

A Machine Learning Approach for Highlighting microRNAs as Biomarkers Linked to Amyotrophic Lateral Sclerosis Diagnosis and Progression



Abstract
:1. Introduction
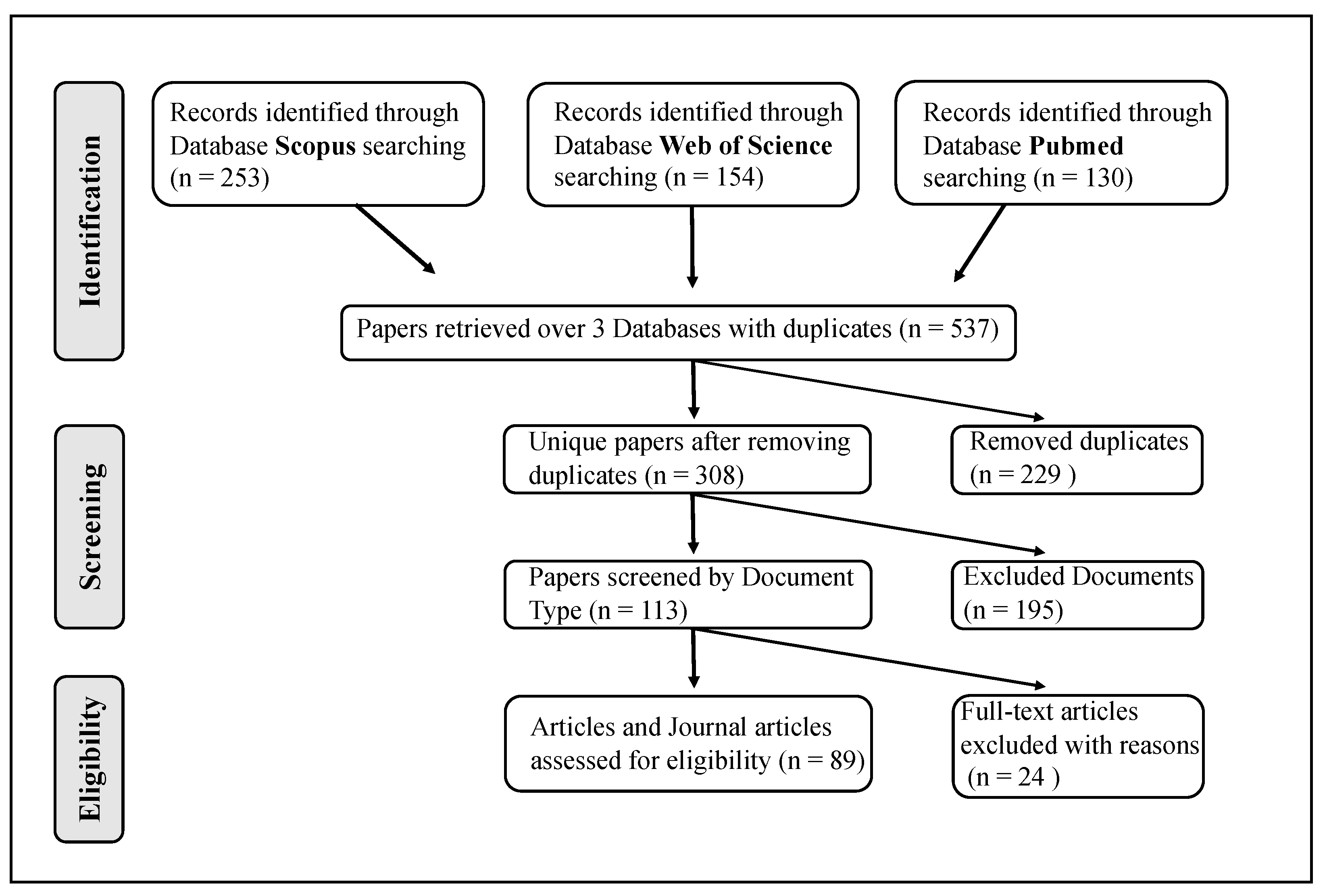
2. Methods
2.1. Paper Location and Selection
2.2. Paper Analysis
- The study was performed on human cells and tissues, in animal models, or in patients with ALS.
- The study measured miRNA expression in blood, serum, plasma, urines, CSF, tissues, or cells.
- The study assessed the association between diagnosis and/or prognosis and miRNA expression.
- The study was a letter to editors, an editorial article, a personal opinion and commentary, a short communication, a meta-analysis, a review, a note, a book chapter, an erratum, or a retracted article.
- A study investigated other types of ALS biomarkers such as proteins, genes, or other types of non-coding RNAs with unknown functions, including long non-coding RNAs, circular RNAs, and small nucleolar RNAs.
- The study tested the prognostic role of target genes or other molecules instead of that of miRNAs.
- k sets of pertinent keywords (each set denotes a topic).
- The document-term matrix, a matrix reporting how much each article is statistically related to a specific topic (known as topic proportion).
2.3. Results Presentation
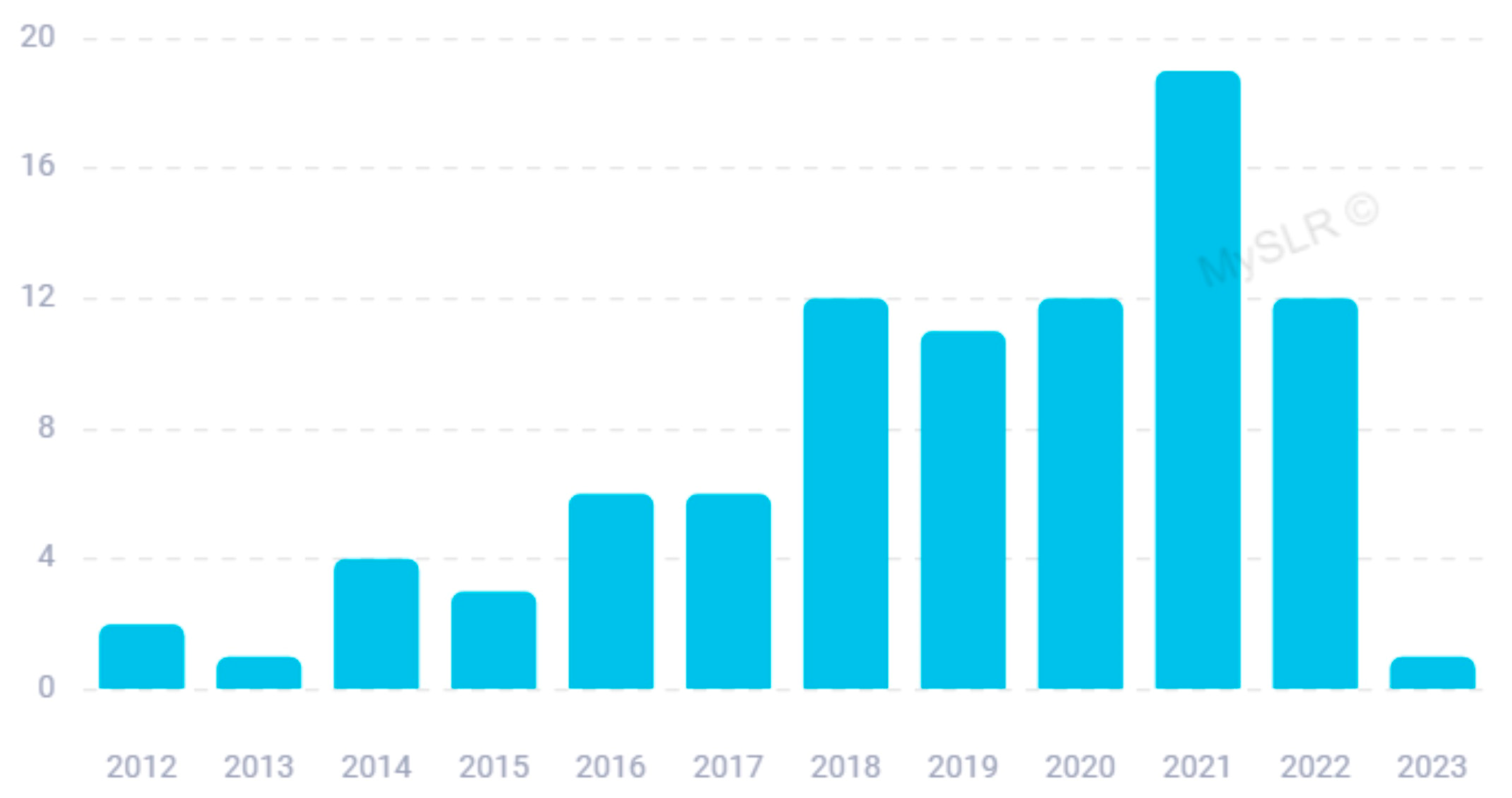
3. Results
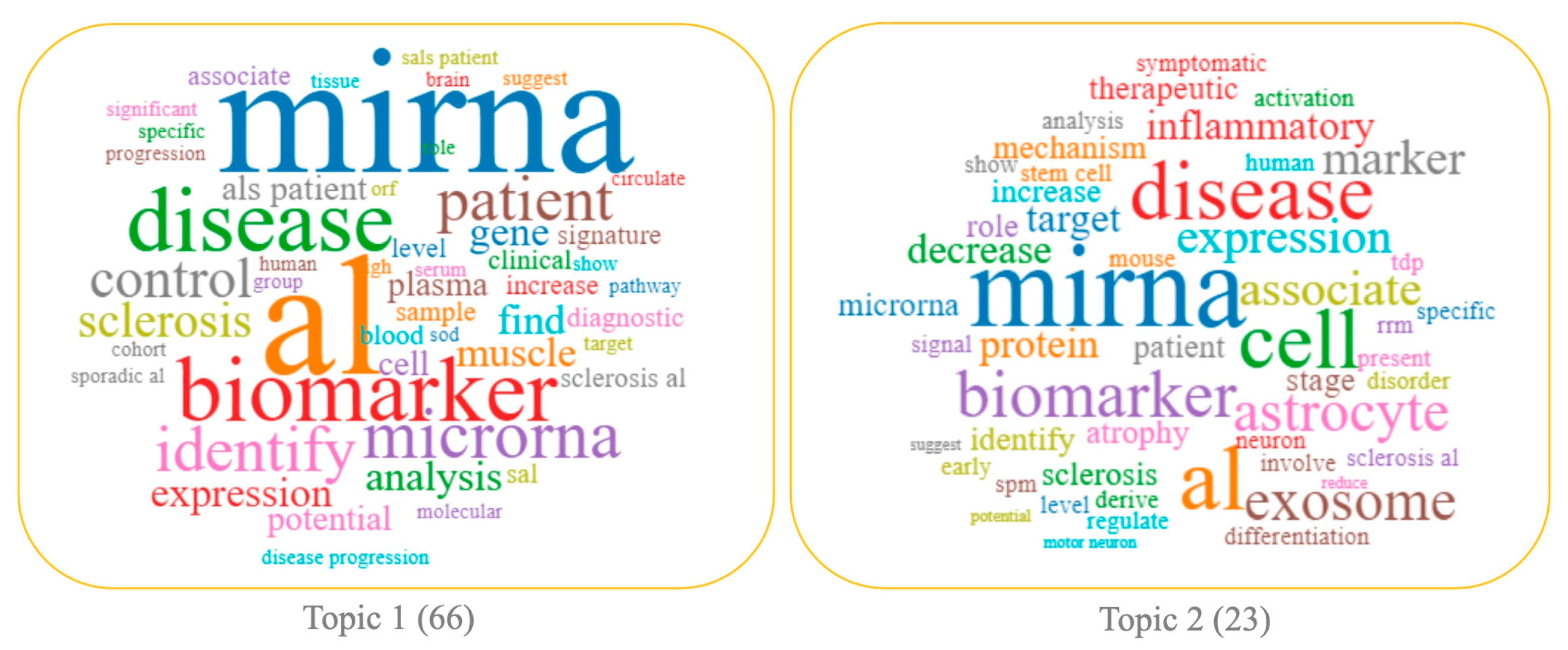
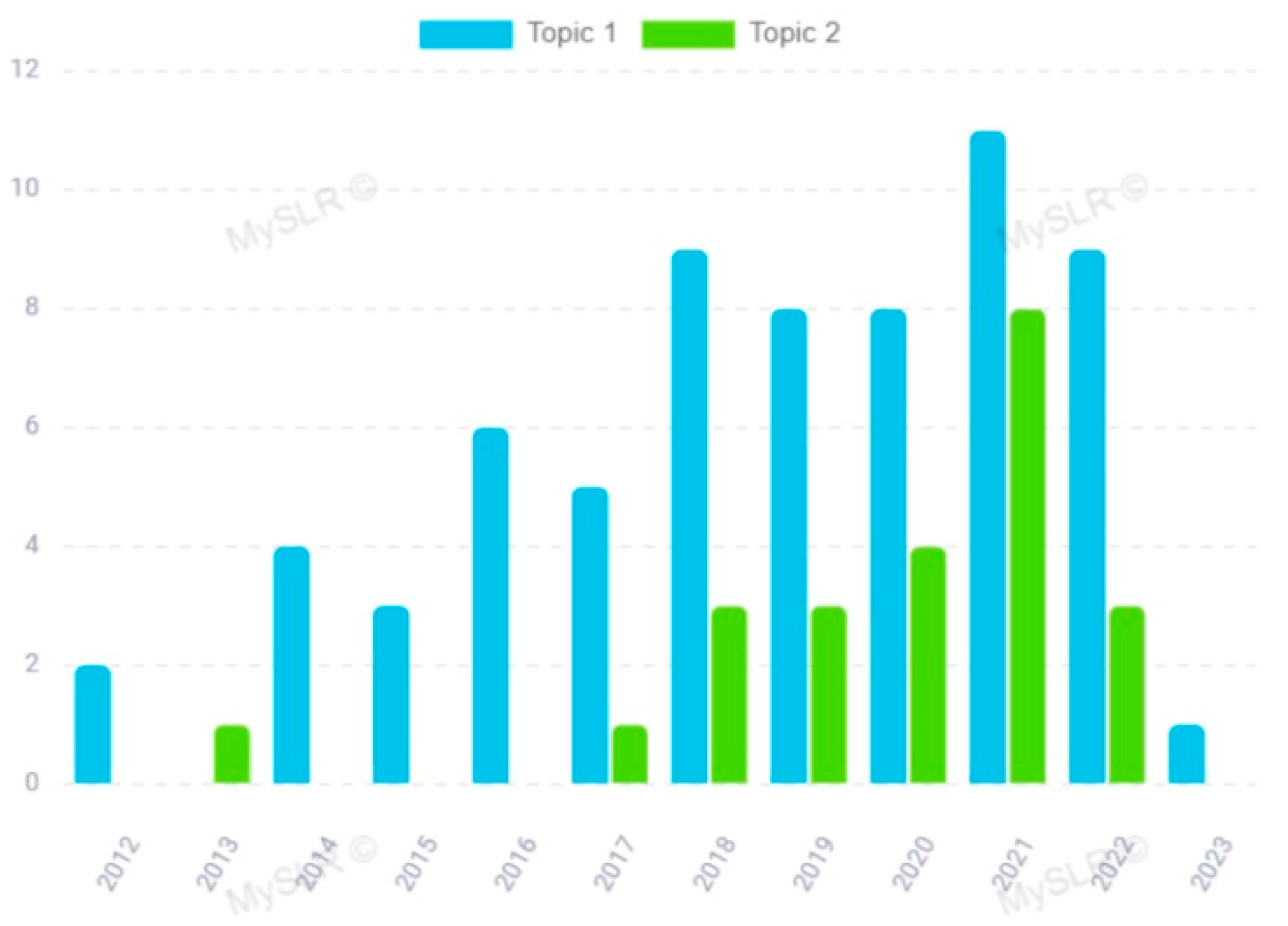
3.1. Topic 2—miRNA in ALS Pathogenesis
3.2. Topic 1—miRNA in the Clinical and Translational Research of ALS

{kind=link}
{kind=link}
{kind=link}
{kind=link}
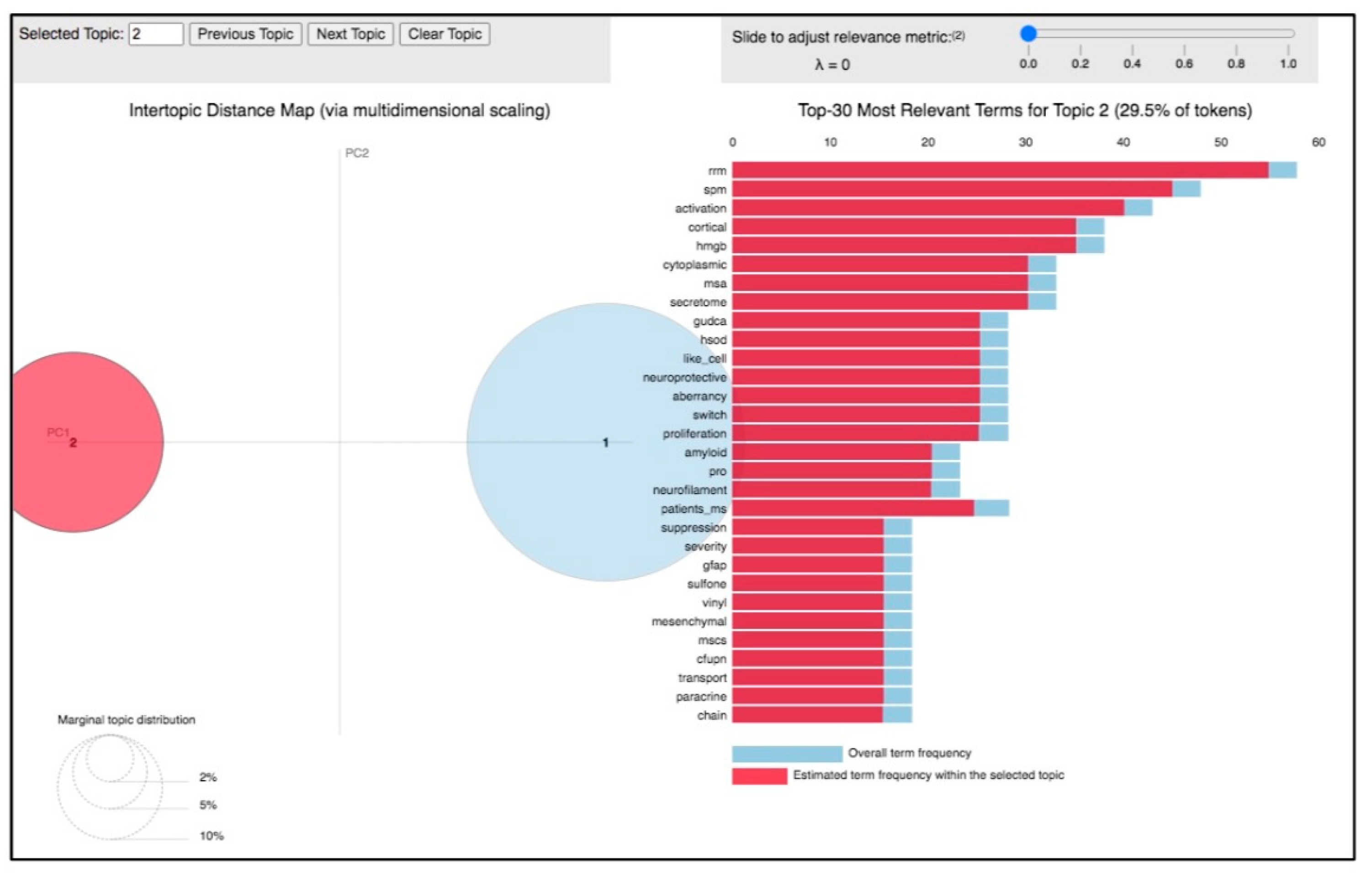{kind=link}
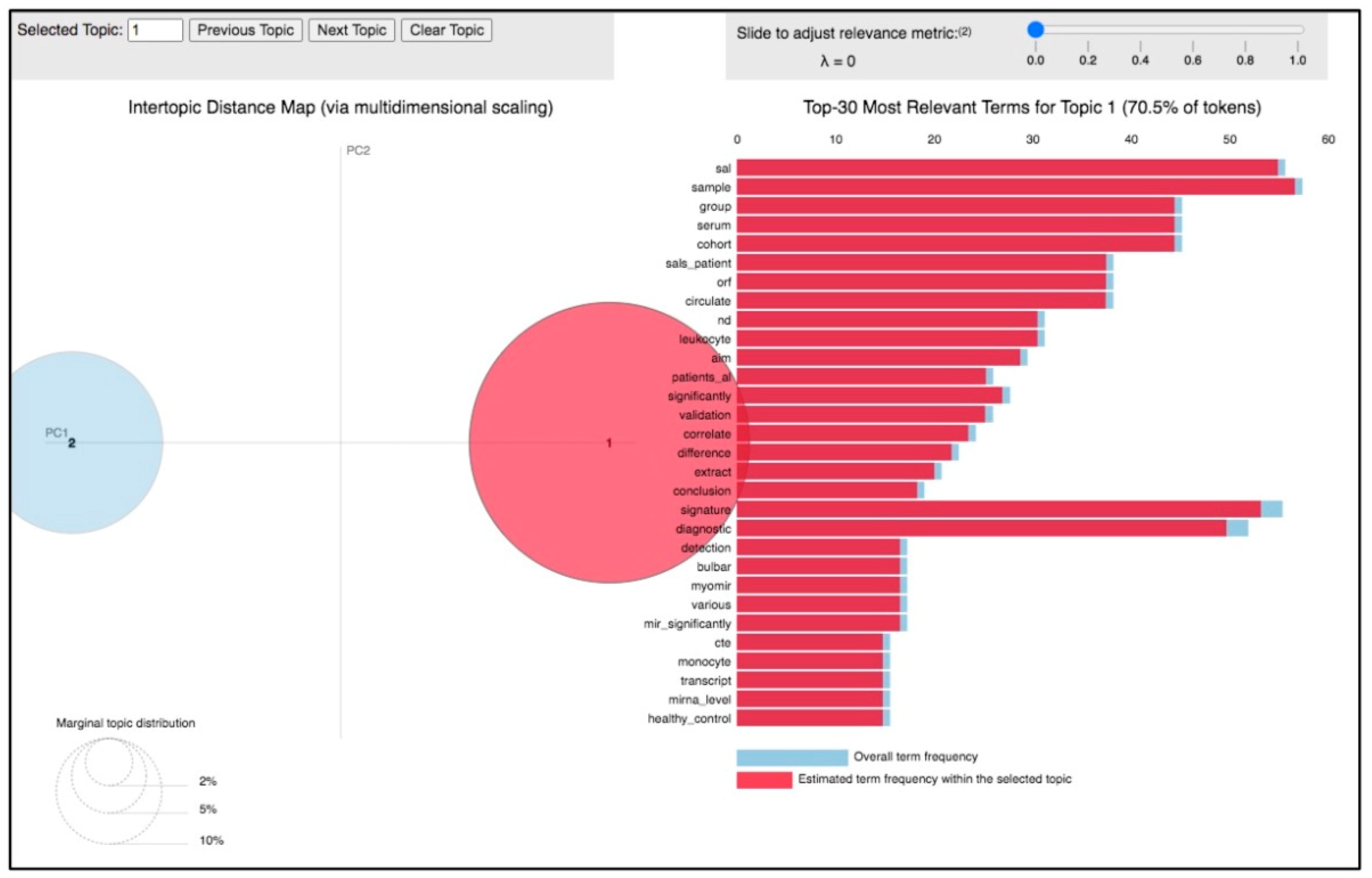{kind=link}
| Dysregulated miRNAs | Biological Sample | Methods for miRNA Profiling | Clinical Value | References |
|---|---|---|---|---|
| ↑ miR-124 | Exosomes of CSF; Blood leukocytes | RT-qPCR | Increased expression has been associated with disease severity and shorter survival | [72,73] |
| ↑ miR-155 | Peripheral Blood; Mononuclear Cells (PBMCs); Spinal cord | Microarray technology; RT-qPCR | Elevated levels have been associated with disease progression and survival | [74,75] |
| ↓ miR-9 | Blood leukocytes; EVs | RT-qPCR; Next-Generation Sequencing | Reduced expression has been associated with disease severity and shorter survival | [68,73] |
| ↑ miR-206 | Serum | RT-qPCR | Higher levels associated with disease progression and muscle atrophy | [58,76,77,78,79,80,81] |
| ↑ miR-181 | Plasma; CSF | Next-Generation Sequencing | High levels predict a greater than two-fold risk of death in independent discovery and replication cohorts | [60,82] |
| ↑ miR-338-3p | Blood leukocytes; CSF; serum and spinal cord | Next-Generation Sequencing; Meta-analysis using rank aggregation (RRA) method, followed by RT-qPCR validation. | Elevated levels have been associated with disease progression and survival | [71,84,85] |
| ↓ miR-338-3p | Blood leukocytes | Microarray technology | Decreased levels correlated with shorter survival and a more aggressive disease course | [73,86] |
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Marra, F.; Lunetti, P.; Curcio, R.; Lasorsa, F.M.; Capobianco, L.; Porcelli, V.; Dolce, V.; Fiermonte, G.; Scarcia, P. An Overview of Mitochondrial Protein Defects in Neuromuscular Diseases. Biomolecules 2021, 11, 1633. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.A.; Lally, C.; Kupelian, V.; Flanders, W.D. Estimated Prevalence and Incidence of Amyotrophic Lateral Sclerosis and Sod1 and C9orf72 Genetic Variants. Neuroepidemiology 2021, 55, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Longinetti, E.; Fang, F. Epidemiology of Amyotrophic Lateral Sclerosis: An Update of Recent Literature. Curr. Opin. Neurol. 2019, 32, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Mead, R.J.; Shan, N.; Reiser, H.J.; Marshall, F.; Shaw, P.J. Amyotrophic Lateral Sclerosis: A Neurodegenerative Disorder Poised for Successful Therapeutic Translation. Nat. Rev. Drug Discov. 2023, 22, 185–212. [Google Scholar] [CrossRef] [PubMed]
