
Unwrap RAP1’s Mystery at Kinetoplastid Telomeres


1
Center for Gene Regulation in Health and Disease, Department of Biological, Geological, and Environmental Sciences, College of Arts and Sciences, Cleveland State University, 2121 Euclid Avenue, Cleveland, OH 44115, USA
2
Case Comprehensive Cancer Center, Case Western Reserve University, 10900 Euclid Avenue, Cleveland, OH 44106, USA
3
Department of Inflammation and Immunity, Lerner Research Institute, Cleveland Clinic, 9500 Euclid Avenue, Cleveland, OH 44195, USA
4
Center for RNA Science and Therapeutics, Case Western Reserve University, 10900 Euclid Avenue, Cleveland, OH 44106, USA
Biomolecules 2024, 14(1), 67; https://doi.org/10.3390/biom14010067
Submission received: 6 December 2023
/
Revised: 27 December 2023
/
Accepted: 27 December 2023
/
Published: 4 January 2024
(This article belongs to the Special Issue Role of Telomere Dynamics in Chromosome Stability and Transcriptional Regulation)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Although located at the chromosome end, telomeres are an essential chromosome component that helps maintain genome integrity and chromosome stability from protozoa to mammals. The role of telomere proteins in chromosome end protection is conserved, where they suppress various DNA damage response machineries and block nucleolytic degradation of the natural chromosome ends, although the detailed underlying mechanisms are not identical. In addition, the specialized telomere structure exerts a repressive epigenetic effect on expression of genes located at subtelomeres in a number of eukaryotic organisms. This so-called telomeric silencing also affects virulence of a number of microbial pathogens that undergo antigenic variation/phenotypic switching. Telomere proteins, particularly the RAP1 homologs, have been shown to be a key player for telomeric silencing. RAP1 homologs also suppress the expression of Telomere Repeat-containing RNA (TERRA), which is linked to their roles in telomere stability maintenance. The functions of RAP1s in suppressing telomere recombination are largely conserved from kinetoplastids to mammals. However, the underlying mechanisms of RAP1-mediated telomeric silencing have many species-specific features. In this review, I will focus on Trypanosoma brucei RAP1’s functions in suppressing telomeric/subtelomeric DNA recombination and in the regulation of monoallelic expression of subtelomere-located major surface antigen genes. Common and unique mechanisms will be compared among RAP1 homologs, and their implications will be discussed.
1. The Telomere Structure and Telomere Functions
Telomeres are nucleoprotein complexes located at chromosome ends [1]. In most eukaryotes, the telomere contains simple repetitive sequences, such as (TTAGGG)n in vertebrates, with the G-rich strand going 5′ to 3′ toward the chromosome end [2,3]. Although most parts of the telomere are double-stranded, the very end of the telomere has a single-stranded 3′ overhang [4,5]. The telomere 3′ overhang can fold back and invade the duplex telomeric DNA and form the T-loop structure, which has been observed in human, Oxytricha fallax (a hypotrichous ciliate), and Trypanosoma brucei (a kinetoplastid parasite) [6,7,8], while a similar loop structure of the telomere chromatin has also been observed in mouse and chicken [9].
Telomeres are essential for maintaining the linear genome stability [10]. They form a specialized structure that protects DNA ends from nucleolytic degradation, prevents natural chromosome ends from being recognized as DNA damage sites, and suppresses illegitimate processes, such as DNA recombination and end joining [11,12,13,14,15]. The T-loop structure can effectively sequestrate and bury the telomere end to protect it from degradation and DNA recombination [13,16]. In addition, telomere proteins play critical roles in chromosome end protection in all eukaryotes that have been investigated so far [17,18], even though telomere proteins are only partially conserved from protozoa to mammals [19,20]. Vertebrate telomeres have both Shelterin and CST protein complexes [21]. Shelterin contains six core telomere proteins: TRF1 and TRF2 bind the duplex TTAGGG repeats [22,23,24], TPP1 and POT1 bind the single-stranded telomere 3′ overhang as a heterodimer [25,26,27,28,29,30], RAP1 interacts with TRF2 [31], and TIN2 interacts with TRF1, TRF2, and TPP1 to link various subunits of Shelterin together [25,27,32,33]. The CST complex includes CTC1, STN1, and TEN1 [34,35]. They bind the telomere 3′ overhang as a heterotrimer [36], which is structurally similar to the RPA complex [34,37,38,39]. Budding yeast Saccaromyces cerevisiae telomere has a sequence of (TG1–3)n [40], which is not a perfect repeat, so its telomere protein complex is quite different from those in vertebrates [41]. ScRap1 binds the duplex telomeric DNA [42,43,44], while the TRF homolog, ScTbf1, binds subtelomeric TTAGGG repeats [45]. TIN2, TPP1, and POT1 homologs are absent in S. cerevisiae, but ScCST (containing Cdc13, Stn1, and Ten1) binds the telomere 3′ overhang and its function is largely conserved as that of mammalian CST [46,47,48]. Telomeres in T. brucei have the same TTAGGG repeat sequence as those in vertebrates [3,49,50], and the T. brucei telomere complex is more conserved to that in vertebrates than in budding yeast. T. brucei has a TRF homolog, TbTRF, that binds the telomeric dsDNA [51], a RAP1 homolog, TbRAP1, that interacts with TbTRF [52], and TbTIF2, which is functionally homologous to TIN2 and also interacts with TbTRF [53]. In addition, a couple of essential DNA polymerases, PolIE and PPL2, have been found to be intrinsic to the telomere chromatin [54,55], and a nonessential protein, TelAP1, has been identified to bind the telomeric DNA [55]. However, T. brucei does not seem to have TPP1, POT1, or CST homologs [54]. Recent studies on PolIE indicate that it has similar functions as CST in that it suppresses telomerase-mediated telomere elongation and stimulates the telomere C-strand fill-in process [20,54], although detailed mechanisms are still unclear.
Telomere repeats serve as a docking site for proteins that bind telomeric DNA directly or indirectly. Critically short telomeres recruit insufficient amounts of telomere proteins, exposing the natural chromosome ends and frequently inducing DNA damage responses, including cell growth arrest [56]. Therefore, maintaining a stable telomere length is essential for telomere end protection, and mammalian and yeast cells with critically short telomeres enter replicative senescence [57,58,59,60]. However, conventional DNA polymerases are incapable of replicating linear DNA molecules completely due to their enzymatic properties (they require a template and a primer and only extend DNA at the 3′ ends), resulting in progressive telomere shortening in proliferating cells [4]. In most eukaryotic cells, telomerase, a specialized reverse transcriptase, can synthesize the G-rich telomeric DNA de novo [1,61,62,63], solving this so-called “end replication problem”. Telomerase has both a protein subunit bearing the reverse transcriptase activity and an RNA subunit that provides the template for de novo telomere DNA synthesis [64,65,66]. On the other hand, telomerase-independent telomere maintenance has been observed in several situations. Drosophila telomeres naturally contain retrotransposon arrays, and transposition is the predominant mechanism for telomere maintenance [67,68]. In telomerase-negative ALT cancer cells, DNA recombination serves as the key mechanism of telomere maintenance [69,70]. In addition, telomerase-null yeast cells can continue to proliferate using DNA recombination-mediated amplification of telomeric/subtelomeric repeats as the mechanism of telomere maintenance [71,72]. Therefore, although telomere recombination is normally a risk factor for genome instability, it can serve as an important telomere maintenance mechanism. In addition, in several eukaryotic pathogens that undergo antigenic variation and harbor their major surface antigen genes at the subtelomere (including Trypanosome brucei that causes human African trypanosomiasis, Plasmodium falciparum that causes malaria, and Pneumocystis jirovecii that causes pneumonia in immune-deficient patients), DNA recombination at the telomere and subtelomere can benefit antigenic variation and enhance pathogen virulence [19,73,74,75,76].
In many eukaryotes, telomeres form a heterochromatic structure and suppress expression of genes located in subtelomeric regions [77,78]. Position effect variegation was originally observed in Drosophila, where euchromatic genes are silenced when they are rearranged or translocated close to the heterochromatin [79]. Subsequently, it was found that in Saccharomyces cerevisiae, genes located near the telomere are repressed, which is termed the telomere position effect or telomeric silencing [78]. The telomeric silencing phenomenon has been observed not only in yeast [41,80] and Drosophila [81] but also in human cells [82,83,84,85,86,87] and protozoan parasites, including T. brucei [88,89,90,91] and P. falciparum [92,93]. Telomeric silencing in microbial pathogens can also be an important player regulating pathogen virulence [52,92,93,94,95,96], and telomere proteins have been shown to play critical roles in telomeric silencing [78].
RAP1 is one of the most conserved telomere proteins, whose homologs have been identified in eukaryotes from kinetoplastids to mammals [31,52,97,98,99,100]. RAP1s have been shown to play essential functions in maintaining telomere stability and in telomeric silencing [101,102]. Particularly, functions of T. brucei RAP1 in telomeric silencing and telomere stability maintenance are intimately involved in the regulation of antigenic variation [52,103,104,105,106,107], an essential pathogenesis mechanism [108], as the major surface antigen VSG is expressed exclusively from subtelomeric loci [109,110,111,112]. All known RAP1 homologs have an N-terminal BRCA1 C-terminus (BRCT) domain that is frequently identified in proteins involved in the DNA damage response or cell cycle checkpoint [113,114], a central Myb domain that typically binds dsDNA [115,116], and a RAP1 C-terminus (RCT) domain that is a protein–protein interaction domain conserved among all RAP1 homologs [31] (Figure 1). Below, I will discuss RAP1’s functions in telomeric silencing and chromosome end protection, focusing on the similarities and differences between TbRAP1 and yeast and mammalian RAP1 homologs.
2. Yeast and Mammalian RAP1 Homologs Are Essential for Telomeric Silencing and Suppress Telomere Recombination
2.1. ScRap1 Is a Key Player of Telomeric Silencing in S. cerevisiae
The telomeric silencing phenomenon has been studied extensively in S. cerevisiae, and ScRap1 is the central factor nucleating the silencing effect [41]. ScRap1 was identified as a DNA binding factor that binds to both transcription activator and repressor elements [97]. It was later shown that ScRap1 binds the yeast duplex telomeric DNA directly [42,43,44], helps protect the chromosome end [117,118,119], plays a critical role in telomere length regulation [98,120], and is essential for telomeric silencing [121,122].
After the crystal structure of the central region of ScRap1 was solved, it was clearly shown that ScRap1 has a Myb-like domain that is nearly identical to its Myb domain (Figure 1), and the Myb/Myb-like region of ScRap1 is responsible for binding dsDNA with two tandem ACAYYY sequences [43]. The two domains provided considerable flexibility [123,124,125], allowing ScRap1 to recognize the imperfect telomere repeats with little structural rearrangements or loss of affinity [44]. ScRap1 is an essential protein, and the C-terminus of its DNA binding domain is indispensable [126]. A recent genomic study comprehensively summarized ScRAP1’s role as a transcription regulator: ScRap1 activates ribosomal protein and RNR genes and suppresses glycolysis genes and homothallic mating loci [127]. On the other hand, ScRap1’s effect on subtelomeric gene expression is predominantly repressive [120,121,128,129,130].
S. cerevisiae telomeres are the largest ScRap1 binding sites, which allow an array of ScRap1 to bind the telomeric dsDNA directly. Telomere-bound ScRap1, through its interaction with Sir3 and Sir4 silencers [128,129,130,131,132,133,134,135,136,137,138], recruits Sir2 [133,139,140,141,142,143,144,145,146] to the telomere. Subsequently, Sir2 removes acetyl groups from histone tails in a NAD+-dependent manner [147,148,149], establishing a heterochromatic structure at the telomere. In addition, Sir3’s broom-adjacent homology (BAH) domain and Sir4 interact with histone H3 and H4 tails [143,150,151,152], which help propagate the heterochromatin from the telomere toward chromosome internal regions [143,153,154,155]. Interestingly, longer telomere repeats in S. cerevisiae are associated with stronger telomeric silencing [121], suggesting that more ScRap1 binding at the telomere can recruit more silencers and establish a more tightly compacted chromatin structure at the telomere. Furthermore, ScRap1, together with its interacting factors (Rif and Sir proteins), suppresses RNA pol II-mediated telomere transcription and the TERRA level [156]. TERRA was initially identified in several protozoan parasites [157] and subsequently in all eukaryotes examined [158,159,160,161,162]. TERRA has been shown to play important roles in telomere protection, length regulation, and recombination in mammalian cells and yeasts [163,164,165]. TERRA can invade the duplex telomeric DNA and form a three-stranded R-loop structure with an RNA:DNA hybrid [166], and R-loops have a propensity to induce DNA breaks [167,168]. In general, TERRA and telomere R-loops are expected to interfere with the replication machinery and disturb the passage of the replication fork and telomere processing [169]. Therefore, ScRap1’s role in suppression of TERRA is linked with its role in telomere stability maintenance (see below).
Interestingly, in budding yeast, Candida glabrata, that causes opportunistic bloodstream, urinary track, and vaginal infections, telomeric silencing also regulates its virulence [96,170,171]. The NAD+-dependent histone deacetylase Sir2 interacts with Sir4, which is recruited to the telomere by CgRap1 and yKu. Deacetylation of the histone tails by Sir2, in turn, helps establish and propagate the telomeric/subtelomeric silent domain over 20 kb [96]. The EPA gene family in C. glabrata is located at subtelomeres and encodes epithelia adhesins required for host–pathogen interaction [95]. With normal telomeric silencing, only selected EPA genes are expressed. However, under the condition of niacin limitation (where niacin is a precursor of NAD+), lowered Sir2 activity leads to weaker telomeric silencing, expression of more EPA genes (such as EPA6), and enhanced C. glabrata adherence to host cells [172].
Telomeric silencing has also been observed in human cells. When a luciferase reporter gene is inserted at a subtelomeric position, it is expressed ~10× lower than when it is inserted at a chromosome internal locus, and longer telomeres induce a stronger silencing effect [83], which depends on histone deacetylation [83]. In addition, expression of a subtelomeric neomycin reporter gene on a Linear Human Artificial Chromosome (L-HAC) is repressed by nearby telomeres and inversely correlated with the telomere length and subtelomeric DNA methylation [173]. Similar to that observed in yeast, human RAP1 can also suppress TERRA expression [174], although the underlying mechanism is unclear. Interestingly, human telomeres can interact with chromosomal internal telomeric sequences via TRF2 [175]. This telomere loop-back can silence genes located over a long distance (a phenomenon termed TPE-OLD), including the hTERT gene located ~1.2 Mb away from the telomere [175]. Telomeric silencing appears to also influence the development of FSHD (facioscapulohumeral muscular dystrophy). Sufficient epigenetic alternation of the D4Z4 array located at the chromosome 4 subtelomere (4q35) can lead to abnormal expression of the nearby DUX4 gene, which is linked to FSHD development [176]. It has been shown that telomeric silencing affects DUX4 expression [86]. In addition, weaker telomeric silencing can allow more inter- and intra-chromosomal subtelomeric rearrangements of the 4q35 locus [177]. Furthermore, shorter telomeres are correlated with D4Z4 locus hypomethylation, and TPE-OLD regulates SORBS2 gene expression in FSHD cells [178].
2.2. RAP1 Homologs in Higher Eukaryotes Suppress Telomere Recombination
ScRap1 has essential functions in protecting the chromosome end. Conditional deletion of ScRap1 leads to yKu70/80, DNA ligase 4, Lif1, and the MRN complex (Mre11/Rad50/Xrs2)-dependent [179,180,181,182,183], non-homologous end-joining (NHEJ)-mediated chromosome end-to-end fusions [117], while the interactions between ScRap1’s RCT domain with Rif2 and Sir4 are both required for this function [118]. Similarly, human RAP1 can inhibit NHEJ together with TRF2 in vitro [184,185] and help suppress telomere end fusions in senescent cells with short telomeres [186]. In addition, RAP1 homologs have a more conserved function in suppressing homologous recombination (HR) at the telomere. The central region of ScRap1 inhibits the recruitment of HR proteins independent of yKu, Cdc13, and Rif1/2 [119]. The Rap1 homolog in Candida albicans is critical to maintain the telomere length and structure by suppressing telomere recombination [187]. In addition, in methylotrophic yeast Hansenula polymorpha DL-1, HpRAP1B (one of the two RAP1 homologs) that binds to the telomere repeats also suppresses telomere recombination [188]. Deletion of mouse RAP1 leads to more homology-directed repair at the telomere (shown as an elevated amount of Telomere-Sister Chromatid Exchanges) [189], as TRF2 and RAP1 suppress PARP1 and SLX4, respectively [190].
3. T. brucei RAP1 Ensures VSG Monoallelic Expression and Suppresses Telomere Recombination through Unusual Mechanisms
3.1. Trypanosoma brucei Undergoes Antigenic Variation to Evade the Host’s Immune Response
T. brucei is a protozoan parasite that causes human African trypanosomiasis, which is frequently fatal without treatment. While proliferating in its mammalian host, T. brucei stays in extracellular spaces and is immediately exposed to the host’s immune surveillance. However, T. brucei sequentially expresses distinct variant surface glycoproteins (VSGs), its major surface antigen, thereby effectively evading the host’s immune response [191]. This antigenic variation is a key pathogenesis mechanism that allows the parasite to establish a long-term infection.
