem proteinopathy. Neuromuscul. Disord. 2018, 28, 491–501. [Google Scholar] [CrossRef]
- Halawani, D.; LeBlanc, A.C.; Rouiller, I.; Michnick, S.W.; Servant, M.J.; Latterich, M. Hereditary inclusion body myopathy-linked p97/VCP mutations in the NH2 domain and the D1 ring modulate p97/VCP ATPase activity and D2 ring conformation. Mol. Cell. Biol. 2009, 29, 4484–4494. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.K.; Xia, D. Altered Intersubunit Communication Is the Molecular Basis for Functional Defects of Pathogenic p97 Mutants. J. Biol. Chem. 2013, 288, 36624–36635. [Google Scholar] [CrossRef]
- Schütz, A.K.; Rennella, E.; Kay, L.E. Exploiting conformational plasticity in the AAA+ protein VCP/p97 to modify function. Proc. Natl. Acad. Sci. USA 2017, 114, E6822–E6829. [Google Scholar] [CrossRef] [PubMed]
- Caffrey, B.; Zhu, X.; Berezuk, A.; Tuttle, K.; Chittori, S.; Subramaniam, S. AAA+ ATPase p97/VCP mutants and inhibitor binding disrupt inter-domain coupling and subsequent allosteric activation. J. Biol. Chem. 2021, 297, 101187. [Google Scholar] [CrossRef] [PubMed]
- Huyton, T.; Pye, V.E.; Briggs, L.C.; Flynn, T.C.; Beuron, F.; Kondo, H.; Ma, J.; Zhang, X.; Freemont, P.S. The crystal structure of murine p97/VCP at 3.6A. J. Struct. Biol. 2003, 144, 337–348. [Google Scholar] [CrossRef]
- Zhang, X.; Shaw, A.; Bates, P.A.; Newman, R.H.; Gowen, B.; Orlova, E.; Gorman, M.A.; Kondo, H.; Dokurno, P.; Lally, J.; et al. Structure of the AAA ATPase p97. Mol. Cell 2000, 6, 1473–1484. [Google Scholar] [CrossRef]
- Blythe, E.; Gates, S.N.; Deshaies, R.J.; Martin, A. Multisystem Proteinopathy Mutations in VCP/p97 Increase NPLOC4·UFD1L Binding and Substrate Processing. Structure 2019, 27, 1820–1829.e4. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Ripstein, Z.A.; Rubinstein, J.L.; Kay, L.E. Cooperative subunit dynamics modulate p97 function. Proc. Natl. Acad. Sci. USA 2019, 116, 158–167. [Google Scholar] [CrossRef]
- Bulfer, S.L.; Chou, T.-F.; Arkin, M.R. p97 Disease Mutations Modulate Nucleotide-Induced Conformation to Alter Protein–Protein Interactions. ACS Chem. Biol. 2016, 11, 2112–2116. [Google Scholar] [CrossRef]
- Fernández-Sáiz, V.; Buchberger, A. Imbalances in p97 co-factor interactions in human proteinopathy. EMBO Rep. 2010, 11, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Mountassif, D.; Fabre, L.; Zaid, Y.; Halawani, D.; Rouiller, I. Cryo-EM of the pathogenic VCP variant R155P reveals long-range conformational changes in the D2 ATPase ring. Biochem. Biophys. Res. Commun. 2015, 468, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Ritz, D.; Vuk, M.; Kirchner, P.; Bug, M.; Schütz, S.; Hayer, A.; Bremer, S.; Lusk, C.; Baloh, R.H.; Lee, H.; et al. Endolysosomal sorting of ubiquitylated caveolin-1 is regulated by VCP and UBXD1 and impaired by VCP disease mutations. Nat. Cell Biol. 2011, 13, 1116–1123. [Google Scholar] [CrossRef]
- Cheng, C.; Weiss, L.; Leinonen, H.; Shmara, A.; Yin, H.Z.; Ton, T.; Do, A.; Lee, J.; Ta, L.; Mohanty, E.; et al. VCP/p97 inhibitor CB-5083 modulates muscle pathology in a mouse model of VCP inclusion body myopathy. J. Transl. Med. 2022, 20, 21. [Google Scholar] [CrossRef]
- Tang, W.K.; Xia, D. Mutations in the Human AAA+ Chaperone p97 and Related Diseases. Front. Mol. Biosci. 2016, 3, 79. [Google Scholar] [CrossRef] [PubMed]
- Costantini, S.; Capone, F.; Polo, A.; Bagnara, P.; Budillon, A. Valosin-Containing Protein (VCP)/p97: A Prognostic Biomarker and Therapeutic Target in Cancer. Int. J. Mol. Sci. 2021, 22, 10177. [Google Scholar] [CrossRef]
