
Omega-3 versus Omega-6: Are We Underestimating the Ecological Significance of Arachidonic Acid in Aquatic Systems?



Abstract
:1. Introduction
2. Methods
2.1. Cultivation of Organisms
2.2. Experimental Setup
2.3. Liposome Preparation
2.4. Chemical Analyses
2.5. Data Analysis
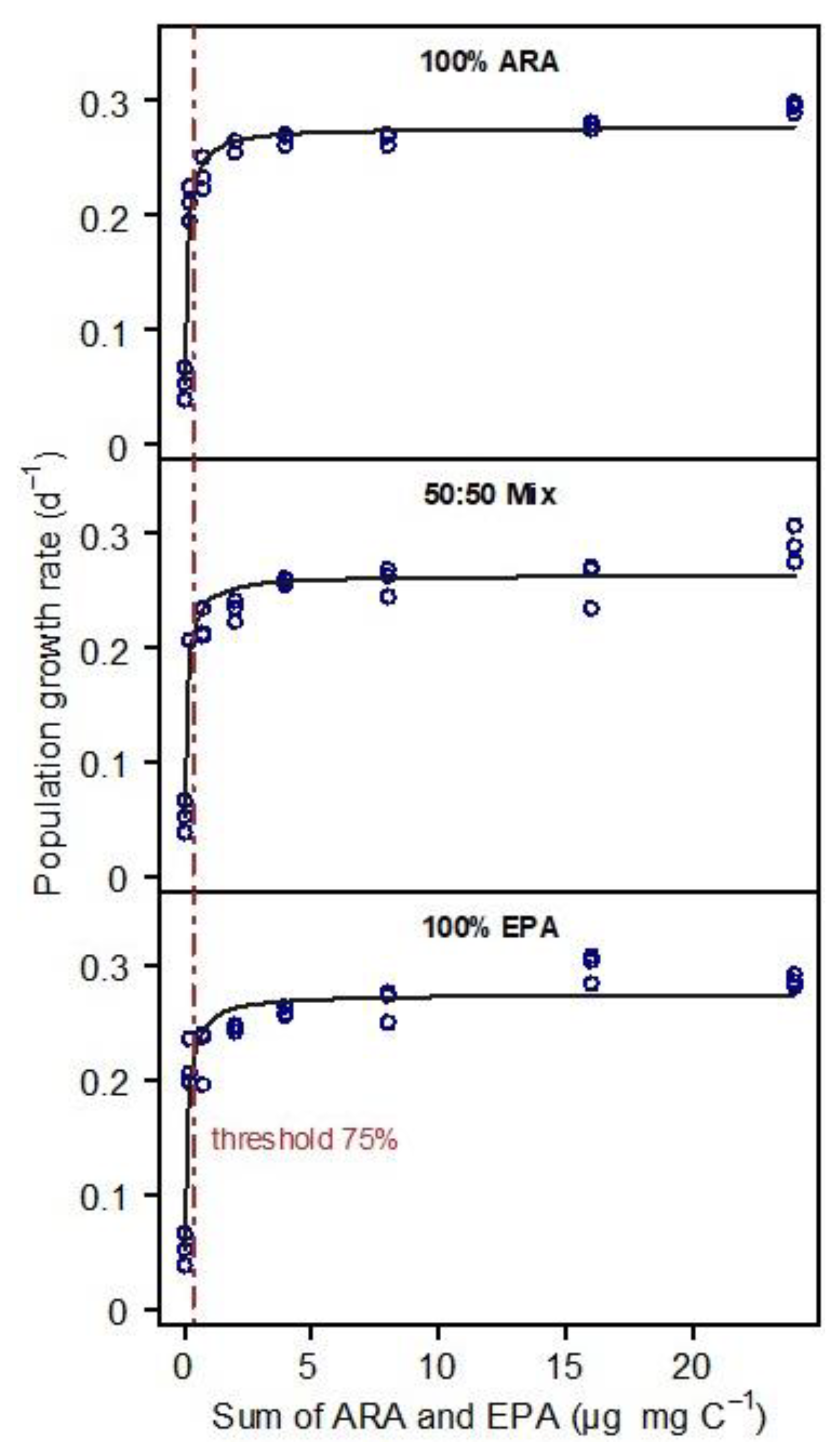
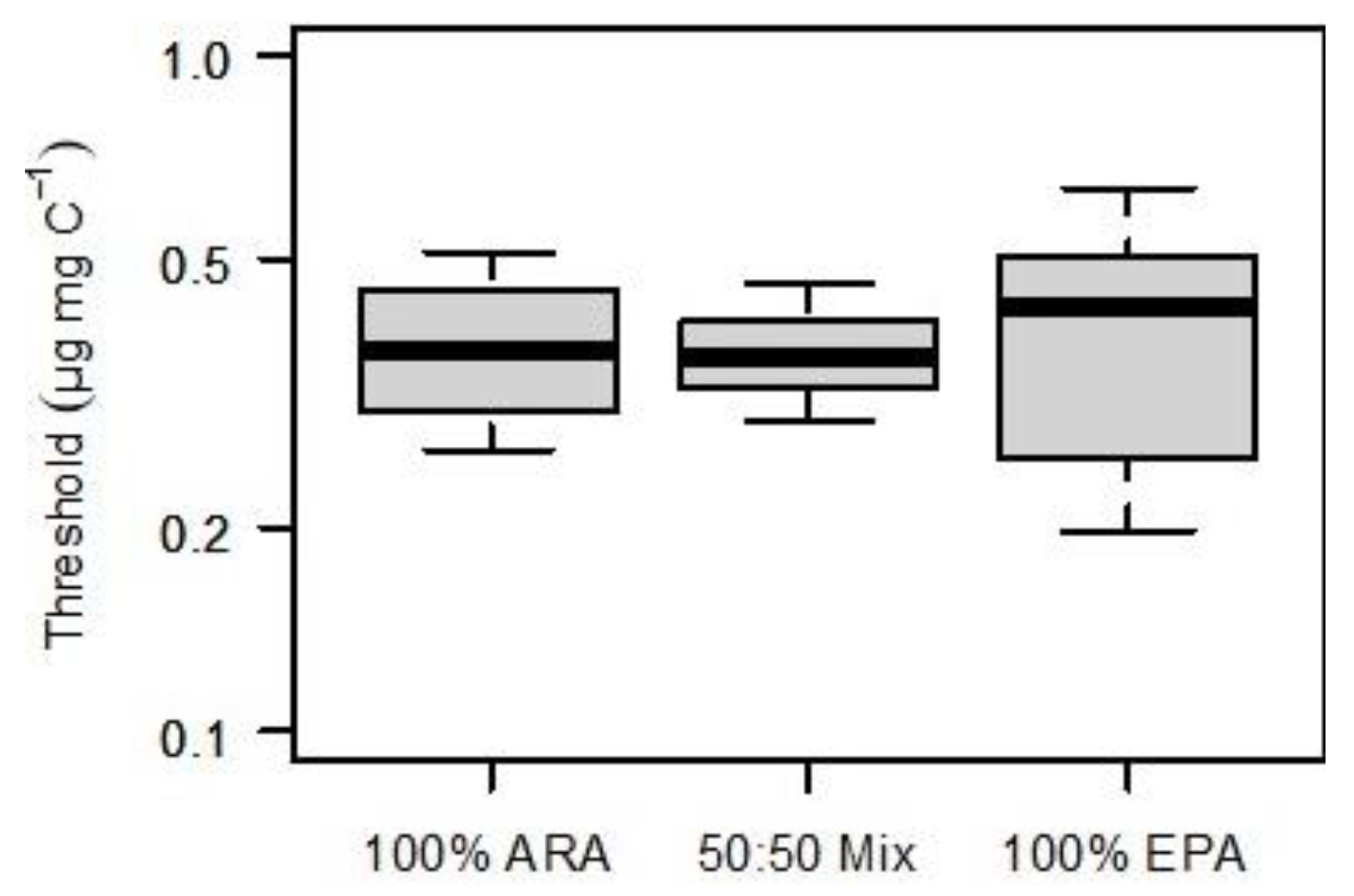
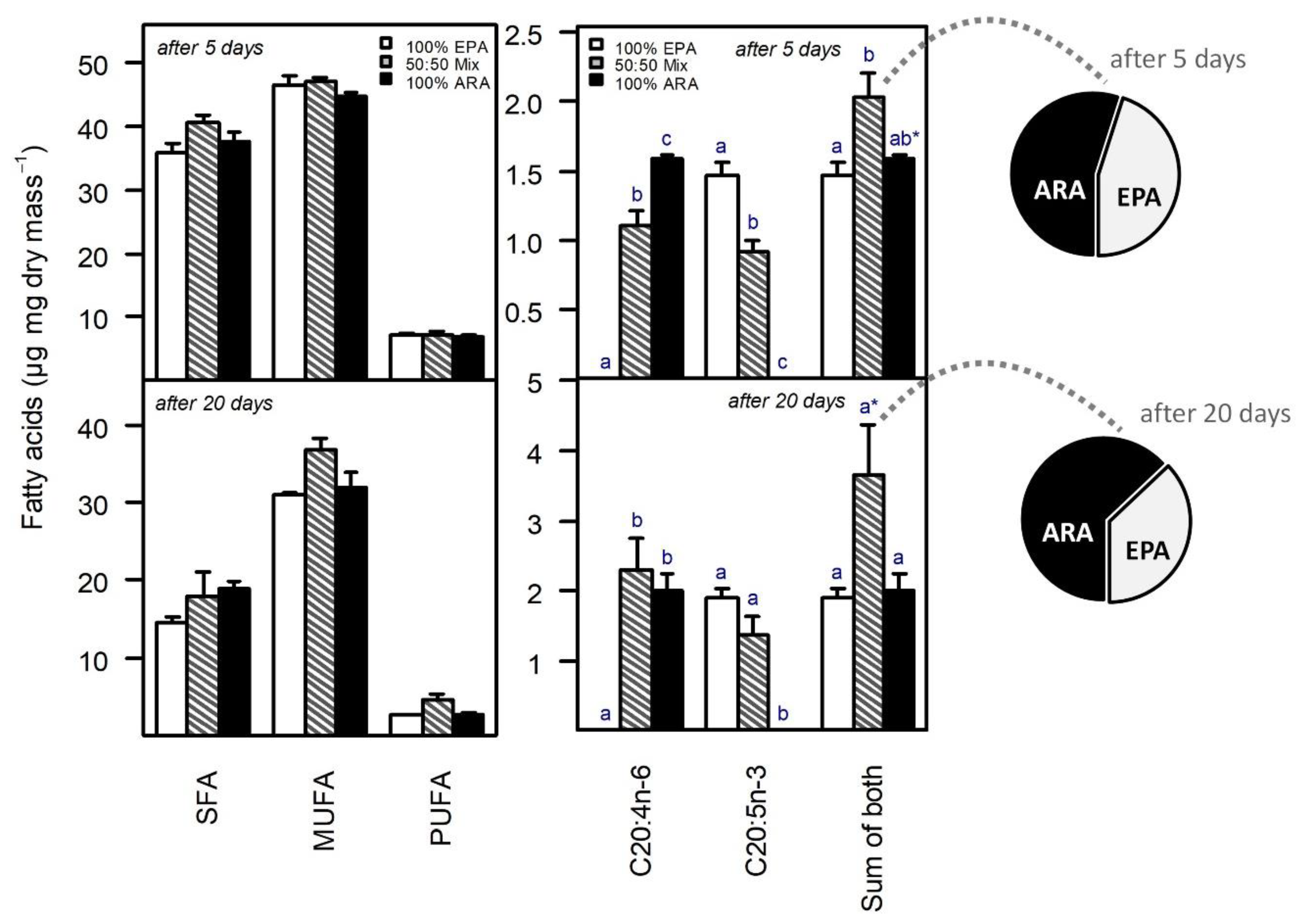
3. Results
4. Discussion
4.1. Substitutability of EPA and ARA
4.2. How Is It Possible That EPA and ARA Can Substitute for Each Other Even Though They Have Distinct Physiological Functions?