T. brucei has a large VSG gene pool, including >2500 VSG genes and pseudogenes [110], and all are located at subtelomeric regions [111,112]. Most VSG genes and pseudogenes are in long VSG gene arrays at subtelomeres of mega-base chromosomes that contain all essential genes [112]. Individual VSG genes are also found in two-thirds of telomeres of the ~100 mini-chromosomes that predominantly consist of repeat sequences [110,192]. However, at the bloodstream form stage (when T. brucei proliferates inside its mammalian host), VSGs are expressed exclusively from bloodstream form VSG expression sites (ESs), which are large polycistronic transcription units with the VSG gene at the end within 2 kb from the telomere repeats and the ES promoter 40–60 kb upstream [111]. The Lister 427 T. brucei strain used in many research laboratories has ~15 different bloodstream form VSG ESs, all with the same gene organization and ~90% sequence identity [111,112]. However, at any moment, only one ES is fully transcribed by RNA pol I, resulting in a single type of VSG being expressed on the cell surface [193,194]. Monoallelic VSG expression is important for T. brucei survival in its mammalian host, as parasites artificially expressing multiple VSGs are more efficiently eliminated by the infected mouse host [195].
VSG switching can occur at the transcription level, where the active ES is silenced while a silent ES is de-repressed (termed in situ switch) [196,197]. In addition, DNA recombination can replace the original active VSG sequence with a new one, resulting in expression of a different VSG [196,197,198,199]. HR in T. brucei appears to be very active, allowing efficient gene targeting [200,201,202]. HR factors, including RAD51, RAD51-3 (a paralogue of RAD51), and BRCA2, have been shown to be important for normal VSG switching [203,204,205,206]. T. brucei lacks the NHEJ machinery [207] but has the microhomology-mediated end-joining (MMEJ) pathway [208,209]. However, whether VSG switching can be mediated solely by MMEJ is unknown. In addition, during a switching event, pieces of various VSG donors can be patched together to form a new functional mosaic VSG in the active ES [210,211]. Several factors involved in DNA replication, DNA damage repair, and DNA recombination have been shown to suppress VSG switching, including TbORC1 [212], RECQ2 [213], TOPO3α [214], and RMI1 [215]. On the other hand, inducing DNA double-strand breaks (DSBs) in or immediately upstream of the active VSG gene can increase the VSG switching rate ~250 fold, and DSBs in the VSG vicinity are a potent trigger of VSG switching [74,75,216,217,218,219]. However, how VSG switching is initiated naturally in T. brucei is less clear.
The telomere structure and telomere proteins also influence the VSG switching rate. T. brucei cells carrying an extremely short telomere downstream of the active ES have a ~10× higher VSG switching rate compared to cells with longer telomeres (10–15 kb, on average) [220]. The active VSG ES-adjacent telomere has been observed to experience frequent truncations during cell proliferation [221]. Presumably, short telomeres have a higher chance to have DNA breaks land in the active VSG or nearby, which in turn induces VSG switching. In addition, T. brucei telomere proteins (including TbTRF, TbRAP1, TbTIF2, and PolIE) suppress VSG switching by maintaining the telomere integrity and stability, although the underlying mechanisms are not identical [53,54,104,222,223].
3.2. Multiple Mechanisms Are Employed to Ensure VSG Monoallelic Expression
Monoallelic gene expression or allelic exclusion has been observed from bacteria to mammals and is important for organism fitness and survival [224,225]. Notable examples include genome imprinting to express one of the two parental alleles, X chromosome inactivation, and random monoallelic expression of autosomal genes in mammals [224,226]. Many monoallelically expressed genes encode cell surface receptors. For example, each αβ T cell expresses one α and one β polypeptide of the T cell receptor, and each human and mouse olfactory sensory neuron expresses only one odorant receptor gene [224]. Feedback signaling involving the gene product (protein) [227] and epigenetic regulation [228] are sometimes employed to achieve monoallelic expression, but detailed mechanisms remain poorly understood.
In T. brucei, VSG monoallelic expression is tightly regulated through multiple mechanisms [229,230,231]. First, the subnuclear localization of the active ES is unique. Transcription of the active ES by RNA pol I occurs at a specialized ES body (ESB) located outside of the nucleolus (where RNA pol I transcribes rRNA) [232]. ESB1 has recently been identified to be essential for the active ES transcription and is responsible for recruiting RNA pol I and forming a local, highly SUMOylated focus at ESB [233], where SUMOylation has been shown to positively regulate VSG expression [234,235]. On the other hand, silent ESs are dispersed in the nucleus, away from ESB [236]. Nearly all genes in T. brucei are organized in polycistronic transcription units [112,237,238], and the polycistronic transcripts are trans-spliced to have a spliced leader added at the 5′ end of the individual gene transcript [239,240]. Consistently, the active ES associates with the spliced leader gene array in Hi-C analysis that examines chromosome conformation [241], and ESB is located adjacent to one of the two splicing centers shown by immunofluorescence analysis [241]. Presumably, the high-level transcription of the active VSG ES is intimately coupled with trans-splicing to improve VSG expression efficiency. Recent studies also showed that VEX1 [242] and VEX2 [243] are essential for VSG monoallelic expression. Both proteins help sustain transcription of the active VSG ES in an allelic exclusive manner [242,243]. VEX1 associates with the spliced leader gene array and one of the splicing centers in the nucleus, while VEX2 associates with the active VSG ES [241,243]. VEX1 can interact with VEX2, but assembly of the VEX complex relies on RNA pol I transcription [243].
Second, VSG ES transcription is regulated at both initiation and elongation steps. The RNA pol I transcription factor complex, CITFA, has been identified [244]. Two essential CITFA subunits occupy the active ES promoter at a much higher level than at those of silent ESs, and the high promoter occupancy of CITFA is correlated with high levels of RNA pol I occupancy and ES transcription [245], indicating that transcription initiation is different in the active and silent ESs. In addition, transcription elongation along ESs is also regulated, as silent ES promotors are also moderately active, but transcription elongation quickly attenuates after a few kbs, effectively blocking transcription of downstream VSGs [246,247].
Third, the chromatin structure plays important roles in VSG ES expression regulation. Although all ESs have ~90% sequence identity [111], silent ESs are packed with nucleosomes, while the active ES is depleted of nucleosomes [248,249,250]. Histone H1 and H3 are required for silencing reporter genes targeted immediately downstream of silent ES promoters, but not the downstream VSG genes [251,252]. The TbISWI complex (including TbISWI, NLP, FYRP, and RCCP) has been identified to suppress reporter genes at the ES promoters, where TbISWI has a highly conserved SWI2/SNF2 family ATPase domain and a SANT domain with DNA binding activity [253,254,255]. In addition, histone chaperones FACT (including TbSpt16 [256] and Pob3 [257]), ASF1A, and CAF-1b have all been identified to be required for ES promoter silencing [252]. Besides chromatin remodeling factors involved in ES promoter silencing, TDP1, an architectural HMG chromatin protein, is enriched at the active ES and rDNA and is essential for full-level VSG and rRNA transcription by RNA pol I, presumably by binding DNA directly and excluding nucleosomes [258]. Furthermore, DOT1b, which trimethylates lysine 76 of histone H3, is required for a tight VSG ES silencing [259].
3.3. Competition between TbRAP1’s DNA and RNA Binding Activities Is Essential for VSG Monoallelic Expression
As VSGs are expressed exclusively from subtelomeric regions, the telomere structure and telomere proteins have been shown to regulate VSG switching and VSG monoallelic expression. T. brucei has the same telomere sequence and terminal 3′ overhang structure as vertebrates [3,49,50,260,261], and the T-loop structure has been observed at both telomeres of the same chromosome [6]. Both the protein and RNA components of T. brucei telomerase, TbTERT and TbTR, have been identified, and the telomerase-mediated telomere synthesis is the predominant telomere maintenance mechanism in T. brucei [260,262,263,264,265]. As mentioned above, TbTRF [51], TbRAP1 [52], and TbTIF2 [53] are Sheltering-equivalent telomere proteins. Several other T. brucei proteins seem to specifically associate with the telomere chromatin but are not homologous to known core telomere proteins in higher eukaryotes, including TelAP1 [55], PolIE [54,266], and PPL2 [54,55,267]. In addition, TbKu70/80 [268,269] and ORC1 [212] are localized at the telomere.
Although most known T. brucei telomere proteins (including TbTRF, TbTIF2, PolIE, and ORC1), when depleted, lead to de-repression of selected VSGs up to 10–20-fold [52,53,54,212,266,270], depletion or conditional knockout of TbRAP1 by far results in the most severe VSG de-repressing phenotype, where silent VSGs are de-repressed up to a thousand-fold, and nearly all VSGs in the genome are affected [52,105,106,107].
Examination of the chromatin structure by Formaldehyde-Assisted Isolation of Regulatory Elements (FAIRE) [271] and micrococcal nuclease (MNase) digestion showed that TbRAP1 helps compact the telomeric and subtelomeric ES chromatin structure, although this effect is more prominent in the insect form of T. brucei that proliferates in the midgut of its insect vector than in the bloodstream form of T. brucei that proliferates in its mammalian host [103]. This observation suggests that TbRAP1-mediated telomeric silencing is an epigenetic effect that relies on modulation of the chromatin structure, which is similar to telomeric silencing in yeast and human, although the underlying mechanism is not fully understood.
Quantitative RT-PCR has been used to measure depression of several genes along the same ES in TbRAP1-depleted cells: a unique VSG gene immediately upstream of the telomere repeats, a unique VSG pseudogene in the middle of the ES, and a luciferase reporter gene inserted immediately downstream of the ES promoter [52]. As expected, the TbRAP1-mediated silencing is strongest for the VSG gene located adjacent to the telomere and weakest for the luciferase gene 40–60 kb upstream [52]. This suggests that the silencing effect is nucleated at the telomere and spreads continuously toward the chromosome internal region, similar to what has been observed in budding yeast at the VII-L telomere with inserted reporter genes [80,272]. Importantly, this observation strongly suggests that TbRAP1’s association with the telomere chromatin is essential for VSG silencing.
Different RAP1 homologs use different means to be localized at the telomere. ScRap1 uses its central Myb and Myb-like domains to bind dsDNA in a sequence-specific manner, as described above [42,43,273]. Similarly, C. albicans RAP1 also has both Myb and Myb-like domains and can bind dsDNA directly [187,274]. However, human and S. pombe RAP1s do not have any DNA binding activities and are only recruited to the telomere through their interaction with TRF2 and SpTaz1, respectively [31,275]. Sequence analysis showed that TbRAP1 has the conserved BRCT, Myb, Myb-like, and RCT domains (Figure 1), but the sequence homology is pretty weak [52]. Although TbRAP1 interacts with TbTRF [52], TbRAP1 is also located at the telomere in TbTRF-depleted cells [107], indicating that TbTRF is not required for recruiting TbRAP1 to the telomere. This is further confirmed by the observation that the TbRAP1∆Myb mutant still associates with the telomere chromatin, while TbTRF interacts with TbRAP1’s Myb domain [106,107]. This observation strongly suggests that if TbRAP1 Myb has any DNA binding activity, it is not essential for targeting TbRAP1 to the telomere.
Interestingly, TbRAP1 has an 737RKRRR741 (R/K) patch in its Myb-like domain and as part of its bipartite nuclear localization signal (NLS) [106,107]. The TbRAP1 Myb-like domain is required for its interaction with Importin α and its nuclear localization, presumably because Importin α interacts with NLS and helps import TbRAP1 into the nucleus [106]. Surprisingly, the TbRAP1 R/K patch has both dsDNA and ssDNA binding activities, which has been demonstrated by NMR titration and EMSA analyses [107]. These DNA binding activities of TbRAP1 are sequence non-specific and electrostatics based. In addition, TbRAP1 binds longer DNA substrates better than shorter ones, and the shortest dsDNA and ssDNA substrates that TbRAP1 can recognize are ~60 bp and 60 nt, respectively [107]. Importantly, this R/K patch is essential for TbRAP1’s association with the telomere chromatin, as mutating the R/K patch to five alanine residues (5A) removes TbRAP1 from the telomere, which is confirmed by both Chromatin IP and Immunofluorescence analyses [107]. Proteomic studies showed that the two serine residues next to the R/K patch, serine 742 and serine 744, can be phosphorylated in T. brucei cells at both the infectious and the insect stages [276,277]. Interestingly, phosphomimetic mutation S742DS744D (2SD) disrupts most of TbRAP1’s dsDNA binding without significantly reducing its ssDNA binding activity, while the S742AS744A (2SA) mutation does not affect TbRAP1’s DNA binding at all [107]. Presumably, phosphorylation of the serine residues brings sufficient negative charges near the R/K patch, which disrupts TbRAP1’s interaction with dsDNA. Importantly, TbRAP1-2SD is no longer localized at the telomere, while TbRAP1-2SA still associates with the telomere chromatin, indicating that the R/K patch-mediated dsDNA binding activity of TbRAP1 is required for its recruitment to the telomere [107]. Furthermore, TbRAP1’s dsDNA binding activity is essential for cell proliferation, VSG silencing, and telomere integrity: both TbRAP1 5A and 2SD mutants experience acute growth arrest when the WT complementary allele is conditionally deleted, approximately ten thousand genes are up- or down-regulated in these mutants, and these mutants have an increased amount of DNA damage in the telomeric and subtelomeric regions [107]. Apparently, TbRAP1’s dsDNA binding activity is essential, which is the same as ScRap1. However, the properties and regulation of TbRAP1 and ScRap1’s DNA binding activities are drastically different.
Interestingly, TbRAP1 and nuclear phosphatidylinositol 5-phosphatase (PIP5Pase) have been identified in the same protein complex [278,279], where PIP5Pase can dephosphorylate PI(3,4,5)P3. A C-terminally HA-tagged TbRAP1 can bind PI(3,4,5)P3 but not PI(4,5)P2, and depletion of PIP5pase also leads to a dramatic global VSG de-repressing phenotype [278,279,280]. The current model is that TbRAP1’s binding to PI(3,4,5)P3 interferes with its association with the telomere chromatin and disrupts the TbRAP1-mediated VSG silencing, while PIP5Pase-mediated dephosphorylation of PI(3,4,5)P3 ensures that TbRAP1’s telomere association is not perturbed. The ability of TbRAP1 to bind phosphoinositol metabolites is unique among RAP1 homologs, and this interaction appears to be important for regulation of TbRAP1’s DNA binding function, although the detailed mechanism is unclear. On the other hand, TbRAP1 does not seem to directly regulate the parasite metabolism, even though RNAseq analysis suggests that TbRAP1 is required for the full-level expression of multiple ribosomal protein genes [105,106,107]. In comparison, ScRap1 is a transcription regulator for ribosomal protein and glycolysis protein genes [97,127], while mouse RAP1 deficiency leads to decreased activation of PPARα and PGC1α, presumably through regulating the expression of genes encoding these enzymes [281,282].
VSG monoallelic expression has two aspects: silencing all but one VSG and expressing the active VSG at its full-expression level. It has been estimated that ~10 million VSG molecules are expressed on the T. brucei cell surface [193], masking invariant surface molecules from the host’s immune surveillance. To make this dense VSG layer, the active VSG gene is transcribed by RNA pol I at a very high level [194], producing VSG RNA that represents ~10% of total cell RNA [283,284]. Comparing RNA levels of the same VSG gene in cells where it is either active or silent indicates that the active VSG RNA level is at least 10,000-fold higher than that of a silent VSG [103]. In addition, VSG is an essential protein, as depleting VSG protein by RNAi leads to a pre-cytokinesis arrest [285], and blocking VSG synthesis triggers a general arrest in translation initiation [286]. Therefore, while TbRAP1’s dsDNA binding activity is essential for VSG silencing, it is important that TbRAP1 allows the active VSG to be expressed at its full level. It turned out that TbRAP1 has another nucleic acid binding activity essential for this function [105].
The NMR structure of the TbRAP1 Myb-like domain has been solved recently [105]. Surprisingly, the N-terminal two-thirds of the Myb-like domain fold into a typical RNA Recognition Motif (RRM) that usually binds RNA or ssDNA [287]. NMR titration and EMSA analyses showed that TbRAP1 RRM binds to RNA but not to ssDNA [105]. Interestingly, TbRAP1 RRM can bind RNA substrates containing the VSG 3′UTR but not TERRA [105]. Essentially all VSG 3′UTRs have a 9-mer and a 16-mer consensus sequence [110,288], and the 16-mer alone is sufficient for TbRAP1’s recognition [105]. TbRAP1 does not bind a 35 nt RNA oligo with a random sequence, indicating that this RNA binding activity is sequence-specific [105]. On the other hand, TbRAP1 RRM also binds a 170 nt RNA substrate without any VSG 3′UTR sequence, suggesting that it can recognize more than one RNA sequence [105]. Aligning TbRAP1 RRM with known RRM domains shows that phenylalanine 655 and phenylalanine 694 are key residues in the conserved RNP2 and RNP1 motifs, respectively, which directly interact with RNA [105]. Indeed, mutating the two phenylalanine residues to alanines, leucines, or glutamines (2FA, 2FL, and 2FQ mutants, respectively) abolishes TbRAP1’s RNA binding activity [105]. Surprisingly, the dsDNA and RNA binding activities of TbRAP1 are mutually exclusive: even though TbRAP1’s dsDNA binding and RNA binding activities are mediated by non-overlapping R/K patch and RRM, respectively, a TbRAP1-dsDNA-RNA ternary complex was not observed in EMSA when the TbRAP1 Myb-like fragment was incubated with both dsDNA and RNA substrates of a similar length [105]. This may be partially due to the fact that the R/K patch also enhances the RNA binding activity of TbRAP1, even though amino acid residues in the R/K patch do not interact directly with the RNA substrate in NMR titration analysis [105].