- Zhang, G.; Li, S.; Cheng, K.-W.; Chou, T.-F. AAA ATPases as therapeutic targets: Structure, functions, and small-molecule inhibitors. Eur. J. Med. Chem. 2021, 219, 113446. [Google Scholar] [CrossRef]
- Zhou, H.-J.; Wang, J.; Yao, B.; Wong, S.; Djakovic, S.; Kumar, B.; Rice, J.; Valle, E.; Soriano, F.; Menon, M.-K.; et al. Discovery of a First-in-Class, Potent, Selective, and Orally Bioavailable Inhibitor of the p97 AAA ATPase (CB-5083). J. Med. Chem. 2015, 58, 9480–9497. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Shinkre, B.A.; Lee, J.G.; Weniger, M.A.; Liu, Y.; Chen, W.; Wiestner, A.; Trenkle, W.C.; Ye, Y. The ERAD inhibitor Eeyarestatin I is a bifunctional compound with a membrane-binding domain and a p97/VCP inhibitory group. PLoS ONE 2010, 5, e15479. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.-F.; Brown, S.J.; Minond, D.; Nordin, B.E.; Li, K.; Jones, A.C.; Chase, P.; Porubsky, P.R.; Stoltz, B.M.; Schoenen, F.J.; et al. Reversible inhibitor of p97, DBeQ, impairs both ubiquitin-dependent and autophagic protein clearance pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 4834–4839. [Google Scholar] [CrossRef]
- Tang, W.K.; Odzorig, T.; Jin, W.; Xia, D. Structural Basis of p97 Inhibition by the Site-Selective Anti-Cancer Compound CB-5083. Mol. Pharmacol. 2018, 95, 286–293. [Google Scholar] [CrossRef]
- Chou, T.F.; Bulfer, S.L.; Weihl, C.C.; Li, K.; Lis, L.G.; Walters, M.A.; Schoenen, F.J.; Lin, H.J.; Deshaies, R.J.; Arkin, M.R. Specific inhibition of p97/VCP ATPase and kinetic analysis demonstrate interaction between D1 and D2 ATPase domains. J. Mol. Biol. 2014, 426, 2886–2899. [Google Scholar] [CrossRef]
- Chou, T.-F.; Li, K.; Frankowski, K.J.; Schoenen, F.J.; Deshaies, R.J. Structure–Activity Relationship Study Reveals ML240 and ML241 as Potent and Selective Inhibitors of p97 ATPase. ChemMedChem 2013, 8, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Alverez, C.; Arkin, M.R.; Bulfer, S.L.; Colombo, R.; Kovaliov, M.; LaPorte, M.G.; Lim, C.; Liang, M.; Moore, W.J.; Neitz, R.J.; et al. Structure–Activity Study of Bioisosteric Trifluoromethyl and Pentafluorosulfanyl Indole Inhibitors of the AAA ATPase p97. ACS Med. Chem. Lett. 2015, 6, 1225–1230. [Google Scholar] [CrossRef]
- Magnaghi, P.; D’Alessio, R.; Valsasina, B.; Avanzi, N.; Rizzi, S.; Asa, D.; Gasparri, F.; Cozzi, L.; Cucchi, U.; Orrenius, C.; et al. Covalent and allosteric inhibitors of the ATPase VCP/p97 induce cancer cell death. Nat. Chem. Biol. 2013, 9, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.J.; Le Moigne, R.; Djakovic, S.; Kumar, B.; Rice, J.; Wong, S.; Wang, J.; Yao, B.; Valle, E.; von Soly, S.K.; et al. Targeting the AAA ATPase p97 as an Approach to Treat Cancer through Disruption of Protein Homeostasis. Cancer Cell 2015, 28, 653–665. [Google Scholar] [CrossRef]
- Leinonen, H.; Cheng, C.; Pitkänen, M.; Sander, C.L.; Zhang, J.; Saeid, S.; Turunen, T.; Shmara, A.; Weiss, L.; Ton, T.; et al. A p97/Valosin-Containing Protein Inhibitor Drug CB-5083 Has a Potent but Reversible Off-Target Effect on Phosphodiesterase-6. J. Pharmacol. Exp. Ther. 2021, 378, 31–41. [Google Scholar] [CrossRef]
- Roux, B.; Vaganay, C.; Vargas, J.D.; Alexe, G.; Benaksas, C.; Pardieu, B.; Fenouille, N.; Ellegast, J.M.; Malolepsza, E.; Ling, F.; et al. Targeting acute myeloid leukemia dependency on VCP-mediated DNA repair through a selective second-generation small-molecule inhibitor. Sci. Transl. Med. 2021, 13, eabg1168. [Google Scholar] [CrossRef] [PubMed]
- Casi Pharmaceutical. Available online: https://www.casipharmaceuticals.com/product-pipeline/cb-5339 (accessed on 12 April 2023).