- Rusina, R.; Vandenberghe, R.; Bruffaerts, R. Cognitive and Behavioral Manifestations in Als: Beyond Motor System Involvement. Diagnostics 2021, 11, 624. [Google Scholar] [CrossRef]
- Dal Bello-Haas, V. Physical Therapy for Individuals with Amyotrophic Lateral Sclerosis: Current Insights. Degener. Neurol. Neuromuscul. Dis. 2018, 8, 45–54. [Google Scholar] [CrossRef]
- Rokade, A.V.; Yelne, P.; Giri, A. Riluzole and Edavarone: The Hope against Amyotrophic Lateral Sclerosis. Cureus J. Med. Sci. 2022, 14, e30035. [Google Scholar] [CrossRef]
- Chiò, A.; Mazzini, L.; D’Alfonso, S.; Corrado, L.; Canosa, A.; Moglia, C.; Manera, U.; Bersano, E.; Brunetti, M.; Barberis, M.; et al. The Multistep Hypothesis of Als Revisited the Role of Genetic Mutations. Neurology 2018, 91, E635–E642. [Google Scholar] [CrossRef]
- Renton, A.E.; Chiò, A.; Traynor, B.J. State of Play in Amyotrophic Lateral Sclerosis Genetics. Nat. Neurosci. 2014, 17, 17–23. [Google Scholar] [CrossRef]
- Ricci, C.; Giannini, F.; Riolo, G.; Bocci, S.; Casali, S.; Battistini, S. A Novel Variant in Superoxide Dismutase 1 Gene (p.V119M) in Als Patients with Pure Lower Motor Neuron Presentation. Genes 2021, 12, 1544. [Google Scholar] [CrossRef]
- Beers, D.R.; Appel, S.H. Immune Dysregulation in Amyotrophic Lateral Sclerosis: Mechanisms and Emerging Therapies. Lancet Neurol. 2019, 18, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Bose, P.; Armstrong, G.A.B.; Drapeau, P. Neuromuscular Junction Abnormalities in a Zebrafish Loss-of-Function Model of Tdp-43. J. Neurophysiol. 2019, 121, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Mitra, J.; Hegde, M.L. A Commentary on Tdp-43 and DNA Damage Response in Amyotrophic Lateral Sclerosis. J. Exp. Neurosci. 2019, 13, 1179069519880166. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.M.; Ji, Y.N.; Wang, W.; Zhang, L.L.; Chen, Z.H.; Yu, M.M.; Shen, Y.T.; Ding, F.; Gu, X.S.; Sun, H.L. Amyotrophic Lateral Sclerosis: Molecular Mechanisms, Biomarkers, Therapeutic Strategies. Antioxidants 2021, 10, 1012. [Google Scholar] [CrossRef]
- Sever, B.; Ciftci, H.; DeMirci, H.; Sever, H.; Ocak, F.; Yulug, B.; Tateishi, H.; Tateishi, T.; Otsuka, M.; Fujita, M.; et al. Comprehensive Research on Past and Future Therapeutic Strategies Devoted to Treatment of Amyotrophic Lateral Sclerosis. Int. J. Mol. Sci. 2022, 23, 2400. [Google Scholar] [CrossRef] [PubMed]
- Tefera, T.W.; Steyn, F.J.; Ngo, S.T.; Borges, K. Cns Glucose Metabolism in Amyotrophic Lateral Sclerosis: A Therapeutic Target? Cell Biosci. 2021, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- DeJesus-Hernandez, M.; Mackenzie, I.R.; Boeve, B.F.; Boxer, A.L.; Baker, M.; Rutherford, N.J.; Nicholson, A.M.; Finch, N.A.; Flynn, H.; Adamson, J.; et al. Expanded Ggggcc Hexanucleotide Repeat in Noncoding Region of Causes Chromosome 9p-Linked Ftd and Als. Neuron 2011, 72, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Renton, A.E.; Majounie, E.; Waite, A.; Simón-Sánchez, J.; Rollinson, S.; Gibbs, J.R.; Schymick, J.C.; Laaksovirta, H.; van Swieten, J.C.; Myllykangas, L.; et al. A hexanucleotide repeat expansion in C9ORF72 is the cause of chromosome 9p21-linked ALS-FTD. Neuron 2011, 72, 257–268. [Google Scholar] [CrossRef]
- Rosen, D.R.; Siddique, T.; Patterson, D.; Figlewicz, D.A.; Sapp, P.; Hentati, A.; Donaldson, D.; Goto, J.; Oregan, J.P.; Deng, H.X.; et al. Mutations in Cu/Zn Superoxide-Dismutase Gene Are Associated with Familial Amyotrophic-Lateral-Sclerosis. Nature 1993, 362, 59–62. [Google Scholar] [CrossRef]