4.3. Body Fatty Acid Level Hints at Substitutability, but Also Complementarity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Carlson, S.E.; Neuringer, M. Polyunsaturated fatty acid status and neurodevelopment: A summary and critical analysis of the literature. Lipids 1999, 34, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Leonard, A.E.; Pereira, S.L.; Sprecher, H.; Huang, Y.-S. Elongation of long-chain fatty acids. Prog. Lipid Res. 2004, 43, 36–54. [Google Scholar] [CrossRef] [PubMed]
- Kabeya, N.; Fonseca, M.M.; Ferrier, D.E.K.; Navarro, J.C.; Bay, L.K.; Francis, D.S.; Tocher, D.R.; Castro, L.F.C.; Monroig, Ó. Genes for de novo biosynthesis of omega-3 polyunsaturated fatty acids are widespread in animals. Sci. Adv. 2018, 4, eaar6849. [Google Scholar] [CrossRef] [PubMed]
- Twining, C.W.; Bernhardt, J.R.; Derry, A.M.; Hudson, C.M.; Ishikawa, A.; Kabeya, N.; Kainz, M.J.; Kitano, J.; Kowarik, C.; Ladd, S.N.; et al. The evolutionary ecology of fatty-acid variation: Implications for consumer adaptation and diversification. Ecol. Lett. 2021, 24, 1709–1731. [Google Scholar] [CrossRef] [PubMed]
- Pilecky, M.; Kämmer, S.K.; Mathieu-Resuge, M.; Wassenaar, L.I.; Taipale, S.J.; Martin-Creuzburg, D.; Kainz, M.J. Hydrogen isotopes (δ2 H) of polyunsaturated fatty acids track bioconversion by zooplankton. Funct. Ecol. 2021, 36, 538–549. [Google Scholar] [CrossRef]
- Hazel, J.R. The role of alterations in membrane lipid composition in enabling physiological adaptation of organisms to their physical environment. Prog. Lipid Res. 1990, 29, 167–227. [Google Scholar] [CrossRef]
- Stillwell, W.; Wassall, S.R. Docosahexaenoic acid: Membrane properties of a unique fatty acid. Chem. Phys. Lipids 2003, 126, 1–27. [Google Scholar] [CrossRef]
- Valentine, R.C.; Valentine, D.L. Omega-3 fatty acids in cellular membranes: A unified concept. Prog. Lipid Res. 2004, 43, 383–402. [Google Scholar] [CrossRef]
- Stanley, D.W. Eicosanoids in invertebrate signal transduction systems. In Eicosanoids in Invertebrate Signal Transduction Systems; Princeton University Press: Princeton, NJ, USA, 1999. [Google Scholar] [CrossRef]
- Heckmann, L.-H.; Sibly, R.M.; Timmermans, M.J.; Callaghan, A. Outlining eicosanoid biosynthesis in the crustacean Daphnia. Front. Zool. 2008, 5, 11. [Google Scholar] [CrossRef]
- Pilecky, M.; Závorka, L.; Arts, M.T.; Kainz, M.J. Omega-3 PUFA profoundly affect neural, physiological, and behavioural competences—Implications for systemic changes in trophic interactions. Biol. Rev. 2021, 96, 2127–2145. [Google Scholar] [CrossRef]
- Stanley-Samuelson, D.W.; Jurenka, R.A.; Cripps, C.; Blomquist, G.J.; de Renobales, M. Fatty acids in insects: Composition, metabolism, and biological significance. Arch. Insect Biochem. Physiol. 1988, 9, 1–33. [Google Scholar] [CrossRef]
- Xu, H.; Meng, X.; Wei, Y.; Ma, Q.; Liang, M.; Turchini, G.M. Arachidonic acid matters. Rev. Aquac. 2022, 14, 1912–1944. [Google Scholar] [CrossRef]
- von Elert, E. Determination of limiting polyunsaturated fatty acids in Daphnia galeata using a new method to enrich food algae with single fatty acids. Limnol. Oceanogr. 2002, 47, 1764–1773. [Google Scholar] [CrossRef]
- Martin-Creuzburg, D.; Sperfeld, E.; Wacker, A. Colimitation of a freshwater herbivore by sterols and polyunsaturated fatty acids. Proc. R. Soc. B: Boil. Sci. 2009, 276, 1805–1814. [Google Scholar] [CrossRef] [PubMed]
- Ravet, J.L.; Persson, J.; Brett, M.T. Threshold dietary polyunsaturated fatty acid concentrations for Daphnia pulex growth and reproduction. Inland Waters 2012, 2, 199–209. [Google Scholar] [CrossRef]
- Twining, C.W.; Brenna, J.T.; Lawrence, P.; Shipley, J.R.; Tollefson, T.N.; Winkler, D.W. Omega-3 long-chain polyunsaturated fatty acids support aerial insectivore performance more than food quantity. Proc. Natl. Acad. Sci. USA 2016, 113, 10920–10925. [Google Scholar] [CrossRef]