- Wang, F.; Li, S.; Gan, T.; Stott, G.M.; Flint, A.; Chou, T. Allosteric p97 Inhibitors Can Overcome Resistance to ATP-Competitive p97 Inhibitors for Potential Anticancer Therapy. ChemMedChem 2020, 15, 685–694. [Google Scholar] [CrossRef]
- Wang, Q.; Li, L.; Ye, Y. Inhibition of p97-dependent Protein Degradation by Eeyarestatin I. J. Biol. Chem. 2008, 283, 7445–7454. [Google Scholar] [CrossRef]
- Wang, Q.; Mora-Jensen, H.; Weniger, M.A.; Perez-Galan, P.; Wolford, C.; Hai, T.; Ron, D.; Chen, W.; Trenkle, W.; Wiestner, A.; et al. ERAD inhibitors integrate ER stress with an epigenetic mechanism to activate BH3-only protein NOXA in cancer cells. Proc. Natl. Acad. Sci. USA 2009, 106, 2200–2205. [Google Scholar] [CrossRef]
- LaPorte, M.G.; Burnett, J.C.; Colombo, R.; Bulfer, S.L.; Alverez, C.; Chou, T.-F.; Neitz, R.J.; Green, N.; Moore, W.J.; Yue, Z.; et al. Optimization of Phenyl Indole Inhibitors of the AAA+ ATPase p97. ACS Med. Chem. Lett. 2018, 9, 1075–1081. [Google Scholar] [CrossRef]
- Pöhler, R.; Krahn, J.H.; van den Boom, J.; Dobrynin, G.; Kaschani, F.; Eggenweiler, H.-M.; Zenke, F.T.; Kaiser, M.; Meyer, H. A Non-Competitive Inhibitor of VCP/p97 and VPS4 Reveals Conserved Allosteric Circuits in Type I and II AAA ATPases. Angew. Chem. Int. Ed. 2018, 57, 1576–1580. [Google Scholar] [CrossRef]
- Figuerola-Conchas, A.; Saarbach, J.; Daguer, J.-P.; Cieren, A.; Barluenga, S.; Winssinger, N.; Gotta, M. Small-Molecule Modulators of the ATPase VCP/p97 Affect Specific p97 Cellular Functions. ACS Chem. Biol. 2019, 15, 243–253. [Google Scholar] [CrossRef]