- Aksoy, Y.A.; Deng, W.; Stoddart, J.; Chung, R.; Guillemin, G.; Cole, N.J.; Neely, G.G.; Hesselson, D. “Stressed Out”: The Role of Fus and Tdp-43 in Amyotrophic Lateral Sclerosis. Int. J. Biochem. Cell Biol. 2020, 126, 105821. [Google Scholar] [CrossRef]
- Prasad, A.; Bharathi, V.; Sivalingam, V.; Girdhar, A.; Patel, B.K. Molecular Mechanisms of Tdp-43 Misfolding and Pathology in Amyotrophic Lateral Sclerosis. Front. Mol. Neurosci. 2019, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Miller, R.G.; Swash, M.; Munsat, T.L.; World Federation of Neurology Research Group on Motor Neuron Diseases. El Escorial Revisited: Revised Criteria for the Diagnosis of Amyotrophic Lateral Sclerosis. Amyotroph. Lateral Scler. Other Mot. Neuron Disord. 2000, 1, 293–299. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, M.; Dengler, R.; Eisen, A.; England, J.D.; Kaji, R.; Kimura, J.; Mills, K.; Mitsumoto, H.; Nodera, H.; Shefner, J.; et al. Electrodiagnostic Criteria for Diagnosis of Als. Clin. Neurophysiol. 2008, 119, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Vucic, S.; Ferguson, T.A.; Cummings, C.; Hotchkin, M.T.; Genge, A.; Glanzman, R.; Roet, K.C.D.; Cudkowicz, M.; Kiernan, M.C. Gold Coast Diagnostic Criteria: Implications for Als Diagnosis and Clinical Trial Enrollment. Muscle Nerve 2021, 64, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Goutman, S.A.; Hardiman, O.; Al-Chalabi, A.; Chió, A.; Savelieff, M.G.; Kiernan, M.C.; Feldman, E.L. Recent Advances in the Diagnosis and Prognosis of Amyotrophic Lateral Sclerosis. Lancet Neurol. 2022, 21, 480–493. [Google Scholar] [CrossRef]
- Richards, D.; Morren, J.A.; Pioro, E.P. Time to Diagnosis and Factors Affecting Diagnostic Delay in Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2020, 417, 117054. [Google Scholar] [CrossRef]
- Fournier, C.N. Considerations for Amyotrophic Lateral Sclerosis (Als) Clinical Trial Design. Neurotherapeutics 2022, 19, 1180–1192. [Google Scholar] [CrossRef]
- Pritchard, C.C.; Cheng, H.H.; Tewari, M. Microrna Profiling: Approaches and Considerations. Nat. Rev. Genet. 2012, 13, 358–369. [Google Scholar] [CrossRef]
- Wu, D.; Raafat, A.; Pak, E.; Clemens, S.; Murashov, A.K. Dicer-microRNA Pathway Is Critical for Peripheral Nerve Regeneration and Functional Recovery in Vivo and Regenerative Axonogenesis in Vitro. Exp. Neurol. 2012, 233, 555–565. [Google Scholar] [CrossRef]
- Gandhi, G.; Abdullah, S.; Foead, A.I.; Yeo, W.W.Y. The Potential Role of Mirna Therapies in Spinal Muscle Atrophy. J. Neurol. Sci. 2021, 427, 117485. [Google Scholar] [CrossRef]
- Godlewski, J.; Lenart, J.; Salinska, E. Microrna in Brain Pathology: Neurodegeneration the Other Side of the Brain Cancer. Non-Coding RNA 2019, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Otaegi, G.; Pollock, A.; Sun, T. An Optimized Sponge for Microrna Mir-9 Affects Spinal Motor Neuron Development. Front. Neurosci. 2012, 5, 146. [Google Scholar] [CrossRef]
- Agostini, M.; Tucci, P.; Steinert, J.R.; Shalom-Feuerstein, R.; Rouleau, M.; Aberdam, D.; Forsythe, I.D.; Young, K.W.; Ventura, A.; Concepcion, C.P.; et al. Microrna-34a Regulates Neurite Outgrowth, Spinal Morphology, Function. Proc. Natl. Acad. Sci. USA 2011, 108, 21099–21104. [Google Scholar] [CrossRef] [PubMed]
- Agostini, M.; Tucci, P.; Killick, R.; Candi, E.; Sayan, B.S.; Cervo, P.R.D.; Nicotera, P.; McKeon, F.; Knight, R.A.; Mak, T.W.; et al. Neuronal Differentiation by Tap73 Is Mediated by Microrna-34a Regulation of Synaptic Protein Targets. Proc. Natl. Acad. Sci. USA 2011, 108, 21093–21098. [Google Scholar] [CrossRef] [PubMed]