In vivo, RNA-CLIP analysis shows that TbRAP1 binds the active VSG RNA, and this binding also depends on the TbRAP1 RRM domain and the conserved F655 and F694 residues [105]. Unexpectedly, the 5A mutation that disrupts TbRAP1’s association with the telomere chromatin also disrupts the TbRAP1–VSG RNA interaction in vivo, even though TbRAP1 RRM alone can bind VSG 3′UTR-containing RNA substrates in EMSA and NMR titration analyses [105]. As TbRAP1 only has a moderate affinity to its RNA substrate, this observation suggests that localizing TbRAP1 in the active VSG vicinity is important for TbRAP1’s access to and interaction with a high concentration of the active VSG RNA. The TbRAP1-5A mutant does not associate with the telomere chromatin [107], so it presumably cannot gain access to the active VSG RNA. On the other hand, mutating F655 and F694 residues does not affect TbRAP1’s DNA binding activities, and all three TbRAP1 point mutants (2FA, 2FL, and 2FQ) are localized at the telomere [105], indicating that TbRAP1’s DNA binding and RNA binding activities are not intrinsically dependent on each other.
Most importantly, the TbRAP1-2FA, 2FL, and 2FQ mutants, which only disrupt TbRAP1’s binding to the active VSG RNA without affecting its association with the telomere chromatin, lead to a ~50% reduction of the active VSG RNA level [105]. In contrast, TbRAP1-5A and ∆DB mutants that lose the DNA binding activities (and do not interact with the telomere chromatin or the VSG RNA in vivo) have only a subtle effect on the active VSG RNA level (<10%). Therefore, the TbRAP1 RRM domain binds the active VSG RNA, and this interaction is essential for the full-level expression of the active VSG. However, the TbRAP1–VSG RNA interaction does not affect the RNA stability [105], suggesting that this interaction may be transient and has a signaling function. These observations suggest that the competition between TbRAP1’s dsDNA and RNA binding activities are critical for maintaining VSG monoallelic expression [105]; on one hand, TbRAP1’s DNA binding activities ensure its association with the telomere chromatin, and hence telomeric silencing, which represses subtelomeric VSG expression globally. On the other hand, TbRAP1’s RNA binding activity enables its interaction with the active VSG RNA only at the active VSG locus where nascent VSG RNA is concentrated [232], which competes for TbRAP1’s binding, prevents TbRAP1 from binding dsDNA, and presumably disrupts the TbRAP1-mediated silencing effect locally, hence permitting a full-level expression of the active VSG. This model can also explain why TbRAP1 DNA binding-defective mutants only affect the active VSG expression subtly [105]: although TbRAP1-5A and ∆DB mutants do not interact with the active VSG RNA in vivo, their dissociation from the telomere chromatin already disrupts the TbRAP1-mediated telomeric silencing in the first place, so the TbRAP1–VSG RNA interaction is no longer necessary to counteract the silencing effect.
These findings answer the question of how TbRAP1 allows a full expression of the active VSG while silencing the other VSGs simultaneously. Identification of TbRAP1’s RRM domain and RNA binding activity further brings the investigation of gene expression regulation into a new paradigm, where both dsDNA and RNA binding activities of the same protein and the competition between the two activities are essential for monoallelic gene expression.
It is worth noting that VSG monoallelic expression employs critical allelic exclusion mechanisms. As described above, VEX2 is associated with ESB and repels silent ES from gaining access to RNA Pol I [241,243]. In addition, the total amount of VSG mRNA appears to be fixed, as expressing an ectopic VSG gene leads to downregulation of the endogenous VSG expression [286,288,289,290,291]. This is possibly regulated through the 16-mer VSG 3′ UTR consensus sequence [288], which can be bound by CFB2 [292,293] and modified with m6A [294], both important for VSG mRNA stability. However, adding stop codons in the VSG gene ORF leads to dramatically increased total VSG mRNA levels, suggesting that translation of VSG mRNA is actually monitored to regulate VSG expression [295]. Regardless of the underlying mechanism, the need to keep a fixed amount of total expressed VSG can explain why TbRAP1 RNA binding-defective mutants also cause de-repression of silent VSGs at slower kinetics than TbRAP1 DNA binding-defective mutants [105], as a dramatic decrease in the active VSG expression can signal for increased expression of silent VSGs. Alternatively, it is also possible that the TbRAP1–VSG RNA interaction may directly signal for allelic exclusion of VSG expression, and loss of this interaction allows de-repression of silent VSGs.
3.4. TbRAP1 Helps Maintain Telomere Stability and Suppresses VSG Switching
As a telomere protein, TbRAP1 is also essential for maintaining telomere integrity and stability: depletion of TbRAP1 by RNAi results in more γH2A (deposited to chromatin at the DNA damage site [296]) and RAD51 associated with the telomere and subtelomere chromatin when analyzed by Chromatin IP, an increased amount of DNA breaks at the subtelomeres when analyzed by Ligation-Mediated PCR, and an elevated VSG switching rate with more VSG gene conversion events when analyzing the status of markers inserted in the active ES [104]. In addition, TbRAP1 point mutants, including 5A and 2SD that disrupt its DNA binding/telomere localization, and 2FQ, 2FL, and 2FA that disrupt its RNA binding, all lead to an increased amount of telomere- and subtelomere-associated γH2A [105,107]. Interestingly, TbRAP1 also suppresses the TERRA level [104], which is an important mechanism of how TbRAP1 maintains telomere integrity and stability.
TERRA was originally detected in T. brucei, which contains the G-rich telomere repeat sequence, and only a small portion of it is poly-adenylated [157]. In T. brucei, TERRA appears to only be transcribed from the active VSG ES-adjacent telomere by RNA pol I, presumably as a read-through product [104,270]. In the G1 phase, most T. brucei cells have a single nuclear TERRA focus. As cells progress through the cell cycle, most cells have 2 or 3 nuclear TERRA foci in S or G2/M phases [270]. Depletion of TbTRF leads to more cells with fewer number of TERRA foci throughout the cell cycle [270]. Both TbRAP1 and TbTRF suppress the TERRA level [104,270]. In addition, depletion of TbRAP1 or TbTRF also results in more telomeric R-loops (TRL), an increased amount of telomere DNA damage, and an elevated VSG switching rate [104,223,270]. Overexpression of an ectopic allele of RNase H1 that degrades RNA in the RNA:DNA hybrid [297,298] can suppress the increased TRL level, more telomeric DNA breaks, and higher VSG switching rate phenotypes in TbRAP1/TbTRF-depleted cells [104,270], confirming that suppressing TERRA and TRL levels is a major underlying mechanism of how TbRAP1 and TbTRF help maintain telomere/subtelomere stability. Therefore, TERRA expression regulation is directly linked to telomere stability through TbRAP1 and TbTRF. However, this regulation, mediated by TbRAP1 and TbTRF, is presumably not at the transcription initiation level, as TERRA is transcribed by RNA pol I downstream of the active VSG ES [104,270], with its promoter 40–60 kb upstream [111]. TbTRF binds TERRA directly using its C-terminal Myb domain [270] that also binds the duplex TTAGGG repeats [51], which may contribute to its role in TERRA suppression. TbRAP1 does not bind TERRA [105], but TERRA species are much longer in TbRAP1-depleted cells [104], suggesting that TbRAP1’s binding to the telomere or TbRAP1-mediated telomere chromatin compaction deters RNA pol I transcription elongation.
4. Discussion
In summary, it is clear that the fundamental telomere functions in different eukaryotes are conserved, as telomere proteins help protect the chromosome ends and maintain telomere integrity and stability, and genes located in subtelomeric regions are frequently silenced by the nearby telomeric chromatin. However, telomere proteins, including their identities, sequences, functional domains, and mechanisms of their action, frequently have species-specific features.
Specifically for RAP1 homologs, their roles in maintaining telomere integrity and stability are conserved (Figure 2), and RAP1 homologs have been shown to suppress HR in the telomere vicinity: TbRAP1 suppresses subtelomeric VSG gene conversion events, and the recombinase RAD51 is recruited to telomeric/subtelomeric DNA damage sites when TbRAP1 is depleted [104]. ScRap1 suppresses recruitment of the Mec1-Ddc2 checkpoint complex and the Rad52, Mre11, and RPA recombination machinery to the telomere [119,299]. Mammalian RAP1 also coordinates with TRF2 to suppress HR-mediated telomere deletion and fusions [189,190], even though they are not essential [281,300,301]. ScRap1 also suppresses NHEJ at the telomere [117,118]. Mammalian RAP1 has been reported to suppress NHEJ in vitro [184,185] and suppress telomere fusions in cells with short telomeres [186]. T. brucei does not have the NHEJ machinery [207], and no telomere end-to-end fusions have been reported in TbRAP1-depleted cells. In addition, T. brucei, yeast, and mammalian RAP1s all suppress TERRA [104,156,160,174], which in turn helps maintain telomere stability. This also links RAP1’s telomeric silencing effect with its chromosome end protection function. However, ScRap1 presumably suppresses TERRA at the transcription initiation step, while TbRAP1 apparently suppresses RNA pol I transcription elongation, even though both may involve chromatin structure remodeling.
RAP1 homologs’ functions in gene transcription regulation appear more divergent (Figure 3). ScRap1’s transcription activation function depends on its binding to the ribosomal protein gene promoter [97], and ScRap1 also represses cryptic or divergent transcription to ensure the transcription goes in the correct direction [302,303]. On the other hand, ScRap1-mediated telomeric silencing depends on its recruitment of Sir proteins to the telomere and a heterochromatic telomere structure [41]. Mouse RAP1 also binds multiple chromosomal internal sites and regulates nearby gene expression [281]. In addition, mouse RAP1 exhibits a repressive effect on the expression of genes located at subtelomeres [281]. Furthermore, cytoplasmic RAP1 interacts with IκB [304], the inhibitory protein partner of NFκB [305]. The RAP1–IκB interaction releases NFκB, which permits NFκB to be activated as a transcription activator [304]. Hence, mammalian RAP1 has an additional extra-telomere gene expression regulation function.
TbRAP1 is clearly required for silencing subtelomeric VSG genes, which appears to involve chromatin remodeling (Figure 3, top) [103]. However, ScRap1-mediated silencing affects RNA pol II access to the subtelomeric gene promoter, while TbRAP1 likely blocks RNA pol I transcription elongation, presumably as a structure protein intrinsic to the telomere chromatin. In addition, RNAseq analysis showed that conditional knockout of TbRAP1 also caused a ~35% decrease in the RNA level of a number of ribosomal protein genes [106,107], although it has not been reported whether these genes are within the same polycistronic transcription units and whether TbRAP1 directly binds their promotor. Interestingly, TbRAP1 also helps ensure full-level expression of the active VSG, which is not due to any canonical transcription activation function (Figure 3, bottom) [105]. Rather, through competing with TbRAP1’s dsDNA binding activity, its RNA binding activity inhibits its silencing effect [105]. Both the RRM domain and RNA binding activity are unique in kinetoplastid RAP1s but absent in yeast or mammalian RAP1 homologs [105]. A recent finding indicates that many human transcription factors have not only DNA binding but also arginine-rich motif (ARM)-mediated RNA binding activities, which help fine-tune transcription regulation [306]. Therefore, many more proteins than we previously knew seem to have both DNA and RNA binding activities that play important roles in gene expression regulation. It would be intriguing to see whether yeast and mammalian RAP1 homologs have similar ARM domains and RNA binding activities.
Identification and characterization of functions of T. brucei telomere proteins revealed that the essential functions of telomere proteins are largely conserved, even though homologs from kinetoplastids to mammals only have limited sequence homology within functional domains, which shed light on telomere protein evolution. Importantly, unique features of TbRAP1 DNA and RNA binding activities and their regulations have been identified that are absent from mammalian RAP1s. Therefore, TbRAP1, an essential telomere protein, is also a promising target for future development of anti-parasite agents.
Funding
This work is supported by NIH grants (R01AI066095, Li and R01AI179972, Li).
Conflicts of Interest
The author declares no conflicts of interest.