- Sasazawa, Y.; Kanagaki, S.; Tashiro, E.; Nogawa, T.; Muroi, M.; Kondoh, Y.; Osada, H.; Imoto, M. Xanthohumol Impairs Autophagosome Maturation through Direct Inhibition of Valosin-Containing Protein. ACS Chem. Biol. 2012, 7, 892–900. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Bienfait, B.; Ertl, P. JSME: A free molecule editor in JavaScript. J. Chemin. 2013, 5, 24. [Google Scholar] [CrossRef] [PubMed]
- Schuttelkopf, A.W.; van Aalten, D.M. PRODRG: A tool for high-throughput crystallography of protein-ligand complexes. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 1355–1363. [Google Scholar] [CrossRef]
- Regensburger, M.; Türk, M.; Pagenstecher, A.; Schröder, R.; Winkler, J. VCP-related multisystem proteinopathy presenting as early-onset Parkinson disease. Neurology 2017, 89, 746–748. [Google Scholar] [CrossRef]
- Adams, J. The development of proteasome inhibitors as anticancer drugs. Cancer Cell 2004, 5, 417–421. [Google Scholar] [CrossRef]
- Vij, N. AAA ATPase p97/VCP: Cellular functions, disease and therapeutic potential. J. Cell. Mol. Med. 2008, 12, 2511–2518. [Google Scholar] [CrossRef] [PubMed]
- Kale, A.J.; Moore, B.S. Molecular Mechanisms of Acquired Proteasome Inhibitor Resistance. J. Med. Chem. 2012, 55, 10317–10327. [Google Scholar] [CrossRef] [PubMed]
- Bursavich, M.G.; Parker, D.P.; Willardsen, J.A.; Gao, Z.-H.; Davis, T.; Ostanin, K.; Robinson, R.; Peterson, A.; Cimbora, D.M.; Zhu, J.-F.; et al. 2-Anilino-4-aryl-1,3-thiazole inhibitors of valosin-containing protein (VCP or p97). Bioorg. Med. Chem. Lett. 2010, 20, 1677–1679. [Google Scholar] [CrossRef] [PubMed]