- Tucci, P.; Agostini, M.; Grespi, F.; Markert, E.K.; Terrinoni, A.; Vousden, K.H.; Muller, P.A.J.; Dötsch, V.; Kehrloesser, S.; Sayan, B.S.; et al. Loss of P63 and Its Microrna-205 Target Results in Enhanced Cell Migration and Metastasis in Prostate Cancer. Proc. Natl. Acad. Sci. USA 2012, 109, 15312–15317. [Google Scholar]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. Mirnas as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [PubMed]
- Dilmaghani, N.A.; Hussen, B.M.; Nateghinia, S.; Taheri, M.; Ghafouri-Fard, S. Emerging Role of Micrornas in the Pathogenesis of Amyotrophic Lateral Sclerosis. Metab. Brain Dis. 2021, 36, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Rinchetti, P.; Rizzuti, M.; Faravelli, I.; Corti, S. Microrna Metabolism and Dysregulation in Amyotrophic Lateral Sclerosis. Mol. Neurobiol. 2018, 55, 2617–2630. [Google Scholar] [CrossRef]
- Nowak, J.S.; Michlewski, G. Mirnas in Development and Pathogenesis of the Nervous System. Biochem. Soc. Trans. 2013, 41, 815–820. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, L.J. Micrornas in Amyotrophic Lateral Sclerosis: From Pathogenetic Involvement to Diagnostic Biomarker and Therapeutic Agent Development. Neurol. Sci. 2020, 41, 3569–3577. [Google Scholar] [CrossRef]
- Panio, A.; Cava, C.; D’Antona, S.; Bertoli, G.; Porro, D. Diagnostic Circulating Mirnas in Sporadic Amyotrophic Lateral Sclerosis. Front. Med. 2022, 9, 861960. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lan, S.; Shi, X.-J.; Fan, F.-C.; Liu, Q.-S.; Cong, L.; Cheng, Y. Systematic Review and Meta-Analysis on Micrornas in Amyotrophic Lateral Sclerosis. Brain Res. Bull. 2023, 194, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Ammirato, S.; Felicetti, A.M.; Rogano, D.; Linzalone, R.; Corvello, V. Digitalising the Systematic Literature Review Process: The Myslr Platform. Knowl. Manag. Res. Pract. 2023, 21, 777–794. [Google Scholar] [CrossRef]
- Bevacqua, E.; Ammirato, S.; Cione, E.; Curcio, R.; Dolce, V.; Tucci, P. The Potential of Micrornas as Non-Invasive Prostate Cancer Biomarkers: A Systematic Literature Review Based on a Machine Learning Approach. Cancers 2022, 14, 5418. [Google Scholar] [CrossRef]
- Blei, D.M.; Ng, A.Y.; Jordan, M.I. Latent Dirichlet Allocation. J. Mach. Learn. Res. 2003, 3, 993–1022. [Google Scholar]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; PRISMA Group. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The Prisma Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Denyer, D.; Tranfield, D. Producing a Systematic Review. In The Sage Handbook of Organizational Research Methods; Sage Publications Ltd.: New York, NY, USA, 2009; pp. 671–689. [Google Scholar]
- Chen, Z.; Liu, B. Topic Modeling Using Topics from Many Domains, Lifelong Learning and Big Data. In Proceedings of the 31st International Conference on International Conference on Machine Learning, Beijing, China, 21–26 June 2014; Volume 32. [Google Scholar]
- Vaz, A.R.; Vizinha, D.; Morais, H.; Colaço, A.R.; Loch-Neckel, G.; Barbosa, M.; Brites, D. Overexpression of Mir-124 in Motor Neurons Plays a Key Role in Als Pathological Processes. Int. J. Mol. Sci. 2021, 22, 6128. [Google Scholar] [CrossRef]
- Li, C.; Wei, Q.; Gu, X.; Chen, Y.; Chen, X.; Cao, B.; Ou, R.; Shang, H. Decreased Glycogenolysis by Mir-338-3p Promotes Regional Glycogen Accumulation within the Spinal Cord of Amyotrophic Lateral Sclerosis Mice. Front. Mol. Neurosci. 2019, 12, 114. [Google Scholar] [CrossRef]
- Barbosa, M.; Gomes, C.; Sequeira, C.; Gonçalves-Ribeiro, J.; Pina, C.C.; Carvalho, L.A.; Moreira, R.; Vaz, S.H.; Vaz, A.R.; Brites, D. Recovery of Depleted Mir-146a in Als Cortical Astrocytes Reverts Cell Aberrancies and Prevents Paracrine Pathogenicity on Microglia and Motor Neurons. Front. Cell Dev. Biol. 2021, 9, 634355. [Google Scholar] [CrossRef]