References
- Shay, J.W.; Wright, W.E. Telomeres and telomerase: Three decades of progress. Nat. Rev. Genet. 2019, 20, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Lyčka, M.; Bubeník, M.; Závodník, M.; Peska, V.; Fajkus, P.; Demko, M.; Fajkus, J.; Fojtová, M. TeloBase: A community-curated database of telomere sequences across the tree of life. Nucleic Acids Res. 2023, gkad672. [Google Scholar] [CrossRef] [PubMed]
- Podlevsky, J.D.; Bley, C.J.; Omana, R.V.; Qi, X.; Chen, J.J. The telomerase database. Nucleic Acids Res. 2008, 36, D339–D343. [Google Scholar] [CrossRef] [PubMed]
- Bonnell, E.; Pasquier, E.; Wellinger, R.J. Telomere Replication: Solving Multiple End Replication Problems. Front. Cell Dev. Biol. 2021, 9, 668171. [Google Scholar] [CrossRef]
- Wellinger, R.J. In the end, what’s the problem. Mol. Cell 2014, 53, 855–856. [Google Scholar] [CrossRef]
- Munoz-Jordan, J.L.; Cross, G.A.M.; de Lange, T.; Griffith, J.D. t-loops at trypanosome telomeres. EMBO J. 2001, 20, 579–588. [Google Scholar] [CrossRef]
- Griffith, J.D.; Comeau, L.; Rosenfield, S.; Stansel, R.M.; Bianchi, A.; Moss, H.; de Lange, T. Mammalian telomeres end in a large duplex loop. Cell 1999, 97, 503–514. [Google Scholar] [CrossRef]
- Murti, K.G.; Prescott, D.M. Telomeres of polytene chromosomes in a ciliated protozoan terminate in duplex DNA loops. Proc. Natl. Acad. Sci. USA 1999, 96, 14436–14439. [Google Scholar] [CrossRef]
- Nikitina, T.; Woodcock, C.L. Closed chromatin loops at the ends of chromosomes. J. Cell Biol. 2004, 166, 161–165. [Google Scholar] [CrossRef]
- Lu, X.; Liu, L. Genome stability from the perspective of telomere length. Trends Genet. 2023. online ahead of print. [Google Scholar] [CrossRef]
- de Lange, T. How telomeres solve the end-protection problem. Science 2009, 326, 948–952. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, R.J.; Karlseder, J. Telomeres: Protecting chromosomes against genome instability. Nat. Rev. Mol. Cell Biol. 2010, 11, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Ruis, P.; Boulton, S.J. The end protection problem—An unexpected twist in the tail. Genes Dev. 2021, 35, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Casari, E.; Gnugnoli, M.; Rinaldi, C.; Pizzul, P.; Colombo, C.V.; Bonetti, D.; Longhese, M.P. To Fix or Not to Fix: Maintenance of Chromosome Ends Versus Repair of DNA Double-Strand Breaks. Cells 2022, 11, 3224. [Google Scholar] [CrossRef] [PubMed]
- de Lange, T. Shelterin-Mediated Telomere Protection. Annu. Rev. Genet. 2018, 52, 223–247. [Google Scholar] [CrossRef] [PubMed]
- Van Ly, D.; Low, R.R.J.; Frölich, S.; Bartolec, T.K.; Kafer, G.R.; Pickett, H.A.; Gaus, K.; Cesare, A.J. Telomere Loop Dynamics in Chromosome End Protection. Mol. Cell 2018, 71, 510–525.e6. [Google Scholar] [CrossRef] [PubMed]
- de Lange, T. Shelterin: The protein complex that shapes and safeguards human telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef]
- Stewart, J.A.; Chaiken, M.F.; Wang, F.; Price, C.M. Maintaining the end: Roles of telomere proteins in end-protection, telomere replication and length regulation. Mutat. Res. 2012, 730, 12–19. [Google Scholar] [CrossRef]
- Li, B.; Zhao, Y. Regulation of antigenic variation by Trypanosoma brucei telomere proteins depends on their unique DNA binding activities. Pathogens 2021, 10, 967. [Google Scholar] [CrossRef]
- Li, B. Telomere maintenance in African trypanosomes. Front. Mol. Biosci. 2023, 10, 1302557. [Google Scholar] [CrossRef]
- Lim, C.J.; Cech, T.R. Shaping human telomeres: From shelterin and CST complexes to telomeric chromatin organization. Nat. Rev. Mol. Cell Biol. 2021, 22, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Chong, L.; van Steensel, B.; Broccoli, D.; Erdjument-Bromage, H.; Hanish, J.; Tempst, P.; de Lange, T. A human telomeric protein. Science 1995, 270, 1663–1667. [Google Scholar] [CrossRef] [PubMed]
- Broccoli, D.; Smogorzewska, A.; Chong, L.; de Lange, T. Human telomeres contain two distinct Myb-related proteins, TRF1 and TRF2. Nat. Genet. 1997, 17, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Bilaud, T.; Brun, C.; Ancelin, K.; Koering, C.E.; Laroche, T.; Gilson, E. Telomeric localization of TRF2, a novel human telobox protein. Nat. Genet. 1997, 17, 236–239. [Google Scholar] [CrossRef] [PubMed]
- Houghtaling, B.R.; Cuttonaro, L.; Chang, W.; Smith, S. A Dynamic Molecular Link between the Telomere Length Regulator TRF1 and the Chromosome End Protector TRF2. Curr. Biol. 2004, 14, 1621–1631. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Safari, A.; O’Connor, M.S.; Chan, D.W.; Laegeler, A.; Qin, J.; Songyang, Z. PTOP interacts with POT1 and regulates its localization to telomeres. Nat. Cell Biol. 2004, 6, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.Z.; Hockemeyer, D.; Krutchinsky, A.N.; Loayza, D.; Hooper, S.M.; Chait, B.T.; de Lange, T. POT1-interacting protein PIP1: A telomere length regulator that recruits POT1 to the TIN2/TRF1 complex. Genes Dev. 2004, 18, 1649–1654. [Google Scholar] [CrossRef]
- Baumann, P.; Podell, E.; Cech, T.R. Human pot1 (protection of telomeres) protein: Cytolocalization, gene structure, and alternative splicing. Mol. Cell. Biol. 2002, 22, 8079–8087. [Google Scholar] [CrossRef]
- Baumann, P.; Cech, T.R. Pot1, the putative telomere end-binding protein in fission yeast and humans. Science 2001, 292, 1171–1175. [Google Scholar] [CrossRef]
- Loayza, D.; de Lange, T. POT1 as a terminal transducer of TRF1 telomere length control. Nature 2003, 424, 1013–1018. [Google Scholar] [CrossRef]
- Li, B.; Oestreich, S.; de Lange, T. Identification of human Rap1: Implications for telomere evolution. Cell 2000, 101, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kaminker, P.; Campisi, J. TIN2, a new regulator of telomere length in human cells. Nat. Genet. 1999, 23, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.Z.; Donigian, J.R.; van Overbeek, M.; Loayza, D.; Luo, Y.; Krutchinsky, A.N.; Chait, B.T.; de Lange, T. TIN2 binds TRF1 and TRF2 simultaneously and stabilizes the TRF2 complex on telomeres. J. Biol. Chem. 2004, 279, 47264–47271. [Google Scholar] [CrossRef] [PubMed]
- Wellinger, R.J. The CST complex and telomere maintenance: The exception becomes the rule. Mol. Cell 2009, 36, 168–169. [Google Scholar] [CrossRef] [PubMed]
- Rice, C.; Skordalakes, E. Structure and function of the telomeric CST complex. Comput. Struct. Biotechnol. J. 2016, 14, 161–167. [Google Scholar] [CrossRef]
- Lim, C.J.; Barbour, A.T.; Zaug, A.J.; Goodrich, K.J.; McKay, A.E.; Wuttke, D.S.; Cech, T.R. The structure of human CST reveals a decameric assembly bound to telomeric DNA. Science 2020, 368, 1081–1085. [Google Scholar] [CrossRef]
- Miyake, Y.; Nakamura, M.; Nabetani, A.; Shimamura, S.; Tamura, M.; Yonehara, S.; Saito, M.; Ishikawa, F. RPA-like mammalian Ctc1-Stn1-Ten1 complex binds to single-stranded DNA and protects telomeres independently of the Pot1 pathway. Mol. Cell 2009, 36, 193–206. [Google Scholar] [CrossRef]
- Olson, C.L.; Barbour, A.T.; Wieser, T.A.; Wuttke, D.S. RPA engages telomeric G-quadruplexes more effectively than CST. Nucleic Acids Res. 2023, 51, 5073–5086. [Google Scholar] [CrossRef]
- Barbour, A.T.; Wuttke, D.S. RPA-like single-stranded DNA-binding protein complexes including CST serve as specialized processivity factors for polymerases. Curr. Opin. Struct. Biol. 2023, 81, 102611. [Google Scholar] [CrossRef]
- Wang, S.S.; Zakian, V.A. Sequencing of Saccharomyces telomeres cloned using T4 DNA polymerase reveals two domains. Mol. Cell Biol. 1990, 10, 4415–4419. [Google Scholar]
- Wellinger, R.J.; Zakian, V.A. Everything you ever wanted to know about Saccharomyces cerevisiae telomeres: Beginning to end. Genetics 2012, 191, 1073–1105. [Google Scholar] [CrossRef] [PubMed]
- Konig, P.; Rhodes, D. Recognition of telomeric DNA. Trends Biochem. Sci. 1997, 22, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Konig, P.; Giraldo, R.; Chapman, L.; Rhodes, D. The crystal structure of the DNA-binding domain of yeast RAP1 in complex with telomeric DNA. Cell 1996, 85, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Wahlin, J.; Cohn, M. Saccharomyces cerevisiae RAP1 binds to telomeric sequences with spatial flexibility. Nucleic Acids Res. 2000, 28, 2292–2301. [Google Scholar] [CrossRef] [PubMed]
- Ribaud, V.; Ribeyre, C.; Damay, P.; Shore, D. DNA-end capping by the budding yeast transcription factor and subtelomeric binding protein Tbf1. EMBO J. 2012, 31, 138–149. [Google Scholar] [CrossRef]
- Bourns, B.D.; Alexander, M.K.; Smith, A.M.; Zakian, V.A. Sir proteins, Rif proteins, and Cdc13p bind Saccharomyces telomeres in vivo. Mol. Cell. Biol. 1998, 18, 5600–5608. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.R.; Weilbaecher, R.G.; Walterscheid, M.; Lundblad, V. Identification of the single-strand telomeric DNA binding domain of the Saccharomyces cerevisiae Cdc13 protein. Proc. Natl. Acad. Sci. USA 2000, 97, 6457–6462. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Wu, Z.; Chen, H.; Zhong, Q.; Shi, S.; Li, G.; Wu, J.; Lei, M. Structural insights into telomere protection and homeostasis regulation by yeast CST complex. Nat. Struct. Mol. Biol. 2020, 27, 752–762. [Google Scholar] [CrossRef]
- Blackburn, E.H.; Challoner, P.B. Identification of a telomeric DNA sequence in Trypanosoma brucei. Cell 1984, 36, 447–457. [Google Scholar] [CrossRef]
- Meyne, J.; Ratliff, R.L.; Moyzis, R.K. Conservation of the human telomere sequence (TTAGGG)n among vertebrates. Proc. Natl. Acad. Sci. USA 1989, 86, 7049–7053. [Google Scholar] [CrossRef]
- Li, B.; Espinal, A.; Cross, G.A.M. Trypanosome telomeres are protected by a homologue of mammalian TRF2. Mol. Cell. Biol. 2005, 25, 5011–5021. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Figueiredo, L.M.; Espinal, A.; Okubo, E.; Li, B. RAP1 is essential for silencing telomeric variant surface glycoprotein genes in Trypanosoma brucei. Cell 2009, 137, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Jehi, S.E.; Wu, F.; Li, B. Trypanosoma brucei TIF2 suppresses VSG switching by maintaining subtelomere integrity. Cell Res. 2014, 24, 870–885. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, M.A.G.; Tonini, M.L.; Afrin, M.; Li, B. POLIE suppresses telomerase-mediated telomere G-strand extension and helps ensure proper telomere C-strand synthesis in trypanosomes. Nucleic Acids Res. 2022, 50, 2036–2050. [Google Scholar] [CrossRef] [PubMed]
- Reis, H.; Schwebs, M.; Dietz, S.; Janzen, C.J.; Butter, F. TelAP1 links telomere complexes with developmental expression site silencing in African trypanosomes. Nucleic Acids Res. 2018, 46, 2820–2833. [Google Scholar] [CrossRef] [PubMed]
- Olovnikov, A.M. A theory of marginotomy. The incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenon. J. Theor. Biol. 1973, 41, 181–190. [Google Scholar] [CrossRef]
- Ferreira, M.G.; Miller, K.M.; Cooper, J.P. Indecent exposure: When telomeres become uncapped. Mol. Cell 2004, 13, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Pizzul, P.; Rinaldi, C.; Bonetti, D. The multistep path to replicative senescence onset: Zooming on triggering and inhibitory events at telomeric DNA. Front. Cell Dev. Biol. 2023, 11, 1250264. [Google Scholar] [CrossRef]
- Lee, J.W.; Ong, E.B.B. Genomic Instability and Cellular Senescence: Lessons from the Budding Yeast. Front. Cell Dev. Biol. 2020, 8, 619126. [Google Scholar] [CrossRef]
- Liu, J.; Wang, L.; Wang, Z.; Liu, J.P. Roles of Telomere Biology in Cell Senescence, Replicative and Chronological Ageing. Cells 2019, 8, 54. [Google Scholar] [CrossRef]
- Greider, C.W.; Blackburn, E.H. A telomeric sequence in the RNA of Tetrahymena telomerase required for telomere repeat synthesis. Nature 1989, 337, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Greider, C.W.; Blackburn, E.H. The telomere terminal transferase of Tetrahymena is a ribonucleoprotein enzyme with two kinds of primer specificity. Cell 1987, 51, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Greider, C.W.; Blackburn, E.H. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell 1985, 43, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, G.E.; Fountain, A.J.; van Roon, A.M.; Rangan, R.; Das, R.; Collins, K.; Nguyen, T.H.D. Structure of human telomerase holoenzyme with bound telomeric DNA. Nature 2021, 593, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; He, Y.; Wang, Y.; Song, H.; Zhou, Z.H.; Feigon, J. Structure of active human telomerase with telomere shelterin protein TPP1. Nature 2022, 604, 578–583. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Feigon, J. Telomerase structural biology comes of age. Curr. Opin. Struct. Biol. 2022, 76, 102446. [Google Scholar] [CrossRef] [PubMed]
- Mason, J.M.; Frydrychova, R.C.; Biessmann, H. Drosophila telomeres: An exception providing new insights. Bioessays 2008, 30, 25–37. [Google Scholar] [CrossRef]
- Pardue, M.L.; DeBaryshe, P.G. Drosophila telomeres: A variation on the telomerase theme. Fly 2008, 2, 101–110. [Google Scholar] [CrossRef]
- Zhang, J.M.; Zou, L. Alternative lengthening of telomeres: From molecular mechanisms to therapeutic outlooks. Cell Biosci. 2020, 10, 30. [Google Scholar] [CrossRef]
- Hou, K.; Yu, Y.; Li, D.; Zhang, Y.; Zhang, K.; Tong, J.; Yang, K.; Jia, S. Alternative Lengthening of Telomeres and Mediated Telomere Synthesis. Cancers 2022, 14, 2194. [Google Scholar] [CrossRef]
- Chen, Q.; Ijpma, A.; Greider, C.W. Two survivor pathways that allow growth in the absence of telomerase are generated by distinct telomere recombination events. Mol. Cell. Biol. 2001, 21, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Lue, N.F.; Yu, E.Y. Telomere recombination pathways: Tales of several unhappy marriages. Curr. Genet. 2017, 63, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Li, B. Keeping balance between genetic stability and plasticity at the telomere and subtelomere of Trypanosoma brucei. Front. Cell Dev. Biol. 2021, 9, 699639. [Google Scholar] [CrossRef] [PubMed]