- Deshaies, R.J. Proteotoxic crisis, the ubiquitin-proteasome system, and cancer therapy. BMC Biol. 2014, 12, 94. [Google Scholar] [CrossRef]
- Chou, T.-F.; Deshaies, R.J. Development of p97 AAA ATPase inhibitors. Autophagy 2011, 7, 1091–1092. [Google Scholar] [CrossRef] [PubMed]
- Richardson, P.G.; Mitsiades, C.; Hideshima, T.; Anderson, K.C. Bortezomib: Proteasome Inhibition as an Effective Anticancer Therapy. Annu. Rev. Med. 2006, 57, 33–47. [Google Scholar] [CrossRef]
- Adams, J.; Kauffman, M. Development of the Proteasome Inhibitor Velcade™ (Bortezomib). Cancer Investig. 2004, 22, 304–311. [Google Scholar] [CrossRef]
- Kuhn, D.J.; Chen, Q.; Voorhees, P.M.; Strader, J.S.; Shenk, K.D.; Sun, C.M.; Demo, S.D.; Bennett, M.K.; van Leeuwen, F.W.B.; Chanan-Khan, A.A.; et al. Potent activity of carfilzomib, a novel, irreversible inhibitor of the ubiquitin-proteasome pathway, against preclinical models of multiple myeloma. Blood 2007, 110, 3281–3290. [Google Scholar] [CrossRef]
- Parlati, F.; Lee, S.J.; Aujay, M.; Suzuki, E.; Levitsky, K.; Lorens, J.B.; Micklem, D.R.; Ruurs, P.; Sylvain, C.; Lu, Y.; et al. Carfilzomib can induce tumor cell death through selective inhibition of the chymotrypsin-like activity of the proteasome. Blood 2009, 114, 3439–3447. [Google Scholar] [CrossRef] [PubMed]
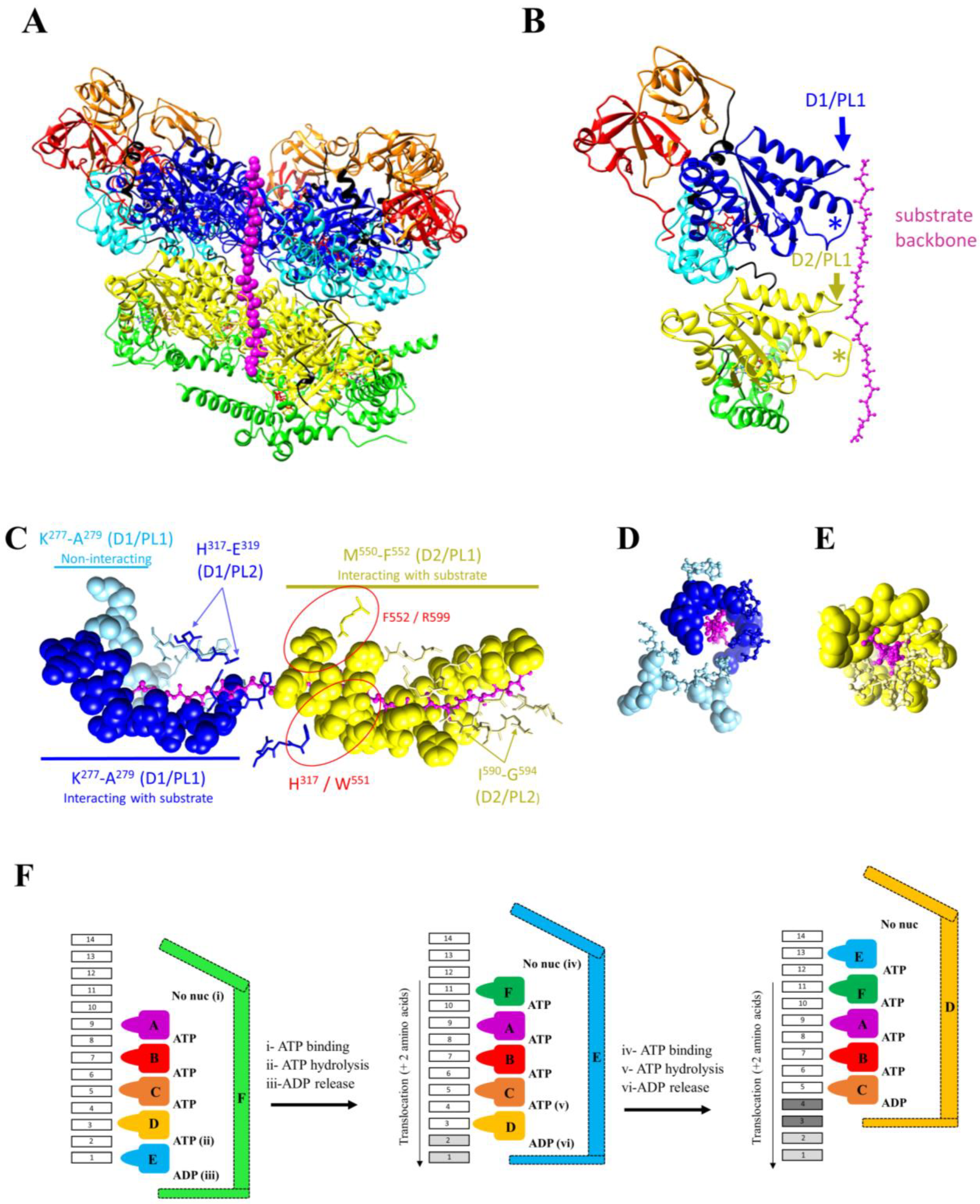
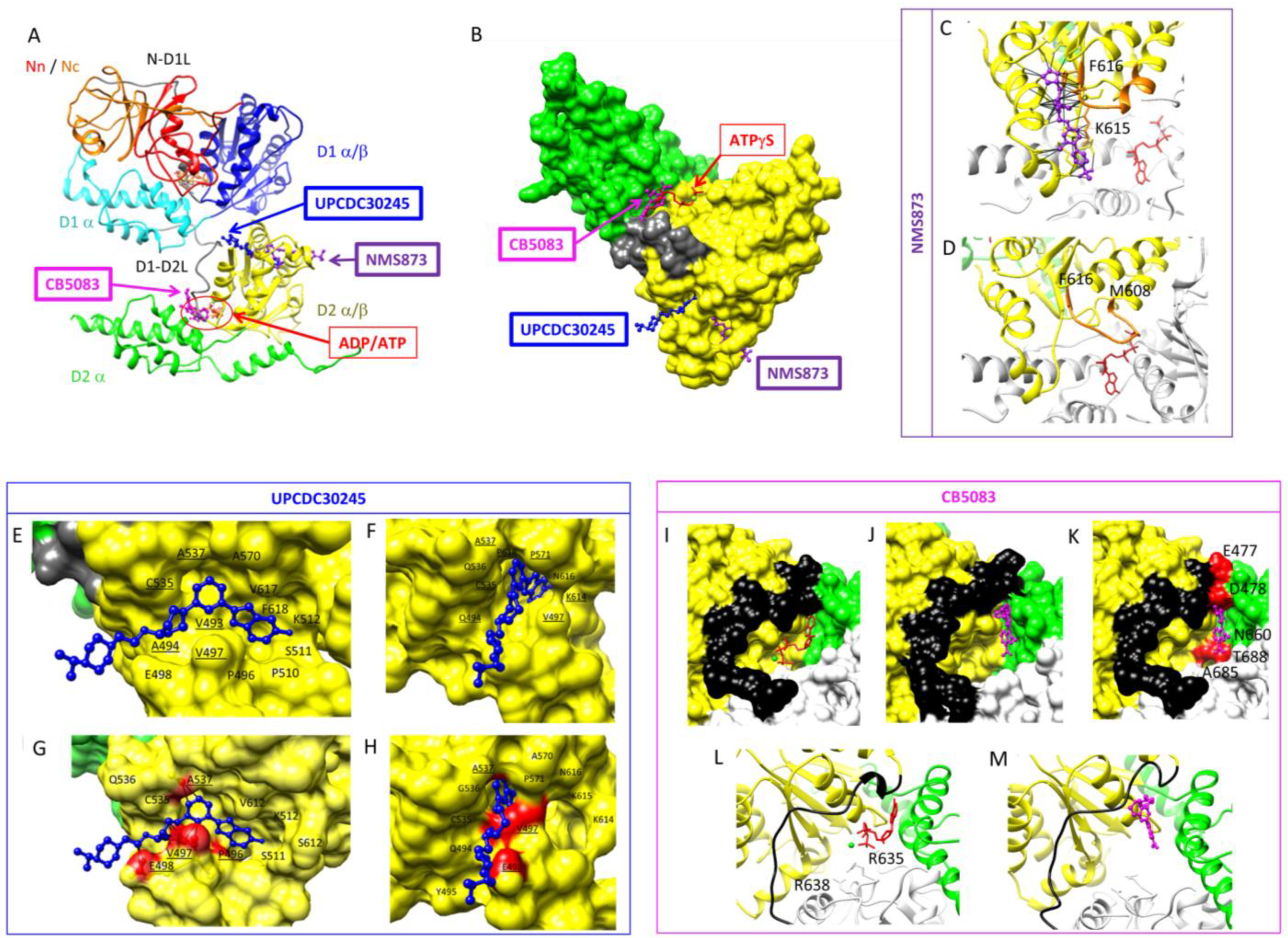
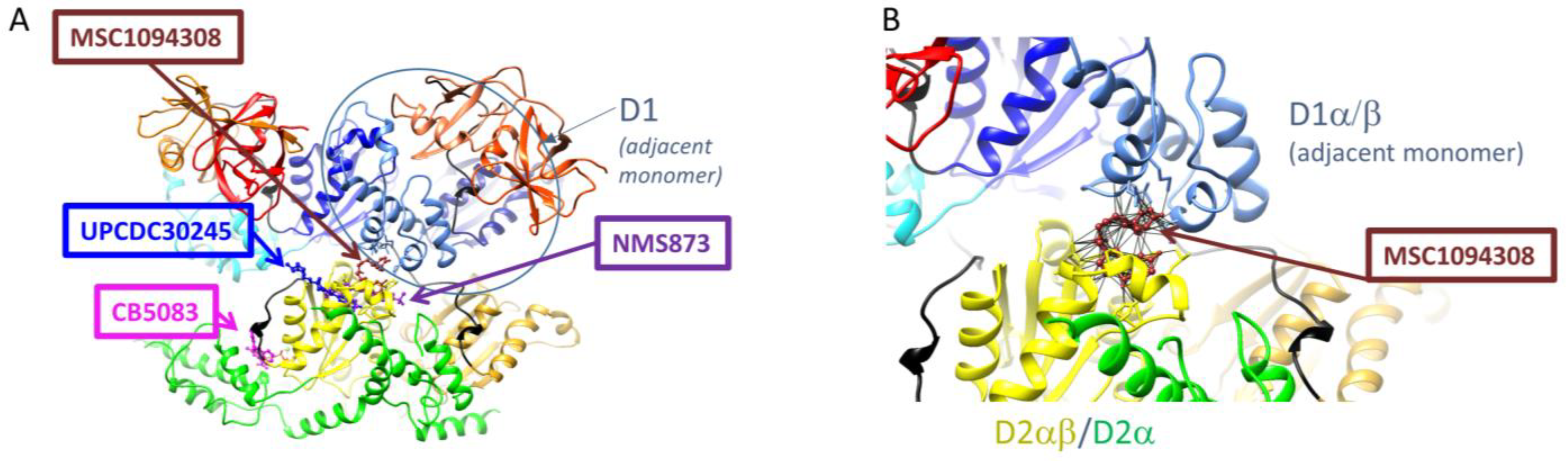

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Construct | PDB/ EMD ID | Method | Resolution (Å) | Ligand D1/D2 | Position of N | Ref. Year |