- Gomes, C.; Cunha, C.; Nascimento, F.; Ribeiro, J.A.; Vaz, A.R.; Brites, D. Cortical Neurotoxic Astrocytes with Early Als Pathology and Mir-146a Deficit Replicate Gliosis Markers of Symptomatic Sod1g93a Mouse Model. Mol. Neurobiol. 2019, 56, 2137–2158. [Google Scholar] [CrossRef]
- Lai, X.; Wang, Y.; Wang, X.; Liu, B.; Rong, L. Mir-146a-5p-Modified Hucmsc-Derived Exosomes Facilitate Spinal Cord Function Recovery by Targeting Neurotoxic Astrocytes. Stem Cell Res. Ther. 2022, 13, 487. [Google Scholar] [CrossRef] [PubMed]
- Vaz, A.R.; Pinto, S.; Ezequiel, C.; Cunha, C.; Carvalho, L.A.; Moreira, R.; Brites, D. Phenotypic Effects of Wild-Type and Mutant Sod1 Expression in N9 Murine Microglia at Steady State, Inflammatory and Immunomodulatory Conditions. Front. Cell. Neurosci. 2019, 13, 109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Almeida, S.; Lu, Y.; Nishimura, A.L.; Peng, L.; Sun, D.; Wu, B.; Karydas, A.M.; Tartaglia, M.C.; Fong, J.C.; et al. Downregulation of Microrna-9 in Ipsc-Derived Neurons of Ftd/Als Patients with Tdp-43 Mutations. PLoS ONE 2013, 8, e76055. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.P.; Wada, S.; Vergani, L.; Hock, M.B.; Lamon, S.; Léger, B.; Ushida, T.; Cartoni, R.; Wadley, G.D.; Hespel, P.; et al. Disruption of Skeletal Muscle Mitochondrial Network Genes and Mirnas in Amyotrophic Lateral Sclerosis. Neurobiol. Dis. 2013, 49, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Pegoraro, V.; Marozzo, R.; Angelini, C. Micrornas and Hdac4 Protein Expression in the Skeletal Muscle of Als Patients. Clin. Neuropathol. 2020, 39, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Waller, R.; Goodall, E.F.; Milo, M.; Cooper-Knock, J.; Da Costa, M.; Hobson, E.; Kazoka, M.; Wollff, H.; Heath, P.R.; Shaw, P.J.; et al. Serum Mirnas Mir-206, 143-3p and 374b-5p as Potential Biomarkers for Amyotrophic Lateral Sclerosis (Als). Neurobiol. Aging 2017, 55, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Potenza, N.; Mosca, N.; Mondola, P.; Damiano, S.; Russo, A.; De Felice, B. Human Mir-26a-5p Regulates the Glutamate Transporter Slc1a1 (Eaat3) Expression. Relevance in Multiple Sclerosis. Biochim. Et Biophys. Acta-Mol. Basis Dis. 2018, 1864, 317–323. [Google Scholar] [CrossRef]
- Benigni, M.; Ricci, C.; Jones, A.R.; Giannini, F.; Al-Chalabi, A.; Battistini, S. Identification of Mirnas as Potential Biomarkers in Cerebrospinal Fluid from Amyotrophic Lateral Sclerosis Patients. Neuromolecular Med. 2016, 18, 551–560. [Google Scholar] [CrossRef]
- Joilin, G.; Gray, E.; Thompson, A.G.; Talbot, K.; Leigh, P.N.; Newbury, S.F.; Turner, M.R.; Hafezparast, M. Profiling Non-Coding Rna Expression in Cerebrospinal Fluid of Amyotrophic Lateral Sclerosis Patients. Ann. Med. 2022, 54, 3069–3078. [Google Scholar] [CrossRef]
- Kmetzsch, V.; Latouche, M.; Saracino, D.; Rinaldi, D.; Camuzat, A.; Gareau, T.; French Research Network on FTD/ALS; Le Ber, I.; Colliot, O.; Becker, E. Microrna Signatures in Genetic Frontotemporal Dementia and Amyotrophic Lateral Sclerosis. Ann. Clin. Transl. Neurol. 2022, 9, 1778–1791. [Google Scholar] [CrossRef]
- Raheja, R.; Regev, K.; Healy, B.C.; Mazzola, M.A.; Beynon, V.; Von Glehn, F.; Paul, A.; Diaz-Cruz, C.; Gholipour, T.; Glanz, B.I.; et al. Correlating Serum Micrornas and Clinical Parameters in Amyotrophic Lateral Sclerosis. Muscle Nerve 2018, 58, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Matamala, J.M.; Arias-Carrasco, R.; Sanchez, C.; Uhrig, M.; Bargsted, L.; Matus, S.; Maracaja-Coutinho, V.; Abarzua, S.; van Zundert, B.; Verdugo, R.; et al. Genome-Wide Circulating Microrna Expression Profiling Reveals Potential Biomarkers for Amyotrophic Lateral Sclerosis. Neurobiol. Aging 2018, 64, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, I.; Hama, Y.; Matsushima, M.; Hirotani, M.; Kano, T.; Hohzen, H.; Yabe, I.; Utsumi, J.; Sasaki, H. Identification of Plasma Micrornas as a Biomarker of Sporadic Amyotrophic Lateral Sclerosis. Mol. Brain 2015, 8, 67. [Google Scholar] [CrossRef]