- Li, B. DNA double-strand breaks and telomeres play important roles in Trypanosoma brucei antigenic variation. Eukaryot. Cell 2015, 14, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Sima, N.; McLaughlin, E.J.; Hutchinson, S.; Glover, L. Escaping the immune system by DNA repair and recombination in African trypanosomes. Open Biol. 2019, 9, 190182. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, R.; Morrison, L.J.; Hall, J.P.J. DNA Recombination Strategies during Antigenic Variation in the African Trypanosome. Microbiol. Spectr. 2015, 3, MDNA3-0016. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, A.; Gilson, E.; Magdinier, F. Telomeric position effect: From the yeast paradigm to human pathologies? Biochimie 2008, 90, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Kim, D.Y.; Kim, W. Regulation of Gene Expression by Telomere Position Effect. Int. J. Mol. Sci. 2021, 22, 12807. [Google Scholar] [CrossRef]
- Elgin, S.C.; Reuter, G. Position-effect variegation; heterochromatin formation, and gene silencing in Drosophila. Cold Spring Harb. Perspect. Biol. 2013, 5, a017780. [Google Scholar] [CrossRef]
- Gottschling, D.E.; Aparicio, O.M.; Billington, B.L.; Zakian, V.A. Position effect at S. cerevisiae telomeres: Reversible repression of pol II transcription. Cell 1990, 63, 751–762. [Google Scholar] [CrossRef]
- Mason, J.M.; Konev, A.Y.; Golubovsky, M.D.; Biessmann, H. Cis- and trans-acting influences on telomeric position effect in Drosophila melanogaster detected with a subterminal transgene. Genetics 2003, 163, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Tennen, R.I.; Bua, D.J.; Wright, W.E.; Chua, K.F. SIRT6 is required for maintenance of telomere position effect in human cells. Nat. Commun. 2011, 2, 433. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Zou, Y.; Shay, J.W.; Wright, W.E. Telomere position effect in human cells. Science 2001, 292, 2075–2077. [Google Scholar] [CrossRef]
- Koering, C.E.; Pollice, A.; Zibella, M.P.; Bauwens, S.; Puisieux, A.; Brunori, M.; Brun, C.; Martins, L.; Sabatier, L.; Pulitzer, J.F.; et al. Human telomeric position effect is determined by chromosomal context and telomeric chromatin integrity. EMBO Rep. 2002, 3, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Robin, J.D.; Ludlow, A.T.; Batten, K.; Magdinier, F.; Stadler, G.; Wagner, K.R.; Shay, J.W.; Wright, W.E. Telomere position effect: Regulation of gene expression with progressive telomere shortening over long distances. Genes Dev. 2014, 28, 2464–2476. [Google Scholar] [CrossRef]
- Stadler, G.; Rahimov, F.; King, O.D.; Chen, J.C.; Robin, J.D.; Wagner, K.R.; Shay, J.W.; Emerson, C.P.; Wright, W.E. Telomere position effect regulates DUX4 in human facioscapulohumeral muscular dystrophy. Nat. Struct. Mol. Biol. 2013, 20, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Ludlow, A.T.; Min, J.; Robin, J.D.; Stadler, G.; Mender, I.; Lai, T.P.; Zhang, N.; Wright, W.E.; Shay, J.W. Regulation of the Human Telomerase Gene TERT by Telomere Position Effect-Over Long Distances (TPE-OLD): Implications for Aging and Cancer. PLoS Biol. 2016, 14, e2000016. [Google Scholar] [CrossRef]
- Glover, L.; Alsford, S.; Beattie, C.; Horn, D. Deletion of a trypanosome telomere leads to loss of silencing and progressive loss of terminal DNA in the absence of cell cycle arrest. Nuc. Acids Res. 2007, 35, 872–880. [Google Scholar] [CrossRef]
- Alsford, S.; Kawahara, T.; Isamah, C.; Horn, D. A sirtuin in the African trypanosome is involved in both DNA repair and telomeric gene silencing but is not required for antigenic variation. Mol. Microbiol. 2007, 63, 724–736. [Google Scholar] [CrossRef]
- Glover, L.; Horn, D. Repression of polymerase I-mediated gene expression at Trypanosoma brucei telomeres. EMBO Rep. 2006, 7, 93–99. [Google Scholar] [CrossRef]
- Horn, D.; Cross, G.A.M. Position-dependent and promoter-specific regulation of gene expression in Trypanosoma brucei. EMBO J. 1997, 16, 7422–7431. [Google Scholar] [CrossRef] [PubMed]
- Freitas-Junior, L.H.; Hernandez-Rivas, R.; Ralph, S.A.; Montiel-Condado, D.; Ruvalcaba-Salazar, O.K.; Rojas-Meza, A.P.; Mancio-Silva, L.; Leal-Silvestre, R.J.; Gontijo, A.M.; Shorte, S.; et al. Telomeric heterochromatin propagation and histone acetylation control mutually exclusive expression of antigenic variation genes in malaria parasites. Cell 2005, 121, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Duraisingh, M.T.; Voss, T.S.; Marty, A.J.; Duffy, M.F.; Good, R.T.; Thompson, J.K.; Freitas-Junior, L.H.; Scherf, A.; Crabb, B.S.; Cowman, A.F. Heterochromatin silencing and locus repositioning linked to regulation of virulence genes in Plasmodium falciparum. Cell 2005, 121, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Green, B.; Benoit, N.; Sobel, J.D.; Schatz, M.C.; Wheelan, S.; Cormack, B.P. Cell wall protein variation, break-induced replication, and subtelomere dynamics in Candida glabrata. Mol. Microbiol. 2021, 116, 260–276. [Google Scholar] [CrossRef]
- López-Fuentes, E.; Gutiérrez-Escobedo, G.; Timmermans, B.; Van Dijck, P.; De Las Peñas, A.; Castaño, I. Candida glabrata’s Genome Plasticity Confers a Unique Pattern of Expressed Cell Wall Proteins. J. Fungi 2018, 4, 67. [Google Scholar] [CrossRef]
- De Las Penas, A.; Juarez-Cepeda, J.; Lopez-Fuentes, E.; Briones-Martin-Del-Campo, M.; Gutierrez-Escobedo, G.; Castano, I. Local and regional chromatin silencing in Candida glabrata: Consequences for adhesion and the response to stress. FEMS Yeast Res. 2015, 15, fov056. [Google Scholar] [CrossRef]
- Shore, D.; Nasmyth, K. Purification and cloning of a DNA binding protein from yeast that binds to both silencer and activator elements. Cell 1987, 51, 721–732. [Google Scholar] [CrossRef]
- Lustig, A.J.; Kurtz, S.; Shore, D. Involvement of the silencer and UAS binding protein RAP1 in regulation of telomere length. Science 1990, 250, 549–553. [Google Scholar] [CrossRef]
- Chikashige, Y.; Hiraoka, Y. Telomere binding of the Rap1 protein is required for meiosis in fission yeast. Curr. Biol. 2001, 11, 1618–1623. [Google Scholar] [CrossRef]
- Kanoh, J.; Ishikawa, F. spRap1 and spRif1, recruited to telomeres by Taz1, are essential for telomere function in fission yeast. Curr. Biol. 2001, 11, 1624–1630. [Google Scholar] [CrossRef]
- Cai, Y.; Kandula, V.; Kosuru, R.; Ye, X.; Irwin, M.G.; Xia, Z. Decoding telomere protein Rap1: Its telomeric and nontelomeric functions and potential implications in diabetic cardiomyopathy. Cell Cycle 2017, 16, 1765–1773. [Google Scholar] [CrossRef] [PubMed]
- Khattar, E.; Tergaonkar, V. Role of Rap1 in DNA damage response: Implications in stem cell homeostasis and cancer. Exp. Hematol. 2020, 90, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Pandya, U.M.; Sandhu, R.; Li, B. Silencing subtelomeric VSGs by Trypanosoma brucei RAP1 at the insect stage involves chromatin structure changes. Nucleic Acids Res. 2013, 41, 7673–7682. [Google Scholar] [CrossRef] [PubMed]
- Nanavaty, V.; Sandhu, R.; Jehi, S.E.; Pandya, U.M.; Li, B. Trypanosoma brucei RAP1 maintains telomere and subtelomere integrity by suppressing TERRA and telomeric RNA:DNA hybrids. Nucleic Acids Res. 2017, 45, 5785–5796. [Google Scholar] [CrossRef] [PubMed]
- Gaurav, A.K.; Afrin, M.; Yang, X.; Saha, A.; Sayeed, S.K.A.; Pan, X.; Ji, Z.; Wong, K.B.; Zhang, M.; Zhao, Y.; et al. The RRM-mediated RNA binding activity in T. brucei RAP1 is essential for VSG monoallelic expression. Nat. Commun. 2023, 14, 1576. [Google Scholar] [CrossRef] [PubMed]
- Afrin, M.; Kishmiri, H.; Sandhu, R.; Rabbani, M.A.G.; Li, B. Trypanosoma brucei RAP1 has essential functional domains that are required for different protein interactions. mSphere 2020, 5, e00027-20. [Google Scholar] [CrossRef] [PubMed]
- Afrin, M.; Gaurav, A.K.; Yang, X.; Pan, X.; Zhao, Y.; Li, B. TbRAP1 has an unusual duplex DNA binding activity required for its telomere localization and VSG silencing. Sci. Adv. 2020, 6, eabc4065. [Google Scholar] [CrossRef]
- Barry, J.D.; McCulloch, R. Antigenic variation in trypanosomes: Enhanced phenotypic variation in a eukaryotic parasite. Adv. Parasitol. 2001, 49, 1–70. [Google Scholar]
- de Lange, T.; Borst, P. Genomic environment of the expression-linked extra copies of genes for surface antigens of Trypanosoma brucei resembles the end of a chromosome. Nature 1982, 299, 451–453. [Google Scholar] [CrossRef]
- Cross, G.A.M.; Kim, H.S.; Wickstead, B. Capturing the variant surface glycoprotein repertoire (the VSGnome) of Trypanosoma brucei Lister 427. Mol. Biochem. Parasitol. 2014, 195, 59–73. [Google Scholar] [CrossRef]
- Hertz-Fowler, C.; Figueiredo, L.M.; Quail, M.A.; Becker, M.; Jackson, A.; Bason, N.; Brooks, K.; Churcher, C.; Fahkro, S.; Goodhead, I.; et al. Telomeric expression sites are highly conserved in Trypanosoma brucei. PLoS ONE 2008, 3, e3527. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.S.M.; Cosentino, R.O.; Förstner, K.U.; Guizetti, J.; Wedel, C.; Kaplan, N.; Janzen, C.J.; Arampatzi, P.; Vogel, J.; Steinbiss, S.; et al. Genome organization and DNA accessibility control antigenic variation in trypanosomes. Nature 2018, 563, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Gerloff, D.L.; Woods, N.T.; Farago, A.A.; Monteiro, A.N. BRCT domains: A little more than kin, and less than kind. FEBS Lett. 2012, 586, 2711–2716. [Google Scholar] [CrossRef] [PubMed]
- Peña-Guerrero, J.; Fernández-Rubio, C.; García-Sosa, A.T.; Nguewa, P.A.; Domains, B.R.T. Functions, and Implications in Disease-New Therapeutic Targets for Innovative Drug Discovery against Infections. Pharmaceutics 2023, 15, 1839. [Google Scholar] [CrossRef] [PubMed]
- Prouse, M.B.; Campbell, M.M. The interaction between MYB proteins and their target DNA binding sites. Biochim. Biophys. Acta 2012, 1819, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Luo, Q.; Li, Y.; Yin, L.; Zhou, N.; Li, X.; Gan, J.; Dong, A. Structural insights into target DNA recognition by R2R3-MYB transcription factors. Nucleic Acids Res. 2020, 48, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Pardo, B.; Marcand, S. Rap1 prevents telomere fusions by nonhomologous end joining. EMBO J. 2005, 24, 3117–3127. [Google Scholar] [CrossRef]
- Marcand, S.; Pardo, B.; Gratias, A.; Cahun, S.; Callebaut, I. Multiple pathways inhibit NHEJ at telomeres. Genes Dev. 2008, 22, 1153–1158. [Google Scholar] [CrossRef]
- Negrini, S.; Ribaud, V.; Bianchi, A.; Shore, D. DNA breaks are masked by multiple Rap1 binding in yeast: Implications for telomere capping and telomerase regulation. Genes Dev. 2007, 21, 292–302. [Google Scholar] [CrossRef]
- Kyrion, G.; Boakye, K.A.; Lustig, A.J. C-terminal truncation of RAP1 results in the deregulation of telomere size, stability, and function in Saccharomyces cerevisiae. Mol. Cell. Biol. 1992, 12, 5159–5173. [Google Scholar]
- Kyrion, G.; Liu, K.; Liu, C.; Lustig, A.J. RAP1 and telomere structure regulate telomere position effects in Saccharomyces cerevisiae. Genes Dev. 1993, 7, 1146–1159. [Google Scholar] [CrossRef] [PubMed]
- Shore, D. RAP1: A protean regulator in yeast. Trends Genet. 1994, 10, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Vignais, M.L.; Huet, J.; Buhler, J.M.; Sentenac, A. Contacts between the factor TUF and RPG sequences. J. Biol. Chem. 1990, 265, 14669–14674. [Google Scholar] [CrossRef] [PubMed]
- Idrissi, F.Z.; Pina, B. Functional divergence between the half-sites of the DNA-binding sequence for the yeast transcriptional regulator Rap1p. Biochem. J. 1999, 341, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Idrissi, F.Z.; Fernández-Larrea, J.B.; Piña, B. Structural and functional heterogeneity of Rap1p complexes with telomeric and UASrpg-like DNA sequences. J. Mol. Biol. 1998, 284, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Matot, B.; Le Bihan, Y.V.; Lescasse, R.; Pérez, J.; Miron, S.; David, G.; Castaing, B.; Weber, P.; Raynal, B.; Zinn-Justin, S.; et al. The orientation of the C-terminal domain of the Saccharomyces cerevisiae Rap1 protein is determined by its binding to DNA. Nucleic Acids Res. 2012, 40, 3197–3207. [Google Scholar] [CrossRef] [PubMed]
- Kalra, S.; Peyser, R.; Ho, J.; Babbin, C.; Bohan, N.; Cortes, A.; Erley, J.; Fatima, M.; Flinn, J.; Horwitz, E.; et al. Genome-wide gene expression responses to experimental manipulation of Saccharomyces cerevisiae repressor activator protein 1 (Rap1) expression level. Genomics 2023, 115, 110625. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Lustig, A.J. Genetic analysis of Rap1p/Sir3p interactions in telomeric and HML silencing in Saccharomyces cerevisiae. Genetics 1996, 143, 81–93. [Google Scholar] [CrossRef]
- Cockell, M.; Palladino, F.; Laroche, T.; Kyrion, G.; Liu, C.; Lustig, A.J.; Gasser, S.M. The carboxy termini of Sir4 and Rap1 affect Sir3 localization: Evidence for a multicomponent complex required for yeast telomeric silencing. J. Cell Biol. 1995, 129, 909–924. [Google Scholar] [CrossRef]
- Liu, C.; Mao, X.; Lustig, A.J. Mutational analysis defines a C-terminal tail domain of RAP1 essential for telomeric silencing in Saccharomyces cerevisiae. Genetics 1994, 138, 1025–1040. [Google Scholar] [CrossRef]
- Hecht, A.; Strahl-Bolsinger, S.; Grunstein, M. Spreading of transcriptional repressor SIR3 from telomeric heterochromatin. Nature 1996, 383, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Moretti, P.; Freeman, K.; Coodly, L.; Shore, D. Evidence that a complex of SIR proteins interacts with the silencer and telomere-binding protein RAP1. Genes Dev. 1994, 8, 2257–2269. [Google Scholar] [CrossRef] [PubMed]
- Moazed, D.; Kistler, A.; Axelrod, A.; Rine, J.; Johnson, A.D. Silent information regulator protein complexes in Saccharomyces cerevisiae—A sir2/sir4 complex and evidence for a regulatory domain in sir4 that inhibits its interaction with sir3. Proc. Natl. Acad. Sci. USA 1997, 94, 2186–2191. [Google Scholar] [CrossRef] [PubMed]
- Moretti, P.; Shore, D. Multiple interactions in sir protein recruitment by Rap1p at silencers and telomeres in yeast. Mol. Cell. Biol. 2001, 21, 8082–8094. [Google Scholar] [CrossRef]
- Luo, K.; Vega-Palas, M.A.; Grunstein, M. Rap1-Sir4 binding independent of other Sir, yKu, or histone interactions initiates the assembly of telomeric heterochromatin in yeast. Genes Dev. 2002, 16, 1528–1539. [Google Scholar] [CrossRef]
- Feeser, E.A.; Wolberger, C. Structural and functional studies of the Rap1 C-terminus reveal novel separation-of-function mutants. J. Mol. Biol. 2008, 380, 520–531. [Google Scholar] [CrossRef]