|---|---|---|---|---|---|---|
| FL | 3CF1 | X-ray | 4.40 | ADP/ADP-ALFx | Down | [30,32] 2003-08 |
| FL | 3CF3 | X-ray | 4.25 | ADP/ADP | Down | [30,32] 2003-08 |
| FL | 3CF2 | X-ray | 3.50 | ADP/ AMP-PNP | Down | [30,32] 2003-08 |
| FL | 1R7R | X-ray | 3.60 | ADP/- | Down | [74] 2003 |
| FL (Δ709–728) | 5C18 | X-ray | 3.30 | ATPγS/ATPγS | Down | [33] 2016 |
| FL (α9-D4) * | 5C19 | X-ray | 4.20 | -/- | Down | [33] 2016 |
| FL (α9-D4) * | 5C1A | X-ray | 3.80 | ATPγS/ATPγS | Down | [33] 2016 |
| FL | 5FTK/3296 | EM | 2.4 | ADP/ADP | Down | [31] 2016 |
| FL | 5FTN/3299 | EM | 3.3 | ATPγS/ATPγS | Up | [31] 2016 |
| FL | 5FTL/3297 | EM | 3.3 | ADP/ADP | Down | [31] 2016 |
| FL | 5FTM/3298 | EM | 3.2 | ADP/ATPγS | Down | [31] 2016 |
| N-D1 | 1E32 | X-ray | 2.90 | ADP | Down | [75] 2001 |
| N-D1 | 5DYI | X-ray | 3.71 | ADP | Down | [71] 2015 |
| FL-Npl4/Ufd1 | 21824, 21825, 21826 | EM | 3.8 3.7 3.9 | [60] 2021 | ||
| FL | 7JY5/22521 | EM | 2.89 | ATPγS/ATPγS | [60] 2021 | |
| FL-Npl4/ Ufd1-Ub-Eos | 21827, 21828, 21829, 21830 | EM | 4.2 4.3 4.5 3.5 | [60] 2021 | ||
| FL-Npl4/Ufd1-Ub6-FOM, 3 classes | 7LMZ/23443 7LN0/23444 7LN1/23445 | EM | 3.06 2.98 3.40 | ATP/ATP ATP/ATP ATP/ATP | Up Up Up | [40] 2021 |
| FL-Npl4/Ufd1-Ubn-Eos-FOM, 3 classes | 7LN2/23446 7LN3/23447 7LN4/23448 | EM | 3.63 3.45 3.00 | ATP/ATP ATP/ATP ATP/ATP | Up Up Up | [40] 2021 |
| FL-Npl4/Ufd1-Ubn-Eos-CHAPSO, 2 classes | 7LN5/23449 7LN6/23450 23451 | EM | 3.09 3.58 3.77 | ATP/ATP ATP/ATP | Up Up | [40] 2021 |