- Banack, S.A.; Dunlop, R.A.; Stommel, E.W.; Mehta, P.; Cox, P.A. Mirna Extracted from Extracellular Vesicles Is a Robust Biomarker of Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2022, 442, 120396. [Google Scholar] [CrossRef] [PubMed]
- McCluskey, G.; Morrison, K.E.; Donaghy, C.; Rene, F.; Duddy, W.; Duguez, S. Extracellular Vesicles in Amyotrophic Lateral Sclerosis. Life 2023, 13, 121. [Google Scholar] [CrossRef] [PubMed]
- Saucier, D.; Wajnberg, G.; Roy, J.; Beauregard, A.-P.; Chacko, S.; Crapoulet, N.; Fournier, S.; Ghosh, A.; Lewis, S.M.; Marrero, A.; et al. Identification of a Circulating Mirna Signature in Extracellular Vesicles Collected from Amyotrophic Lateral Sclerosis Patients. Brain Res. 2019, 1708, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Katsu, M.; Hama, Y.; Utsumi, J.; Takashina, K.; Yasumatsu, H.; Mori, F.; Wakabayashi, K.; Shoji, M.; Sasaki, H. Microrna Expression Profiles of Neuron-Derived Extracellular Vesicles in Plasma from Patients with Amyotrophic Lateral Sclerosis. Neurosci. Lett. 2019, 708, 134176. [Google Scholar] [CrossRef]
- Giagnorio, E.; Malacarne, C.; Mantegazza, R.; Bonanno, S.; Marcuzzo, S. Myomirs and Their Multifaceted Regulatory Roles in Muscle Homeostasis and Amyotrophic Lateral Sclerosis. J. Cell Sci. 2021, 134, jcs258349. [Google Scholar] [CrossRef]
- De Felice, B.; Annunziata, A.; Fiorentino, G.; Borra, M.; Biffali, E.; Coppola, C.; Cotrufo, R.; Brettschneider, J.; Giordana, M.L.; Dalmay, T.; et al. Mir-338-3p Is over-Expressed in Blood, Cfs, Serum and Spinal Cord from Sporadic Amyotrophic Lateral Sclerosis Patients. Neurogenetics 2014, 15, 243–253. [Google Scholar] [CrossRef]
- Yelick, J.; Men, Y.; Jin, S.; Seo, S.; Espejo-Porras, F.; Yang, Y. Elevated Exosomal Secretion of Mir-124-3p from Spinal Neurons Positively Associates with Disease Severity in Als. Exp. Neurol. 2020, 333, 113414. [Google Scholar] [CrossRef]
- Vrabec, K.; Boštjančič, E.; Koritnik, B.; Leonardis, L.; Grošelj, L.D.; Zidar, J.; Rogelj, B.; Glavač, D.; Ravnik-Glavač, M. Differential Expression of Several Mirnas and the Host Genes Aatk and Dnm2 in Leukocytes of Sporadic Als Patients. Front. Mol. Neurosci. 2018, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.; Santos, C.; Gomes, C.; Fernandes, A.; Correia, A.M.; Sebastião, A.M.; Vaz, A.R.; Brites, D. Downregulated Glia Interplay and Increased Mirna-155 as Promising Markers to Track Als at an Early Stage. Mol. Neurobiol. 2018, 55, 4207–4224. [Google Scholar] [CrossRef] [PubMed]
- Butovsky, O.; Jedrychowski, M.P.; Cialic, R.; Krasemann, S.; Murugaiyan, G.; Fanek, Z.; Greco, D.J.; Wu, P.M.; Doykan, C.E.; Kiner, O.; et al. Targeting Mir-155 Restores Abnormal Microglia and Attenuates Disease in Sod1 Mice. Ann. Neurol. 2015, 77, 75–99. [Google Scholar] [CrossRef] [PubMed]
- de Andrade, H.M.; de Albuquerque, M.; Avansini, S.H.; Rocha, C.d.S.; Dogini, D.B.; Nucci, A.; Carvalho, B.; Lopes-Cendes, I.; França, M.C. Micrornas-424 and 206 Are Potential Prognostic Markers in Spinal Onset Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2016, 368, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, L.; Baranzini, M.; Berardinelli, M.G.; Lattanzi, W.; Monforte, M.; Tasca, G.; Conte, A.; Logroscino, G.; Michetti, F.; Ricci, E.; et al. Potential Therapeutic Targets for Als: And Are Modulated During Disease Progression in the Skeletal Muscle of Patients. Sci. Rep. 2017, 7, 872–893. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.H.; Valdez, G.; Moresi, V.; Qi, X.X.; McAnally, J.; Elliott, J.L.; Bassel-Duby, R.; Sanes, J.R.; Olson, E.N. Microrna-206 Delays Als Progression and Promotes Regeneration of Neuromuscular Synapses in Mice. Science 2009, 326, 1549–1554. [Google Scholar] [CrossRef] [PubMed]