- Chen, Y.; Rai, R.; Zhou, Z.R.; Kanoh, J.; Ribeyre, C.; Yang, Y.; Zheng, H.; Damay, P.; Wang, F.; Tsujii, H.; et al. A conserved motif within RAP1 has diversified roles in telomere protection and regulation in different organisms. Nat. Struct. Mol. Biol. 2011, 18, 213–221. [Google Scholar] [CrossRef]
- Buck, S.W.; Shore, D. Action of a RAP1 carboxy-terminal silencing domain reveals an underlying competition between HMR and telomeres in yeast. Genes Dev. 1995, 9, 370–384. [Google Scholar] [CrossRef]
- Straight, A.F.; Shou, W.Y.; Dowd, G.J.; Turck, C.W.; Deshaies, R.J.; Johnson, A.D.; Moazed, D. Net1, a Sir2-associated nucleolar protein required for rDNA silencing and nucleolar integrity. Cell 1999, 97, 245–256. [Google Scholar] [CrossRef]
- Tanny, J.C.; Kirkpatrick, D.S.; Gerber, S.A.; Gygi, S.P.; Moazed, D. Budding yeast silencing complexes and regulation of Sir2 activity by protein-protein interactions. Mol. Cell. Biol. 2004, 24, 6931–6946. [Google Scholar] [CrossRef]
- Rudner, A.D.; Hall, B.E.; Ellenberger, T.; Moazed, D. A nonhistone protein-protein interaction required for assembly of the SIR complex and silent chromatin. Mol. Cell. Biol. 2005, 25, 4514–4528. [Google Scholar] [CrossRef] [PubMed]
- Cubizolles, F.; Martino, F.; Perrod, S.; Gasser, S.M. A homotrimer-heterotrimer switch in Sir2 structure differentiates rDNA and telomeric silencing. Mol. Cell 2006, 21, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Buchberger, J.R.; Onishi, M.; Li, G.; Seebacher, J.; Rudner, A.D.; Gygi, S.P.; Moazed, D. Sir3-nucleosome interactions in spreading of silent chromatin in Saccharomyces cerevisiae. Mol. Cell. Biol. 2008, 28, 6903–6918. [Google Scholar] [CrossRef] [PubMed]
- Strahl-Bolsinger, S.; Hecht, A.; Luo, K.; Grunstein, M. SIR2 and SIR4 interactions differ in core and extended telomeric heterochromatin in yeast. Genes Dev. 1997, 11, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, G.J.; Tanny, J.C.; Rudner, A.D.; Gerber, S.A.; Danaie, S.; Gygi, S.P.; Moazed, D. Steps in assembly of silent chromatin in yeast: Sir3-independent binding of a Sir2/Sir4 complex to silencers and role for Sir2-dependent deacetylation. Mol. Cell. Biol. 2002, 22, 4167–4180. [Google Scholar] [CrossRef]
- Ghidelli, S.; Donze, D.; Dhillon, N.; Kamakaka, R.T. Sir2p exists in two nucleosome-binding complexes with distinct deacetylase activities. EMBO J. 2001, 20, 4522–4535. [Google Scholar] [CrossRef]
- Landry, J.; Sutton, A.; Tafrov, S.T.; Heller, R.C.; Stebbins, J.; Pillus, L.; Sternglanz, R. The silencing protein SIR2 and its homologs are NAD-dependent protein deacetylases. Proc. Natl. Acad. Sci. USA 2000, 97, 5807–5811. [Google Scholar] [CrossRef]
- Hickman, M.A.; Froyd, C.A.; Rusche, L.N. Reinventing heterochromatin in budding yeasts: Sir2 and the origin recognition complex take center stage. Eukaryot. Cell 2011, 10, 1183–1192. [Google Scholar] [CrossRef]
- Bi, X. Heterochromatin structure: Lessons from the budding yeast. IUBMB Life 2014, 66, 657–666. [Google Scholar] [CrossRef]
- Hecht, A.; Laroche, T.; Strahl-Bolsinger, S.; Gasser, S.M.; Grunstein, M. Histone H3 and H4 N-termini interact with SIR3 and SIR4 proteins: A molecular model for the formation of heterochromatin in yeast. Cell 1995, 80, 583–592. [Google Scholar] [CrossRef]
- Sampath, V.; Yuan, P.; Wang, I.X.; Prugar, E.; van Leeuwen, F.; Sternglanz, R. Mutational analysis of the Sir3 BAH domain reveals multiple points of interaction with nucleosomes. Mol. Cell. Biol. 2009, 29, 2532–2545. [Google Scholar] [CrossRef] [PubMed]
- Carmen, A.A.; Milne, L.; Grunstein, M. Acetylation of the yeast histone H4 N-terminus regulates its binding to heterochromatin protein SIR3. J. Biol. Chem. 2001, 19, 19. [Google Scholar] [CrossRef] [PubMed]
- Georgel, P.T.; DeBeer, M.A.P.; Pietz, G.; Fox, C.A.; Hansen, J.C. Sir3-dependent assembly of supramolecular chromatin structures in vitro. Proc. Natl. Acad. Sci. USA 2001, 98, 8584–8589. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.; Utley, R.T.; Lacoste, N.; Tan, S.; Briggs, S.D.; Cote, J. Interplay of chromatin modifiers on a short basic patch of histone H4 tail defines the boundary of telomeric heterochromatin. Mol. Cell 2007, 28, 1002–1014. [Google Scholar] [CrossRef] [PubMed]
- Martino, F.; Kueng, S.; Robinson, P.; Tsai-Pflugfelder, M.; van Leeuwen, F.; Ziegler, M.; Cubizolles, F.; Cockell, M.M.; Rhodes, D.; Gasser, S.M. Reconstitution of yeast silent chromatin: Multiple contact sites and O-AADPR binding load SIR complexes onto nucleosomes in vitro. Mol. Cell 2009, 33, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, N.; Redon, S.; Pfeiffer, V.; Dees, M.; Lingner, J.; Luke, B. Subtelomeric repetitive elements determine TERRA regulation by Rap1/Rif and Rap1/Sir complexes in yeast. EMBO Rep. 2011, 12, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, G.; Van der Ploeg, L.H. Transcription of telomere repeats in protozoa. EMBO J. 1989, 8, 2633–2638. [Google Scholar] [CrossRef]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric repeat containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef]
- Schoeftner, S.; Blasco, M.A. Developmentally regulated transcription of mammalian telomeres by DNA-dependent RNA polymerase II. Nat. Cell Biol. 2008, 10, 228–236. [Google Scholar] [CrossRef]
- Luke, B.; Panza, A.; Redon, S.; Iglesias, N.; Li, Z.; Lingner, J. The Rat1p 5’ to 3’ exonuclease degrades telomeric repeat-containing RNA and promotes telomere elongation in Saccharomyces cerevisiae. Mol. Cell 2008, 32, 465–477. [Google Scholar] [CrossRef]
- Bah, A.; Wischnewski, H.; Shchepachev, V.; Azzalin, C.M. The telomeric transcriptome of Schizosaccharomyces pombe. Nucleic Acids Res. 2012, 40, 2995–3005. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Nanavaty, V.P.; Li, B. Telomere and subtelomere R-loops and antigenic variation in trypanosomes. J. Mol. Biol. 2019, 432, 4167–4185. [Google Scholar] [CrossRef] [PubMed]
- Bettin, N.; Pegorar, C.O.; Cusanelli, E. The Emerging Roles of TERRA in Telomere Maintenance and Genome Stability. Cells 2019, 8, 246. [Google Scholar] [CrossRef] [PubMed]
- Kroupa, M.; Tomasova, K.; Kavec, M.; Skrobanek, P.; Buchler, T.; Kumar, R.; Vodickova, L.; Vodicka, P. TElomeric repeat-containing RNA (TERRA): Physiological functions and relevance in cancer. Front. Oncol. 2022, 12, 913314. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, M.; Chartrand, P. TERRA, a Multifaceted Regulator of Telomerase Activity at Telomeres. J. Mol. Biol. 2020, 432, 4232–4243. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.V.; Feretzaki, M.; Lingner, J. The makings of TERRA R-loops at chromosome ends. Cell Cycle 2021, 20, 1745–1759. [Google Scholar] [CrossRef]
- Crossley, M.P.; Bocek, M.; Cimprich, K.A. R-Loops as Cellular Regulators and Genomic Threats. Mol. Cell 2019, 73, 398–411. [Google Scholar] [CrossRef]
- Hegazy, Y.A.; Fernando, C.M.; Tran, E.J. The balancing act of R-loop biology: The good, the bad, and the ugly. J. Biol. Chem. 2020, 295, 905–913. [Google Scholar] [CrossRef]
- Toubiana, S.; Selig, S. DNA:RNA hybrids at telomeres—When it is better to be out of the (R) loop. FEBS J. 2018, 285, 2552–2566. [Google Scholar] [CrossRef]
- Castano, I.; Pan, S.J.; Zupancic, M.; Hennequin, C.; Dujon, B.; Cormack, B.P. Telomere length control and transcriptional regulation of subtelomeric adhesins in Candida glabrata. Mol. Microbiol. 2005, 55, 1246–1258. [Google Scholar] [CrossRef]
- De Las Penas, A.; Pan, S.J.; Castano, I.; Alder, J.; Cregg, R.; Cormack, B.P. Virulence-related surface glycoproteins in the yeast pathogen Candida glabrata are encoded in subtelomeric clusters and subject to RAP1- and SIR-dependent transcriptional silencing. Genes Dev. 2003, 17, 2245–2258. [Google Scholar] [CrossRef] [PubMed]
- Domergue, R.; Castano, I.; De Las Penas, A.; Zupancic, M.; Lockatell, V.; Hebel, J.R.; Johnson, D.; Cormack, B.P. Nicotinic acid limitation regulates silencing of Candida adhesins during UTI. Science 2005, 308, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Weuts, A.; Voet, T.; Verbeeck, J.; Lambrechts, N.; Wirix, E.; Schoonjans, L.; Danloy, S.; Marynen, P.; Froyen, G. Telomere length homeostasis and telomere position effect on a linear human artificial chromosome are dictated by the genetic background. Nucleic Acids Res. 2012, 40, 11477–11489. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, Z.; Liu, X.; Wang, S.; Zhang, Y.; He, X.; Sun, S.; Ma, S.; Shyh-Chang, N.; Liu, F.; et al. Telomere-dependent and telomere-independent roles of RAP1 in regulating human stem cell homeostasis. Protein Cell 2019, 10, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Shay, J.W. Long-range telomere regulation of gene expression: Telomere looping and telomere position effect over long distances (TPE-OLD). Differentiation 2018, 99, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sidlauskaite, E.; Le Gall, L.; Mariot, V.; Dumonceaux, J. DUX4 Expression in FSHD Muscles: Focus on Its mRNA Regulation. J. Pers. Med. 2020, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Lemmers, R.J.; van der Maarel, S.M.; van Deutekom, J.C.; van der Wielen, M.J.; Deidda, G.; Dauwerse, H.G.; Hewitt, J.; Hofker, M.; Bakker, E.; Padberg, G.W.; et al. Inter- and intrachromosomal sub-telomeric rearrangements on 4q35: Implications for facioscapulohumeral muscular dystrophy (FSHD) aetiology and diagnosis. Hum. Mol. Genet. 1998, 7, 1207–1214. [Google Scholar] [CrossRef] [PubMed]
- Robin, J.D.; Ludlow, A.T.; Batten, K.; Gaillard, M.C.; Stadler, G.; Magdinier, F.; Wright, W.E.; Shay, J.W. SORBS2 transcription is activated by telomere position effect-over long distance upon telomere shortening in muscle cells from patients with facioscapulohumeral dystrophy. Genome Res. 2015, 25, 1781–1790. [Google Scholar] [CrossRef]
- Milne, G.T.; Jin, S.; Shannon, K.B.; Weaver, D.T. Mutations in two Ku homologs define a DNA end-joining repair pathway in Saccharomyces cerevisiae. Mol. Cell. Biol. 1996, 16, 4189–4198. [Google Scholar] [CrossRef]
- Teo, S.H.; Jackson, S.P. Identification of Saccharomyces cerevisiae DNA ligase IV: Involvement in DNA double-strand break repair. Embo J. 1997, 16, 4788–4795. [Google Scholar] [CrossRef]
- Wilson, T.E.; Grawunder, U.; Lieber, M.R. Yeast DNA ligase IV mediates non-homologous DNA end joining. Nature 1997, 388, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, G.; Lindahl, T.; Schar, P. Saccharomyces cerevisiae LIF1: A function involved in DNA double-strand break repair related to mammalian XRCC4. Embo. J. 1998, 17, 4188–4198. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Trujillo, K.; Ramos, W.; Sung, P.; Tomkinson, A.E. Promotion of Dnl4-catalyzed DNA end-joining by the Rad50/Mre11/Xrs2 and Hdf1/Hdf2 complexes. Mol. Cell 2001, 8, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Sarthy, J.; Bae, N.S.; Scrafford, J.; Baumann, P. Human RAP1 inhibits non-homologous end joining at telomeres. EMBO J. 2009, 28, 3390–3399. [Google Scholar] [CrossRef] [PubMed]
- Bae, N.S.; Baumann, P. A RAP1/TRF2 complex inhibits nonhomologous end-joining at human telomeric DNA ends. Mol. Cell 2007, 26, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Lototska, L.; Yue, J.X.; Li, J.; Giraud-Panis, M.J.; Songyang, Z.; Royle, N.J.; Liti, G.; Ye, J.; Gilson, E.; Mendez-Bermudez, A. Human RAP1 specifically protects telomeres of senescent cells from DNA damage. EMBO Rep. 2020, 21, e49076. [Google Scholar] [CrossRef]
- Yu, E.Y.; Yen, W.F.; Steinberg-Neifach, O.; Lue, N.F. Rap1 in Candida albicans: An unusual structural organization and a critical function in suppressing telomere recombination. Mol. Cell. Biol. 2010, 30, 1254–1268. [Google Scholar] [CrossRef]
- Malyavko, A.N.; Petrova, O.A.; Zvereva, M.I.; Dontsova, O.A. Functional duplication of Rap1 in methylotrophic yeasts. Sci. Rep. 2019, 9, 7196. [Google Scholar] [CrossRef]
- Sfeir, A.; Kabir, S.; van Overbeek, M.; Celli, G.B.; de Lange, T. Loss of Rap1 induces telomere recombination in the absence of NHEJ or a DNA damage signal. Science 2010, 327, 1657–1661. [Google Scholar] [CrossRef]
- Rai, R.; Chen, Y.; Lei, M.; Chang, S. TRF2-RAP1 is required to protect telomeres from engaging in homologous recombination-mediated deletions and fusions. Nat. Commun. 2016, 7, 10881. [Google Scholar] [CrossRef]
- Mugnier, M.R.; Stebbins, C.E.; Papavasiliou, F.N. Masters of Disguise: Antigenic Variation and the VSG Coat in Trypanosoma brucei. PLoS Pathog. 2016, 12, e1005784. [Google Scholar] [CrossRef]
- Wickstead, B.; Ersfeld, K.; Gull, K. The small chromosomes of Trypanosoma brucei involved in antigenic variation are constructed around repetitive palindromes. Genome Res. 2004, 14, 1014–1024. [Google Scholar] [CrossRef] [PubMed]
- Cross, G.A.M. Identification, purification and properties of clone-specific glycoprotein antigens constituting the surface coat of Trypanosoma brucei. Parasitology 1975, 71, 393–417. [Google Scholar] [CrossRef] [PubMed]
- Gunzl, A.; Bruderer, T.; Laufer, G.; Schimanski, B.; Tu, L.C.; Chung, H.M.; Lee, P.T.; Lee, M.G. RNA polymerase I transcribes procyclin genes and variant surface glycoprotein gene expression sites in Trypanosoma brucei. Eukaryot. Cell 2003, 2, 542–551. [Google Scholar] [CrossRef]
- Aresta-Branco, F.; Sanches-Vaz, M.; Bento, F.; Rodrigues, J.A.; Figueiredo, L.M. African trypanosomes expressing multiple VSGs are rapidly eliminated by the host immune system. Proc. Natl. Acad. Sci. USA 2019, 116, 20725–20735. [Google Scholar] [CrossRef]
- Myler, P.; Nelson, R.G.; Agabian, N.; Stuart, K. Two mechanisms of expression of a variant antigen gene of Trypanosoma brucei. Nature 1984, 309, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Myler, P.J.; Allison, J.; Agabian, N.; Stuart, K. Antigenic variation in African trypanosomes by gene replacement or activation of alternative telomeres. Cell 1984, 39, 203–211. [Google Scholar] [CrossRef]
- Rudenko, G.; McCulloch, R.; Dirksmulder, A.; Borst, P. Telomere exchange can be an important mechanism of variant surface glycoprotein gene switching in Trypanosoma brucei. Mol. Biochem. Parasitol. 1996, 80, 65–75. [Google Scholar] [CrossRef]
- Horn, D.; McCulloch, R. Molecular mechanisms underlying the control of antigenic variation in African trypanosomes. Curr. Opin. Microbiol. 2010, 13, 700–705. [Google Scholar] [CrossRef]
- Eid, J.; Sollner-Webb, B. Stable integrative transformation of Trypanosoma brucei that occurs exclusively by homologous recombination. Proc. Natl. Acad. Sci. USA 1991, 88, 2118–2121. [Google Scholar] [CrossRef]