- Copeman, L.; Parrish, C.; Brown, J.; Harel, M. Effects of docosahexaenoic, eicosapentaenoic, and arachidonic acids on the early growth, survival, lipid composition and pigmentation of yellowtail flounder (Limanda ferruginea): A live food enrichment experiment. Aquaculture 2002, 210, 285–304. [Google Scholar] [CrossRef]
- Gladyshev, M.I.; Sushchik, N.N.; Dubovskaya, O.P.; Makhutova, O.N.; Kalachova, G.S. Influence of sestonic elemental and essential fatty acid contents in a eutrophic reservoir in Siberia on population growth of Daphnia (longispina group). J. Plankton Res. 2006, 28, 907–917. [Google Scholar] [CrossRef]
- Wenzel, A.; Bergström, A.K.; Jansson, M.; Vrede, T. Poor Direct Exploitation of Terrestrial Particulate Organic Material from Peat Layers by Daphnia galeata. Can. J. Fish. Aquat. Sci. 2021, 69, 1870–1880. [Google Scholar] [CrossRef]
- Martin-Creuzburg, D.; von Elert, E. Good food versus bad food: The role of sterols and polyunsaturated fatty acids in determining growth and reproduction of Daphnia magna. Aquat. Ecol. 2009, 43, 943–950. [Google Scholar] [CrossRef]
- Martin-Creuzburg, D.; Wacker, A.; Basena, T. Interactions between limiting nutrients: Consequences for somatic and population growth of Daphnia magna. Limnol. Oceanogr. 2010, 55, 2597–2607. [Google Scholar] [CrossRef]
- Martin-Creuzburg, D.; Wacker, A.; Ziese, C.; Kainz, M.J. Dietary lipid quality affects temperature-mediated reaction norms of a freshwater key herbivore. Oecologia 2011, 168, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Schlotz, N.; Pester, M.; Freese, H.M.; Martin-Creuzburg, D. A dietary polyunsaturated fatty acid improves consumer performance during challenge with an opportunistic bacterial pathogen. FEMS Microbiol. Ecol. 2014, 90, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Ilić, M.; Werner, C.; Fink, P. Equal relevance of omega-3 and omega-6 polyunsaturated fatty acids for the fitness of Daphnia spp. Limnol. Oceanogr. 2019, 64, 2512–2525. [Google Scholar] [CrossRef]
- Schmitz, G.; Ecker, J. The opposing effects of n−3 and n−6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. Available online: https://www.sciencedirect.com/science/article/pii/S0024320518302303 (accessed on 25 March 2023). [CrossRef]
- Sperfeld, E.; Martin-Creuzburg, D.; Wacker, A. Multiple resource limitation theory applied to herbivorous consumers: Liebig’s minimum rule vs. interactive co-limitation. Ecol. Lett. 2011, 15, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Lampert, W. The dynamics of Daphnia magna in a shallow lake. Congr. Int. Assoc Theor. Appl. Limnol. 1991, 24, 795–798. [Google Scholar] [CrossRef]
- Juttner, F.; Leonhardt, J.; Mohren, S. Environmental Factors Affecting The Formation of Mesityloxide, Dimethylallylic Alcohol And Other Volatile Compounds Excreted By Anabaena cylindrica. Microbiology 1983, 129, 407–412. [Google Scholar] [CrossRef]
- Lukas, M.; Wacker, A. Daphnia’s dilemma of adjusting carbon budgets when facing limitations by food quantity and the essential organic compound cholesterol. J. Exp. Biol. 2013, 217, 1079–1086. [Google Scholar] [CrossRef]
- Martin-Creuzburg, D.; Oexle, S.; Wacker, A. Thresholds for Sterol-Limited Growth of Daphnia magna: A Comparative Approach Using 10 Different Sterols. J. Chem. Ecol. 2014, 40, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Martin-Creuzburg, D.; von Elert, E.; Hoffmann, K.H. Nutritional constraints at the cyanobacteria-Daphnia magna interface: The role of sterols. Limnol. Oceanogr. 2008, 53, 456–468. [Google Scholar] [CrossRef]
- Sperfeld, E.; Wacker, A. Temperature- and cholesterol-induced changes in eicosapentaenoic acid limitation of Daphnia magna determined by a promising method to estimate growth saturation thresholds. Limnol. Oceanogr. 2011, 56, 1273–1284. [Google Scholar] [CrossRef]
- R Core Development Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; p. 2020.