- Toivonen, J.M.; Manzano, R.; Oliván, S.; Zaragoza, P.; García-Redondo, A.; Osta, R. Microrna-206: A Potential Circulating Biomarker Candidate for Amyotrophic Lateral Sclerosis. PLoS ONE 2014, 9, e89065. [Google Scholar] [CrossRef]
- Dobrowolny, G.; Martone, J.; Lepore, E.; Casola, I.; Petrucci, A.; Inghilleri, M.; Morlando, M.; Colantoni, A.; Scicchitano, B.M.; Calvo, A.; et al. A Longitudinal Study Defined Circulating Micrornas as Reliable Biomarkers for Disease Prognosis and Progression in Als Human Patients. Cell Death Discov. 2021, 7, 4. [Google Scholar] [CrossRef]
- Joilin, G.; Gray, E.; Thompson, A.G.; Bobeva, Y.; Talbot, K.; Weishaupt, J.; Ludolph, A.; Malaspina, A.; Leigh, P.N.; Newbury, S.F.; et al. Identification of a Potential Non-Coding Rna Biomarker Signature for Amyotrophic Lateral Sclerosis. Brain Commun. 2020, 2, fcaa053. [Google Scholar] [CrossRef]
- Magen, I.; Yacovzada, N.S.; Yanowski, E.; Coenen-Stass, A.; Grosskreutz, J.; Lu, C.-H.; Greensmith, L.; Malaspina, A.; Fratta, P.; Hornstein, E. Circulating Mir-181 Is a Prognostic Biomarker for Amyotrophic Lateral Sclerosis. Nat. Neurosci. 2021, 24, 1534–1541. [Google Scholar] [CrossRef]
- Verde, F.; Otto, M.; Silani, V. Neurofilament Light Chain as Biomarker for Amyotrophic Lateral Sclerosis and Frontotemporal Dementia. Front. Neurosci. 2021, 15, 679199. [Google Scholar] [CrossRef] [PubMed]
- De Felice, B.; Manfellotto, F.; Fiorentino, G.; Annunziata, A.; Biffali, E.; Pannone, R.; Federico, A. Wide-Ranging Analysis of Microrna Profiles in Sporadic Amyotrophic Lateral Sclerosis Using Next-Generation Sequencing. Front. Genet. 2018, 9, 310. [Google Scholar] [CrossRef] [PubMed]
- Daneshafrooz, N.; Joghataei, M.T.; Mehdizadeh, M.; Alavi, A.; Barati, M.; Panahi, B.; Teimourian, S.; Zamani, B. Identification of Let-7f and Mir-338 as Plasma-Based Biomarkers for Sporadic Amyotrophic Lateral Sclerosis Using Meta-Analysis and Empirical Validation. Sci. Rep. 2022, 12, 1373. [Google Scholar] [CrossRef] [PubMed]
- De Felice, B.; Guida, M.; Guida, M.; Coppola, C.; De Mieri, G.; Cotrufo, R. A Mirna Signature in Leukocytes from Sporadic Amyotrophic Lateral Sclerosis. Gene 2012, 508, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Freischmidt, A.; Müller, K.; Ludolph, A.C.; Weishaupt, J.H. Systemic Dysregulation of Tdp-43 Binding Micrornas in Amyotrophic Lateral Sclerosis. Acta Neuropathol. Commun. 2013, 1, 42. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, A. Extracellular Mirna Biomarkers in Neurologic Disease: Is Cerebrospinal Fluid Helpful? Biomark. Med. 2021, 15, 1377–1388. [Google Scholar] [CrossRef] [PubMed]
- Sjoqvist, S.; Otake, K. Saliva and Saliva Extracellular Vesicles for Biomarker Candidate Identification-Assay Development and Pilot Study in Amyotrophic Lateral Sclerosis. Int. J. Mol. Sci. 2023, 24, 5237. [Google Scholar] [CrossRef] [PubMed]
- Vinaiphat, A.; Sze, S.K. Clinical Implications of Extracellular Vesicles in Neurodegenerative Diseases. Expert Rev. Mol. Diagn. 2019, 19, 813–824. [Google Scholar] [CrossRef]
- Pasetto, L.; Callegaro, S.; Corbelli, A.; Fiordaliso, F.; Ferrara, D.; Brunelli, L.; Sestito, G.; Pastorelli, R.; Bianchi, E.; Cretich, M.; et al. Decoding Distinctive Features of Plasma Extracellular Vesicles in Amyotrophic Lateral Sclerosis. Mol. Neurodegener. 2021, 16, 52. [Google Scholar] [CrossRef]
- Sproviero, D.; Gagliardi, S.; Zucca, S.; Arigoni, M.; Giannini, M.; Garofalo, M.; Olivero, M.; Dell’Orco, M.; Pansarasa, O.; Bernuzzi, S.; et al. Different Mirna Profiles in Plasma Derived Small and Large Extracellular Vesicles from Patients with Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 2737. [Google Scholar] [CrossRef]