| FL-Npl4/Ufd1-Ub6-NMS-873-FOM | 7LMY/23442 | EM | 2.40 | ATP/ATP | Up | [40] 2021 |
| FL-Npl4/Ufd1-Ub6-NMS-873-CHAPSO | 23452 23453 23454 23455 23456 23457 23458 | EM | 2.87 3.69 6.00 3.58 4.25 3.47 3.65 | [40] 2021 | ||
| Dodecamer FL/FL | 7VCS/31894 | EM | 3.32 | ATPγS/ATPγS ATPγS/ATPγS | Up/up | [49] 2022 |
| Dodecamer FL/FL | 7VCU/31896 | EM | 3.15 | ATPγS/ADP ATPγS/ADP | Down/ down | [49] 2022 |
| FL | 7VCT/31895 | EM | 3.21 | ATPγS/ADP | Down | [49] 2022 |
| FL | 7VCV/31897 | EM | 3.21 | ATPγS/ATPγS | Up | [49] 2022 |
| FL | 7VCX/31899 | EM | 3.24 | ATPγS/ATPγS | Down | [49] 2022 |
| Construct | PDB/ EMD ID | Method | Resolution (Å) | Mutation | Ligand D1/D2 | Position of N | Ref. Year |
|---|---|---|---|---|---|---|---|
| FL/UN | 20730 | EM | 4.26 | A232E | ? | Up | [76] 2019 |
| FL | - | EM | 3.7 | R95G | ? | Mixture | [77] 2019 |
| N-D1 | 4KO8 | X-ray | 1.98 | R155H | ATPγS(D1) | Up | [71] 2013 |
| N-D1 | 4KOD | X-ray | 2.96 | R155H | ADP(D1) | Down | [71] 2013 |
| N-D1 | 4KLN | X-ray | 2.62 | A232E | ATPγS(D1) | Up | [71] 2013 |
| N-D1 | 5DYG | X-ray | 2.20 | L198W | ADP (D1) | Down | [44] 2015 |
| N-D1 | 3HU1 | X-ray | 2.81 | R95G | ATPγS(D1) | Up | [45] 2010 |
| N-D1 | 3HU1 | X-ray | 2.85 | R86A | ATPγS(D1) | Up | [45] 2010 |
| N-D1 | 3HU3 | X-ray | 2.20 | R155H | ATPγS(D1) | Up | [45] 2010 |
| FL | 7RL6/24518 | EM | 3.7 | R155H | ADP (D1) ADP (D2) | Down | [73] 2021 |
| FL | 7RL7/24519 | EM | 3.0 | R155H | ATPγS (D1) ATPγS (D2) | Up | [73] 2021 |
| FL | 7RL9/24522 | EM | 3.3 | R191Q | ADP (D1) ADP (D2) | Up | [73] 2021 |