- Lee, M.-S.; van der Ploeg, L.H.T. Homologous recombination and stable transfection in the parasitic protozoan Trypanosoma brucei. Science 1990, 250, 1583–1587. [Google Scholar] [CrossRef] [PubMed]
- Barnes, R.L.; McCulloch, R. Trypanosoma brucei homologous recombination is dependent on substrate length and homology, though displays a differential dependence on mismatch repair as substrate length decreases. Nucleic Acids Res. 2007, 35, 3478–3493. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, R.; Barry, J.D. A role for RAD51 and homologous recombination in Trypanosoma brucei antigenic variation. Genes Dev. 1999, 13, 2875–2888. [Google Scholar] [CrossRef] [PubMed]
- Dobson, R.; Stockdale, C.; Lapsley, C.; Wilkes, J.; McCulloch, R. Interactions among Trypanosoma brucei RAD51 paralogues in DNA repair and antigenic variation. Mol. Microbiol. 2011, 81, 434–456. [Google Scholar] [CrossRef] [PubMed]
- Hartley, C.L.; McCulloch, R. Trypanosoma brucei BRCA2 acts in antigenic variation and has undergone a recent expansion in BRC repeat number that is important during homologous recombination. Mol. Microbiol. 2008, 68, 1237–1251. [Google Scholar] [CrossRef] [PubMed]
- Girasol, M.J.; Krasilnikova, M.; Marques, C.A.; Damasceno, J.D.; Lapsley, C.; Lemgruber, L.; Burchmore, R.; Beraldi, D.; Carruthers, R.; Briggs, E.M.; et al. RAD51-mediated R-loop formation acts to repair transcription-associated DNA breaks driving antigenic variation in Trypanosoma brucei. Proc. Natl. Acad. Sci. USA 2023, 120, e2309306120. [Google Scholar] [CrossRef] [PubMed]
- Burton, P.; McBride, D.J.; Wilkes, J.M.; Barry, J.D.; McCulloch, R. Ku heterodimer-independent end joining in Trypanosoma brucei cell extracts relies upon sequence microhomology. Eukaryot. Cell 2007, 6, 1773–1781. [Google Scholar] [CrossRef]
- Conway, C.; Proudfoot, C.; Burton, P.; Barry, J.D.; McCulloch, R. Two pathways of homologous recombination in Trypanosoma brucei. Mol. Microbiol. 2002, 45, 1687–1700. [Google Scholar] [CrossRef]
- Glover, L.; McCulloch, R.; Horn, D. Sequence homology and microhomology dominate chromosomal double-strand break repair in African trypanosomes. Nucleic Acids Res. 2008, 36, 2608–2618. [Google Scholar] [CrossRef]
- Barbet, A.F.; Kamper, S.M. The importance of mosaic genes to trypanosome survival. Parasitol. Today 1993, 9, 63–66. [Google Scholar] [CrossRef]
- Kamper, S.M.; Barbet, A.F. Surface epitope variation via mosaic gene formation is potential key to long-term survival of Trypanosoma brucei. Mol. Biochem. Parasitol. 1992, 53, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Benmerzouga, I.; Concepcion-Acevedo, J.; Kim, H.S.; Vandoros, A.V.; Cross, G.A.M.; Klingbeil, M.M.; Li, B. Trypanosoma brucei Orc1 is essential for nuclear DNA replication and affects both VSG silencing and VSG switching. Mol. Microbiol. 2013, 87, 196–210. [Google Scholar] [CrossRef]
- Devlin, R.; Marques, C.A.; Paape, D.; Prorocic, M.; Zurita-Leal, A.C.; Campbell, S.J.; Lapsley, C.; Dickens, N.; McCulloch, R. Mapping replication dynamics in Trypanosoma brucei reveals a link with telomere transcription and antigenic variation. Elife 2016, 5, e12765. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Cross, G.A.M. TOPO3alpha influences antigenic variation by monitoring expression-site-associated VSG switching in Trypanosoma brucei. PLoS Pathog. 2010, 6, e1000992. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Cross, G.A.M. Identification of Trypanosoma brucei RMI1/BLAP75 homologue and its roles in antigenic variation. PLoS ONE 2011, 6, e25313. [Google Scholar] [CrossRef] [PubMed]
- Boothroyd, C.E.; Dreesen, O.; Leonova, T.; Ly, K.I.; Figueiredo, L.M.; Cross, G.A.M.; Papavasiliou, F.N. A yeast-endonuclease-generated DNA break induces antigenic switching in Trypanosoma brucei. Nature 2009, 459, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Glover, L.; Alsford, S.; Horn, D. DNA break site at fragile subtelomeres determines probability and mechanism of antigenic variation in African trypanosomes. PLoS Pathog. 2013, 9, e1003260. [Google Scholar] [CrossRef] [PubMed]
- Thivolle, A.; Mehnert, A.K.; Tihon, E.; McLaughlin, E.; Dujeancourt-Henry, A.; Glover, L. DNA double strand break position leads to distinct gene expression changes and regulates VSG switching pathway choice. PLoS Pathog. 2021, 17, e1010038. [Google Scholar] [CrossRef]
- Alsford, S.; Horn, D.; Glover, L. DNA breaks as triggers for antigenic variation in African trypanosomes. Genome Biol. 2009, 10, 223. [Google Scholar] [CrossRef]
- Hovel-Miner, G.A.; Boothroyd, C.E.; Mugnier, M.; Dreesen, O.; Cross, G.A.M.; Papavasiliou, F.N. Telomere length affects the frequency and mechanism of antigenic variation in Trypanosoma brucei. PLoS Pathog. 2012, 8, e1002900. [Google Scholar] [CrossRef]
- Bernards, A.; Michels, P.A.M.; Lincke, C.R.; Borst, P. Growth of chromosome ends in multiplying trypanosomes. Nature 1983, 303, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Jehi, S.E.; Nanavaty, V.; Li, B. Trypanosoma brucei TIF2 and TRF suppress VSG switching using overlapping and independent mechanisms. PLoS ONE 2016, 11, e0156746. [Google Scholar] [CrossRef] [PubMed]
- Jehi, S.E.; Li, X.; Sandhu, R.; Ye, F.; Benmerzouga, I.; Zhang, M.; Zhao, Y.; Li, B. Suppression of subtelomeric VSG switching by Trypanosoma brucei TRF requires its TTAGGG repeat-binding activity. Nucleic Acids Res. 2014, 42, 12899–12911. [Google Scholar] [CrossRef]
- Khamlichi, A.A.; Feil, R. Parallels between Mammalian Mechanisms of Monoallelic Gene Expression. Trends Genet. 2018, 34, 954–971. [Google Scholar] [CrossRef] [PubMed]
- Chess, A. Monoallelic Gene Expression in Mammals. Annu. Rev. Genet. 2016, 50, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Oppikofer, M.; Kueng, S.; Gasser, S.M. SIR-nucleosome interactions: Structure-function relationships in yeast silent chromatin. Gene 2013, 527, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Tarutani, Y.; Takayama, S. Monoallelic gene expression and its mechanisms. Curr. Opin. Plant Biol. 2011, 14, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Massah, S.; Beischlag, T.V.; Prefontaine, G.G. Epigenetic events regulating monoallelic gene expression. Crit. Rev. Biochem. Mol. Biol. 2015, 50, 337–358. [Google Scholar] [CrossRef]
- Gunzl, A.; Kirkham, J.K.; Nguyen, T.N.; Badjatia, N.; Park, S.H. Mono-allelic VSG expression by RNA polymerase I in Trypanosoma brucei: Expression site control from both ends? Gene 2015, 556, 68–73. [Google Scholar] [CrossRef]
- Cestari, I.; Stuart, K. Transcriptional regulation of telomeric expression sites and antigenic variation in trypanosomes. Curr. Genom. 2018, 19, 119–132. [Google Scholar] [CrossRef]
- Faria, J.; Briggs, E.M.; Black, J.A.; McCulloch, R. Emergence and adaptation of the cellular machinery directing antigenic variation in the African trypanosome. Curr. Opin. Microbiol. 2022, 70, 102209. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.; Gull, K. A pol I transcriptional body associated with VSG mono-allelic expression in Trypanosoma brucei. Nature 2001, 414, 759–763. [Google Scholar] [CrossRef] [PubMed]
- López-Escobar, L.; Hänisch, B.; Halliday, C.; Ishii, M.; Akiyoshi, B.; Dean, S.; Sunter, J.D.; Wheeler, R.J.; Gull, K. Stage-specific transcription activator ESB1 regulates monoallelic antigen expression in Trypanosoma brucei. Nat. Microbiol. 2022, 7, 1280–1290. [Google Scholar] [CrossRef]
- Lopez-Farfan, D.; Bart, J.M.; Rojas-Barros, D.I.; Navarro, M. SUMOylation by the E3 Ligase TbSIZ1/PIAS1 Positively Regulates VSG Expression in Trypanosoma brucei. PLoS Pathog. 2014, 10, e1004545. [Google Scholar] [CrossRef] [PubMed]
- Saura, A.; Iribarren, P.A.; Rojas-Barros, D.; Bart, J.M.; López-Farfán, D.; Andrés-León, E.; Vidal-Cobo, I.; Boehm, C.; Alvarez, V.E.; Field, M.C.; et al. SUMOylated SNF2PH promotes variant surface glycoprotein expression in bloodstream trypanosomes. EMBO Rep. 2019, 20, e48029. [Google Scholar] [CrossRef] [PubMed]
- Field, M.C. Deviating from the norm: Nuclear organisation in trypanosomes. Curr. Opin. Cell Biol. 2023, 85, 102234. [Google Scholar] [CrossRef] [PubMed]
- Berriman, M.; Ghedin, E.; Hertz-Fowler, C.; Blandin, G.; Renauld, H.; Bartholomeu, D.C.; Lennard, N.J.; Caler, E.; Hamlin, N.E.; Haas, B.; et al. The genome of the African trypanosome Trypanosoma brucei. Science 2005, 309, 416–422. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, N.M.; Myler, P.J.; Blandin, G.; Berriman, M.; Crabtree, J.; Aggarwal, G.; Caler, E.; Renauld, H.; Worthey, E.A.; Hertz-Fowler, C.; et al. Comparative genomics of trypanosomatid parasitic protozoa. Science 2005, 309, 404–409. [Google Scholar] [CrossRef]
- Michaeli, S. Trans-splicing in trypanosomes: Machinery and its impact on the parasite transcriptome. Future Microbiol. 2011, 6, 459–474. [Google Scholar] [CrossRef]
- Gunzl, A. The pre-mRNA splicing machinery of trypanosomes: Complex or simplified? Eukaryot. Cell 2010, 9, 1159–1170. [Google Scholar] [CrossRef]
- Faria, J.; Luzak, V.; Müller, L.S.M.; Brink, B.G.; Hutchinson, S.; Glover, L.; Horn, D.; Siegel, T.N. Spatial integration of transcription and splicing in a dedicated compartment sustains monogenic antigen expression in African trypanosomes. Nat. Microbiol. 2021, 6, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Glover, L.; Hutchinson, S.; Alsford, S.; Horn, D. VEX1 controls the allelic exclusion required for antigenic variation in trypanosomes. Proc. Natl. Acad. Sci. USA 2016, 113, 7225–7230. [Google Scholar] [CrossRef] [PubMed]
- Faria, J.; Glover, L.; Hutchinson, S.; Boehm, C.; Field, M.C.; Horn, D. Monoallelic expression and epigenetic inheritance sustained by a Trypanosoma brucei variant surface glycoprotein exclusion complex. Nat. Commun. 2019, 10, 3023. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.; Nguyen, B.N.; Lee, J.H.; Panigrahi, A.K.; Gunzl, A. Characterization of a novel class I transcription factor A (CITFA) subunit that is indispensable for transcription by the multifunctional RNA polymerase I of Trypanosoma brucei. Eukaryot. Cell 2012, 11, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.; Muller, L.S.; Park, S.H.; Siegel, T.N.; Gunzl, A. Promoter occupancy of the basal class I transcription factor A differs strongly between active and silent VSG expression sites in Trypanosoma brucei. Nucleic Acids Res. 2014, 42, 3164–3176. [Google Scholar] [CrossRef] [PubMed]
- Kassem, A.; Pays, E.; Vanhamme, L. Transcription is initiated on silent variant surface glycoprotein expression sites despite monoallelic expression in Trypanosoma brucei. Proc. Natl. Acad. Sci. USA 2014, 111, 8943–8948. [Google Scholar] [CrossRef] [PubMed]
- Vanhamme, L.; Poelvoorde, P.; Pays, A.; Tebabi, P.; Van Xong, H.; Pays, E. Differential RNA elongation controls the variant surface glycoprotein gene expression sites of Trypanosoma brucei. Mol. Microbiol. 2000, 36, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, G. Epigenetics and transcriptional control in African trypanosomes. Essays Biochem. 2010, 48, 201–219. [Google Scholar]
- Stanne, T.M.; Rudenko, G. Active VSG expression sites in Trypanosoma brucei are depleted of nucleosomes. Eukaryot. Cell 2010, 9, 136–147. [Google Scholar] [CrossRef]
- Figueiredo, L.M.; Cross, G.A.M. Nucleosomes are depleted at the VSG expression site transcribed by RNA polymerase I in African trypanosomes. Eukaryot. Cell 2010, 9, 148–154. [Google Scholar] [CrossRef]
- Povelones, M.L.; Gluenz, E.; Dembek, M.; Gull, K.; Rudenko, G. Histone H1 plays a role in heterochromatin formation and VSG expression site silencing in Trypanosoma brucei. PLoS Pathog. 2012, 8, e1003010. [Google Scholar] [CrossRef] [PubMed]
- Alsford, S.; Horn, D. Cell-cycle-regulated control of VSG expression site silencing by histones and histone chaperones ASF1A and CAF-1b in Trypanosoma brucei. Nucleic Acids Res. 2012, 40, 10150–10160. [Google Scholar] [CrossRef] [PubMed]
- Hughes, K.; Wand, M.; Foulston, L.; Young, R.; Harley, K.; Terry, S.; Ersfeld, K.; Rudenko, G. A novel ISWI is involved in VSG expression site downregulation in African trypanosomes. EMBO J. 2007, 26, 2400–2410. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, M.S.; Kushwaha, M.; Ersfeld, K.; Fullbrook, A.; Stanne, T.M.; Rudenko, G. NLP is a novel transcription regulator involved in VSG expression site control in Trypanosoma brucei. Nucleic Acids Res. 2011, 39, 2018–2031. [Google Scholar] [CrossRef]
- Stanne, T.M.; Narayanan, M.S.; Ridewood, S.; Ling, A.; Witmer, K.; Kushwaha, M.; Wiesler, S.; Wickstead, B.; Wood, J.; Rudenko, G. Identification of the ISWI chromatin remodeling complex of the early branching eukaryote Trypanosoma brucei. J. Biol. Chem. 2015, 290, 29760. [Google Scholar]
- Denninger, V.; Fullbrook, A.; Bessat, M.; Ersfeld, K.; Rudenko, G. The FACT subunit TbSpt16 is involved in cell cycle specific control of VSG expression sites in Trypanosoma brucei. Mol. Microbiol. 2010, 78, 459–474. [Google Scholar] [CrossRef] [PubMed]
- Denninger, V.; Rudenko, G. FACT plays a major role in histone dynamics affecting VSG expression site control in Trypanosoma brucei. Mol. Microbiol. 2014, 94, 945–962. [Google Scholar] [CrossRef]
- Narayanan, M.S.; Rudenko, G. TDP1 is an HMG chromatin protein facilitating RNA polymerase I transcription in African trypanosomes. Nucleic Acids Res. 2013, 41, 2981–2992. [Google Scholar] [CrossRef]
- Figueiredo, L.M.; Janzen, C.J.; Cross, G.A.M. A histone methyltransferase modulates antigenic variation in African trypanosomes. PLoS Biol. 2008, 6, e161. [Google Scholar] [CrossRef]
- Sandhu, R.; Li, B. Telomerase activity is required for the telomere G-overhang structure in Trypanosoma brucei. Sci. Rep. 2017, 7, 15983. [Google Scholar] [CrossRef]
- Sandhu, R.; Li, B. Examination of the telomere G-overhang structure in Trypanosoma brucei. J. Vis. Exp. 2011, 47, 1959. [Google Scholar]
- Dreesen, O.; Li, B.; Cross, G.A.M. Telomere structure and shortening in telomerase-deficient Trypanosoma brucei. Nucleic Acids Res. 2005, 33, 4536–4543. [Google Scholar] [CrossRef]