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Sage Publications: Thousand Oaks, CA, USA, 2018. [Google Scholar]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Emmeans: Estimated Marginal Means, Aka Least-Squares Means; The R Project for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Becker, C.; Boersma, M. Differential effects of phosphorus and fatty acids on Daphnia magna growth and reproduction. Limnol. Oceanogr. 2005, 50, 388–397. [Google Scholar] [CrossRef]
- Martin-Creuzburg, D.; Massier, T.; Wacker, A. Sex-Specific Differences in Essential Lipid Requirements of Daphnia magna. Front. Ecol. Evol. 2018, 6, 89. [Google Scholar] [CrossRef]
- Gladyshev, M.I.; Sushchik, N.N.; Dubovskaya, O.P.; Makhutova, O.N.; Kalachova, G.S. Growth Rate of Daphnia Feeding on Seston in a Siberian Reservoir: The Role of Essential Fatty Acid. Aquat. Ecol. 2008, 42, 617–627. [Google Scholar] [CrossRef]
- Sperfeld, E.; Wacker, A. Effects of temperature and dietary sterol availability on growth and cholesterol allocation of the aquatic keystone species Daphnia. J. Exp. Biol. 2009, 212, 3051–3059. [Google Scholar] [CrossRef]
- Müller-Navarra, D.C.; Brett, M.T.; Liston, A.M.; Goldman, C.R. A highly unsaturated fatty acid predicts carbon transfer between primary producers and consumers. Nature 2000, 403, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Wacker, A.; von Elert, E. Polyunsaturated fatty acids: Evidence for non-substitutable biochemical resources in Daphnia galeata. Ecology 2001, 82, 2507–2520. [Google Scholar] [CrossRef]
- Sperfeld, E.; Wacker, A. Temperature affects the limitation of Daphnia magna by eicosapentaenoic acid, and the fatty acid composition of body tissue and eggs. Freshw. Biol. 2011, 57, 497–508. [Google Scholar] [CrossRef]
- Niebylski, C.; Salem, N. A calorimetric investigation of a series of mixed-chain polyunsaturated phosphatidylcholines: Effect of sn-2 chain length and degree of unsaturation. Biophys. J. 1994, 67, 2387–2393. [Google Scholar] [CrossRef]
- Hazel, J.R.; Prosser, C.L. Molecular mechanisms of temperature compensation in poikilotherms. Physiol. Rev. 1974, 54, 620–677. [Google Scholar] [CrossRef]
- Sinensky, M. Homeoviscous Adaptation—A Homeostatic Process that Regulates the Viscosity of Membrane Lipids in Escherichia coli. Proc. Natl. Acad. Sci. USA 1974, 71, 522–525. [Google Scholar] [CrossRef] [PubMed]
- Schlotz, N.; Sørensen, J.G.; Martin-Creuzburg, D. The potential of dietary polyunsaturated fatty acids to modulate eicosanoid synthesis and reproduction in Daphnia magna: A gene expression approach. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 162, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Kim, Y. Prostaglandins and Other Eicosanoids in Insects: Biosynthesis and Biological Actions. Front. Physiol. 2018, 9, 1927. [Google Scholar] [CrossRef]
- Morgan, E.L.; Maskrey, B.H.; Rowley, A.F. At what stage in metazoan evolution did leukotriene generation first appear?—Key insights from cartilaginous fish. Dev. Comp. Immunol. 2005, 29, 53–59. Available online: https://www.sciencedirect.com/science/article/pii/S0145305X04000771 (accessed on 25 March 2023). [CrossRef] [PubMed]
- Yuan, D.; Zou, Q.; Yu, T.; Song, C.; Huang, S.; Chen, S.; Ren, Z.; Xu, A. Ancestral genetic complexity of arachidonic acid metabolism in Metazoa. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2014, 1841, 1272–1284. [Google Scholar] [CrossRef] [PubMed]
- Lands, W.E.M. Biochemistry and physiology of n-3 fatty acids. FASEB J. 1992, 6, 2530–2536. [Google Scholar] [CrossRef] [PubMed]