| FL | 7RLA/24523 | EM | 3.1 | R191Q | ATPγS (D1) ATPγS (D2) | Up | [73] 2021 |
| FL | 7RLB/24524 | EM | 3.3 | A232E | ADP (D1) ADP (D2) | Down | [73] 2021 |
| FL | 7RLC/24525 | EM | 3.2 | A232E | ATPγS (D1) ATPγS (D2) | Up | [73] 2021 |
| FL | 7RLD/24526 | EM | 3.4 | E470D | ADP (D1) ADP (D2) | Down | [73] 2021 |
| FL | 7RLF/24528 | EM | 3.1 | E470D | ATPγS (D1) ATPγS (D2) | Down | [73] 2021 |
| FL | 7RLG/24529 | EM | 3.7 | D592N | ADP (D1) ADP (D2) | Down | [73] 2021 |
| FL | 7RLH/24530 | EM | 3.7 | D592N | ATPγS (D1) ATPγS (D2 | Down | [73] 2021 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valimehr, S.; Sethi, A.; Shukla, M.; Bhattacharyya, S.; Kazemi, M.; Rouiller, I. Molecular Mechanisms Driving and Regulating the AAA+ ATPase VCP/p97, an Important Therapeutic Target for Treating Cancer, Neurological and Infectious Diseases. Biomolecules 2023, 13, 737. https://doi.org/10.3390/biom13050737
Valimehr S, Sethi A, Shukla M, Bhattacharyya S, Kazemi M, Rouiller I. Molecular Mechanisms Driving and Regulating the AAA+ ATPase VCP/p97, an Important Therapeutic Target for Treating Cancer, Neurological and Infectious Diseases. Biomolecules. 2023; 13(5):737. https://doi.org/10.3390/biom13050737
Chicago/Turabian StyleValimehr, Sepideh, Ashish Sethi, Manjari Shukla, Sudipta Bhattacharyya, Mohsen Kazemi, and Isabelle Rouiller. 2023. "Molecular Mechanisms Driving and Regulating the AAA+ ATPase VCP/p97, an Important Therapeutic Target for Treating Cancer, Neurological and Infectious Diseases" Biomolecules 13, no. 5: 737. https://doi.org/10.3390/biom13050737