- Dunlop, R.A.; Banack, S.A.; Cox, P.A. L1cam Immunocapture Generates a Unique Extracellular Vesicle Population with a Reproducible Mirna Fingerprint. RNA Biol. 2023, 20, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Norman, M.; Ter-Ovanesyan, D.; Trieu, W.; Lazarovits, R.; Kowal, E.J.K.; Lee, J.H.; Chen-Plotkin, A.S.; Regev, A.; Church, G.M.; Walt, D.R. L1cam Is Not Associated with Extracellular Vesicles in Human Cerebrospinal Fluid or Plasma. Nat. Methods 2021, 18, 631–634. [Google Scholar] [CrossRef] [PubMed]






| Dysregulated miRNAs | Target Genes | Main Regulatory Effects | References |
|---|---|---|---|
| ↑ miR-124 | C/EBPα (CCAAT/enhancer-binding protein alpha), STAT3 (Signal transducer and activator of transcription 3) | Regulates neuronal differentiation and maturation; dysregulation may contribute to neuroinflammation and neurodegeneration in ALS | [49] |
| ↑ miR-338-3p | PYGB (Glycogen Phosphorylase Brain) | Dysregulation leads to decreased glycogenolysis and subsequent aberrant accumulation of glycogen that causes glucotoxicity and impairs neuronal functions, leading to neurodegeneration | [50] |
| ↑ miR-146a | IRAK1 (Interleukin-1 receptor-associated kinase 1), TRAF6 (TNF receptor-associated factor 6) | Regulates immune responses and inflammation; dysregulation may contribute to neuroinflammatory processes in ALS | [51,52,53] |
| ↑ miR-155 | TAB2 (TGF-beta-activated kinase 1/MAP3K7-binding protein 2), SOCS1 (Suppressor of cytokine signaling 1) | Involved in immune regulation and neuroinflammation; dysregulation may contribute to glial cell dysfunction and disease progression in ALS | [54] |
| ↓ miR-9 | TDP-43 (Transactive response DNA-binding protein 43) | Dysregulation may disrupt the normal clearance of TDP-43, leading to its accumulation and subsequent neuroinflammation and neurotoxicity | [55] |
| ↑ miR-23a | PGC-1α (Skeletal muscle peroxisome proliferator-activated receptor γ coactivator-1α) | Regulates signaling networks involved in mitochondrial biogenesis and function | [56] |
| ↑ miR-206 | HDAC4 (Histone deacetylase 4), Cx43 (Connexin 43) | Regulates muscle regeneration and repair, and its dysregulation may contribute to muscle atrophy and impaired motor function in ALS | [57] |
| ↑ miR-143 | FUS (Fused in sarcoma), SMAD3 (Mothers against decapentaplegic homolog 3) | Involved in RNA metabolism and cellular stress response; dysregulation may contribute to the accumulation of abnormal protein aggregates and neuronal cell death in ALS | [58] |
| ↑ miR-26a | SLC1A1 (Glutamate transporter) | Regulates the glutamate receptor signaling pathway. Dysregulation leads to high levels of glutamate in the CNS, causing inflammation and neurodegeneration | [59] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lauria, G.; Curcio, R.; Tucci, P. A Machine Learning Approach for Highlighting microRNAs as Biomarkers Linked to Amyotrophic Lateral Sclerosis Diagnosis and Progression. Biomolecules 2024, 14, 47. https://doi.org/10.3390/biom14010047
Lauria G, Curcio R, Tucci P. A Machine Learning Approach for Highlighting microRNAs as Biomarkers Linked to Amyotrophic Lateral Sclerosis Diagnosis and Progression. Biomolecules. 2024; 14(1):47. https://doi.org/10.3390/biom14010047
Chicago/Turabian StyleLauria, Graziantonio, Rosita Curcio, and Paola Tucci. 2024. "A Machine Learning Approach for Highlighting microRNAs as Biomarkers Linked to Amyotrophic Lateral Sclerosis Diagnosis and Progression" Biomolecules 14, no. 1: 47. https://doi.org/10.3390/biom14010047
APA StyleLauria, G., Curcio, R., & Tucci, P. (2024). A Machine Learning Approach for Highlighting microRNAs as Biomarkers Linked to Amyotrophic Lateral Sclerosis Diagnosis and Progression. Biomolecules, 14(1), 47. https://doi.org/10.3390/biom14010047