- Tootle, T.L.; Spradling, A.C. Drosophila Pxt: A cyclooxygenase-like facilitator of follicle maturation. Development 2008, 135, 839–847. Available online: https://journals.biologists.com/dev/article-abstract/135/5/839/65029 (accessed on 25 March 2023). [CrossRef]
- Vrablik, T.L.; Watts, J.L. Polyunsaturated fatty acid derived signaling in reproduction and development: Insights from Caenorhabditis elegans and Drosophila melanogaster. Mol. Reprod. Dev. 2013, 80, 244–259. [Google Scholar] [CrossRef]
- Schlotz, N.; Roulin, A.; Ebert, D.; Martin-Creuzburg, D. Combined effects of dietary polyunsaturated fatty acids and parasite exposure on eicosanoid-related gene expression in an invertebrate model. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 201, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Schlotz, N.; Ebert, D.; Martin-Creuzburg, D. Dietary supply with polyunsaturated fatty acids and resulting maternal effects influence host—Parasite interactions. BMC Ecol. 2013, 13, 41. [Google Scholar] [CrossRef] [PubMed]
- Bownes, M. Expression of the Genes Coding for Vitellogenin (Yolk Protein). Annu. Rev. EÈntomol. 1986, 31, 507–531. [Google Scholar] [CrossRef]
- Fink, P.; Windisch, H.S. The essential omega-3 fatty acid EPA affects expression of genes involved in the metabolism of omega-6-derived eicosanoids in Daphnia magna. Hydrobiologia 2018, 846, 5–16. [Google Scholar] [CrossRef]
- Henrotte, E.; Milla, S.; Mandiki, S.N.M.; Kestemont, P. Arachidonic Acid Induces Production of 17,20β-Dihydroxy-4-pregnen-3-one (DHP) via a Putative PGE2 Receptor in Fish Follicles from the Eurasian Perch. Lipids 2010, 46, 179–187. [Google Scholar] [CrossRef]
- Taipale, S.J.; Kainz, M.J.; Brett, M.T. Diet-switching experiments show rapid accumulation and preferential retention of highly unsaturated fatty acids in Daphnia. Oikos 2011, 120, 1674–1682. [Google Scholar] [CrossRef]
- Hartwich, M.; Martin-Creuzburg, D.; Wacker, A. Seasonal changes in the accumulation of polyunsaturated fatty acids in zooplankton. J. Plankton Res. 2013, 35, 121–134. [Google Scholar] [CrossRef]
- Wacker, A.; Martin-Creuzburg, D. Allocation of essential lipids in Daphnia magna during exposure to poor food quality. Funct. Ecol. 2007, 21, 738–747. [Google Scholar] [CrossRef]
- Putman, A.; Martin-Creuzburg, D.; Panis, B.; De Meester, L. A comparative analysis of the fatty acid composition of sexual and asexual eggs of Daphnia magna and its plasticity as a function of food quality. J. Plankton Res. 2015, 37, 752–763. [Google Scholar] [CrossRef]
- Persson, J.; Vrede, T. Polyunsaturated fatty acids in zooplankton: Variation due to taxonomy and trophic position. Freshw. Biol. 2006, 51, 887–900. [Google Scholar] [CrossRef]
- Kainz, M.J.; Perga, M.-E.; Arts, M.T.; Mazumder, A. Essential fatty acid concentrations of different seston sizes and zooplankton: A field study of monomictic coastal lakes. J. Plankton Res. 2009, 31, 635–645. Available online: https://academic.oup.com/plankt/article-abstract/31/6/635/1389888 (accessed on 25 March 2023). [CrossRef]
- Lau, D.C.P.; Jonsson, A.; Isles, P.D.F.; Creed, I.F.; Bergström, A. Lowered nutritional quality of plankton caused by global environmental changes. Glob. Chang. Biol. 2021, 27, 6294–6306. [Google Scholar] [CrossRef]
- Kainz, M.; Arts, M.T.; Mazumder, A. Essential fatty acids in the planktonic food web and their ecological role for higher trophic levels. Limnol. Oceanogr. 2004, 49, 1784–1793. [Google Scholar] [CrossRef]