- Sandhu, R.; Sanford, S.; Basu, S.; Park, M.; Pandya, U.M.; Li, B.; Chakrabarti, K. A trans-spliced telomerase RNA dictates telomere synthesis in Trypanosoma brucei. Cell Res. 2013, 23, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Kolet, L.; Doniger, T.; Biswas, V.K.; Unger, R.; Tzfati, Y.; Michaeli, S. The Trypanosoma brucei telomerase RNA (TER) homologue binds core proteins of the C/D snoRNA family. FEBS Lett. 2013, 587, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.; Monroy-Eklund, A.; Klotz, K.; Saha, A.; Davis, J.; Li, B.; Laederach, A.; Chakrabarti, K. In vivo architecture of the telomerase RNA catalytic core in Trypanosoma brucei. Nucleic Acids Res. 2021, 49, 12445–12466. [Google Scholar] [CrossRef] [PubMed]
- Leal, A.Z.; Schwebs, M.; Briggs, E.; Weisert, N.; Reis, H.; Lemgruber, L.; Luko, K.; Wilkes, J.; Butter, F.; McCulloch, R.; et al. Genome maintenance functions of a putative Trypanosoma brucei translesion DNA polymerase include telomere association and a role in antigenic variation. Nucleic Acids Res. 2020, 48, 9660–9680. [Google Scholar] [CrossRef]
- Rudd, S.G.; Glover, L.; Jozwiakowski, S.K.; Horn, D.; Doherty, A.J. PPL2 translesion polymerase is essential for the completion of chromosomal DNA replication in the African trypanosome. Mol. Cell 2013, 52, 554–565. [Google Scholar] [CrossRef]
- Janzen, C.J.; Lander, F.; Dreesen, O.; Cross, G.A.M. Telomere length regulation and transcriptional silencing in KU80-deficient Trypanosoma brucei. Nucleic Acids Res. 2004, 32, 6575–6584. [Google Scholar] [CrossRef]
- Conway, C.; McCulloch, R.; Ginger, M.L.; Robinson, N.P.; Browitt, A.; Barry, J.D. Ku is important for telomere maintenance, but not for differential expression of telomeric VSG genes, in African trypanosomes. J. Biol. Chem. 2002, 277, 21269–21277. [Google Scholar] [CrossRef]
- Saha, A.; Gaurav, A.K.; Pandya, U.M.; Afrin, M.; Sandhu, R.; Nanavaty, V.; Schnur, B.; Li, B. TbTRF suppresses the TERRA level and regulates the cell cycle-dependent TERRA foci number with a TERRA binding activity in its C-terminal Myb domain. Nucleic Acids Res. 2021, 49, 5637–5653. [Google Scholar] [CrossRef]
- Giresi, P.G.; Lieb, J.D. Isolation of active regulatory elements from eukaryotic chromatin using FAIRE (Formaldehyde Assisted Isolation of Regulatory Elements). Methods 2009, 48, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Renauld, H.; Aparicio, O.M.; Zierath, P.D.; Billington, B.L.; Chhablani, S.K.; Gottschling, D.E. Silent domains are assembled continuously from the telomere and are defined by promoter distance and strength, and by SIR3 dosage. Genes Dev. 1993, 7, 1133–1145. [Google Scholar] [CrossRef]
- Konig, P.; Fairall, L.; Rhodes, D. Sequence-specific DNA recognition by the Myb-like domain of the human telomere binding protein TRF1: A model for the protein-DNA complex. Nucleic Acids Res. 1998, 26, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Biswas, K.; Rieger, K.J.; Morschhauser, J. Functional analysis of CaRAP1, encoding the Repressor/activator protein 1 of Candida albicans. Gene 2003, 307, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Fujita, I.; Tanaka, M.; Kanoh, J. Identification of the functional domains of the telomere protein Rap1 in Schizosaccharomyces pombe. PLoS ONE 2012, 7, e49151. [Google Scholar] [CrossRef] [PubMed]
- Nett, I.R.; Martin, D.M.; Miranda-Saavedra, D.; Lamont, D.; Barber, J.D.; Mehlert, A.; Ferguson, M.A. The phosphoproteome of bloodstream form Trypanosoma brucei, causative agent of African sleeping sickness. Mol. Cell Proteom. 2009, 8, 1527–1538. [Google Scholar] [CrossRef]
- Urbaniak, M.D.; Martin, D.M.; Ferguson, M.A. Global quantitative SILAC phosphoproteomics reveals differential phosphorylation is widespread between the procyclic and bloodstream form lifecycle stages of Trypanosoma brucei. J. Proteome Res. 2013, 12, 2233–2244. [Google Scholar] [CrossRef]
- Cestari, I.; McLeland-Wieser, H.; Stuart, K. Nuclear phosphatidylinositol 5-phosphatase is essential for allelic exclusion of variant surface glycoprotein genes in trypanosomes. Mol. Cell. Biol. 2019, 39, e00395-18. [Google Scholar] [CrossRef]
- Cestari, I.; Stuart, K. Inositol phosphate pathway controls transcription of telomeric expression sites in trypanosomes. Proc. Natl. Acad. Sci. USA 2015, 112, E2803–E2812. [Google Scholar] [CrossRef]
- Touray, A.O.; Rajesh, R.; Isebe, T.; Sternlieb, T.; Loock, M.; Kutova, O.; Cestari, I. PI(3,4,5)P3 allosteric regulation of repressor activator protein 1 controls antigenic variation in trypanosomes. Elife 2023, 12, RP89331. [Google Scholar] [CrossRef] [PubMed]
- Martinez, P.; Gomez-Lopez, G.; Garcia, F.; Mercken, E.; Mitchell, S.; Flores, J.M.; de Cabo, R.; Blasco, M.A. RAP1 protects from obesity through its extratelomeric role regulating gene expression. Cell Rep. 2013, 3, 2059–2074. [Google Scholar] [CrossRef]
- Yeung, F.; Ramírez, C.M.; Mateos-Gomez, P.A.; Pinzaru, A.; Ceccarini, G.; Kabir, S.; Fernández-Hernando, C.; Sfeir, A. Nontelomeric role for Rap1 in regulating metabolism and protecting against obesity. Cell Rep. 2013, 3, 1847–1856. [Google Scholar] [CrossRef] [PubMed]
- Budzak, J.; Jones, R.; Tschudi, C.; Kolev, N.G.; Rudenko, G. An assembly of nuclear bodies associates with the active VSG expression site in African trypanosomes. Nat. Commun. 2022, 13, 101. [Google Scholar] [CrossRef] [PubMed]
- Budzak, J.; Rudenko, G. Pedal to the Metal: Nuclear Splicing Bodies Turbo-Charge VSG mRNA Production in African Trypanosomes. Front. Cell Dev. Biol. 2022, 10, 876701. [Google Scholar] [CrossRef] [PubMed]
- Sheader, K.; Vaughan, S.; Minchin, J.; Hughes, K.; Gull, K.; Rudenko, G. Variant surface glycoprotein RNA interference triggers a precytokinesis cell cycle arrest in African trypanosomes. Proc. Natl. Acad. Sci. USA 2005, 102, 8716–8721. [Google Scholar] [CrossRef]
- Smith, T.K.; Vasileva, N.; Gluenz, E.; Terry, S.; Portman, N.; Kramer, S.; Carrington, M.; Michaeli, S.; Gull, K.; Rudenko, G. Blocking variant surface glycoprotein synthesis in Trypanosoma brucei triggers a general arrest in translation initiation. PLoS ONE 2009, 4, e7532. [Google Scholar] [CrossRef]
- Maris, C.; Dominguez, C.; Allain, F.H. The RNA recognition motif, a plastic RNA-binding platform to regulate post-transcriptional gene expression. FEBS J. 2005, 272, 2118–2131. [Google Scholar] [CrossRef]
- Ridewood, S.; Ooi, C.P.; Hall, B.; Trenaman, A.; Wand, N.V.; Sioutas, G.; Scherwitzl, I.; Rudenko, G. The role of genomic location and flanking 3’UTR in the generation of functional levels of variant surface glycoprotein in Trypanosoma brucei. Mol. Microbiol. 2017, 106, 614–634. [Google Scholar] [CrossRef]
- Munoz-Jordan, J.L.; Davies, K.P.; Cross, G.A.M. Stable expression of mosaic coats of variant surface glycoproteins in Trypanosoma brucei. Science 1996, 272, 1795–1797. [Google Scholar] [CrossRef]
- Batram, C.; Jones, N.G.; Janzen, C.J.; Markert, S.M.; Engstler, M. Expression site attenuation mechanistically links antigenic variation and development in Trypanosoma brucei. Elife 2014, 3, e02324. [Google Scholar] [CrossRef]
- Zimmermann, H.; Subota, I.; Batram, C.; Kramer, S.; Janzen, C.J.; Jones, N.G.; Engstler, M. A quorum sensing-independent path to stumpy development in Trypanosoma brucei. PLoS Pathog. 2017, 13, e1006324. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, L.M.D.; Egler, F.; Arnold, K.; Papavasiliou, N.; Clayton, C.; Erben, E. Functional insights from a surface antigen mRNA-bound proteome. Elife 2021, 10, e68136. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, G.B.; Tinti, M.; Ridgway, M.; Horn, D. Control of Variant Surface Glycoprotein Expression by CFB2 in Trypanosoma brucei and Quantitative Proteomic Connections to Translation and Cytokinesis. mSphere 2022, 7, e0006922. [Google Scholar] [CrossRef] [PubMed]
- Viegas, I.J.; de Macedo, J.P.; Serra, L.; De Niz, M.; Temporão, A.; Pereira, S.S.; Mirza, A.H.; Bergstrom, E.; Rodrigues, J.A.; Aresta-Branco, F.; et al. N6-methyladenosine in poly(A) tails stabilize VSG transcripts. Nature 2022, 604, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Maudlin, I.E.; Kelly, S.; Schwede, A.; Carrington, M. VSG mRNA levels are regulated by the production of functional VSG protein. Mol. Biochem. Parasitol. 2020, 241, 111348. [Google Scholar] [CrossRef] [PubMed]
- Glover, L.; Horn, D. Trypanosomal histone gammaH2A and the DNA damage response. Mol. Biochem. Parasitol. 2012, 183, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Cerritelli, S.M.; Crouch, R.J. Ribonuclease H: The enzymes in eukaryotes. FEBS J. 2009, 276, 1494–1505. [Google Scholar] [CrossRef]
- Briggs, E.; Crouch, K.; Lemgruber, L.; Lapsley, C.; McCulloch, R. Ribonuclease H1-targeted R-loops in surface antigen gene expression sites can direct trypanosome immune evasion. PLoS Genet. 2018, 14, e1007729. [Google Scholar] [CrossRef]
- Khadaroo, B.; Teixeira, M.T.; Luciano, P.; Eckert-Boulet, N.; Germann, S.M.; Simon, M.N.; Gallina, I.; Abdallah, P.; Gilson, E.; Géli, V.; et al. The DNA damage response at eroded telomeres and tethering to the nuclear pore complex. Nat. Cell Biol. 2009, 11, 980–987. [Google Scholar] [CrossRef]
- Kabir, S.; Hockemeyer, D.; de Lange, T. TALEN gene knockouts reveal no requirement for the conserved human shelterin protein Rap1 in telomere protection and length regulation. Cell Rep. 2014, 9, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Martinez, P.; Thanasoula, M.; Carlos, A.R.; Gomez-Lopez, G.; Tejera, A.M.; Schoeftner, S.; Dominguez, O.; Pisano, D.G.; Tarsounas, M.; Blasco, M.A. Mammalian Rap1 controls telomere function and gene expression through binding to telomeric and extratelomeric sites. Nat. Cell Biol. 2010, 12, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.C.K.; Patel, H.; Chia, M.; Moretto, F.; Frith, D.; Snijders, A.P.; van Werven, F.J. Repression of Divergent Noncoding Transcription by a Sequence-Specific Transcription Factor. Mol. Cell 2018, 72, 942–954.e7. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.C.K.; Van Werven, F.J. Transcribe this way: Rap1 confers promoter directionality by repressing divergent transcription. Transcription 2019, 10, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Teo, H.; Ghosh, S.; Luesch, H.; Ghosh, A.; Wong, E.T.; Malik, N.; Orth, A.; de Jesus, P.; Perry, A.S.; Oliver, J.D.; et al. Telomere-independent Rap1 is an IKK adaptor and regulates NF-kappaB-dependent gene expression. Nat. Cell Biol. 2010, 12, 758–767. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef]
- Oksuz, O.; Henninger, J.E.; Warneford-Thomson, R.; Zheng, M.M.; Erb, H.; Vancura, A.; Overholt, K.J.; Hawken, S.W.; Banani, S.F.; Lauman, R.; et al. Transcription factors interact with RNA to regulate genes. Mol. Cell 2023, 83, 2449–2463.e13. [Google Scholar] [CrossRef]
Figure 1.
Domain structures for four RAP1 homologs: TbRAP1, Trypanosoma brucei RAP1; HsRAP1, Homo sapiens RAP1; ScRap1, Saccharomyces cerevisiae Rap1; SpRap1, Schizosaccharomyces pombe Rap1. BRCT, BRCA1 C-terminus; RRM, RNA Recognition Motif; DB, DNA binding; MYBLike, Myb-like; RCT, RAP1 C-terminus. Numbers stand for amino acid positions.
Figure 1.
Domain structures for four RAP1 homologs: TbRAP1, Trypanosoma brucei RAP1; HsRAP1, Homo sapiens RAP1; ScRap1, Saccharomyces cerevisiae Rap1; SpRap1, Schizosaccharomyces pombe Rap1. BRCT, BRCA1 C-terminus; RRM, RNA Recognition Motif; DB, DNA binding; MYBLike, Myb-like; RCT, RAP1 C-terminus. Numbers stand for amino acid positions.

Figure 2.
RAP1 homologs suppress telomeric homologous recombination. RAP1 homologs have been shown to suppress the TERRA level, where TERRA can form telomeric R-loops that induce telomere instability, and homologous recombination is a major DNA DSB repair pathway, particularly in T. brucei. Red bolts represent DNA damage. Blue hexagons in RAP1s represent DNA binding or the critical domain required for its recruitment to the telomere. Budding yeast and human RAP1s also suppress NHEJ under special conditions (not shown).
Figure 2.
RAP1 homologs suppress telomeric homologous recombination. RAP1 homologs have been shown to suppress the TERRA level, where TERRA can form telomeric R-loops that induce telomere instability, and homologous recombination is a major DNA DSB repair pathway, particularly in T. brucei. Red bolts represent DNA damage. Blue hexagons in RAP1s represent DNA binding or the critical domain required for its recruitment to the telomere. Budding yeast and human RAP1s also suppress NHEJ under special conditions (not shown).
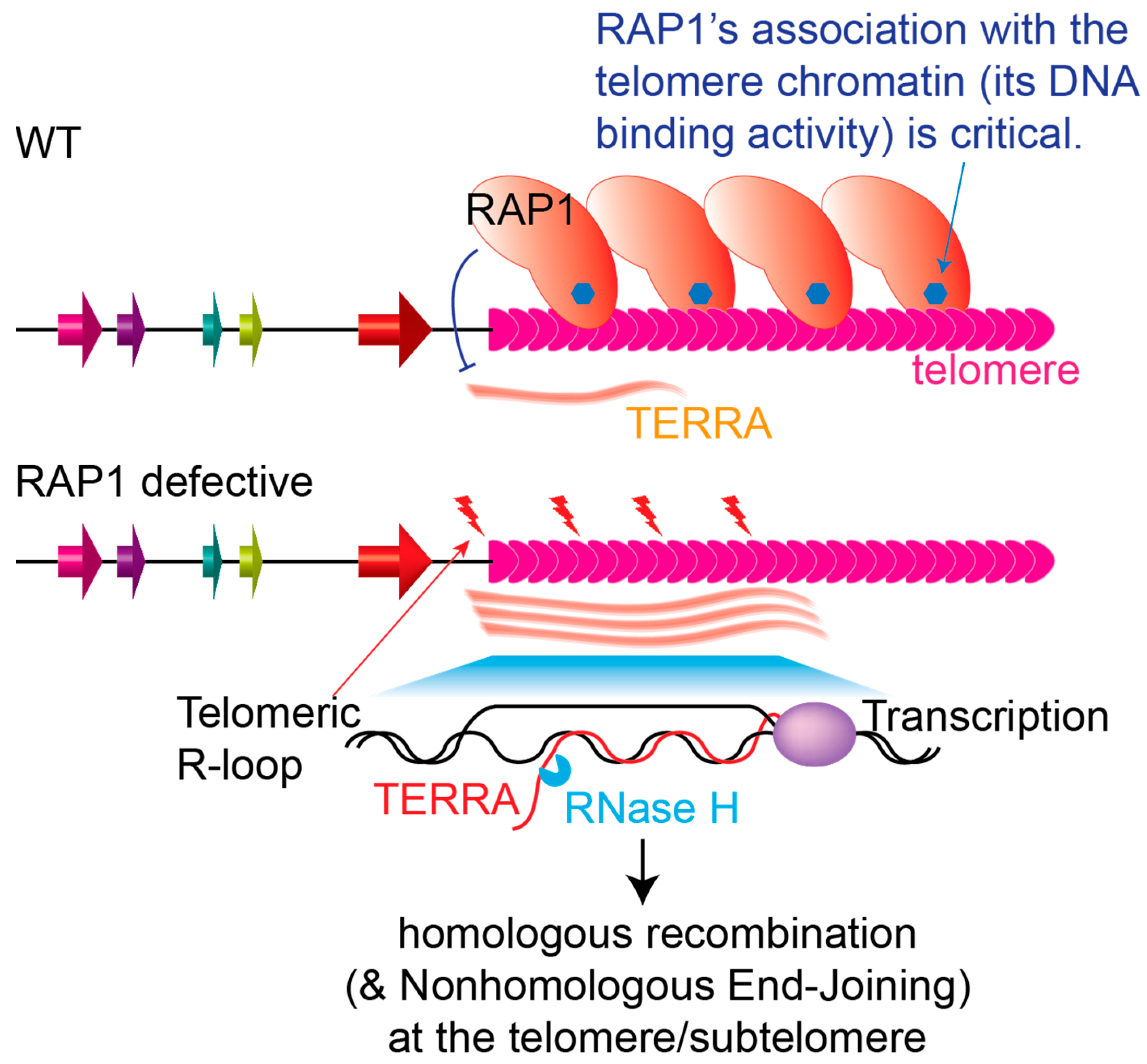
Figure 3.
RAP1 homologs are essential for telomeric silencing. Top: RAP1s help establish/maintain telomeric silencing, presumably by modulating the chromatin structure through recruiting silencers and chromatin remodelers. Curved lines with a short bar at the end represent the silencing effect. The thickness of the lines represents the strength of the silencing effect. Blue hexagons in RAP1 homologs represent DNA binding or the critical domain required for its recruitment to the telomere. Bottom: TbRAP1 has a unique RNA binding RRM domain (represented by a green star), and the competition between RNA and DNA binding ensures that the active VSG is not suppressed by TbRAP1.
Figure 3.
RAP1 homologs are essential for telomeric silencing. Top: RAP1s help establish/maintain telomeric silencing, presumably by modulating the chromatin structure through recruiting silencers and chromatin remodelers. Curved lines with a short bar at the end represent the silencing effect. The thickness of the lines represents the strength of the silencing effect. Blue hexagons in RAP1 homologs represent DNA binding or the critical domain required for its recruitment to the telomere. Bottom: TbRAP1 has a unique RNA binding RRM domain (represented by a green star), and the competition between RNA and DNA binding ensures that the active VSG is not suppressed by TbRAP1.
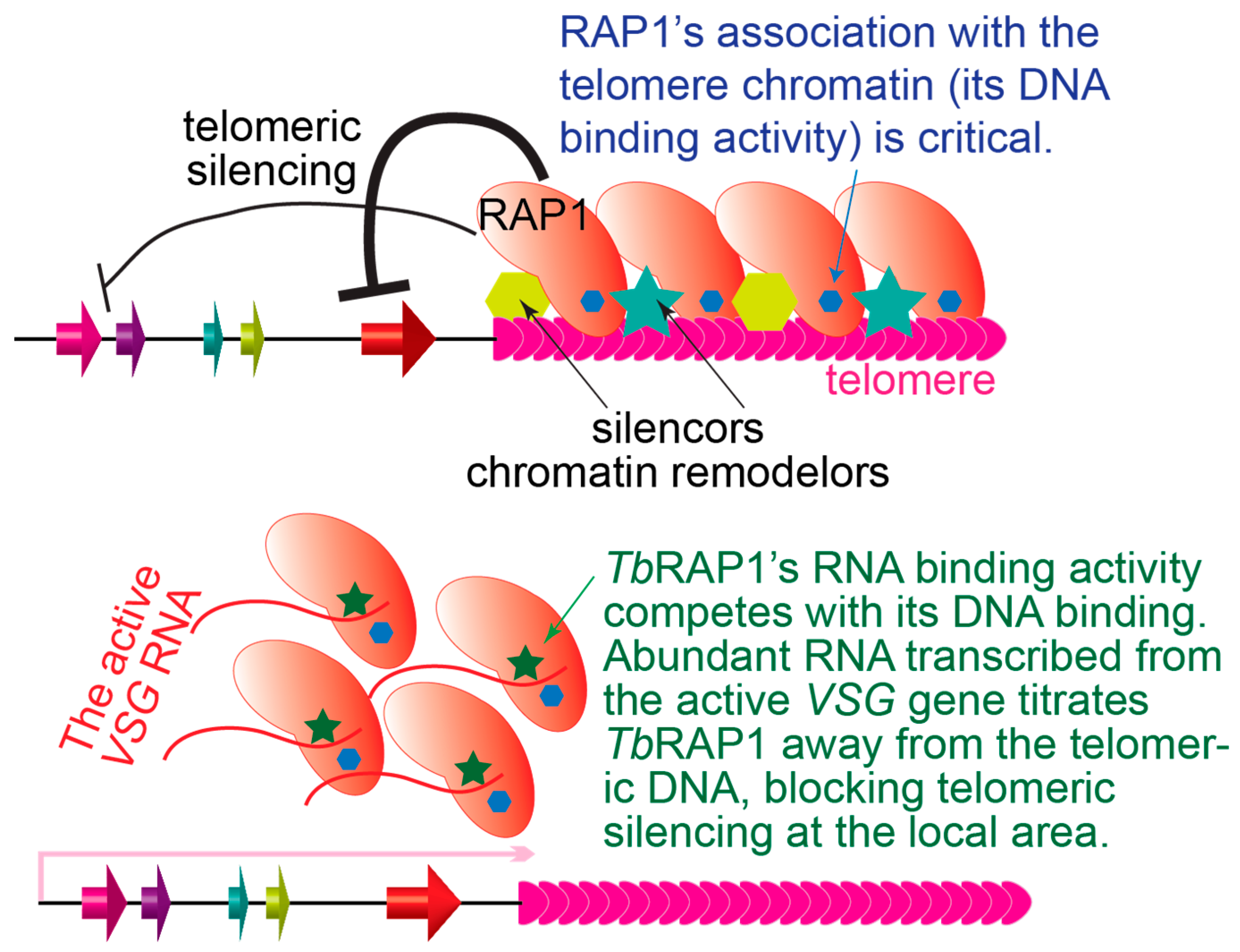
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, B. Unwrap RAP1’s Mystery at Kinetoplastid Telomeres. Biomolecules 2024, 14, 67. https://doi.org/10.3390/biom14010067
AMA Style
Li B. Unwrap RAP1’s Mystery at Kinetoplastid Telomeres. Biomolecules. 2024; 14(1):67. https://doi.org/10.3390/biom14010067
Chicago/Turabian StyleLi, Bibo. 2024. "Unwrap RAP1’s Mystery at Kinetoplastid Telomeres" Biomolecules 14, no. 1: 67. https://doi.org/10.3390/biom14010067
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.