- Taipale, S.J.; Kainz, M.J.; Brett, M.T. A low ω-3:ω-6 ratio in Daphnia indicates terrestrial resource utilization and poor nutritional condition. J. Plankton Res. 2015, 37, 596–610. [Google Scholar] [CrossRef]
- Twining, C.W.; Taipale, S.J.; Ruess, L.; Bec, A.; Martin-Creuzburg, D.; Kainz, M.J. Stable isotopes of fatty acids: Current and future perspectives for advancing trophic ecology. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190641. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Day 5 | |||
|---|---|---|---|
| 100% EPA | 50:50 Mix | 100% ARA | |
| C13:0 | 397.3 ± 48.5 | 528.6 ± 35.5 | 451.3 ± 50.8 |
| C14:0 | 8941.2 ± 798.3 | 12,591.5 ± 1055.6 | 10,523.5 ± 1105.2 |
| C14:1n-7 | 510.6 ± 55.3 | 629.2 ± 41.4 | 556.7 ± 52.2 |
| C15:0 | 943.6 ± 47.0 | 1187.4 ± 21.1 | 1052.6 ± 46.9 |
| C16:0 | 23,586.4 ± 1225.5 | 24,512.0 ± 458.2 | 23,704.7 ± 929.6 |
| C16:1n-7 | 20,211.5 ± 1280.7 | 22,857.5 ± 734.5 | 20,961.0 ± 294.9 |
| C17:1n-7 | 990.8 ± 84.3 | 990.8 ± 11.0 | 905.2 ± 23.8 |
| C18:0 | 1923.0 ± 181.9 | 1945.4 ± 21.1 | 1861.6 ± 64.9 |
| C18:1n-9 | 24,540.4 ± 1047.3 | 22,620.7 ± 334.2 | 22,494.2 ± 167.5 |
| C18:2n-6 (LIN) | 1104.0 ± 96.2 | 1030.5 ± 99.3 | 1012.9 ± 79.0 |
| C18:3n-6 | 88.1 ± 124.6 | 218.7 ± 155.8 | 224.7 ± 159.4 |
| C18:3n-3 (ALA) | 4079.0 ± 247.4 | 3487.2 ± 481.0 | 3632.2 ± 312.6 |
| C18:4n-3 | 457.5 ± 29.5 | 417.1 ± 33.1 | 388.3 ± 21.8 |
| C20:4n-6 (ARA) | 0.0 ± 0.0 | 1114.3 ± 137.7 | 1586.9 ± 35.7 |
| C20:5n-3 (EPA) | 1471.7 ± 125.1 | 915.9 ± 122.7 | 0.0 ± 0.0 |
| C22:6n-3 (DHA) | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Day 20 | |||
| 100% EPA | 50:50 Mix | 100% ARA | |
| C13:0 | 0.0 ± 0.0 | 214.4 ± 156.9 | 245.9 ± 26.8 |
| C14:0 | 3180.2 ± 513.9 | 5670.2 ± 2167.9 | 5939.0 ± 358.6 |
| C14:1n-7 | 165.6 ± 118.0 | 319.2 ± 236.5 | 372.0 ± 52.4 |
| C15:0 | 536.0 ± 49.1 | 682.5 ± 143.8 | 658.4 ± 55.0 |
| C16:0 | 9705.1 ± 733.0 | 10,205.9 ± 1877.7 | 10,989.7 ± 897.5 |
| C16:1n-7 | 14,138.9 ± 842.8 | 17,924.4 ± 2783.1 | 16,809.0 ± 785.8 |
| C17:1n-7 | 750.7 ± 15.0 | 310.2 ± 438.6 | 677.8 ± 91.4 |
| C18:0 | 1093.6 ± 30.6 | 1188.3 ± 134.3 | 978.4 ± 103.8 |
| C18:1n-9 | 15,736.4 ± 881.4 | 18,057.9 ± 2177.3 | 13,915.1 ± 1812.5 |
| C18:2n-6 (LIN) | 223.6 ± 14.4 | 230.6 ± 175.2 | 202.8 ± 38.7 |
| C18:3n-6 | 0.0 ± 0.0 | 0.0 ± 0.0 | 168.6 ± 27.9 |
| C18:3n-3 (ALA) | 367.2 ± 28.5 | 595.8 ± 134.4 | 329.2 ± 62.0 |
| C18:4n-3 | 60.7 ± 85.8 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| C20:4n-6 (ARA) | 0.0 ± 0.0 | 2297.6 ± 633.8 | 2003.1 ± 326.7 |
| C20:5n-3 (EPA) | 1897.0 ± 176.7 | 1357.8 ± 390.5 | 0.0 ± 0.0 |
| C22:6n-3 (DHA) | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiene, M.; Wacker, A.; Martin-Creuzburg, D. Omega-3 versus Omega-6: Are We Underestimating the Ecological Significance of Arachidonic Acid in Aquatic Systems? Biomolecules 2023, 13, 791. https://doi.org/10.3390/biom13050791
Kiene M, Wacker A, Martin-Creuzburg D. Omega-3 versus Omega-6: Are We Underestimating the Ecological Significance of Arachidonic Acid in Aquatic Systems? Biomolecules. 2023; 13(5):791. https://doi.org/10.3390/biom13050791
Chicago/Turabian StyleKiene, Marvin, Alexander Wacker, and Dominik Martin-Creuzburg. 2023. "Omega-3 versus Omega-6: Are We Underestimating the Ecological Significance of Arachidonic Acid in Aquatic Systems?" Biomolecules 13, no. 5: 791. https://doi.org/10.3390/biom13050791
APA StyleKiene, M., Wacker, A., & Martin-Creuzburg, D. (2023). Omega-3 versus Omega-6: Are We Underestimating the Ecological Significance of Arachidonic Acid in Aquatic Systems? Biomolecules, 13(5), 791. https://doi.org/10.3390/biom13050791